
The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Morphological, Biochemical, and Blood Minerals Parameters
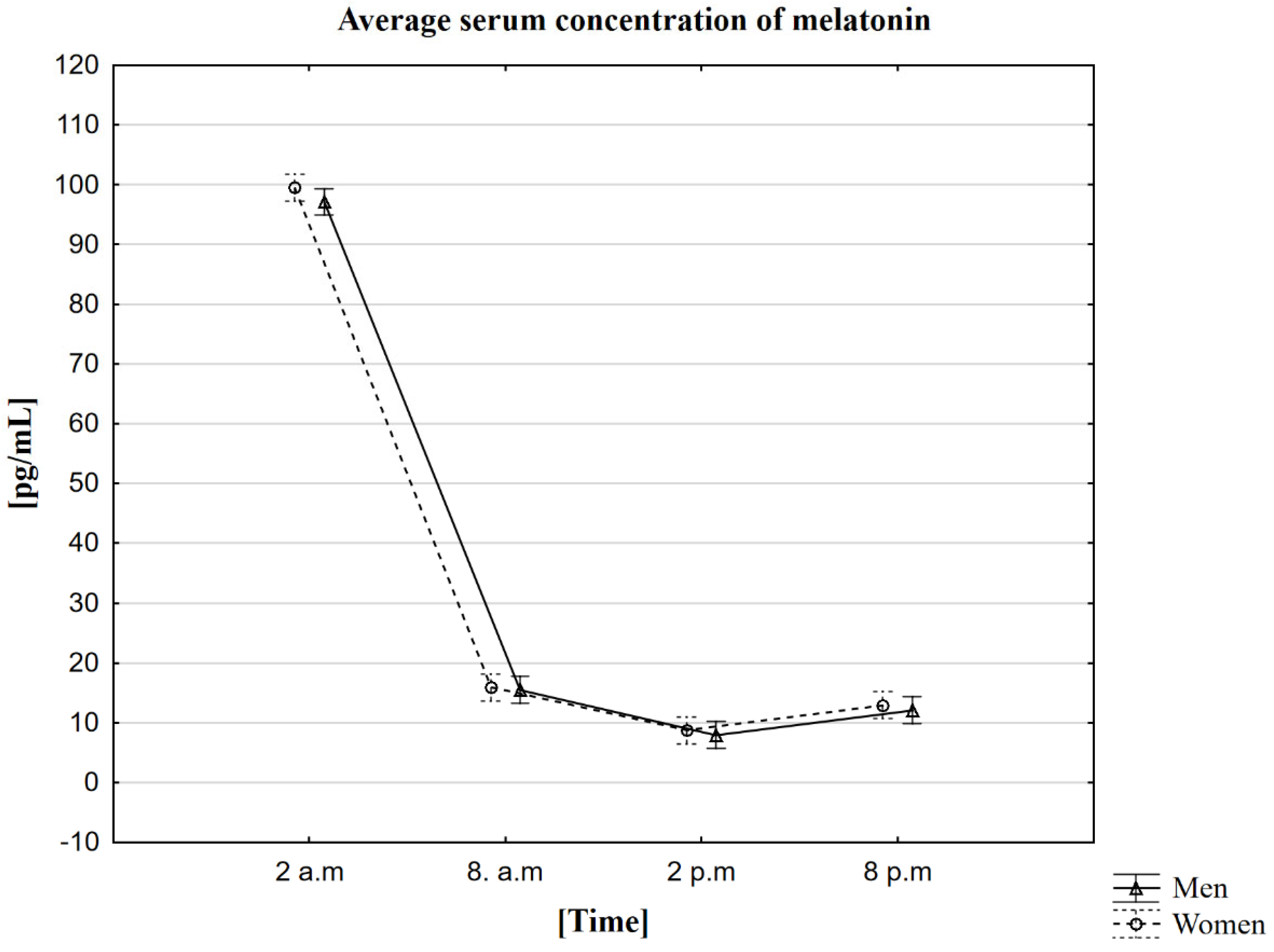
2.2. Analysis of the Influence of the Circadian Rhythm on the Concentration of Melatonin in the Blood Serum
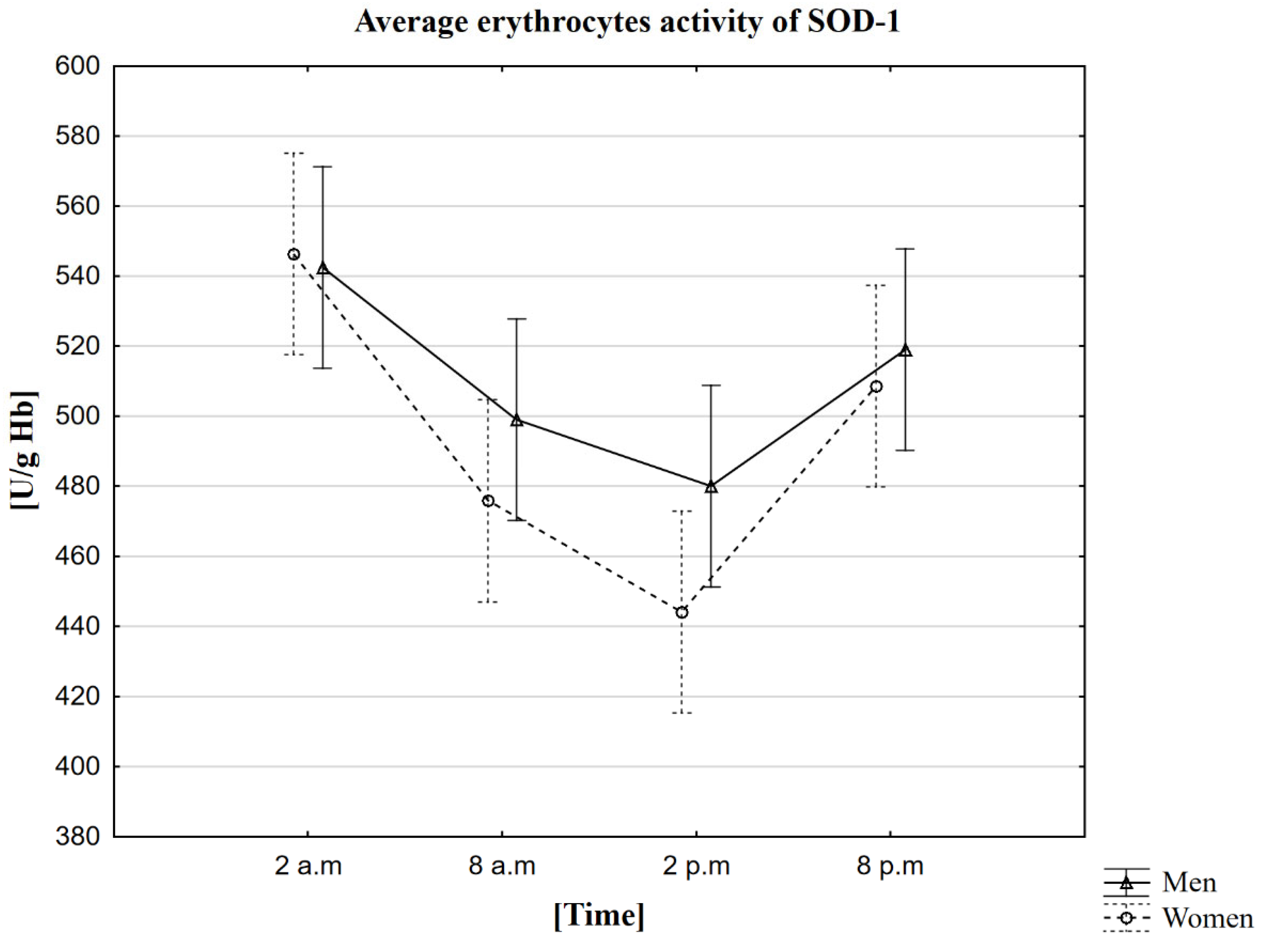
2.3. Analysis of the Influence of Circadian Rhythm on the Activity of Superoxide Dismutase (SOD-1)
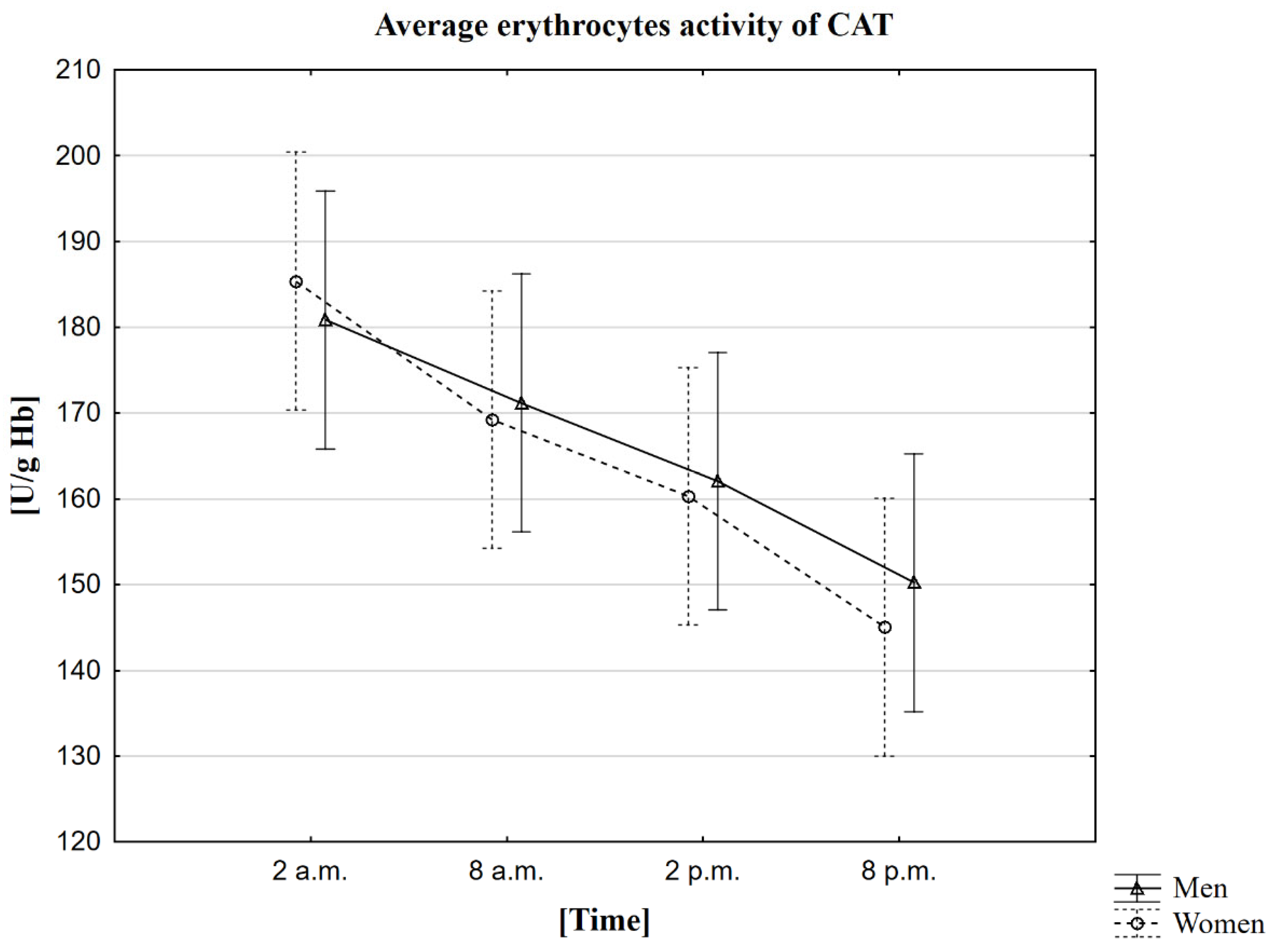
2.4. Analysis of the Influence of the Circadian Rhythm on the Activity of Catalase (CAT)
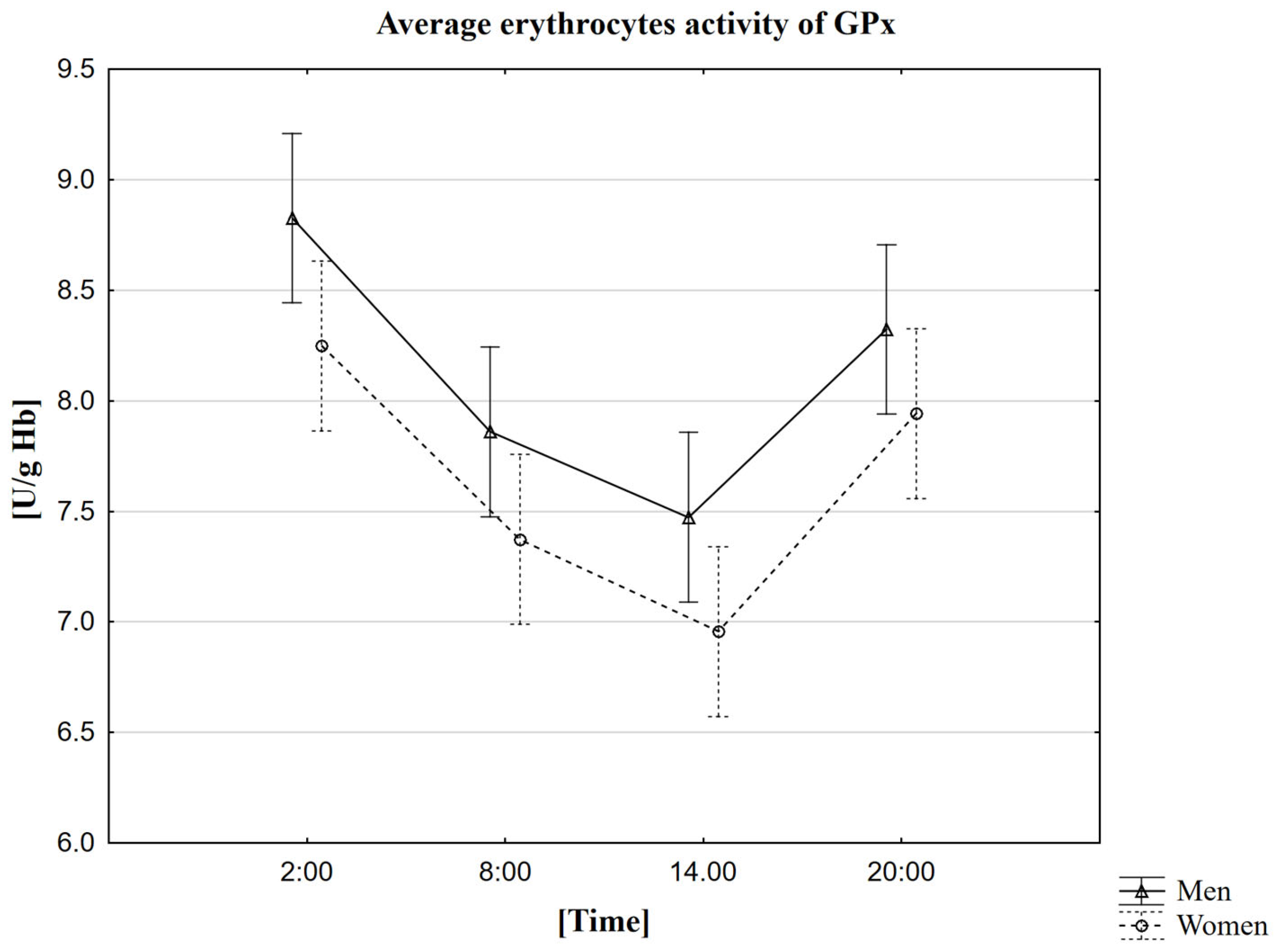
2.5. Analysis of the Influence of Circadian Rhythm on the Activity of Glutathione Peroxidase (GPx)
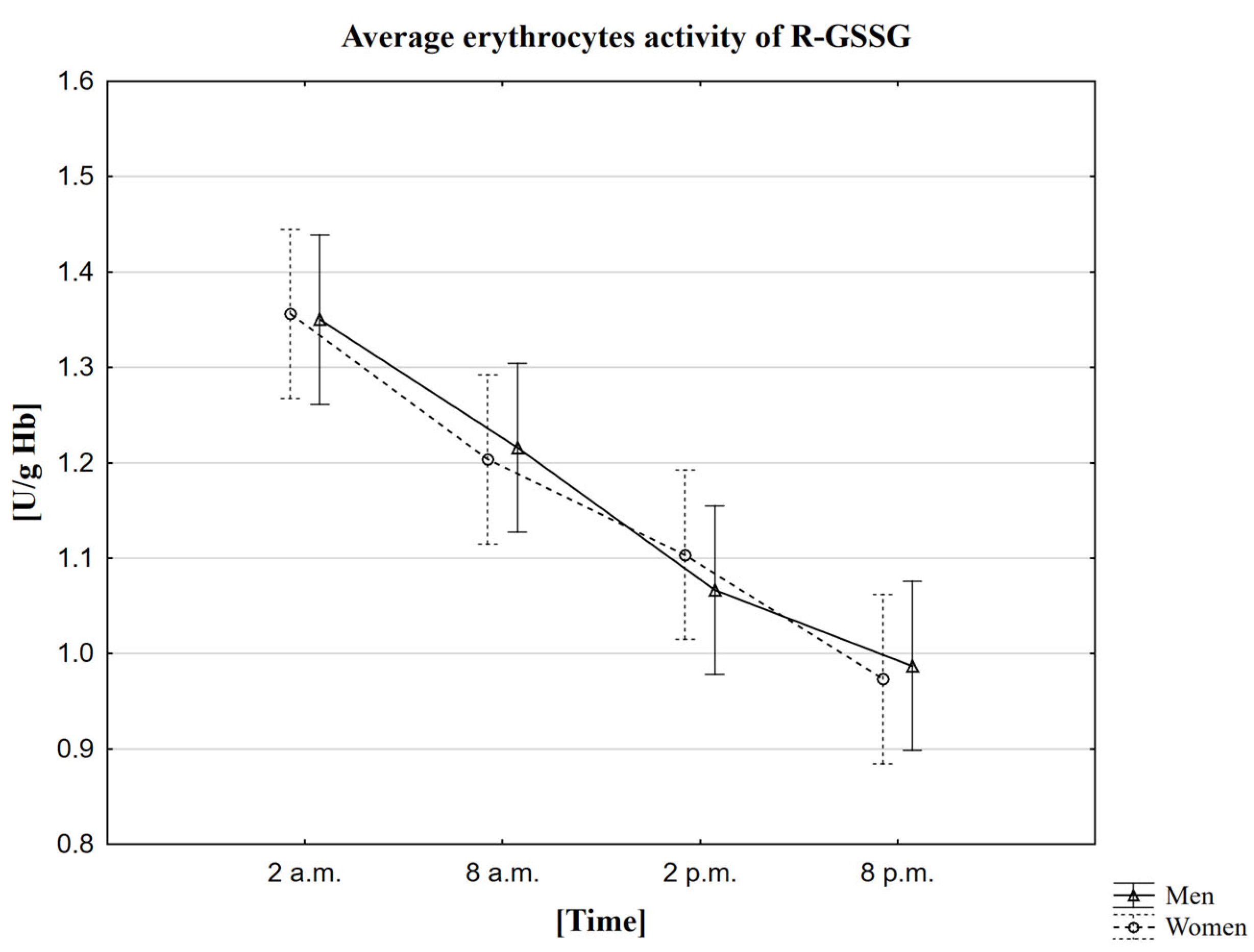
2.6. Analysis of the Influence of Circadian Rhythm on the Activity of Glutathione Reductase (R-GSSG)
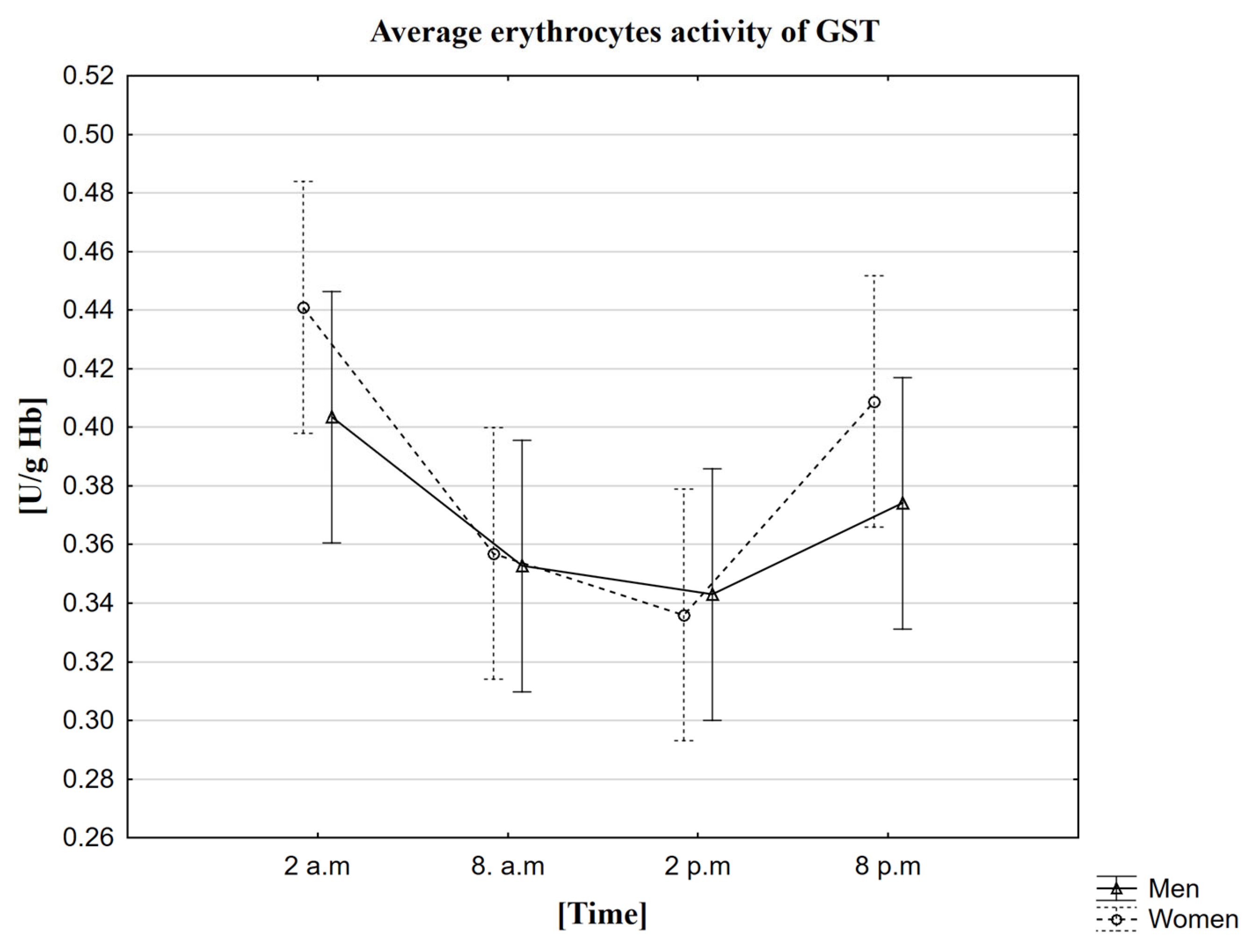
2.7. Analysis of the Influence of Circadian Rhythm on the Activity of Glutathione Transferase (GST)
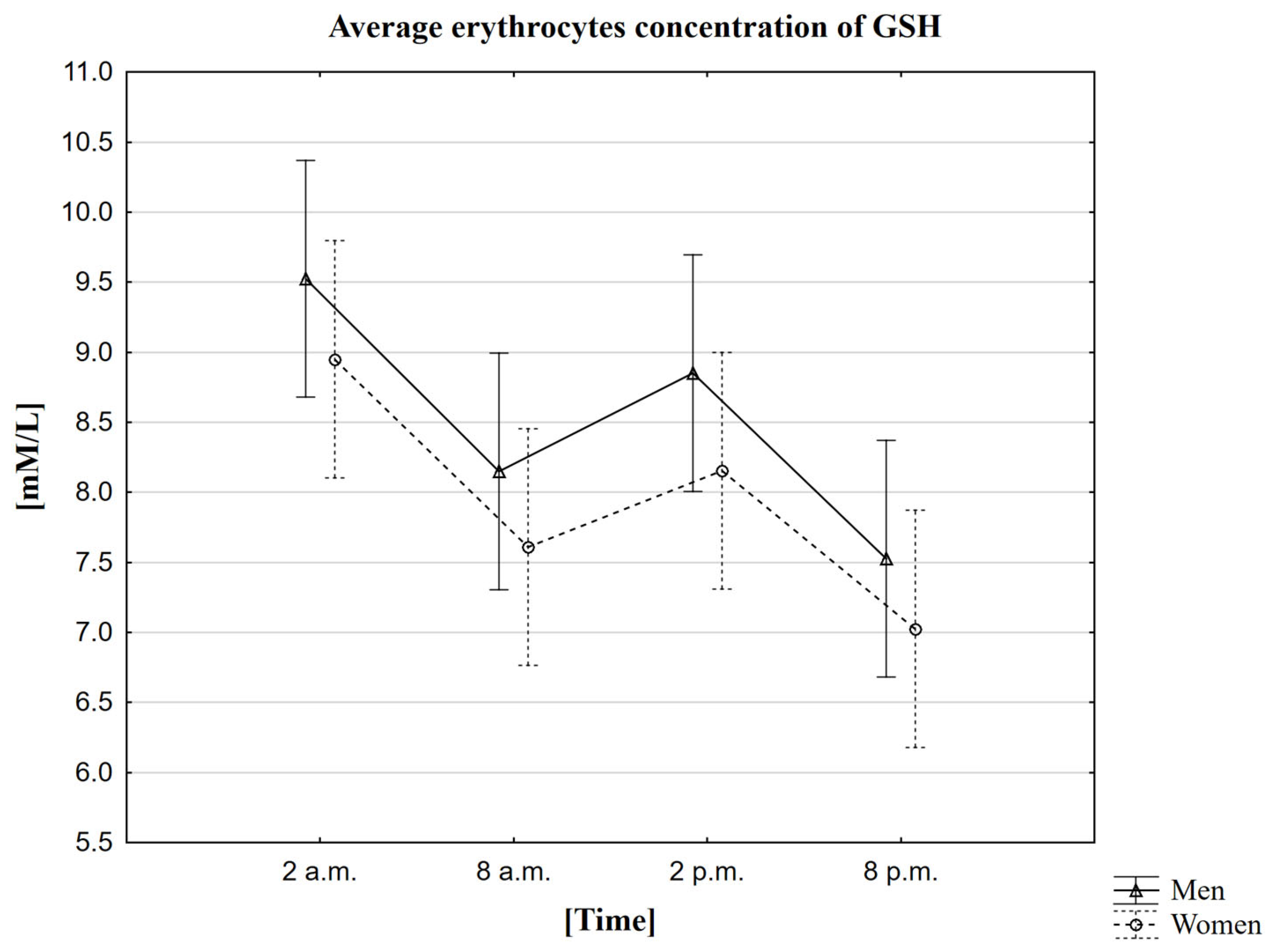
2.8. Analysis of the Influence of the Rhythm on the Concentration of Glutathione (GSH)
3. Discussion
3.1. Morphological, Biochemical, and Blood Minerals Parameters
3.2. The Circadian Rhythm of Melatonin: Influence of Melatonin Concentration on the Activity of Tested Oxidative Stress Enzymes 9SOD-1, CAT, GPx, R-GSSG, CST) and GSH Concentration
3.3. The Circadian Rhythm of SOD-1
3.4. The Circadian Rhythm of CAT
3.5. The Circadian Rhythm of GPx
3.6. The Circadian Rhythm of R-GSSG
3.7. The Circadian Rhythm of GST
3.8. The Circadian Rhythm of GSH
3.9. Relationship between Circadian Rhythms and the Body’s Oxidative Stress—Conclusion
4. Materials and Methods
4.1. Study Group
4.2. Study Material
4.3. Determination of Melatonin Concentration
4.4. Determination of Hemoglobin Concentration by the Drabkin Method in Erythrocyte Hemolysates
4.5. Determination of Superoxide Dismutase Activity (CuZn-SOD—SOD-1) in Erythrocytes
4.6. Determination of Catalase Activity (CAT) in Erythrocytes
4.7. Determination of Glutathione Peroxidase (GPx) Activity in Erythrocytes
4.8. Determination of Glutathione Reductase (R-GSSG) Activity in Erythrocytes
4.9. Determination of Glutathione Transferase (GST) Activity in Erythrocytes
4.10. Determination of Glutathione (GSH) Concentration in Erythrocytes
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leger, D.; Metlaine, A.; Gronfier, C.; Metlaine, A. Consensus Chronobiologie et sommeil de la Société française de recherche et médecine du sommeil (SFRMS). Physiology of the biological clock. Presse Med. 2018, 47, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Kohsaka, A.; Bhawal, U.K.; Muragaki, Y. Potential Roles of Dec and Bmal1 Genes in Interconnecting Circadian Clock and Energy Metabolism. Int. J. Mol. Sci. 2018, 19, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, F.C.; Driver, H.S. Circadian rhythms, sleep, and the menstrual cycle. Sleep Med. 2007, 8, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta 1998, 1366, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Cassone, V.M. Effects of melatonin on vertebrate circadian systems. Trends Neurosci. 1990, 13, 457–464. [Google Scholar] [CrossRef]
