
Acute Stress Induces Different Changes on the Expression of BDNF and trkB in the Mesocorticolimbic System of Two Lines of Rats Differing in Their Response to Stressors
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Behavioral Measures during FS
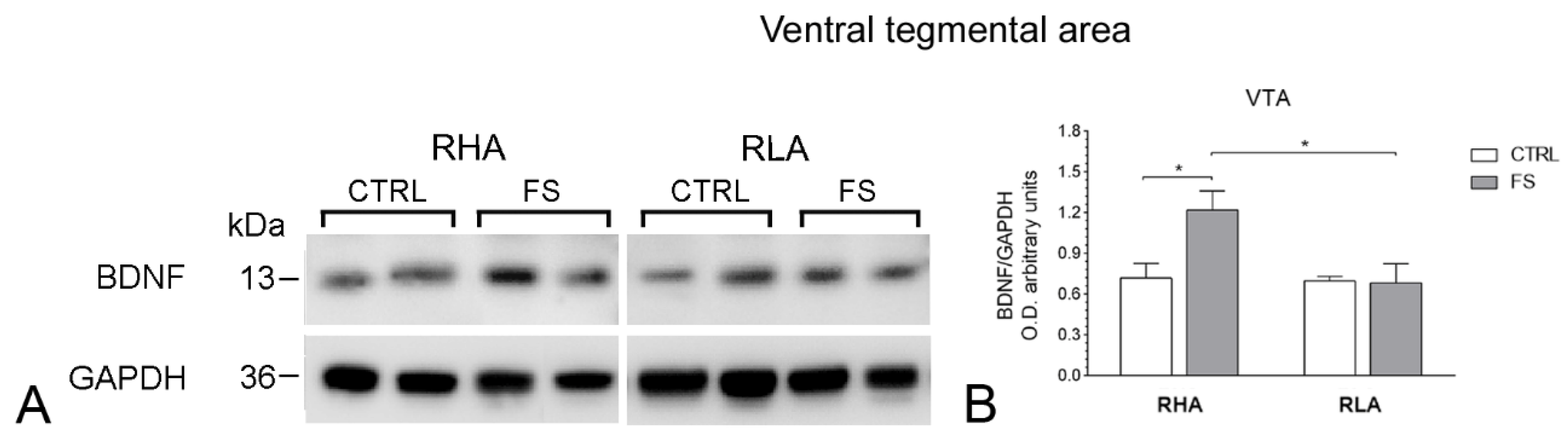
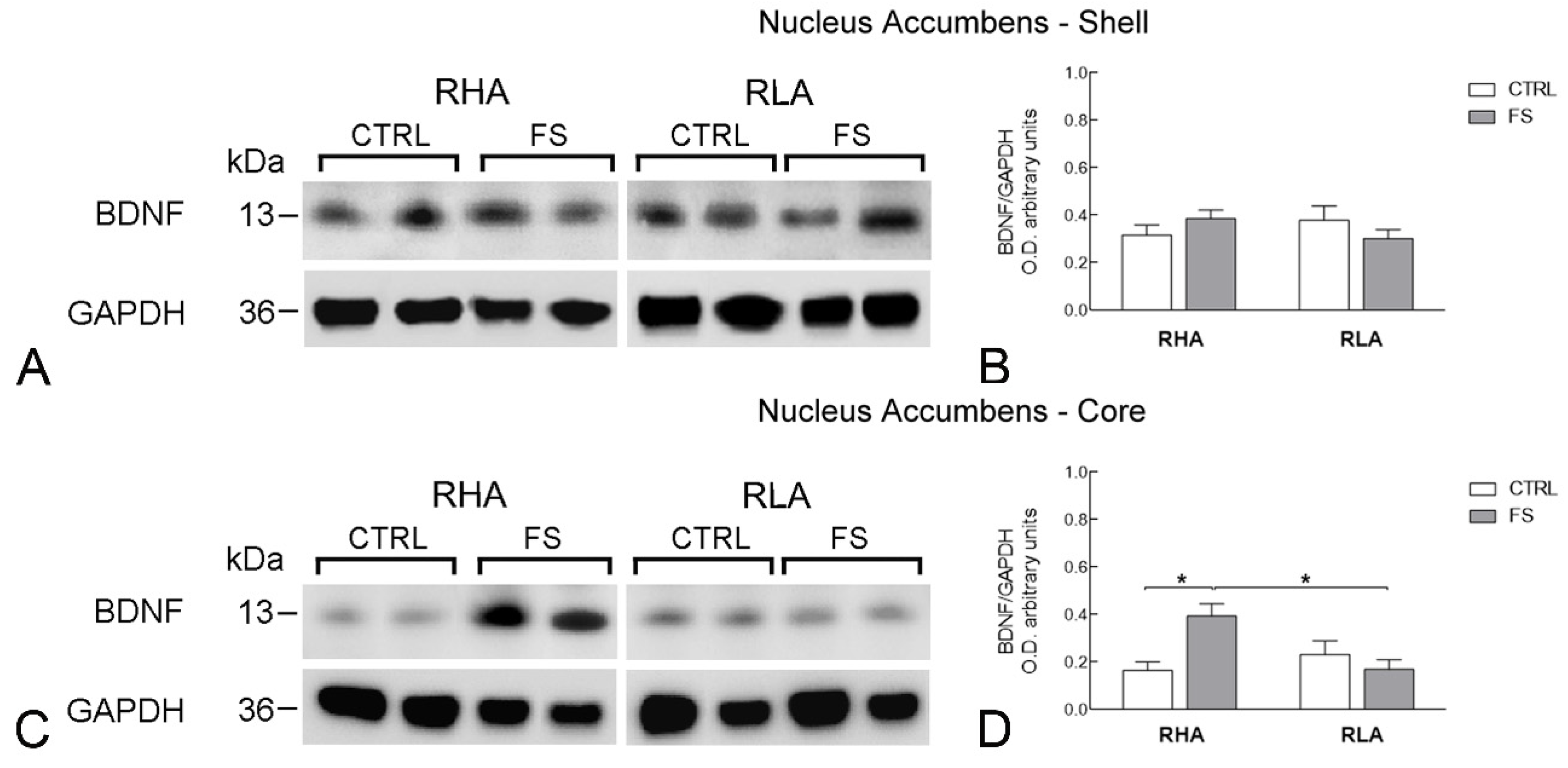
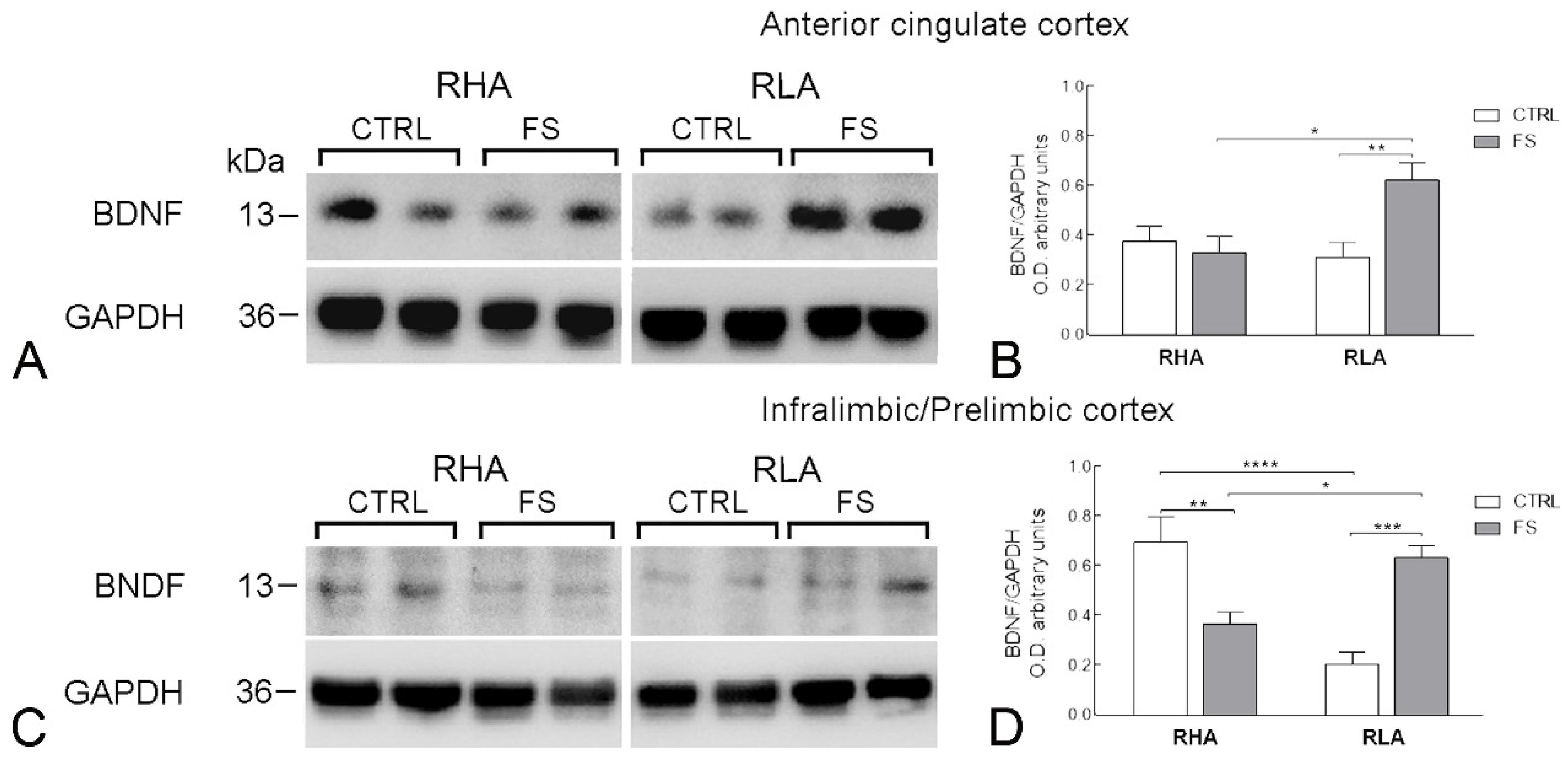
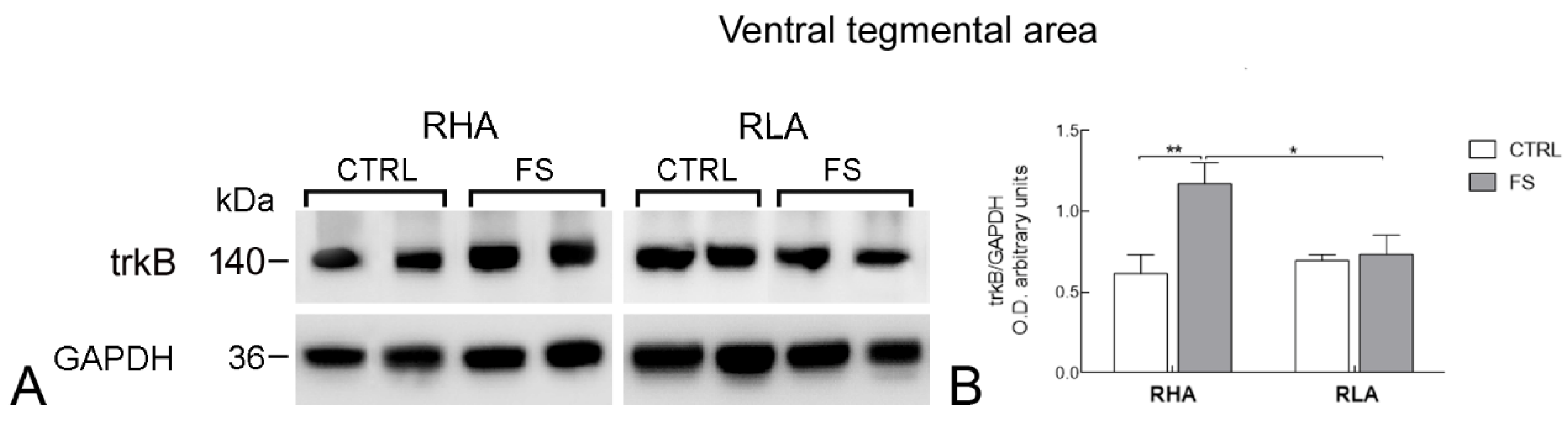
2.2. Western Blot
2.2.1. BDNF Protein Levels
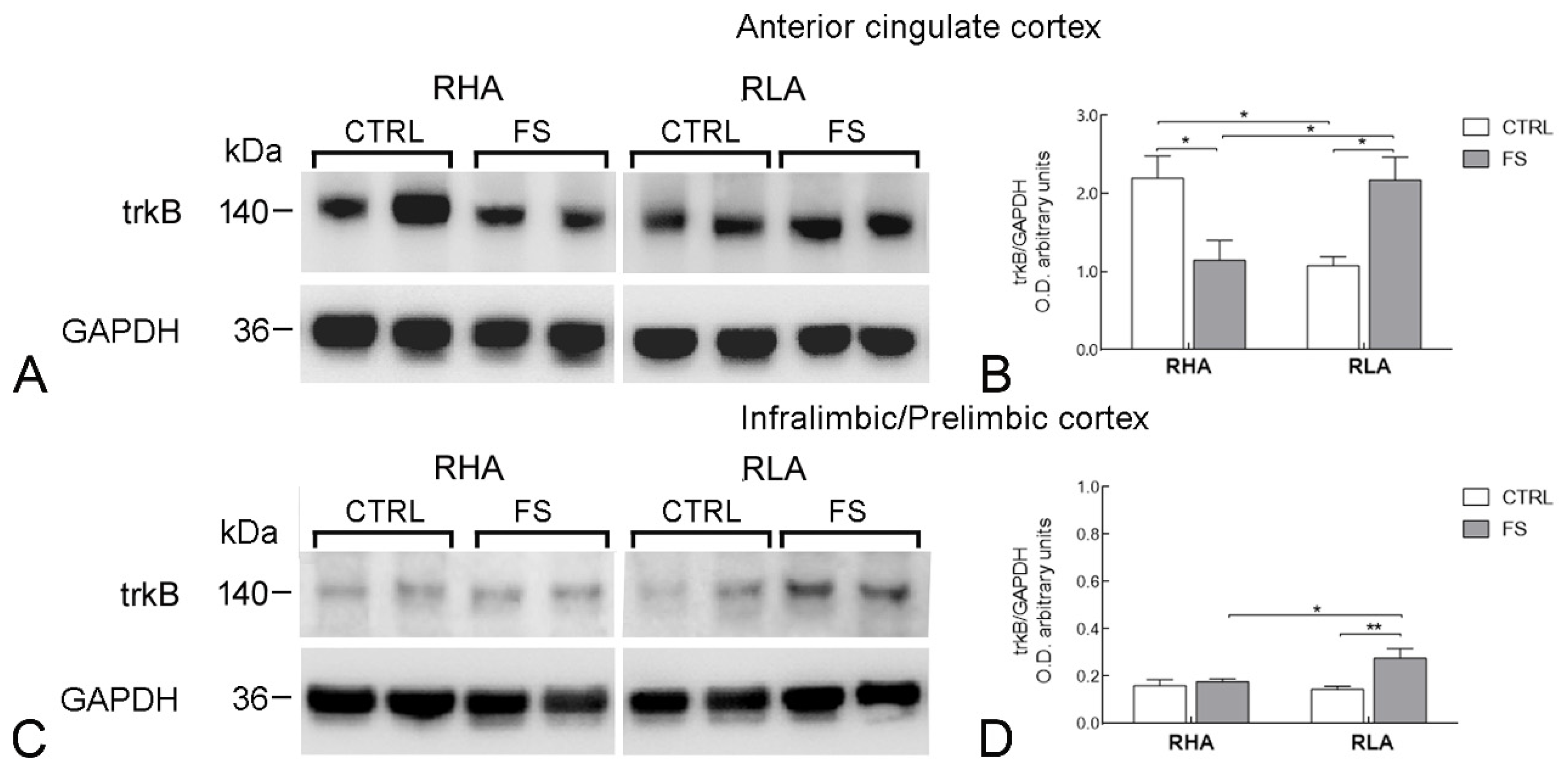
2.2.2. trkB Protein Levels
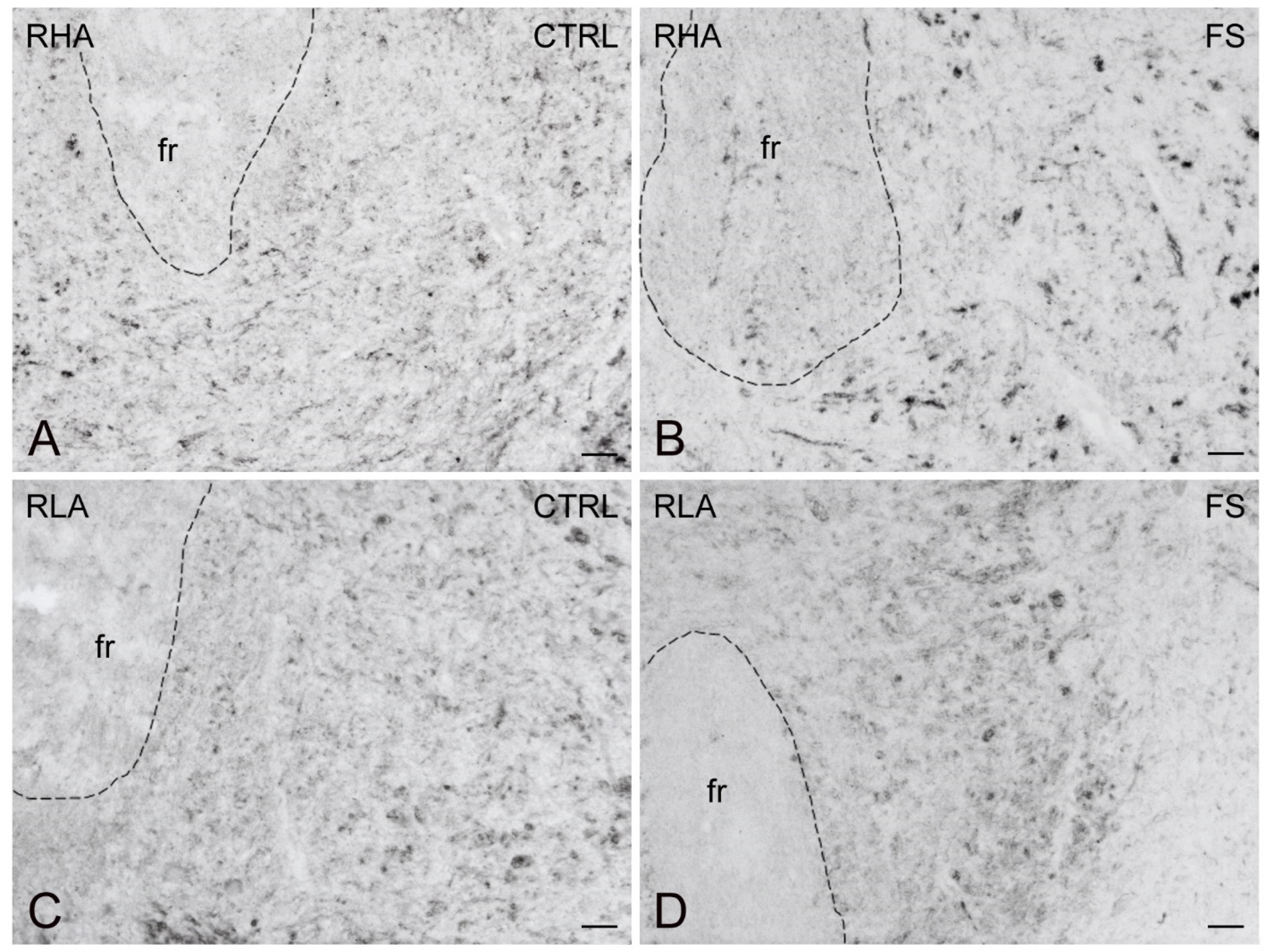
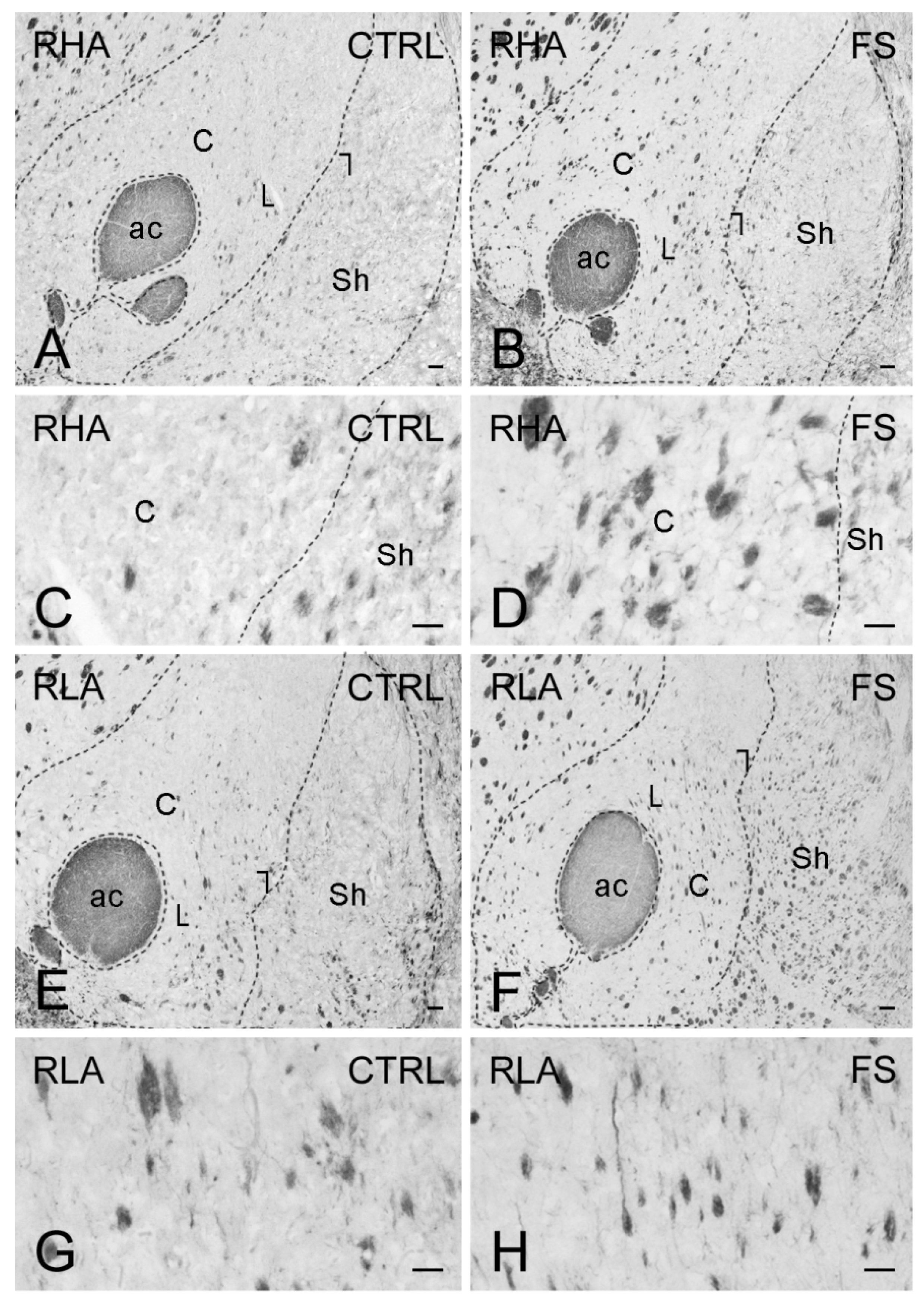
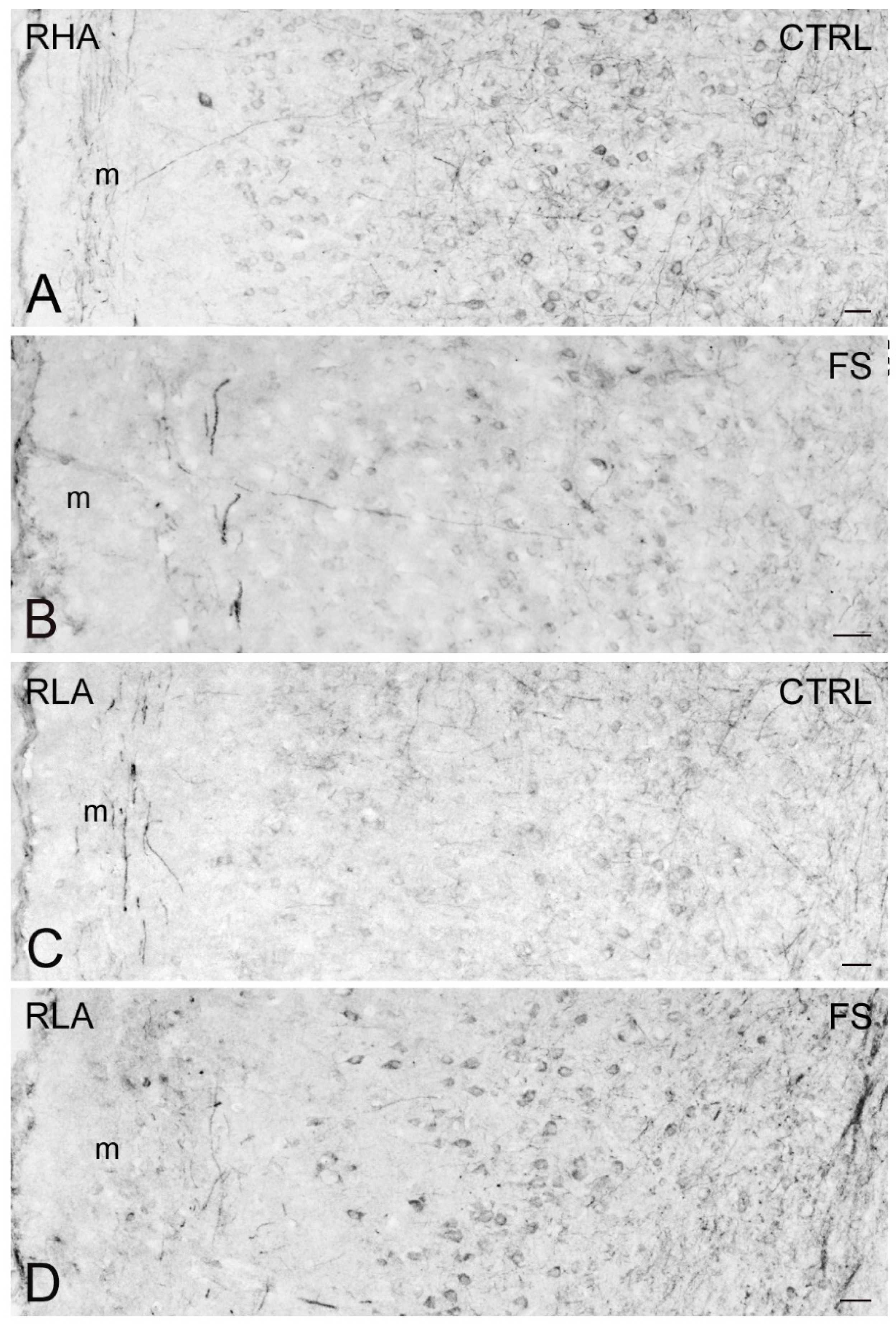
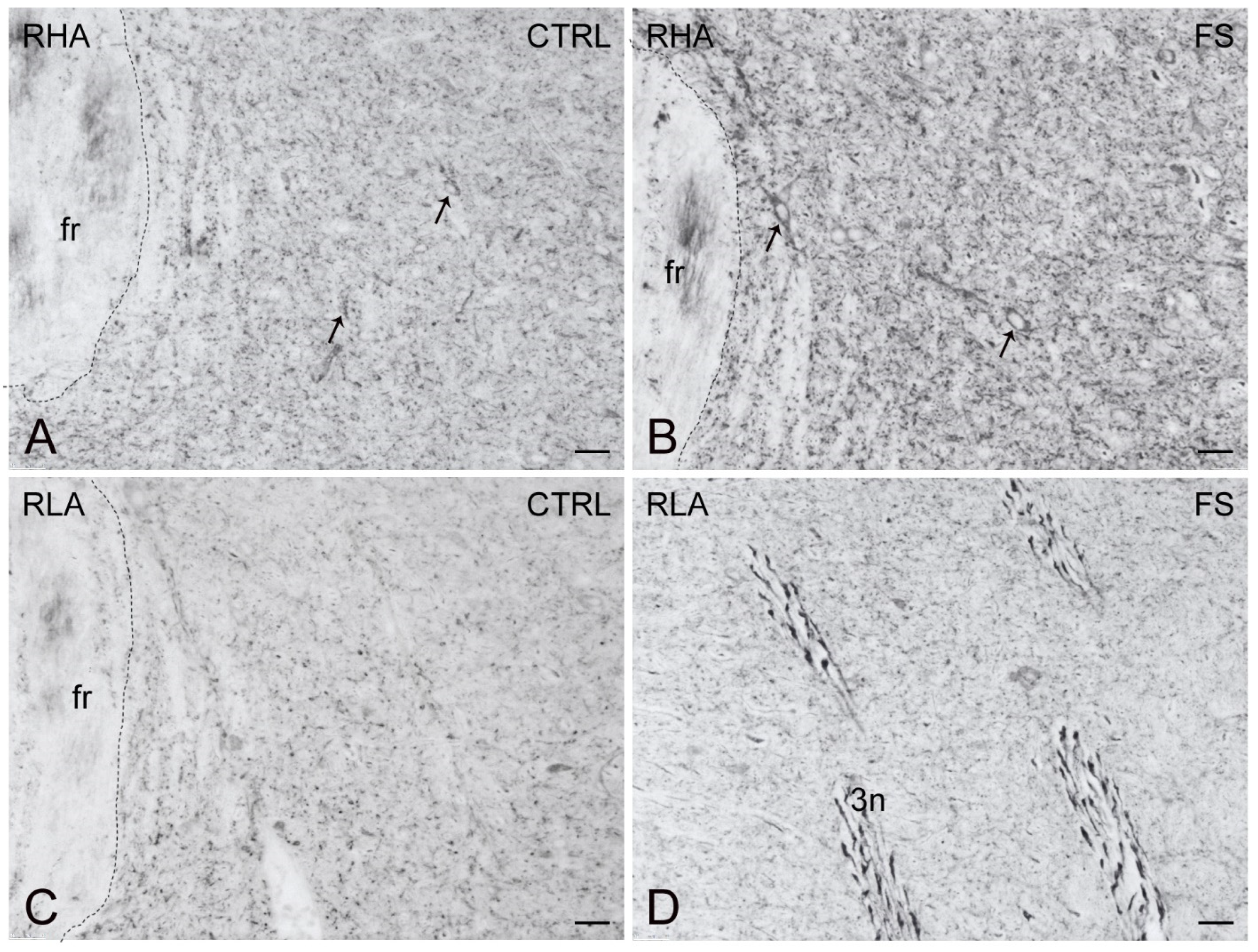
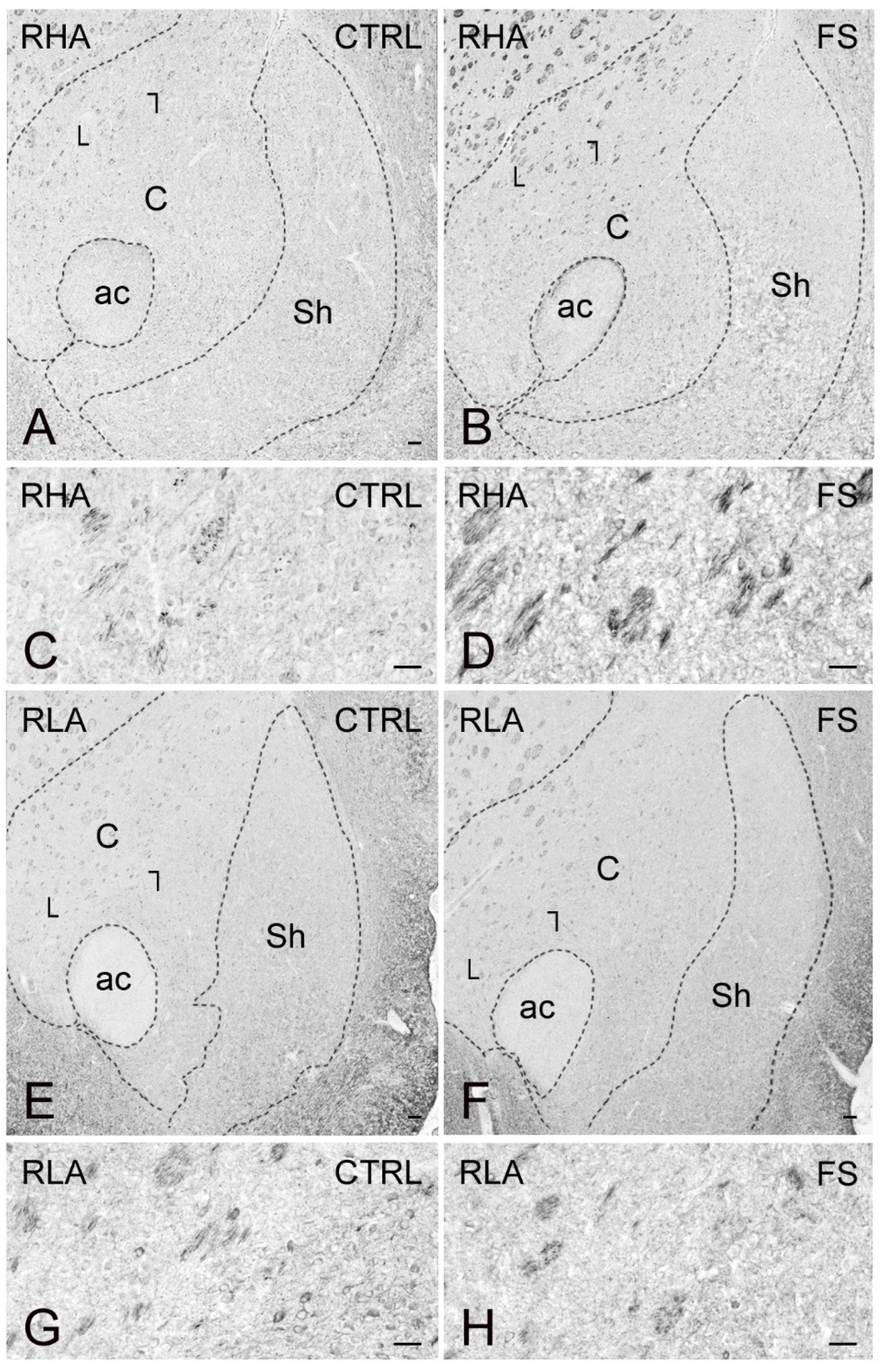
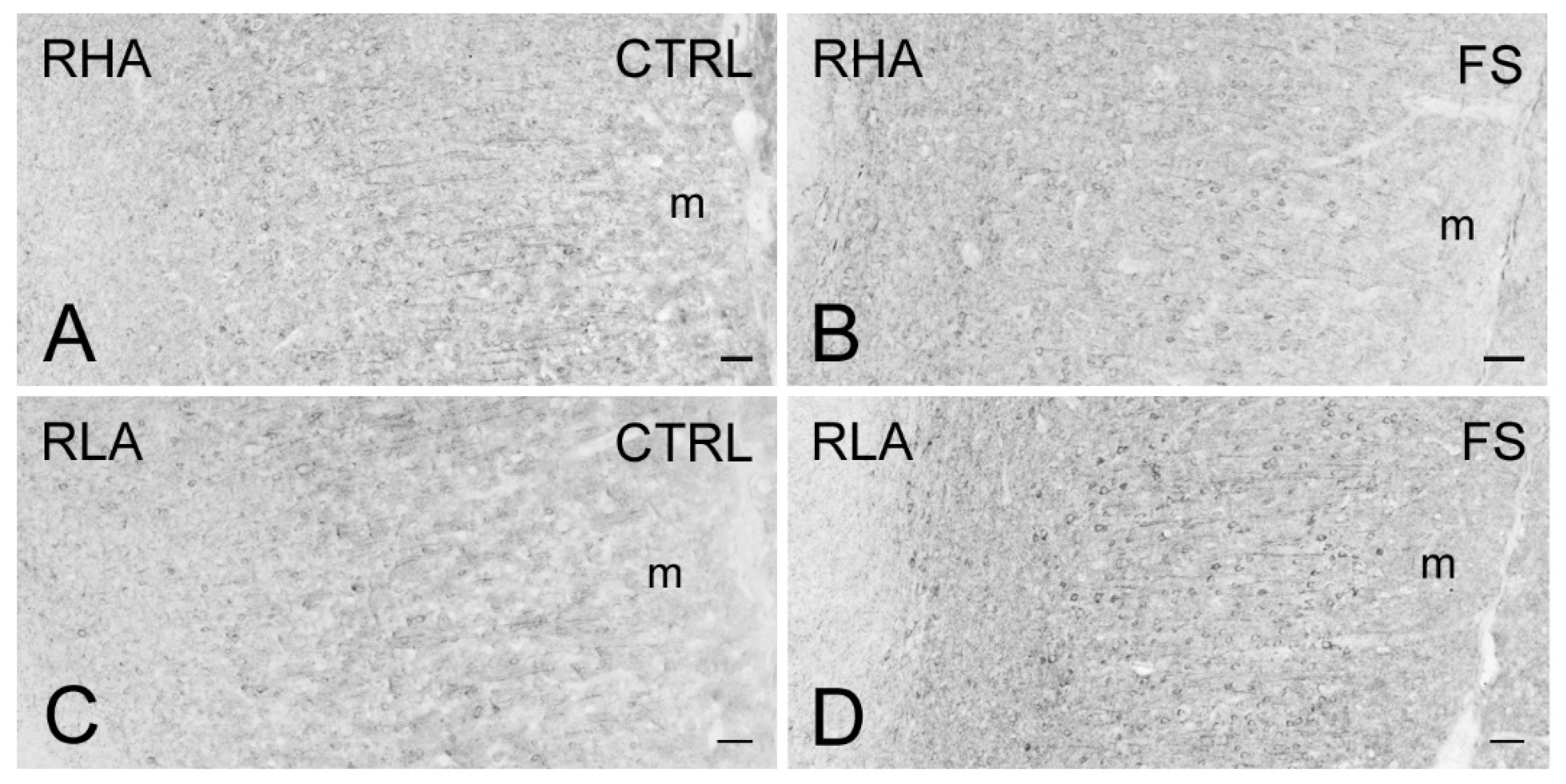
2.3. Immunohistochemistry
3. Discussion
3.1. RLA and RHA Rats Display Different Coping Strategies in Response to FS
3.2. Role of the BDNF/trkB System and the Dopaminergic Mesocorticolimbic Pathways in Stress-Induced Depression-Like Phenotypes
3.3. Differential Effects of Stress on BDNF and trkB Expression in the Mesolimbic Pathway of RLA and RHA Rats
3.4. Differential Effects of Stress on BDNF and trkB Expression in the Mesocortical Pathway of RLA and RHA Rats
3.5. Final Remarks
4. Materials and Methods
4.1. Animals
4.2. FS and Behavioral Measurements
4.3. Sampling
4.4. Western Blot
4.5. Immunohistochemistry
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | avidin–biotin–peroxidase complex |
| Acb | nucleus accumbens |
| ACg | anterior cingulate |
| AMYG | amygdala |
| ANOVA | analysis of variance |
| BDNF | brain derived neurotrophic factor |
| CSDS | chronic social defeat stress |
| CUS | chronic unpredictable stress |
| DA | dopamine |
| dHC | dorsal hippocampus |
| FS | forced swimming |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HC | hippocampus |
| IL | infralimbic |
| LI | prelimbic |
| mw | molecular weight |
| mTORC1 | mammalian target of rapamycin complex 1 |
| O.D. | relative optical density |
| PBS | phosphate buffered saline |
| PFA | paraformaldehyde |
| PFC | prefrontal cortex |
| PL | prelimbic |
| RHA | roman high avoidance |
| RLA | roman low avoidance |
