
Molecular Characteristics of Subgenomic RNAs and the Cap-Dependent Translational Advantage Relative to Corresponding Genomic RNAs of Tomato spotted wilt virus
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
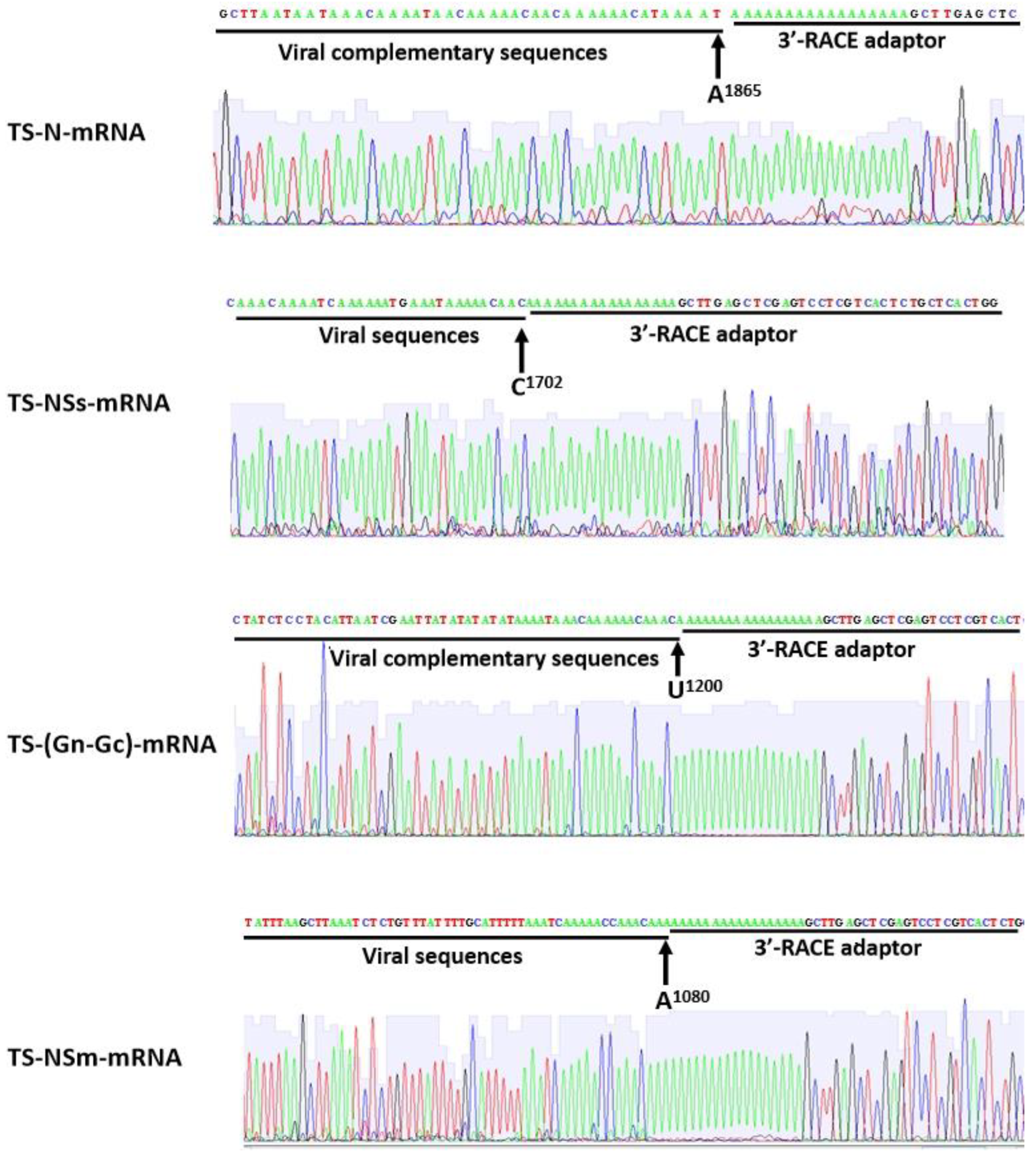
2.1. Mapping of 3′ Terminal Nucleotide in the TSWV sgRNAs
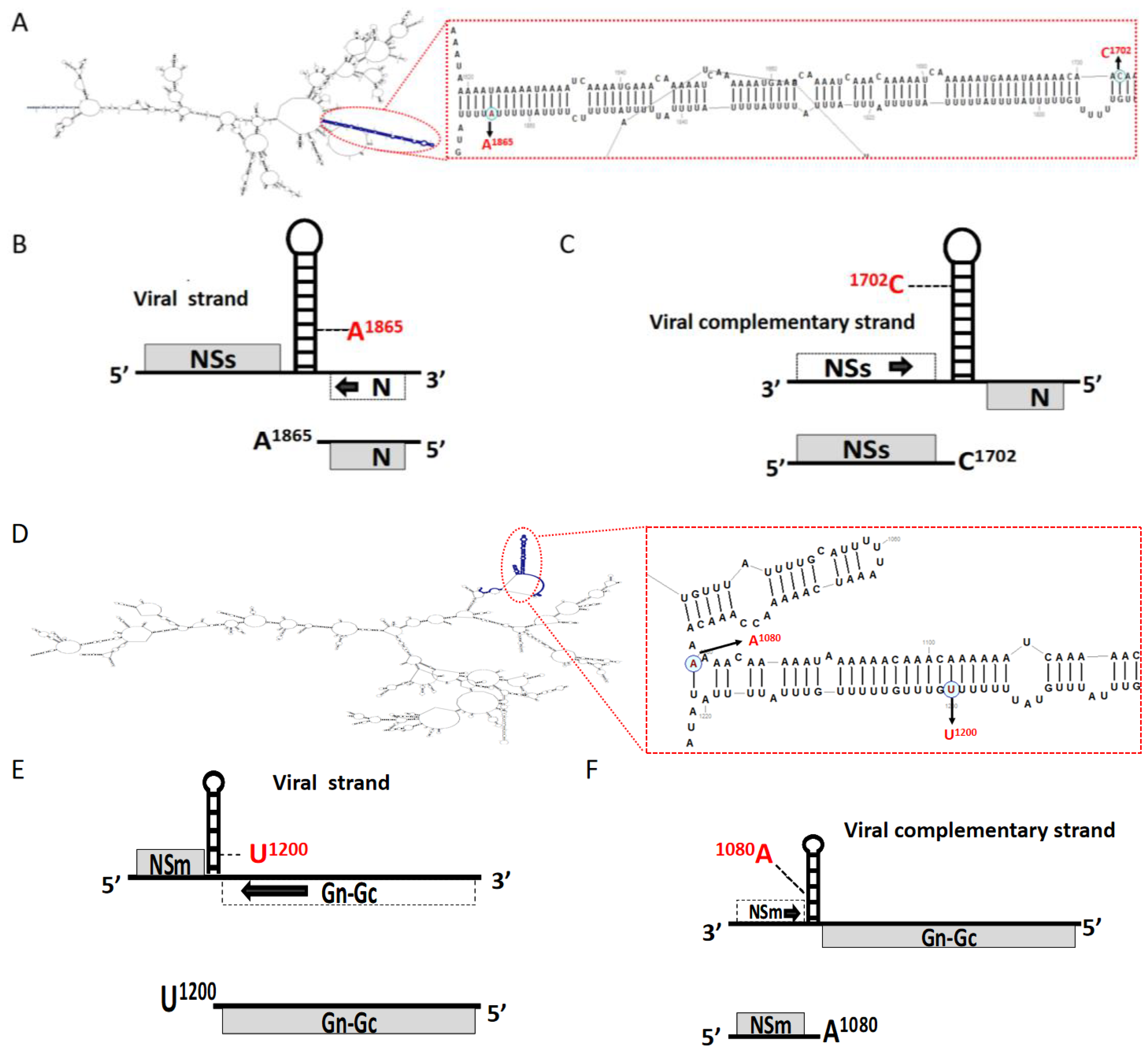
2.2. The 3′ End of TSWV sgRNAs Located at the Side of the A-Rich Region of IGR
2.3. Conservation Analysis of TSWV RNA M and S IGRs
2.4. Effect of 5′ and 3′ UTR of TSWV gRNA and sgRNA on Cap-Dependent or Cap-Independent Translation
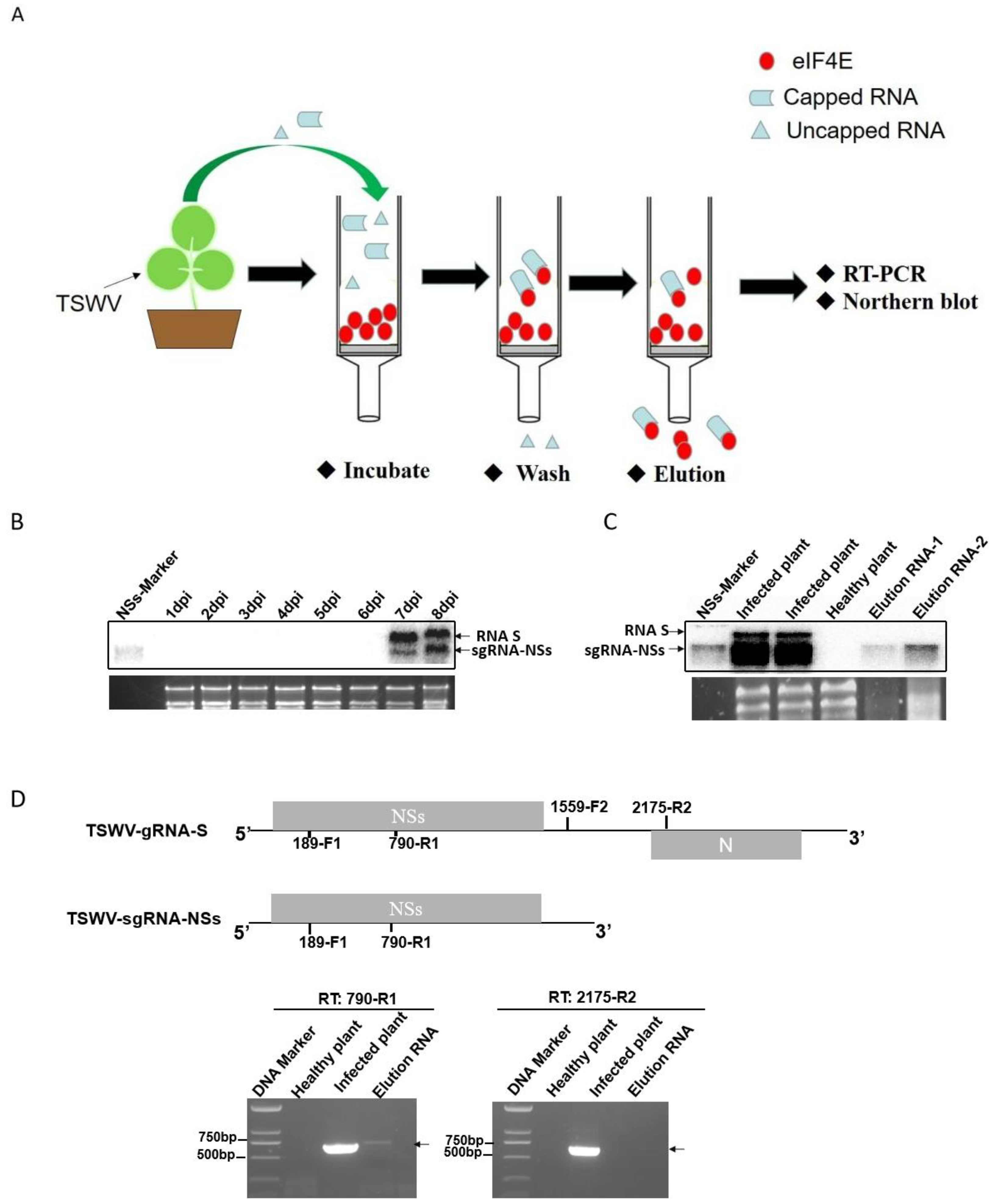
2.5. The TSWV sgRNA-NSs, but Not RNA S, Is 5′ Capped In Vivo
3. Discussion
3.1. Molecular Characteristics of TSWV sgRNAs Superior to gRNAs in Viral Proteins Expression
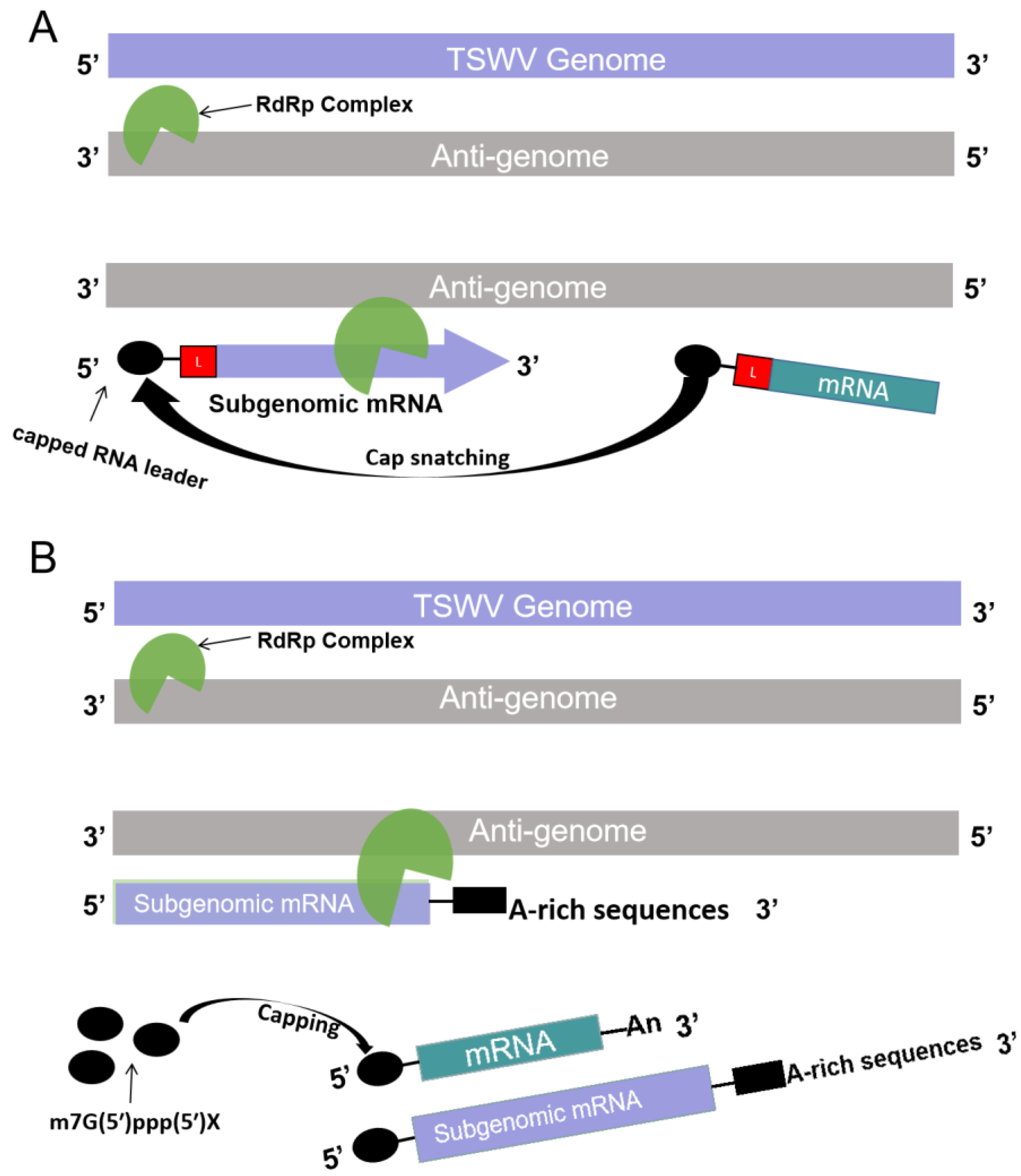
3.2. Possible Mechanisms of sgRNA Synthesis of Ambisense RNA Viruses
4. Materials and Methods
4.1. Rapid Amplification of cDNA Ends (3′RACE)
4.2. Plasmid Construction
4.3. RNA Preparation
4.4. In Vitro Luciferase Translation Assays
4.5. In Vitro Translation of TSWV gRNAs and sgRNAs
4.6. eIF4E Expression and Purification
4.7. RT-PCR Detection of TSWV gRNAs and sgRNAs
4.8. Northern Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, F.; Kasprzak, W.K.; Szarko, C.; Shapiro, B.A.; Simon, A.E. The 3′ untranslated region of pea enation mosaic virus contains two T-shaped, ribosome-binding, cap-independent translation enhancers. J. Virol. 2014, 88, 11696–11712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Embarc-Buh, A.; Francisco-Velilla, R.; Martinez-Salas, E. RNA-binding proteins at the host-pathogen interface targeting viral regulatory elements. Viruses 2021, 13, 952. [Google Scholar] [CrossRef] [PubMed]