- Wagner, G.C.; Johnston, J.D.; Tournier, B.B.; Ebling, F.J.; Hazlerigg, D.G. Melatonin induces gene-specific effects on rhythmic mRNA expression in the pars tuberalis of the Siberian hamster (Phodopus sungorus). Eur. J. Neurosci. 2007, 25, 485–490. [Google Scholar] [CrossRef]
- Weaver, D.R.; Stehle, J.H.; Stopa, E.G.; Reppert, S.M. Melatonin receptors in human hypothalamus and pituitary: Implications for circadian and reproductive responses to melatonin. J. Clin. Endocrinol. Metab. 1993, 76, 295–301. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Cabrera, J.; D’Arpa, D.; Sainz, R.M.; Mayo, J.C.; Ramos, S. The oxidant/antioxidant network: Role of melatonin. Biol. Signals Recept. 1999, 8, 56–63. [Google Scholar] [CrossRef]
- Baydas, G.; Ercel, E.; Canatan, H.; Donder, E.; Akyol, A. Effect of melatonin on oxidative status of rat brain, liver and kidney tissues under constant light exposure. Cell Biochem. Funct. 2001, 19, 37–41. [Google Scholar] [CrossRef]
- Hirata, F.; Hayaishi, O.; Tokuyama, T.; Seno, S. In vitro and in vivo formation of two new metabolites of melatonin. J. Biol. Chem. 1974, 249, 1311–1313. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin.l A mammalian pineal hormone. Endocr. Rev. 1981, 2, 327–346. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Acuna-Castroviejo, D.; Tan, D.X.; Burkhardt, S. Free radical-mediated molecular damage. Mechanisms for the protective actions of melatonin in the central nervous system. Ann. N. Y. Acad. Sci. 2001, 939, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Pablos, M.I.; Agapito, M.T.; Gutierrez, R.; Recio, J.M.; Reiter, R.J.; Barlow-Walden, L.; Acuna-Castroviejo, D.; Menendez-Pelaez, A. Melatonin stimulates the activity of the detoxifying enzyme glutathione peroxidase in several tissues of chicks. J. Pineal Res. 1995, 19, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Pablos, M.I.; Reiter, R.J.; Ortiz, G.G.; Guerrero, J.M.; Agapito, M.T.; Chuang, J.I.; Sewerynek, E. Rhythms of glutathione peroxidase and glutathione reductase in brain of chick and their inhibition by light. Neurochem. Int. 1998, 32, 69–75. [Google Scholar] [CrossRef]
