
Increased Prevalence of Unstable HLA-C Variants in HIV-1 Rapid-Progressor Patients

 , ,
, ,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Studied Patient’s Cohort
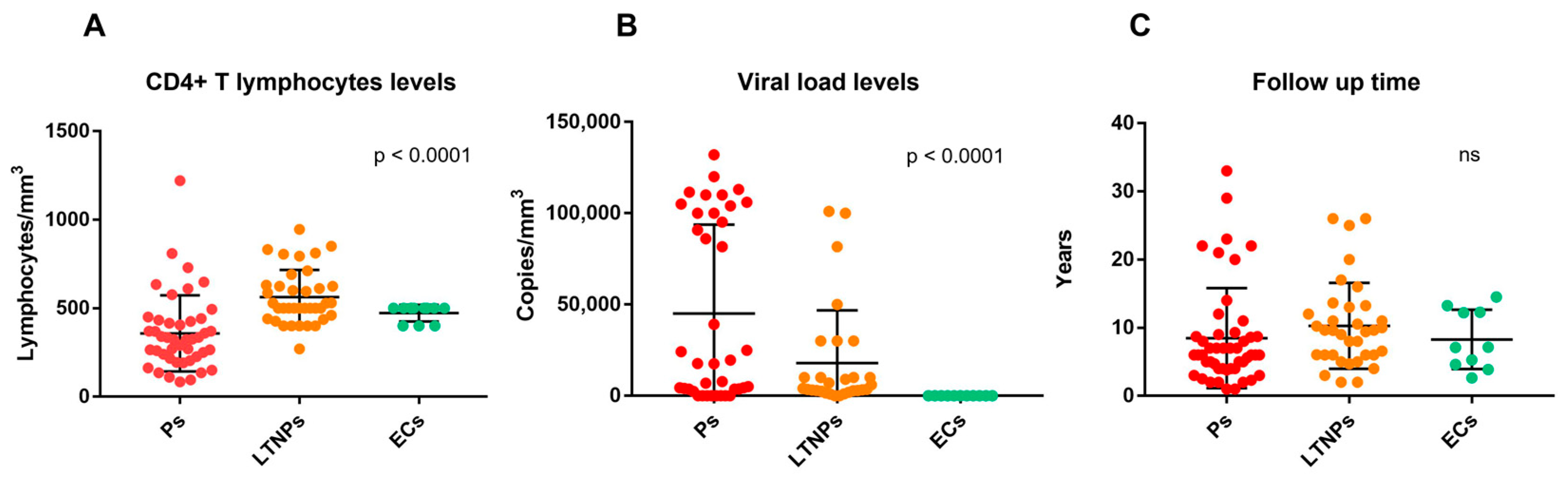
2.2. Immuno-Virological Characteristics of the Studied Cohort
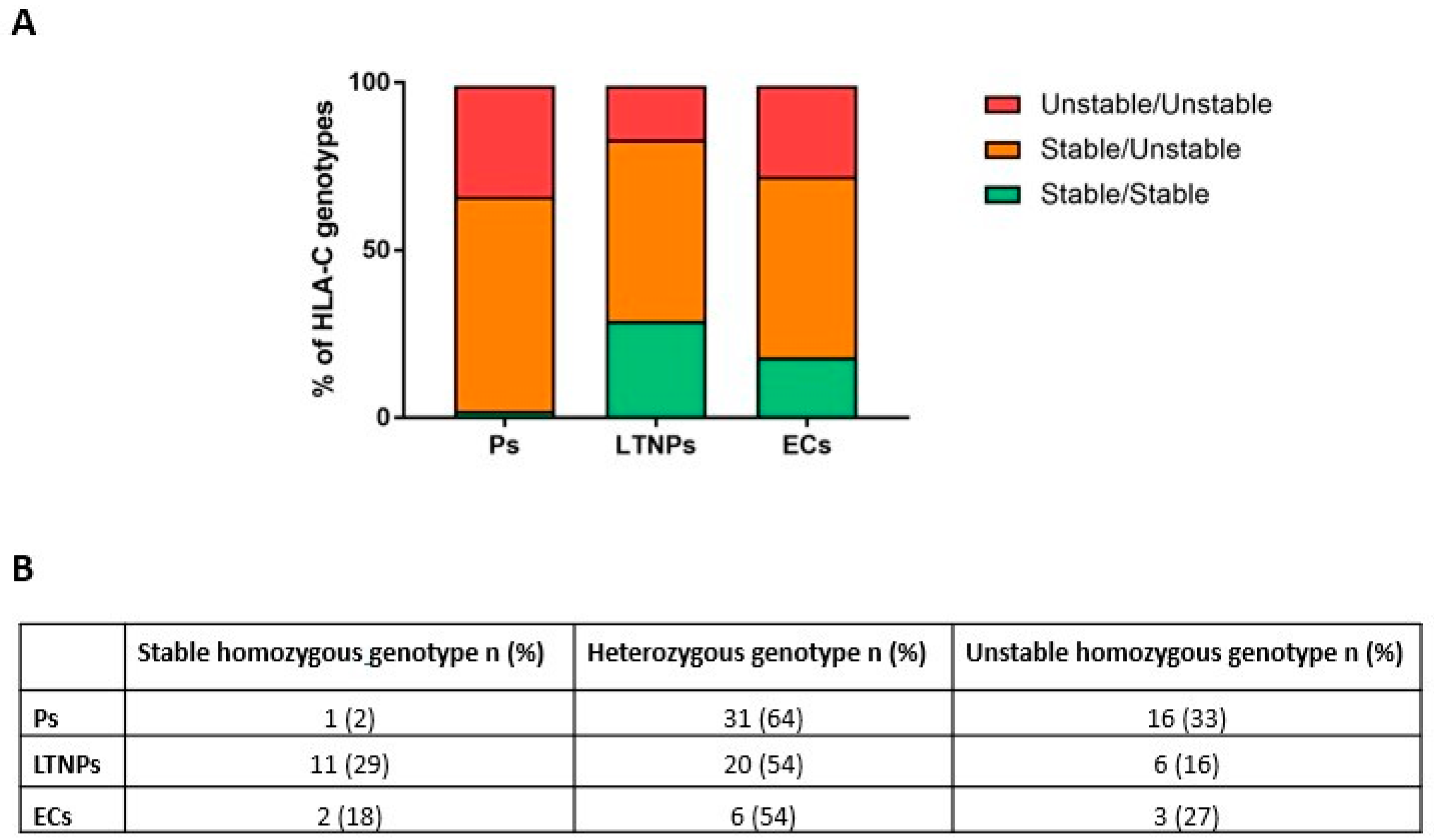
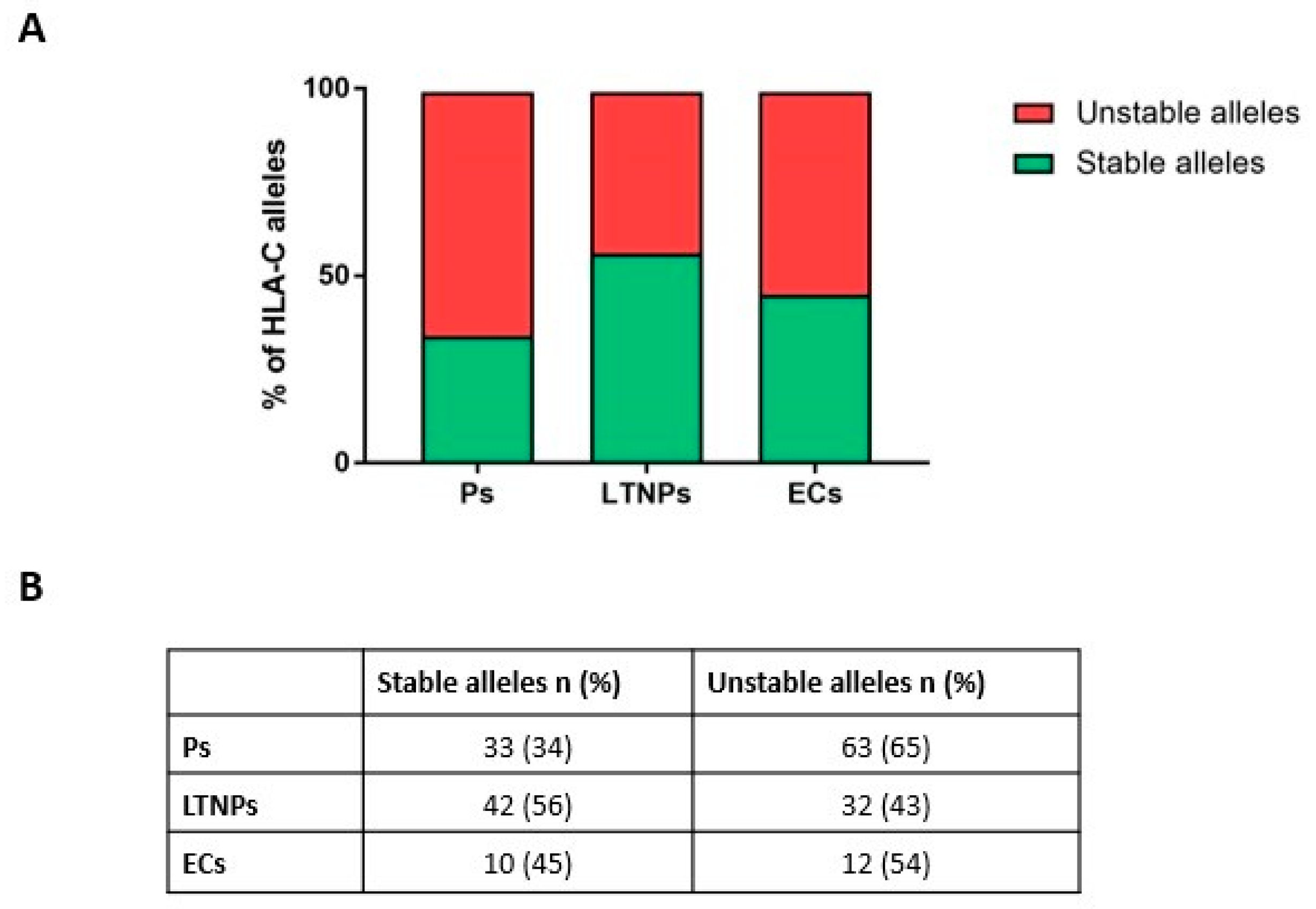
2.3. Unstable HLA-C Variants Correlate with HIV-1 Progression
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. DNA Extraction and HLA-C Genotyping
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallo, R.C.; Salahuddin, S.Z.; Popovic, M.; Shearer, G.M.; Kaplan, M.; Haynes, B.F.; Palker, T.J.; Redfield, R.; Oleske, J.; Safai, B.; et al. Frequent Detection and Isolation of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and at Risk for AIDS. Science 1984, 224, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Clapham, P.R.; McKnight, Á. HIV-1 Receptors and Cell Tropism. Br. Med. Bull. 2001, 58, 43–59. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. Global HIV & AIDS Statistics—Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 15 December 2021).
- Hemelaar, J. The Origin and Diversity of the HIV-1 Pandemic. Trends Mol. Med. 2012, 18, 182–192. [Google Scholar] [CrossRef]
- Kaur, G.; Mehra, N. Genetic Determinants of HIV-1 Infection and Progression to AIDS: Susceptibility to HIV Infection. Tissue Antigens 2009, 73, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Mbondji-Wonje, C.; Dong, M.; Zhao, J.; Wang, X.; Nanfack, A.; Ragupathy, V.; Sanchez, A.M.; Denny, T.N.; Hewlett, I. Genetic Variability of the U5 and Downstream Sequence of Major HIV-1 Subtypes and Circulating Recombinant Forms. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Muñoz, M.A.; Restrepo, C.; Benito, J.M.; Rallón, N. Elite Controllers: A Heterogeneous Group of HIV-Infected Patients. Virulence 2020, 11, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Mandalia, S.; Westrop, S.J.; Beck, E.J.; Nelson, M.; Gazzard, B.G.; Imami, N. Are Long-Term Non-Progressors Very Slow Progressors? Insights from the Chelsea and Westminster HIV Cohort, 1988–2010. PLoS ONE 2012, 7, e29844. [Google Scholar] [CrossRef] [Green Version]