| SDS-PAGE | sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| TBS-T | tris base, sodium chloride, tween 2 |
| trkB | tyrosine receptor kinase b |
| vHC | ventral hippocampus |
| VTA | ventral tegmental area |
| WB | western blot |
References
- Schildkraut, J.J. The catecholamine hypothesis of affective disorders: A review of supporting evidence. Am. J. Psychiatry 1965, 122, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Holsboer, F. Antidepressant drug discovery in the postgenomic era. World J. Biol. Psychiatry 2001, 2, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H. Depression and immunity: A role for T cells? Brain Behav. Immun. 2010, 24, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Malberg, J.; Thome, J. Neural plasticity to stress and antidepressant treatment. Biol. Psychiatry 1999, 46, 1181–1191. [Google Scholar] [CrossRef]
- Nestler, E.J.; Gould, E.; Manji, H.; Buncan, M.; Duman, R.S.; Greshenfeld, H.K.; Hen, R.; Koester, S.; Lederhendler, I.; Meaney, M.; et al. Preclinical models: Status of basic research in depression. Biol. Psychiatry 2002, 52, 503–528. [Google Scholar] [CrossRef]
- Koo, J.W.; Chaudhury, D.; Han, M.-H.; Nestler, E.J. Role of Mesolimbic Brain-Derived Neurotrophic Factor in Depression. Biol. Psychiatry 2019, 86, 738–748. [Google Scholar] [CrossRef]
- Fanous, S.; Hammer, R.P., Jr.; Nikulina, E.M. Short- and long-term effects of intermittent social defeat stress on brain-derived neurotrophic factor expression in mesocorticolimbic brain regions. Neuroscience 2010, 167, 598–607. [Google Scholar] [CrossRef] [Green Version]
- Shirayama, Y.; Chen, A.C.; Nakagawa, S.; Russell, D.S.; Duman, R.S. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J. Neurosci. 2002, 22, 223251–223261. [Google Scholar] [CrossRef] [Green Version]
- Eisch, A.J.; Bolaños, C.A.; de Wit, J.; Simonak, R.D.; Pudiak, C.M.; Barrot, M.; Verhaagen, J.; Nestler, E.J. Brain-derived neurotrophic factor in the ventral midbrain-nucleus accumbens pathway: A role in depression. Biol. Psychiatry 2003, 54, 994–1005. [Google Scholar] [CrossRef]
- Broadhurt, P.L.; Bignami, G. Correlative effects of psychogenetic selection: A study of the Roman high and low avoidance strains of rats. Behav. Res. Ther. 1965, 3, 273–280. [Google Scholar] [CrossRef]
