
Computational Repurposing of Mitoxantrone-Related Structures against Monkeypox Virus: A Molecular Docking and 3D Pharmacophore Study

 , , , and
, , , and
Abstract
:1. Introduction
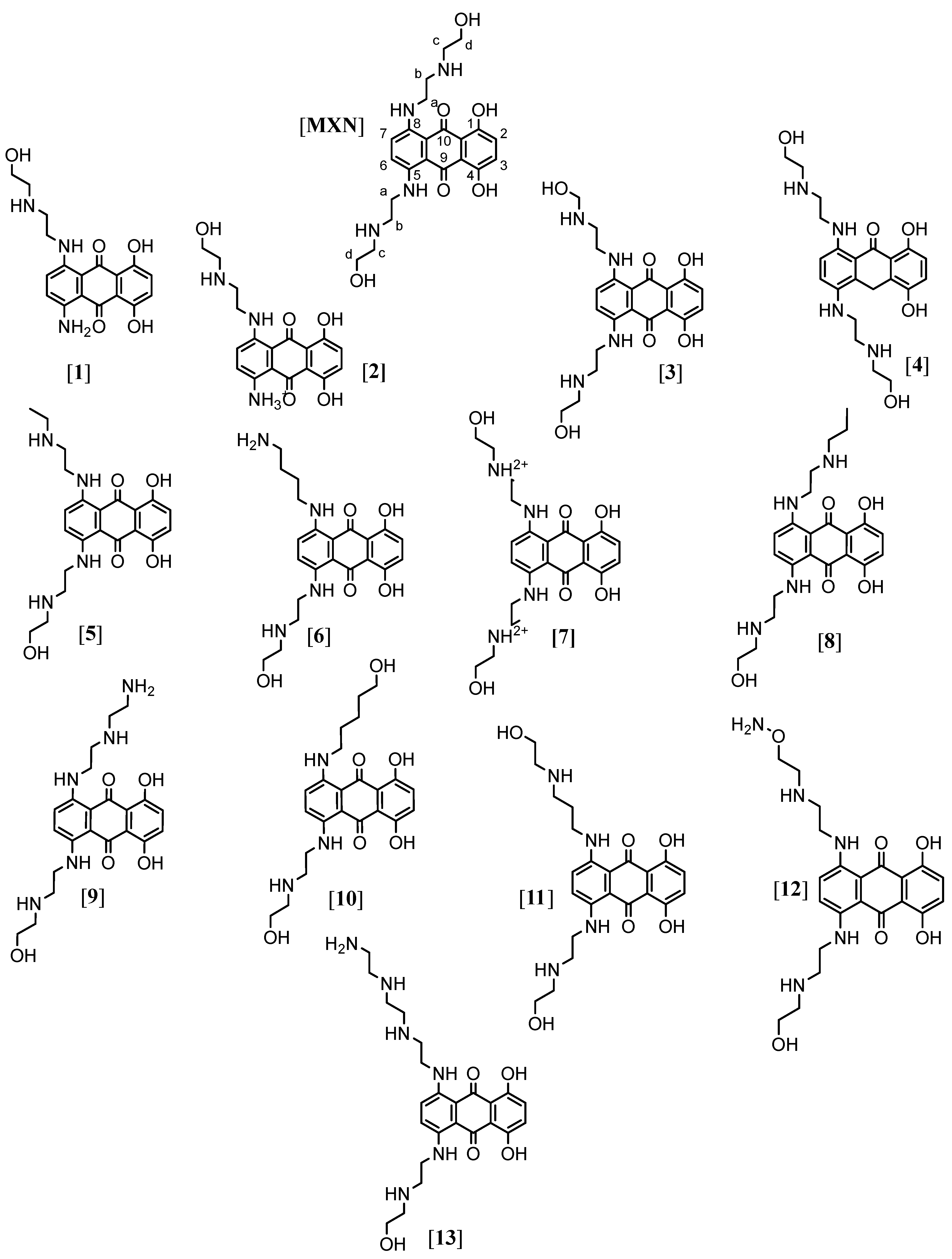
2. Results and Discussion
2.1. Determination of Antiviral Activity Using Molecular Docking
2.2. Pharmacophore Evaluation
3. Materials and Methods
3.1. Origin of Compounds
3.2. Molecular Docking
3.3. 3D Pharmacophore Model Generation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacNeil, A.; Reynolds, M.; Braden, Z.; Carroll, D.S.; Bostik, V.; Karem, K.; Smith, S.K.; Davidson, W.; Li, Y.; Moundeli, A.; et al. Transmission of atypical varicella-zoster virus infections involving palm and sole manifestations in an area with monkeypox endemicity. Clin. Infect Dis. 2009, 48, e6–e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect Genet. Evol. 2014, 21, 15–40. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.; Leggat, P.A. Human Monkeypox: Current State of Knowledge and Implications for the Future. Trop. Med. Infect Dis. 2016, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Vallee, A.; Farfour, E.; Zucman, D. Monkeypox virus: A novel sexually transmitted disease? A case report from France. Travel. Med. Infect. Dis. 2022, 49, 102394. [Google Scholar] [CrossRef] [PubMed]
- Tesini, B.L. Monkeypox. Available online: https://www.msdmanuals.com/en-sg/professional/infectious-diseases/pox-viruses/monkeypox (accessed on 13 August 2022).