- Di, H.; McIntyre, A.A.; Brinton, M.A. New insights about the regulation of Nidovirus subgenomic mRNA synthesis. Virology 2018, 517, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Wongsurawat, T.; Jenjaroenpun, P.; Taylor, M.K.; Lee, J.; Tolardo, A.L.; Parvathareddy, J.; Kandel, S.; Wadley, T.D.; Kaewnapan, B.; Athipanyasilp, N.; et al. Rapid sequencing of multiple RNA viruses in their native form. Front. Microbiol. 2019, 10, 260. [Google Scholar] [CrossRef]
- Kanodia, P.; Miller, W.A. Effects of the noncoding subgenomic RNA of red clover necrotic mosaic virus in Virus Infection. J. Virol. 2022, 96, e01815-21. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; Zhang, Q.; Zhang, B.; Xia, H.; Yuan, Z. Homologous RNA secondary structure duplications in 3′ untranslated region influence subgenomic RNA production and replication of dengue virus. Virology 2018, 524, 114–126. [Google Scholar] [CrossRef]
- Ilyas, M.; Du, Z.; Simon, A.E. Opium poppy mosaic virus has an Xrn-resistant, translated subgenomic RNA and a BTE 3′ CITE. J. Virol. 2021, 95, e02109-20. [Google Scholar] [CrossRef]
- Dinesh-Kumar, S.P.; Brault, V.; Allen, W. Precise mapping and in vitro translation of a trifunctional subgenomic RNA of barley yellow dwarf virus. Virology 1992, 187, 711–722. [Google Scholar] [CrossRef]
- Guo, S.; Wong, S.M. Disruption of a stem-loop structure located upstream of pseudoknot domain in tobacco mosaic virus enhanced its infectivity and viral RNA accumulation. Virology 2018, 519, 170–179. [Google Scholar] [CrossRef]
- Rasekhian, M.; Roohvand, F.; Habtemariam, S.; Marzbany, M.; Kazemimanesh, M. The role of 3′UTR of RNA viruses on mRNA stability and translation enhancement. Mini Rev. Med. Chem. 2021, 21, 2389–2398. [Google Scholar] [CrossRef]