- Driscoll, P.; Bättig, K. Behavioral, emotional and neurochemical profiles of rats selected for extreme differences in active, two-way avoidance performance. In Genetics of the Brain, 1st ed.; Lieblich, I., Ed.; Elsevier Biomedical Press: Amsterdam, NL, USA, 1982; pp. 95–123. [Google Scholar]
- Fernández-Teruel, A.; Escorihuela, R.M.; Gray, J.A.; Aguilar, R.; Gil, L.; Giménez-Llort, L.; Tobeña, A.; Bhomra, A.; Nicod, A.; Mott, R.; et al. A quantitative trait locus influencing anxiety in the laboratory rat. Genome Res. 2002, 12, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steimer, T.; Python, A.; Schulz, P.E.; Aubry, J.M. Plasma corticosterone, dexamethasone (DEX) suppression and DEX/CRH tests in a rat model of genetic vulnerability to depression. Psychoneuroendocrinology 2007, 32, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, O.; Lecca, D.; Piras, G.; Driscoll, P.; Corda, M.G. Dissociation between mesocortical dopamine release and fear-related behaviours in two psychogenetically selected lines of rats that differ in coping strategies to aversive conditions. Eur. J. Neurosci. 2003, 17, 2716–2726. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.P.; Poddighe, L.; Boi, M.; Sanna, F.; Piludu, M.A.; Corda, M.G.; Giorgi, O.; Quartu, M. Expression of BDNF and trkB in the hippocampus of a rat genetic model of vulnerability (Roman low-avoidance) and resistance (Roman high-avoidance) to stress-induced depression. Brain Behav. 2017, 7, e00861. [Google Scholar] [CrossRef]
- Serra, M.P.; Poddighe, L.; Boi, M.; Sanna, F.; Piludu, M.A.; Sanna, F.; Corda, M.G.; Giorgi, O.; Quartu, M. Effect of Acute Stress on the Expression of BDNF, trkB, and PSA-NCAM in the Hippocampus of the Roman Rats: A Genetic Model of Vulnerability/Resistance to Stress-Induced Depression. Int. J. Mol. Sci. 2018, 19, 3745. [Google Scholar] [CrossRef] [Green Version]
- Drevets, W.C.; Price, J.L.; Simpson, J.R., Jr.; Todd, R.D.; Reich, T.; Vannier, M.; Raichle, M.E. Subgenual prefrontal cortex abnormalities in mood disorders. Nature 1997, 24, 824–827. [Google Scholar] [CrossRef]