- Cecerska-Heryc, E.; Surowska, O.; Heryc, R.; Serwin, N.; Napiontek-Balinska, S.; Dolegowska, B. Are antioxidant enzymes essential markers in the diagnosis and monitoring of cancer patients—A review. Clin. Biochem. 2021, 93, 1–8. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species (ROS), oxygen radicals and antioxidants: Where are we now, where is the field going and where should we go? Biochem. Biophys. Res. Commun. 2022, 633, 17–19. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Cecerska-Heryc, E.; Polikowska, A.; Serwin, N.; Roszak, M.; Grygorcewicz, B.; Heryc, R.; Michalczyk, A.; Dolegowska, B. Importance of oxidative stress in the pathogenesis, diagnosis, and monitoring of patients with neuropsychiatric disorders, a review. Neurochem. Int. 2022, 153, 105269. [Google Scholar] [CrossRef]
- Cecerska-Heryc, E.; Heryc, R.; Dutkiewicz, G.; Michalczyk, A.; Grygorcewicz, B.; Serwin, N.; Napiontek-Balinska, S.; Dolegowska, B. Xanthine oxidoreductase activity in platelet-poor and rich plasma as a oxidative stress indicator in patients required renal replacement therapy. BMC Nephrol. 2022, 23, 35. [Google Scholar] [CrossRef]
- Ji, M.; Barnwell, C.V.; Grunden, A.M. Characterization of recombinant glutathione reductase from the psychrophilic Antarctic bacterium Colwellia psychrerythraea. Extremophiles 2015, 19, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P.; Lund, L.G. Cellular reducing equivalents and oxidative stress. Free Radic. Biol. Med. 1994, 17, 65–75. [Google Scholar] [CrossRef]
- Janda, K.; Kasprzak, M.; Wolska, J. Vitamin C- structure, properties, occurrence and functions. Pomeranian J. Life Sci. 2015, 61, 419–425. [Google Scholar] [PubMed]
- Bose, K.S.; Vyas, P.; Singh, M. Plasma non-enzymatic antioxidants-vitamin C, E, beta-carotenes, reduced glutathione levels and total antioxidant activity in oral sub mucous fibrosis. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 530–532. [Google Scholar]
- Lee, G.Y.; Han, S.N. The Role of Vitamin E in Immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Bivona, J.J., 3rd; Patel, S.; Vajdy, M. Induction of cellular and molecular Immunomodulatory pathways by vitamin E and vitamin C. Expert Opin. Biol. Ther. 2017, 17, 1539–1551. [Google Scholar] [CrossRef]
- Kipp, A.P.; Frombach, J.; Deubel, S.; Brigelius-Flohe, R. Selenoprotein W as biomarker for the efficacy of selenium compounds to act as source for selenoprotein biosynthesis. Methods Enzymol. 2013, 527, 87–112. [Google Scholar] [CrossRef]
- Rutz, J.K.; Borges, C.D.; Zambiazi, R.C.; da Rosa, C.G.; da Silva, M.M. Elaboration of microparticles of carotenoids from natural and synthetic sources for applications in food. Food Chem. 2016, 202, 324–333. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef]
- Mata-Perez, C.; Spoel, S.H. Thioredoxin-mediated redox signalling in plant immunity. Plant. Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef]