- Olson, A.D.; Guiguet, M.; Zangerle, R.; Gill, J.; Perez-Hoyos, S.; Lodi, S.; Ghosn, J.; Dorrucci, M.; Johnson, A.; Sannes, M.; et al. Evaluation of Rapid Progressors in HIV Infection as an Extreme Phenotype. J. Acquir. Immune. Defic. Syndr. 2014, 67, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Kulpa, D.A.; Collins, K.L. The Emerging Role of HLA-C in HIV-1 Infection. Immunology 2011, 134, 116. [Google Scholar] [CrossRef]
- Biassoni, R.; Falco, M.; Cambiaggi, A.; Costa, P.; Verdiani, S.; Pende, D.; Conte, R.; Di Donato, C.; Parharn, P.; Moretta, L. Amino Acid Substitutions Can Influence the Natural Killer (Nk)-Mediated Recognition of Hla-c Molecules. Role of Serine-77 and Lysine-80 in the Target Cell Protection from Lysis Mediated by “Group 2” or “Group 1” Nk Clones. J. Exp. Med. 1995, 182, 605–609. [Google Scholar] [CrossRef]
- Körner, C.; Simoneau, C.R.; Schommers, P.; Granoff, M.; Ziegler, M.; Hölzemer, A.; Lunemann, S.; Chukwukelu, J.; Corleis, B.; Naranbhai, V.; et al. HIV-1-Mediated Downmodulation of HLA-C Impacts Target Cell Recognition and Antiviral Activity of NK Cells. Cell Host Microbe 2017, 22, 111–119.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipeto, D.; Beretta, A. HLA-C and HIV-1: Friends or Foes? Retrovirology 2012, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Costello, C.; Keet, I.P.M.; Rivers, C.; Leblanc, S.; Karita, E.; Allen, S.; Kaslow, R.A. HLA Class I Homozygosity Accelerates Disease Progression in Human Immunodeficiency Virus Type 1 Infection. AIDS Res. Hum. Retroviruses 1999, 15, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Carrington, M.; Nelson, G.W.; Martin, M.P.; Kissner, T.; Vlahov, D.; Goedert, J.J.; Kaslow, R.; Buchbinder, S.; Hoots, K.; O’Brien, S.J. HLA and HIV-1: Heterozygote Advantage and B*35-Cw*04 Disadvantage. Science 1999, 283, 1748–1752. [Google Scholar] [CrossRef]