- Drevets, W.C. Functional anatomical abnormalities in limbic and prefrontal cortical structures in major depression. Prog. Brain Res. 2000, 126, 413–431. [Google Scholar] [CrossRef]
- Piras, G.; Giorgi, O.; Corda, M.G. Effects of antidepressants on the performance in the forced swim test of two psychogenetically selected lines of rats that differ in coping strategies to aversive conditions. Psychopharmacology 2010, 211, 403–414. [Google Scholar] [CrossRef]
- Piras, G.; Piludu, M.A.; Giorgi, O.; Corda, M.G. Effects of chronic antidepressant treatments in a putative genetic model of vulnerability (Roman low-avoidance rats) and resistance (Roman high-avoidance rats) to stress-induced depression. Psychopharmacology 2014, 231, 43–53. [Google Scholar] [CrossRef]
- Rosenthal, A.; Goeddel, D.V.; Nguyen, T.; Martin, E.; Burton, L.E.; Shih, A.; Laramee, G.R.; Wurm, F.; Mason, A.; Nikolics, K.; et al. Primary structure and biological activity of human brain-derived neurotrophic factor. Endocrinology 1991, 129, 1289–1294. [Google Scholar] [CrossRef]
- Klein, R.; Parada, L.F.; Coulier, F.; Barbacid, M. trkB, a novel tyrosine protein kinase receptor expressed during mouse neural development. EMBO J. 1989, 8, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 15, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Heninger, G.R.; Nestler, E.J. A molecular and cellular theory of depression. Arch. Gen. Psychiatry 1997, 54, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Charney, D.S.; Manji, H.K. Life Stress, Genes, and Depression: Multiple Pathways Lead to Increased Risk and New Opportunities for Intervention. Sci. STKE 2004, 2004, re5. [Google Scholar] [CrossRef] [Green Version]
- Gold, P.W.; Chrousos, G.P. Organization of the stress system and its dysregulation in melancholic and atypical depression: High vs. low CRH/NE states. Mol. Psychiatry 2002, 7, 254–275. [Google Scholar] [CrossRef] [Green Version]
- Grønli, J.; Bramham, C.; Murison, R.; Kanhema, T.; Fiske, E.; Bjorvatn, B.; Ursin, R.; Portas, C.M. Chronic mild stress inhibits BDNF protein expression and CREB activation in the dentate gyrus but not in the hippocampus proper. Pharmacol. Biochem. Behav. 2006, 85, 842–849. [Google Scholar] [CrossRef]