- Ouyang, Y.; Peng, Y.; Li, J.; Holmgren, A.; Lu, J. Modulation of thiol-dependent redox system by metal ions via thioredoxin and glutaredoxin systems. Metallomics 2018, 10, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Li Volti, G.; Giallongo, C.; Grasso, S.; Tomassoni, D.; Anfuso, C.D.; Lupo, G.; Amenta, F.; Avola, R.; Bramanti, V. Biochemical and clinical relevance of alpha lipoic acid: Antioxidant and anti-inflammatory activity, molecular pathways and therapeutic potential. Inflamm. Res. 2017, 66, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Majumder, D.; Das, A.; Saha, C. Catalase inhibition an anti cancer property of flavonoids: A kinetic and structural evaluation. Int. J. Biol. Macromol. 2017, 104, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Tahara, Y.; Aoyama, S.; Shibata, S. The mammalian circadian clock and its entrainment by stress and exercise. J. Physiol. Sci. 2017, 67, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.; Davis, A.J.; Giebultowicz, J.M. Circadian regulation of response to oxidative stress in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2008, 374, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Dembińska–Kieć, A.; Naskalski, J.W.; Solnica, B. Diagnostyka Laboratoryjna z Elementami Biochemii Klinicznej, 4th ed.; Edra Urban & Partner: Wrocław, Poland, 2017; pp. 274–283. [Google Scholar]
- Bunn, H.F. Erythropoietin. Cold Spring Harb. Perspect. Med. 2013, 3, a011619. [Google Scholar] [CrossRef] [Green Version]
- Wallach, J. Badania Analitów Krwi i Ich Zmiany w Różnych Chorobach. Interpretacja Badań Laboratoryjnych, 2nd ed.; Franek, E., Kokot, F., Pietruczuk, M., Eds.; MediPage: Warszawa, Poland, 2011; pp. 55,97. [Google Scholar]
- Ohlander, S.J.; Varghese, B.; Pastuszak, A.W. Erythrocytosis Following Testosterone Therapy. Sex Med. Rev. 2018, 6, 77–85. [Google Scholar] [CrossRef]
- Neumeister, B.; Basenthal, I.; Böhm, B.O. Diagnostyka Laboratoryjna; Pietruczuk, M., Bartoszko-Tyczkowska, A., Eds.; Elsevier Urban & Partner: Wrocław, Poland, 2013; pp. 192,193. [Google Scholar]
- Itoh, K.; Masumori, S.; Mukai, D.; Sakakibara, H.; Yasuda, M.; Shimoi, K. Dosage time affects alkylating agents induced micronuclei in mouse peripheral blood reticulocytes through the function of erythropoietin. J. Toxicol. Sci. 2019, 44, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Pagana, D.K.; Pagana, T.J. Triglicerydy. In Testy Laboratoryjne i Badania Diagnostyczne w Medycynie, 1st ed.; Pietruczuk, M., Ed.; Elsevier Urban & Partner: Wrocław, Poland, 2013; pp. 872–873. [Google Scholar]
- Claustrat, B.; Leston, J. Melatonin: Physiological effects in humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Pevet, P. Basic aspects of melatonin action. Sleep Med. Rev. 1998, 2, 175–190. [Google Scholar] [CrossRef]