- Neefjes, J.J.; Ploegh, H.L. Allele and Locus-Specific Differences in Cell Surface Expression and the Association of HLA Class I Heavy Chain with Β2-Microglobulin: Differential Effects of Inhibition of Glycosylation on Class I Subunit Association. Eur. J. Immunol. 1988, 18, 801–810. [Google Scholar] [CrossRef]
- Apps, R.; Del Prete, G.Q.; Chatterjee, P.; Lara, A.; Brumme, Z.L.; Brockman, M.A.; Neil, S.; Pickering, S.; Schneider, D.K.; Piechocka-Trocha, A.; et al. HIV-1 Vpu Mediates HLA-C Downregulation. Cell Host Microbe 2016, 19, 686. [Google Scholar] [CrossRef] [Green Version]
- Geyer, M.; Fackler, O.T.; Peterlin, B.M. Structure--Function Relationships in HIV-1 Nef. EMBO Rep. 2001, 2, 580–585. [Google Scholar] [CrossRef]
- Langer, S.; Hammer, C.; Hopfensperger, K.; Klein, L.; Hotter, D.; Jesus, P.D.D.; Herbert, K.M.; Pache, L.; Smith, N.; Der Merwe, J.A.V.; et al. HIV-1 Vpu Is a Potent Transcriptional Suppressor of NF-ΚB-Elicited Antiviral Immune Responses. Elife 2019, 8, e41930. [Google Scholar] [CrossRef]
- Xu, Z.; Lodge, R.; Power, C.; Cohen, E.A.; Hobman, T.C. The HIV-1 Accessory Protein Vpu Downregulates Peroxisome Biogenesis. MBio 2020, 11, e03395-19. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.; Savan, R.; Qi, Y.; Gao, X.; Yuki, Y.; Bass, S.E.; Martin, M.P.; Hunt, P.; Deeks, S.G.; Telenti, A.; et al. Differential MicroRNA Regulation of HLA-C Expression and Its Association with HIV Control. Nature 2011, 472, 495. [Google Scholar] [CrossRef]
- Malnati, M.S.; Biswas, P.; Ugolotti, E.; Di Marco, E.; Sironi, F.; Parolini, F.; Garbarino, L.; Mazzocco, M.; Zipeto, D.; Biassoni, R. A Fast and Reliable Method for Detecting SNP Rs67384697 (Hsa-MiR-148a Binding Site) by a Single Run of Allele-Specific Real-Time PCR. HLA 2020, 96, 312–322. [Google Scholar] [CrossRef] [PubMed]