- Thompson Ray, M.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, trkB-TK+ and GAD67 mRNA expression in the hippocampus of individuals with schizophrenia and mood disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Chen, Z. The role of BDNF in depression on the basis of its location in the neural circuitry. Acta Pharmacol. Sin. 2011, 32, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Lammel, S.; Tye, K.M.; Warden, M.R. Progress in understanding mood disorders: Optogenetic dissection of neural circuits. Genes Brain Behav. 2014, 13, 38–51. [Google Scholar] [CrossRef]
- Nestler, E.J.; Carlezon, W.A., Jr. The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Koo, J.W.; Labonté, B.; Engmann, O.; Calipari, E.S.; Juarez, B.; Lorsch, Z.; Walsh, J.J.; Friedman, A.K.; Yorgason, J.T.; Han, M.H.; et al. Essential Role of Mesolimbic Brain-Derived Neurotrophic Factor in Chronic Social Stress-Induced Depressive Behaviors. Biol. Psychiatry 2016, 15, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2016, 24, 78–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 10, 864–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef]
- Liu, D.; Tang, Q.-Q.; Yin, C.; Song, Y.; Liu, Y.; Yang, J.-X.; Liu, H.; Zhang, Y.-M.; Wu, S.-Y.; Song, Y.; et al. BDNF-mediated projection-specific regulation of depressive-like and nociceptive behaviors in mesolimbic reward circuitry. Pain 2018, 159, 175. [Google Scholar] [CrossRef]
- Jongen-Rêlo, A.L.; Kaufmann, S.; Feldon, J. A differential involvement of the shell and core subterritories of the nucleus accumbens of rats in attentional processes. Neuroscience 2002, 111, 95–109. [Google Scholar] [CrossRef]
- Cassaday, H.J.; Horsley, R.R.; Norman, C. Electrolytic lesions to nucleus accumbens core and shell have dissociable effects on conditioning to discrete and contextual cues in aversive and appetitive procedures respectively. Behav. Brain Res. 2005, 28, 222–235. [Google Scholar] [CrossRef]