- Michalik, A.; Bobinski, R. “Jet-lag”—Pathophysiology and methods of prevention and treatment. Przegl. Epidemiol. 2009, 63, 589–595. [Google Scholar] [PubMed]
- Ding, K.; Wang, H.; Xu, J.; Li, T.; Zhang, L.; Ding, Y.; Zhu, L.; He, J.; Zhou, M. Melatonin stimulates antioxidant enzymes and reduces oxidative stress in experimental traumatic brain injury: The Nrf2-ARE signaling pathway as a potential mechanism. Free Radic. Biol. Med. 2014, 73, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nozik-Grayck, E.; Suliman, H.B.; Piantadosi, C.A. Extracellular superoxide dismutase. Int. J. Biochem. Cell Biol. 2005, 37, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; St. Clair, D.K. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krych-Madej, J.; Gebicka, L. Interactions of nitrite with catalase: Enzyme activity and reaction kinetics studies. J. Inorg. Biochem. 2017, 171, 10–17. [Google Scholar] [CrossRef]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—An evolutionary overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef]
- Diaz-Munoz, M.; Hernandez-Munoz, R.; Suarez, J.; Chagoya de Sanchez, V. Day-night cycle of lipid peroxidation in rat cerebral cortex and their relationship to the glutathione cycle and superoxide dismutase activity. Neuroscience 1985, 16, 859–863. [Google Scholar] [CrossRef]
- Martin, V.; Sainz, R.M.; Mayo, J.C.; Antolin, I.; Herrera, F.; Rodriguez, C. Daily rhythm of gene expression in rat superoxide dismutases. Endocr. Res. 2003, 29, 83–95. [Google Scholar] [CrossRef]
- Singh, R.; Singh, R.K.; Tripathi, A.K.; Cornelissen, G.; Schwartzkopff, O.; Otsuka, K.; Halberg, F. Chronomics of circulating plasma lipid peroxides and anti-oxidant enzymes and other related molecules in cirrhosis of liver. In the memory of late Shri Chetan Singh. Biomed. Pharmacother. 2005, 59, S229–S235. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.H.; Bozigian, H.P.; Merrick, B.A.; Birt, D.F.; Schnell, R.C. Circadian variations in glutathione-S-transferase and glutathione peroxidase activities in the mouse. Toxicol. Lett. 1983, 19, 23–27. [Google Scholar] [CrossRef]
- Kedrowski, B.L.; Gutow, J.H.; Stock, G.; Smith, M.; Jordan, C.; Masterson, D.S. Glutathione reductase activity with an oxidized methylated glutathione analog. J. Enzyme Inhib. Med. Chem. 2014, 29, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Voehringer, D.W.; Meyn, R.E. Redox aspects of Bcl-2 function. Antioxid. Redox Signal. 2000, 2, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Bhatla, S.C. Melatonin and nitric oxide modulate glutathione content and glutathione reductase activity in sunflower seedling cotyledons accompanying salt stress. Nitric Oxide 2016, 59, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, R.K.; Masood, T.; Tripathi, A.K.; Mahdi, A.A.; Singh, R.K.; Schwartzkopff, O.; Cornelissen, G. Circadian time structure of circulating plasma lipid peroxides, antioxidant enzymes and other small molecules in peptic ulcers. Clin. Chim. Acta 2015, 451, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Gupta, S. The multifaceted role of glutathione S-transferases in cancer. Cancer Lett. 2018, 433, 33–42. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Marengo, B.; Nitti, M.; Furfaro, A.L.; Colla, R.; Ciucis, C.D.; Marinari, U.M.; Pronzato, M.A.; Traverso, N.; Domenicotti, C. Redox Homeostasis and Cellular Antioxidant Systems: Crucial Players in Cancer Growth and Therapy. Oxid. Med. Cell. Longev. 2016, 2016, 6235641. [Google Scholar] [CrossRef] [Green Version]
- Tew, K.D.; Townsend, D.M. Glutathione-s-transferases as determinants of cell survival and death. Antioxid. Redox Signal. 2012, 17, 1728–1737. [Google Scholar] [CrossRef] [Green Version]
- Inoue, N.; Imai, K.; Aimoto, T. Circadian variation of hepatic glutathione S-transferase activities in the mouse. Xenobiotica 1999, 29, 43–51. [Google Scholar] [CrossRef]
- Chakravarty, S.; Rizvi, S.I. Day and Night GSH and MDA Levels in Healthy Adults and Effects of Different Doses of Melatonin on These Parameters. Int. J. Cell Biol. 2011, 2011, 404591. [Google Scholar] [CrossRef] [PubMed]
- Ulanczyk, Z.; Grabowicz, A.; Cecerska-Heryc, E.; Sleboda-Taront, D.; Krytkowska, E.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Dolegowska, B.; Machalinska, A. Dietary and Lifestyle Factors Modulate the Activity of the Endogenous Antioxidant System in Patients with Age-Related Macular Degeneration: Correlations with Disease Severity. Antioxidants 2020, 9, 954. [Google Scholar] [CrossRef] [PubMed]
- Cecerska-Heryc, E.; Krauze, K.; Szczesniak, A.; Goryniak-Mikolajczyk, A.; Serwin, N.; Sleboda-Taront, D.; Jacek, R.; Heryc, R.; Michalczyk, A.; Dolegowska, B. Activity of erythrocyte antioxidant enzymes in healthy women depends on age, BMI, physical activity, and diet. J. Health Popul. Nutr. 2022, 41, 35. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Women (n = 33) | Men (n = 33) | p-Value |
|---|---|---|---|
| Value (Mean ± SD) [NR] | Value (Mean ± SD) [NR] | ||
| RBC [1012/L] | 4.5 ± 0.4 [3.8–5.2] | 5.3 ± 0.5 [4.0–5.9] | <0.0001 |
| Hb [mM/L] | 7.9 ± 0.6 [7.4–9.9] | 9.2 ± 0.6 [8.7–11.2] | <0.0001 |
| Hct [%] | 40 ± 3 [33–44] | 46 ± 3 [41–50] | <0.0001 |
| WBC [109/L] | 6.0 ± 1,4 [4.5–11.0] | 5.8 ± 1,1 [4.5–11.0] | 0.9929 |
| PLT [103/µL] | 252 ± 41 [150–450] | 253 ± 40 [150–450] | 0.9715 |
| GLC [mg/dL] | 90 ± 8 [70–99] | 92 ± 7 [70–99] | 0.1311 |
| IP [mM/L] | 1.5 ± 0.2 [1.12–1.45] | 1.5 ± 0.1 [1.12–1.45] | 0.9359 |
| tMG [mM/L] | 0.92 ± 0.03 [0.85–1.10] | 0.93 ± 0.04 [0.85–1.10] | 0.6487 |
| TCAL [mM/L] | 2.3 ± 0.2 [2.2–2.6] | 2.4 ± 0.1 [2.2–2.6] | 0.7141 |