- O’Huigin, C.; Kulkarni, S.; Xu, Y.; Deng, Z.; Kidd, J.; Kidd, K.; Gao, X.; Carrington, M. The Molecular Origin and Consequences of Escape from MiRNA Regulation by HLA-C Alleles. Am. J. Hum. Genet. 2011, 89, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Sibilio, L.; Martayan, A.; Setini, A.; Monaco, E.L.; Tremante, E.; Butler, R.H.; Giacomini, P. A Single Bottleneck in HLA-C Assembly. J. Biol. Chem. 2008, 283, 1267–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolini, F.; Biswas, P.; Serena, M.; Sironi, F.; Muraro, V.; Guizzardi, E.; Cazzoletti, L.; Scupoli, M.T.; Gibellini, D.; Ugolotti, E.; et al. Stability and Expression Levels of HLA-C on the Cell Membrane Modulate HIV-1 Infectivity. J. Virol. 2018, 92, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serena, M.; Parolini, F.; Biswas, P.; Sironi, F.; Blanco Miranda, A.; Zoratti, E.; Scupoli, M.T.; Ziglio, S.; Valenzuela-Fernandez, A.; Gibellini, D.; et al. HIV-1 Env Associates with HLA-C Free-Chains at the Cell Membrane Modulating Viral Infectivity. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Song, M.J.; Lee, C.W.; Kim, J.H.; Lee, S.J.; Kim, C.J.; Hur, S.Y.; Park, T.C.; Kim, T.G.; Park, J.S. Association of KIR Genes and HLA-C Alleles with HPV-Related Uterine Cervical Disease in Korean Women. Tissue Antigens 2013, 81, 164–170. [Google Scholar] [CrossRef]
- Ursu, L.; Calenic, B.; Diculescu, M.; Dima, A.; Constantinescu, I. HLA Alleles and KIR Genes in Romanian Patients with Chronic Hepatitis C. J. Gastrointestin. Liver Dis. 2020, 29, 595–601. [Google Scholar] [CrossRef]
- Yengo, C.K.; Torimiro, J.; Kowo, M.; Lebon, P.A.; Tiedeu, B.A.; Luma, H.; Njoya, O.; Rowland-Jones, S.; Yindom, L.M. Variation of HLA Class I (-A and -C) Genes in Individuals Infected with Hepatitis B or Hepatitis C Virus in Cameroon. Heliyon 2020, 6, e05232. [Google Scholar] [CrossRef]
- Zipeto, D.; Serena, M.; Mutascio, S.; Parolini, F.; Diani, E.; Guizzardi, E.; Muraro, V.; Lattuada, E.; Rizzardo, S.; Malena, M.; et al. HIV-1-Associated Neurocognitive Disorders: Is HLA-C Binding Stability to Β2-Microglobulin a Missing Piece of the Pathogenetic Puzzle? Front. Neurol. 2018, 9, 791. [Google Scholar] [CrossRef] [Green Version]