- Yong, S.E.F.; Ng, O.T.; Ho, Z.J.M.; Mak, T.M.; Marimuthu, K.; Vasoo, S.; Yeo, T.W.; Ng, Y.K.; Cui, L.; Ferdous, Z.; et al. Imported Monkeypox, Singapore. Emerg. Infect Dis. 2020, 26, 1826–1830. [Google Scholar] [CrossRef]
- Vaughan, A.; Aarons, E.; Astbury, J.; Balasegaram, S.; Beadsworth, M.; Beck, C.R.; Chand, M.; O’Connor, C.; Dunning, J.; Ghebrehewet, S.; et al. Two cases of monkeypox imported to the United Kingdom, September 2018. Eurosurveillance 2018, 23, 1800509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erez, N.; Achdout, H.; Milrot, E.; Schwartz, Y.; Wiener-Well, Y.; Paran, N.; Politi, B.; Tamir, H.; Israely, T.; Weiss, S.; et al. Diagnosis of Imported Monkeypox, Israel, 2018. Emerg. Infect Dis. 2019, 25, 980–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. U.S. Monkeypox Outbreak 2022: Situation Summary. 2022. Available online: https://stacks.cdc.gov/view/cdc/118618 (accessed on 14 June 2022).
- Damon, I.K. Status of human monkeypox: Clinical disease, epidemiology and research. Vaccine 2011, 29 (Suppl. 4), D54–D59. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.G.; McCollum, A.M.; Nguete, B.; Shongo Lushima, R.; Petersen, B.W. Improving the Care and Treatment of Monkeypox Patients in Low-Resource Settings: Applying Evidence from Contemporary Biomedical and Smallpox Biodefense Research. Viruses 2017, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Fox, E.J. Mechanism of action of mitoxantrone. Neurology 2004, 63 (Suppl. 6), S15–S18. [Google Scholar] [CrossRef]
- Deng, L.; Dai, P.; Ciro, A.; Smee, D.F.; Djaballah, H.; Shuman, S. Identification of novel antipoxviral agents: Mitoxantrone inhibits vaccinia virus replication by blocking virion assembly. J. Virol. 2007, 81, 13392–13402. [Google Scholar] [CrossRef] [Green Version]
- Altmann, S.E.; Smith, A.L.; Dyall, J.; Johnson, R.F.; Dodd, L.E.; Jahrling, P.B.; Paragas, J.; Blaney, J.E. Inhibition of cowpox virus and monkeypox virus infection by mitoxantrone. Antivir. Res. 2012, 93, 305–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Xia, D.; Syed, A.A.S.; Wang, Z.; Shi, Y. Highly accurate protein structure prediction and drug screen of monkeypox virus proteome. J. Infect. 2022. [CrossRef]
- Lam, H.Y.I.; Guan, J.S.; Mu, Y. In Silico Repurposed Drugs against Monkeypox Virus. Molecules 2022, 27, 5277. [Google Scholar] [CrossRef]
- Aiman, S.; Alhamhoom, Y.; Ali, F.; Rahman, N.; Rastrelli, L.; Khan, A.; Farooq, Q.U.; Ahmed, A.; Khan, A.; Li, C. Multi-epitope chimeric vaccine design against emerging Monkeypox virus via reverse vaccinology techniques- a bioinformatics and immunoinformatics approach. Front. Immunol. 2020, 13, 985450. [Google Scholar] [CrossRef] [PubMed]
- Abdi, S.A.H.; Ali, A.; Sayed, S.F.; Abutahir; Ali, A.; Alam, P. Multi-Epitope-Based Vaccine Candidate for Monkeypox: An In Silico Approach. Vaccines 2022, 10, 1564. [Google Scholar] [CrossRef] [PubMed]