| TC [mg/dL] | 179 ± 23 [125–200] | 187 ± 22 [125–200] | 0.1555 |
| TG [mg/dL] | 92 ± 19 [<150] | 111 ± 35 [<150] | 0.0207 |
| LDL-C [mg/dL] | 89 ± 23 [50–100] | 92 ± 25 [50–100] | 0.7141 |
| HDL-C [mg/dL] | 70 ± 11 [50–90] | 62 ± 14 [45–70] | 0.3669 |
| TP [g/dL] | 6.5 ± 0.4 [6–8] | 6.6 ± 0.5 [6–8] | 0.2491 |
| Alb [g/dL] | 3.9 ± 0.3 [3.5–5.0] | 4.0 ± 0.3 [3.5–5.0] | 0.0525 |
| CRE [mg/dL] | 0.9 ± 0.2 [0.6–1.1] | 1.0 ± 0.2 [0.7–1.3] | 0.0012 |
| UA [mg/dL] | 4.5 ± 0.7 [2.7–7.3] | 5.3 ± 1.0 [4.0–8.5] | 0.0005 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 99.5 | 15.9 | 8.7 | 12.9 | 97.1 | 15.5 | 8.0 | 12.1 |
| Standard deviation | ±11.2 | ±3.8 | ±2.5 | ±4.7 | ±11.7 | ±4.4 | ±2.6 | ±3.0 |
| Median | 98.4 | 15.3 | 9.1 | 12.2 | 98.1 | 14.5 | 8.1 | 12.2 |
| Minimum | 76.2 | 8.3 | 3.6 | 4.7 | 72.1 | 8.2 | 3.9 | 5.7 |
| Maximum | 135.8 | 22.8 | 13.4 | 27.1 | 118.9 | 23.8 | 15.0 | 19.0 |
| Upper quartile | 105.3 | 18.2 | 10.3 | 15.1 | 104.6 | 18.8 | 10.1 | 13.8 |
| Lower quartile | 94.6 | 13.9 | 6.9 | 9.4 | 89.4 | 12.0 | 5.8 | 10.3 |
| Interquartile range | 10.7 | 4.3 | 3.4 | 5.7 | 15.2 | 6.8 | 4.3 | 3.5 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 546 | 476 | 444 | 509 | 542 | 499 | 480 | 519 |
| Standard deviation | ±85 | ±74 | ±76 | ±78 | ±91 | ±87 | ±85 | ±93 |
| Median | 544 | 473 | 427 | 501 | 536 | 501 | 479 | 522 |
| Minimum | 398 | 371 | 315 | 382 | 388 | 364 | 356 | 370 |
| Maximum | 753 | 737 | 715 | 763 | 758 | 724 | 713 | 740 |
| Upper quartile | 595 | 529 | 492 | 558 | 580 | 532 | 512 | 551 |
| Lower quartile | 514 | 418 | 399 | 453 | 477 | 440 | 432 | 447 |
| Interquartile range | 81 | 111 | 94 | 105 | 103 | 92 | 80 | 104 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 185 | 169 | 160 | 145 | 181 | 171 | 162 | 150 |
| Standard deviation | ±49 | ±43 | ±43 | ±40 | ±45 | ±44 | ±43 | ±43 |
| Median | 173 | 159 | 149 | 136 | 176 | 160 | 150 | 142 |
| Minimum | 122 | 108 | 92 | 91 | 99 | 95 | 86 | 83 |
| Maximum | 285 | 275 | 271 | 231 | 281 | 268 | 267 | 248 |
| Upper quartile | 210 | 201 | 186 | 165 | 213 | 201 | 192 | 179 |
| Lower quartile | 155 | 140 | 131 | 117 | 152 | 142 | 137 | 123 |
| Interquartile range | 55 | 61 | 55 | 48 | 61 | 59 | 55 | 56 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 8.2 | 7.4 | 7.0 | 7.9 | 8.8 | 7.9 | 7.5 | 8.3 |
| Standard deviation | ±1.4 | ±1.2 | ±1.1 | ±1.3 | ±1.0 | ±1.0 | ±0.9 | ±1.0 |
| Median | 8.6 | 7.6 | 7.3 | 8.2 | 8.9 | 7.7 | 7.5 | 8.3 |
| Minimum | 5.5 | 5.3 | 5.1 | 5.6 | 7.2 | 6.3 | 5.5 | 6.8 |
| Maximum | 10.6 | 9.5 | 9.1 | 10.0 | 10.8 | 10.0 | 9.4 | 10.1 |
| Upper quartile | 9.2 | 8.2 | 7.6 | 9.0 | 10.0 | 8.4 | 8.1 | 8.2 |
| Lower quartile | 7.3 | 6.3 | 6.0 | 7.0 | 8.1 | 7.1 | 7.0 | 7.6 |