- Apps, R.; Qi, Y.; Carlson, J.M.; Chen, H.; Gao, X.; Thomas, R.; Yuki, Y.; Del Prete, G.Q.; Goulder, P.; Brumme, Z.L.; et al. Influence of HLA-C Expression Level on HIV Control. Science 2013, 340, 87. [Google Scholar] [CrossRef]
- Bunce, M.; O’Neill, C.M.; Barnardo, M.C.N.M.; Krausa, P.; Browning, M.J.; Morris, P.J.; Welsh, K.I. Phototyping: Comprehensive DNA Typing for HLA-A, B, C, DRB1, DRB3, DRB4, DRB5 & DQB1 by PCR with 144 Primer Mixes Utilizing Sequence-Specific Primers (PCR-SSP). Tissue Antigens 1995, 46, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Bunce, M.; Barnardo, M.C.N.M.; Procter, J.; Marsh, S.G.E.; Vilches, C.; Welsh, K.I. High Resolution HLA-C Typing by PCR-SSP: Identification of Allelic Frequencies and Linkage Disequilibria in 604 Unrelated Random UK Caucasoids and a Comparison with Serology. Tissue Antigens 1996, 48, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, D.S.; Pasquinelli, A.E. BstUI and DpnII RFLPs at the COL5A1 Gene. Hum. Mol. Genet. 1994, 3, 385. [Google Scholar] [CrossRef] [PubMed]
- Sangalli, A.; Malerba, G.; Tessari, G.; Rodolfo, M.; Gomez-Lira, M. Melanoma Risk Alleles Are Associated with Downregulation of the MTAP Gene and Hypermethylation of a CpG Island Upstream of the Gene in Dermal Fibroblasts. Exp. Dermatol. 2017, 26, 733–736. [Google Scholar] [CrossRef] [Green Version]
- Lazaro, A.; Tu, B.; Yang, R.; Xiao, Y.; Kariyawasam, K.; Ng, J.; Hurley, C.K. Human Leukocyte Antigen (HLA) Typing by DNA Sequencing. Methods Mol. Biol. 2013, 1034, 161–195. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ps (n = 48) * | LTNPs (n = 37) * | ECs (n = 11) * | p-Value | ||
|---|---|---|---|---|---|
| Age, median (IQR), years | 40.5 (30–52.5) | 43 (33.5–52.5) | 33 (27–41) | Ps vs. LTNPs p = 0.438 Ps vs. ECs p = 0.112 ECs vs. LTNPs p = 0.0143 | |
| Male, n (%) | 37 (77) | 27 (72.9) | 7 (63.6) | p = 0.6471 | |
| Female, n (%) | 11 (23) | 10 (27.1) | 4 (36.4) | ||
| Ethnicity, n (%) | Caucasian | 44 (91.6) | 31 (83.8) | 7 (63.6) | p = 0.0558 |
| Black | 4 (8.4) | 6 (16.2) | 4 (36.4) | ||
| Origin, n (%) | Brazil | 28 (58.4) | 19 (51.4) | - | p = 0.0009 |