- Poxviruses, M.B. Virology, 2nd ed.; Fields, B.N., Chanock, R.M., Hirsch, M.S., Melnick, J., Monath, T.P., Roizman, B., Eds.; Raven Press: New York, NY, USA, 1990; Volume 2, pp. 2079–2111. [Google Scholar]
- Caillat, C.; Topalis, D.; Agrofoglio, L.A.; Pochet, S.; Balzarini, J.; Deville-Bonne, D.; Meyer, P. Crystal structure of poxvirus thymidylate kinase: An unexpected dimerization has implications for antiviral therapy. Proc. Natl. Acad. Sci. USA 2008, 105, 16900–16905. [Google Scholar] [CrossRef] [Green Version]
- Minasov, G.; Shuvalova, L.; Dubrovska, I.; Flores, K.; Grimshaw, S.; Kwon, K.; Anderson, W.F.; Center for Structural Genomics of Infectious Diseases (CSGID). 1.52 Angstrom Crystal Structure of A42R Profilin-like Protein from Monkeypox Virus Zaire-96-I-16. Available online: https://www.wwpdb.org/pdb?id=pdb_00004qwo (accessed on 25 August 2022).
- Garriga, D.; Headey, S.; Accurso, C.; Gunzburg, M.; Scanlon, M.; Coulibaly, F. Structural basis for the inhibition of poxvirus assembly by the antibiotic rifampicin. Proc. Natl. Acad. Sci. USA 2018, 115, 8424–8429. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2020, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Wolber, G.; Thierry, L. LigandScout: 3-D pharmacophores derived from protein-bound ligands and their use as virtual screening filters. J. Chem. Inf. Model 2005, 45, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera- a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- SAMSON: Software for Adaptive Modeling and Simulation Of Nanosystems. Available online: https://www.samson-connect.net (accessed on 26 August 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (Pubchem ID) | Docking Score (−) (kcal/mol) | Docking Score (−) (kcal/mol) | Docking Score (−) (kcal/mol) |
|---|---|---|---|
| PDB ID: 2V54 | PDB ID: 4QWO | PDB ID: 6BED | |
| (Thymidylate Kinase) | (A42R Profilin) | (D13 Protein) | |
| MXN (4212) (Standard) | 6.9 | 6.7 | 7.3 |
| (1) (13276605) | 7.2 | 6.6 | 7.9 |
| (2) (153640723) | 7.2 | 6.8 | 8 |
| (3) (44275839) | 6.7 | 6.4 | 7.6 |
| (4) (71044822) | 6.9 | 6.6 | 7.3 |
| (5) (142963046) | 6.9 | 6.1 | 7.3 |
| (6) (44316536) | 6.8 | 6.2 | 7.5 |
| (7) (24848320) | 6.9 | 6.8 | 7.4 |
| (8) (59835539) | 7.2 | 6 | 7.9 |
| (9) (44541200) | 7 | 6.1 | 7.4 |
| (10) (58102019) | 7 | 6.8 | 7.5 |
| (11) (143270488) | 7.2 | 6.6 | 7.4 |
| (12) (145293737) | 7 | 6.8 | 8.3 |
| (13) (44541201) | 6.8 | 6.3 | 7.5 |
| Residues | MXN | Compound (2) | Compound (7) | Compound (10) | Compound (12) |
|---|---|---|---|---|---|
| Arg41(B) | x | x | xx | x | x |
| Arg93(B) | xx | x | xx | xx | xx |
| Asn37(B) | xx | xx | xx | x | xx |
| Asp13(B) | xx | xx | xx | xx | |
| Asp92(B) | xx | x | x | ||
| Glu142(B) | xx | xx | x | x | xx |
| Glu145(B) | x | xx | x | x | |