- Sõmera, M.; Massart, S.; Tamisier, L.; Sooväli, P.; Sathees, K.; Kvarnheden, A. A survey using high-throughput sequencing suggests that the diversity of cereal and barley yellow dwarf viruses is underestimated. Front. Microbiol. 2021, 12, 673218. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. Translational control of cellular and viral mRNAs. Plant Mol. Biol. 1996, 32, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Allen, E.M.; Miller, W.A. Base-pairing between untranslated regions facilitates translation of uncapped, nonpolyadenylated viral RNA. Mol. Cell 2001, 7, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Merrick, W.C. Cap-dependent and cap-independent translation in eukaryotic systems. Gene 2004, 332, 1–11. [Google Scholar] [CrossRef]
- Kneller, E.L.P.; Rakotondrafara, A.M.; Miller, W.A. Cap-independent translation of plant viral RNAs. Virus Res. 2006, 119, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Weingarten-Gabbay, S.; Elias-Kirma, S.; Nir, R.; Gritsenko, A.A.; Stern-Ginossar, N.; Yakhini, Z.; Weinberger, A.; Segal, E. Systematic discovery of cap-independent translation sequences in human and viral genomes. Science 2016, 351, aad4939. [Google Scholar] [CrossRef]
- Geng, G.; Wang, D.; Liu, Z.; Wang, Y.; Zhu, M.; Cao, X.; Yu, C.; Yuan, X. Translation of plant RNA viruses. Viruses 2021, 13, 2499. [Google Scholar] [CrossRef]
- Fauquet, C.M.; Mayo, M.A.; Maniloff, J.; Desselberger, U.; Ball, L.A. 8th report of the international committee on taxonomy of viruses. In Virus Taxonomy; Elsevier Academic Press: San Diego, CA, USA, 2005. [Google Scholar]