| Canada | 14 (37.8) | 14 (37.8) | 11 (100) | ||
| USA | 6 (12.5) | 4 (10.8) | - |
| Stable Alleles | Unstable Alleles | |
|---|---|---|
| Brazil | 37 | 57 |
| Canada | 37 | 41 |
| USA | 11 | 9 |
| Ps (n = 44) * | LTNPs (n = 37) * | ECs (n = 11) * | p-Value | |
| CD4, median (IQR), lymphocytes/mm3 | 324 (219.5–431) | 500 (451–627) | 500 (400–500) | Ps vs. LTNPs p < 0.0001 Ps vs. ECs p = 0.0017 ECs vs. LTNPs p = 0.0528 |
| Ps (n = 39) * | LTNPs (n = 29) * | ECs (n = 11) * | ||
| Viral load, median (IQR), copies/mm3 | 17,698 (3688–100,000) | 4119 (2691–20,000) | 50 (50–50) | Ps vs. LTNPs p = 0.0773 Ps vs. ECs p < 0.0001 ECs vs. LTNPs p < 0.0001 |
| Ps (n = 48) * | LTNPs (n = 35) * | ECs (n = 10) * | ||
| Follow up, median (IQR), years | 6 (4–8.9) | 9.6 (6–13) | 7.14 (4.4–12.5) | Ps vs. LTNPs p = 0.0235 Ps vs. ECs p = 0.505 ECs vs. LTNPs p = 0.513 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefani, C.; Sangalli, A.; Locatelli, E.; Federico, T.; Malerba, G.; Romanelli, M.G.; Argañaraz, G.A.; Da Silva, B.C.M.; Da Silva, A.J.D.; Casseb, J.; et al. Increased Prevalence of Unstable HLA-C Variants in HIV-1 Rapid-Progressor Patients. Int. J. Mol. Sci. 2022, 23, 14852. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314852
Stefani C, Sangalli A, Locatelli E, Federico T, Malerba G, Romanelli MG, Argañaraz GA, Da Silva BCM, Da Silva AJD, Casseb J, et al. Increased Prevalence of Unstable HLA-C Variants in HIV-1 Rapid-Progressor Patients. International Journal of Molecular Sciences. 2022; 23(23):14852. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314852
Chicago/Turabian StyleStefani, Chiara, Antonella Sangalli, Elena Locatelli, Tania Federico, Giovanni Malerba, Maria Grazia Romanelli, Gustavo Adolfo Argañaraz, Bosco Christiano Maciel Da Silva, Alberto Jose Duarte Da Silva, Jorge Casseb, and et al. 2022. "Increased Prevalence of Unstable HLA-C Variants in HIV-1 Rapid-Progressor Patients" International Journal of Molecular Sciences 23, no. 23: 14852. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314852