- Goto, Y.; Grace, A.A. Limbic and cortical information processing in the nucleus accumbens. Trends Neurosci. 2008, 31, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Sesack, S.R.; Grace, A.A. Cortico-Basal Ganglia reward network: Microcircuitry. Neuropsychopharmacology 2010, 35, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, O.; Piras, G.; Corda, M.G. The psychogenetically selected Roman high-and low-avoidance rat lines: A model to study the individual vulnerability to drug addiction. Neurosci. Biobehav. Rev. 2007, 31, 148–163. [Google Scholar] [CrossRef]
- Giorgi, O.; Corda, M.G.; Fernández-Teruel, A.A. Genetic model of impulsivity, vulnerability to drug abuse and schizophrenia-relevant symptoms with translational potential: The roman high- vs. low-avoidance rats. Front. Behav. Neurosci. 2019, 5, 145. [Google Scholar] [CrossRef] [PubMed]
- Heidbreder, C.A.; Groenewegen, H.J. The medial prefrontal cortex in the rat: Evidence for a dorso-ventral distinction based upon functional and anatomical characteristics. Neurosci. Biobehav. Rev. 2003, 27, 555–579. [Google Scholar] [CrossRef] [PubMed]
- Berendse, H.W.; Groenewegen, H.J. Lohman, A.H.M. Compartmental distribution of ventral striatal neurons projecting to the ventral mesencephalon in the rat. J. Neurosci. 1992, 12, 2079–2103. [Google Scholar] [CrossRef] [Green Version]
- Holly, E.N.; Miczek, K.A. Ventral tegmental area dopamine revisited: Effects of acute and repeated stress. Psychopharmacology 2016, 233, 163–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, D.L.; Wessendorf, M.W.; Williams, J.T. A subset of ventral tegmental area neurons is inhibited by dopamine, 5-hydroxytryptamine and opioids. Neuroscience 1997, 77, 155–166. [Google Scholar] [CrossRef]
- Grace, A.A.; Onn, S.P. Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J. Neurosci. 1989, 9, 3463–3481. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.W.; North, R.A. Opioids excite dopamine neurons by hyperpolarization of local interneurons. J. Neurosci. 1992, 12, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Lammel, S.; Ion, D.I.; Roeper, J.; Malenka, R.C. Projection-specific modulation of dopamine neuron synapses by aversive and rewarding stimuli. Neuron 2011, 70, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Teruel, A.; Driscoll, P.; Gil, L.; Aguilar, R.; Tobeña, A.; Escorihuela, R.M. Enduring effects of environmental enrichment on novelty seeking, saccharin and ethanol intake in two rat lines (RHA/Verh and RLA/Verh) differing in incentive-seeking behavior. Biochem. Behav. 2002, 73, 225–231. [Google Scholar] [CrossRef]