| Gly16(B) | x | x | x | x | |
| Leu53(B) | x | x | x | x | |
| Lys14(B) | x | xx | x | x | x |
| Lys17(B) | x | x | x | x | x |
| Phe38(B) | x | ||||
| Phe68(B) | x | x | |||
| Pro39 (B) | x | ||||
| Ser15(B) | xx | xx | xx | x | |
| Ser97(B) | xx | ||||
| Thr18(B) | xx | xx | x | xx | x |
| Tyr35(B) | x | ||||
| Tyr101(B) | x | x | x | x | |
| Tyr144(B) | x | x | x | x |
| Residues | MXN | Compound (2) | Compound (7) | Compound (10) | Compound (12) |
|---|---|---|---|---|---|
| Ala129(B) | x | x | x | ||
| Ala130(B) | x | x | x | x | |
| Arg119(A) | x | x | x | x | x |
| Arg127(A) | x | x | x | x | |
| Arg129(B) | x | ||||
| Asn14(A) | xx | xx | xx | xx | xx |
| Asn78(B) | x | x | x | x | x |
| Asp10(A) | x | x | x | x | x |
| Asp116(A) | x | x | x | x | x |
| Asp123(A) | x | x | x | x | x |
| Glu77(B) | x | xx | x | x | |
| His100(B) | x | x | |||
| Lys16(A) | x | x | x | x | x |
| Phe17(A) | x | x | x | x | x |
| Thr120(A) | xx | xx | x | xx | x |
| Thr126(B) | x | x | |||
| Tyr80(B) | x | x |
| Residues | MXN | Compound (2) | Compound (7) | Compound (10) | Compound (12) |
|---|---|---|---|---|---|
| Asn117(A) | x | xx | x | x | x |
| Asn118(A) | xx | x | x | x | xx |
| Asn121(A) | xx | x | xx | xx | xx |
| Asn464(A) | xx | xx | xx | xx | |
| Asn530(A) | xx | xx | x | x | x |
| Glu114(A) | x | xx | x | x | |
| Glu230(A) | x | xx | xx | ||
| Gly473(A) | x | ||||
| Ile110(A) | x | ||||
| Lys159(A) | xx | xx | xx | xx | |
| Lys484(A) | x | x | x | ||
| Pro161(A) | x | x | x | ||
| Pro232(A) | x | x | x | ||
| Ser254(A), | x | x | x | x | x |
| Ser256(A) | x | x | xx | xx | xx |
| Ser470(A) | x | ||||
| Thr468(A) | x | x | x | ||
| Thr474(A) | xx | xx | xx | x | |
| Thr464(A) | x | xx | |||
| Thr468(A) | x | x | |||
| Trp108(A) | xx | x | x | ||
| Val528(A) | x | x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preet, G.; Oluwabusola, E.T.; Milne, B.F.; Ebel, R.; Jaspars, M. Computational Repurposing of Mitoxantrone-Related Structures against Monkeypox Virus: A Molecular Docking and 3D Pharmacophore Study. Int. J. Mol. Sci. 2022, 23, 14287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214287
Preet G, Oluwabusola ET, Milne BF, Ebel R, Jaspars M. Computational Repurposing of Mitoxantrone-Related Structures against Monkeypox Virus: A Molecular Docking and 3D Pharmacophore Study. International Journal of Molecular Sciences. 2022; 23(22):14287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214287
Chicago/Turabian StylePreet, Gagan, Emmanuel T. Oluwabusola, Bruce Forbes Milne, Rainer Ebel, and Marcel Jaspars. 2022. "Computational Repurposing of Mitoxantrone-Related Structures against Monkeypox Virus: A Molecular Docking and 3D Pharmacophore Study" International Journal of Molecular Sciences 23, no. 22: 14287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214287