- Huang, H.; Huang, S.; Li, J.; Wang, H.; Zhao, Y.; Feng, M.; Dai, J.; Wang, T.; Zhu, M.; Tao, X. Stepwise artificial evolution of an Sw-5b immune receptor extends its resistance spectrum against resistance-breaking isolates of tomato spotted wilt virus. Plant Biotechnol. J. 2021, 19, 2164–2176. [Google Scholar] [CrossRef]
- Qi, S.; Shen, Y.; Wang, X.; Zhang, S.; Li, Y.; Islam, M.; Wang, J.; Zhao, P.; Zhan, X.; Zhang, F.; et al. A new NLR gene for resistance to tomato spotted wilt virus in tomato (Solanum lycopersicum). Theor. Appl. Genet. 2022, 135, 1493–1509. [Google Scholar] [CrossRef]
- Kormelink, R.; Storms, M.; van Lent, J.; Peters, D.; Goldbach, R. Expression and subcellular location of the NSm protein of tomato spotted wilt virus (TSWV), a putative viral movement protein. Virology 1994, 200, 56–65. [Google Scholar] [CrossRef]
- Kormelink, R.; Van Poelwijk, F.; Peters, D.; Goldbach, R. Non-viral heterogeneous sequences at the 5′ ends of tomato spotted wilt virus mRNAs. J. Gen. Virol. 1992, 73, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; Kiima, E.W.; Haana, P.D. The nonstructural protein(NSs) encoded bythe ambisen~S RNA segment of tomao spoted wilt virus is associated with fibrous structures in infected plant cells. Virology 1991, 181, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.; Haenni, A.L. Expression strategies of ambisense viruses. Virus Res. 2003, 93, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Tsompana, M.; Abad, J.; Purugganan, M.; Moyer, J.W. The molecular population genetics of the tomato spotted wilt virus (TSWV) genome. Mol. Ecol. 2005, 14, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duijsings, D.; Kormelink, R.; Goldbach, R. Alfalfa mosaic virus RNAs serve as cap donors for tomato spotted wilt virus transcription during coinfection of Nicotiana benthamiana. J. Virol. 