- Sanna, F.; Piludu, M.A.; Corda, M.G.; Melis, M.R.; Giorgi, O.; Argiolas, A. Involvement of dopamine in the differences in sexual behaviour between Roman high- and low-avoidance rats: An intracerebral microdialysis study. Behav. Brain Res. 2014, 281, 177–186. [Google Scholar] [CrossRef]
- Corda, M.G.; Piras, G.; Piludu, M.A.; Giorgi, O. Differential effects of voluntary ethanol consumption on dopamine output in the nucleus accumbens shell of Roman high- and low-avoidance rats: A behavioral and brain microdialysis study. World J. Neurosci. 2014, 4, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Lecca, D.; Piras, G.; Driscoll, P.; Giorgi, O.; Corda, M.G. A differential activation of dopamine output in the shell and core of the nucleus accumbens is associated with the motor responses to addictive drugs: A brain dialysis study in Roman high- and low-avoidance rats. Neuropharmacology 2004, 46, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998; p. 237. ISBN 10: 0125476191. [Google Scholar]
- Palkovits, M. Punch sampling biopsy technique. Methods Enzymol. 1983, 103, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurements with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Poddighe, L.; Serra, M.P.; Boi, M.; Bratzu, J.; Sanna, F.; Corda, M.G.; Giorgi, O.; Melis, M.R.; Argiolas, A.; et al. c-Fos, ΔFosB, BDNF, trkB and Arc expression in the limbic system of male Roman High and Low Avoidance rats that show differences in sexual behaviour: Effect of sexual activity. Neuroscience 2018, 396, 1–23. [Google Scholar] [CrossRef]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The Role of BDNF on Neural Plasticity in Depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Measures | RHA | RLA |
|---|---|---|
| Immobility latency (s) | 291 ± 45 | 124 ± 21 * |
| Immobility time (s) | 256 ± 19 | 538 ± 22 *** |
| Swimming (s) | 299 ± 28 | 211 ± 36 * |
| Climbing (s) | 286 ± 31 | 132 ± 12 *** |
| Diving (s) | 59 ± 8 | 19 ± 2 *** |