| Interquartile range | 1.9 | 1.9 | 1.6 | 2.0 | 1.9 | 1.3 | 1.0 | 1.2 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 1.4 | 1.2 | 1.1 | 1.0 | 1.4 | 1.2 | 1.1 | 1.0 |
| Standard deviation | ±0.3 | ±0.3 | ±0.3 | ±0.2 | ±0.3 | ±0.3 | ±0.2 | ±0.2 |
| Median | 1.3 | 1.1 | 1.0 | 0.9 | 1.3 | 1.2 | 1.1 | 0.9 |
| Minimum | 0.8 | 0.8 | 0.6 | 0.6 | 0.8 | 0.7 | 0.6 | 0.6 |
| Maximum | 2.0 | 1.8 | 1.7 | 1.6 | 1.9 | 2.0 | 1.5 | 1.6 |
| Upper quartile | 1.4 | 1.3 | 1.2 | 1.1 | 1.5 | 1.4 | 1.2 | 1.1 |
| Lower quartile | 1.2 | 1.0 | 0.9 | 0.9 | 1.2 | 1.1 | 1.0 | 0.8 |
| Interquartile range | 0.2 | 0.3 | 0.3 | 0.2 | 0.3 | 0.3 | 0.2 | 0.3 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 0.44 | 0.36 | 0.34 | 0.41 | 0.40 | 0.35 | 0.34 | 0.37 |
| Standard deviation | ±0.14 | ±0.12 | ±0.13 | ±0.17 | ±0.09 | ±0.10 | ±0.12 | ±0.11 |
| Median | 0.44 | 0.35 | 0.32 | 0.40 | 0.39 | 0.34 | 0.34 | 0.32 |
| Minimum | 0.19 | 0.15 | 0.11 | 0.16 | 0.28 | 0.19 | 0.11 | 0.25 |
| Maximum | 0.78 | 0.66 | 0.59 | 0.99 | 0.66 | 0.59 | 0.59 | 0.69 |
| Upper quartile | 0.50 | 0.42 | 0.45 | 0.44 | 0.43 | 0.42 | 0.41 | 0.43 |
| Lower quartile | 0.34 | 0.30 | 0.25 | 0.30 | 0.34 | 0.27 | 0.27 | 0.30 |
| Interquartile range | 0.16 | 0.12 | 0.20 | 0.14 | 0.09 | 0.15 | 0.15 | 0.14 |
| Gender | Women (n = 33) | Men (n = 33) | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Blood Collection | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. | 2 a.m. | 8 a.m. | 2 p.m. | 8 p.m. |
| Mean | 9.0 | 7.6 | 8.2 | 7.0 | 9.5 | 8.2 | 8.9 | 7.5 |
| Standard deviation | ±2.7 | ±2.5 | ±2.7 | ±2.6 | ±2.4 | ±2.2 | ±2.4 | ±2.1 |
| Median | 7.9 | 6.7 | 7.2 | 6.0 | 8.9 | 7.4 | 7.8 | 6.5 |
| Minimum | 5.5 | 5.2 | 5.4 | 5.0 | 6.1 | 5.1 | 5.3 | 5.1 |
| Maximum | 16.1 | 14.5 | 15.6 | 14.2 | 13.9 | 12.4 | 13.5 | 12.3 |
| Upper quartile | 9.7 | 7.9 | 8.4 | 7.2 | 11.1 | 10.2 | 10.8 | 9.7 |
| Lower quartile | 7.1 | 6.1 | 6.4 | 5.3 | 7.8 | 6.5 | 7.2 | 6.1 |
| Interquartile range | 2.6 | 1.8 | 2.0 | 1.9 | 3.3 | 3.7 | 3.6 | 3.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budkowska, M.; Cecerska-Heryć, E.; Marcinowska, Z.; Siennicka, A.; Dołęgowska, B. The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes. Int. J. Mol. Sci. 2022, 23, 14275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214275
Budkowska M, Cecerska-Heryć E, Marcinowska Z, Siennicka A, Dołęgowska B. The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes. International Journal of Molecular Sciences. 2022; 23(22):14275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214275
Chicago/Turabian StyleBudkowska, Marta, Elżbieta Cecerska-Heryć, Zuzanna Marcinowska, Aldona Siennicka, and Barbara Dołęgowska. 2022. "The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes" International Journal of Molecular Sciences 23, no. 22: 14275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214275