| Fecal boli (n) | 3 ± 2 | 7 ± 1 * |
| Line | FS | Line × FS | ||||||
|---|---|---|---|---|---|---|---|---|
| Brain Area | Marker | F Value | p Value | F Value | p Value | F Value | p Value | d.f. |
| VTA | BDNF | 6.052 | 0.021 | 4.630 | 0.041 | 5.146 | 0.0318 | 1, 26 |
| trkB | 2.764 | ns | 7.563 | 0.0103 | 5.794 | 0.0229 | 1, 28 | |
| Acb s | BDNF | 0.055 | ns | 0.011 | ns | 2.684 | ns | 1, 28 |
| trkB | 5.517 | 0.0261 | 0.1088 | ns | 0.0647 | ns | 1, 28 | |
| Acb c | BDNF | 2.723 | ns | 2.984 | ns | 9.224 | 0.0051 | 1, 28 |
| trkB | 1.717 | ns | 1.550 | ns | 14.49 | 0.0007 | 1, 28 | |
| ACg cortex | BDNF | 3.266 | 0.0082 | 4.267 | 0.048 | 7.956 | 0.009 | 1, 28 |
| trkB | 0.035 | ns | 0.009 | ns | 18,98 | 0.0002 | 1, 28 | |
| IL/PL cortex | BDNF | 2.829 | ns | 0.548 | ns | 32.45 | <0.0001 | 1, 28 |
| trkB | 3.641 | ns | 6.481 | 0.0167 | 8.165 | 0.008 | 1, 28 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, M.P.; Sanna, F.; Boi, M.; Poddighe, L.; Secci, L.; Trucas, M.; Fernández-Teruel, A.; Corda, M.G.; Giorgi, O.; Quartu, M. Acute Stress Induces Different Changes on the Expression of BDNF and trkB in the Mesocorticolimbic System of Two Lines of Rats Differing in Their Response to Stressors. Int. J. Mol. Sci. 2022, 23, 14995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314995
Serra MP, Sanna F, Boi M, Poddighe L, Secci L, Trucas M, Fernández-Teruel A, Corda MG, Giorgi O, Quartu M. Acute Stress Induces Different Changes on the Expression of BDNF and trkB in the Mesocorticolimbic System of Two Lines of Rats Differing in Their Response to Stressors. International Journal of Molecular Sciences. 2022; 23(23):14995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314995
Chicago/Turabian StyleSerra, Maria Pina, Francesco Sanna, Marianna Boi, Laura Poddighe, Lorenzo Secci, Marcello Trucas, Alberto Fernández-Teruel, Maria Giuseppa Corda, Osvaldo Giorgi, and Marina Quartu. 2022. "Acute Stress Induces Different Changes on the Expression of BDNF and trkB in the Mesocorticolimbic System of Two Lines of Rats Differing in Their Response to Stressors" International Journal of Molecular Sciences 23, no. 23: 14995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314995