1999, 73, 5172–5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Zhou, Y.; Moffett, P. Alterations in cellular RNA de capping dynamics affect tomato spotted wilt virus cap snatching and infection in Arabidopsis. New Phytol. 2019, 224, 789–803. [Google Scholar] [CrossRef]
- Duijsings, D.; Kormelink, R.; Goldbach, R. In vivo analysis of the TSWV cap-snatching mechanism: Single base complementarity and primer length requirements. EMBO J. 2001, 20, 2545–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Knippenberg, I.; Goldbach, R.; Kormelink, R. In vitro transcription of tomato spotted wilt virus is independent of translation. J. Gen. Virol. 2004, 85, 1335–1338. [Google Scholar] [CrossRef]
- van Knippenberg, I.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus S-segment mRNAs have overlapping 3′-ends containing a predicted stem-loop structure and conserved sequence motif. Virus Res. 2005, 110, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Heinze, C.; Willingmann, P.; Schwach, F.; Adam, G. An unusual large intergenic region in the S-RNA of a bulgarian tomato spotted wilt virus isolate. Arch. Virol. 2003, 148, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Geerts-Dimitriadou, C.; Lu, Y.Y.; Geertsema, C.; Goldbach, R.; Kormelink, R. Analysis of the tomato spotted wilt virus ambisense S RNA-encoded hairpin structure in translation. PLoS ONE 2012, 7, e31013. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Cheng, R.; Chen, M.; Guo, R.; Li, L.; Feng, Z.; Wu, J.; Xie, L.; Hong, J.; Zhang, Z.; et al. Rescue of tomato spotted wilt virus entirely from complementary DNA clones. Proc. Natl. Acad. Sci. USA 2020, 117, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Koundal, V.; Williams, S.; Pappu, H. Differential expression of tomato spotted wilt virus-derived viral small RNAs in infected commercial and experimental host plants. PLoS ONE 2013, 8, e76276. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Rakotondrafara, A.M.; Miller, W.A. Trans regulation of cap-independent translation by a viral subgenomic RNA. J. Virol. 2006, 80, 10045–10054. [Google Scholar] [CrossRef] [Green Version]
- Sztuba-Solińska, J.; Stollar, V.; Bujarski, J.J. Subgenomic messenger RNAs: Mastering regulation of (+)-strand RNA virus life cycle. Virology 2011, 412, 245–255. [Google Scholar] [CrossRef] [Green Version]
- van Knippenberg, I.; Goldbach, R.; Kormelink, R. Purified tomato spotted wilt virus particles support both genome replication and transcription in vitro. Virology 2002, 303, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Gallie, D.R. Cap-independent translation conferred by the 5′ leader of tobacco etch virus is eukaryotic initiation factor 4G dependent. J. Virol. 2001, 75, 12141–12152. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.A.; Koev, G. Synthesis of subgenomic RNAs by positive-strand RNA viruses. Virology 2000, 273, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Mazur, M.; Gulickx, N.; Hong, H.; Overmars, H.; Tao, X.; Kormelink, R. Bunyaviral N proteins localize at RNA processing bodies and stress granules: The enigma of cytoplasmic sources of capped RNA for cap Snatching. Viruses 2022, 14, 1679. [Google Scholar] [CrossRef]
- Koev, G.; Miller, W.A. A positive-strand RNA virus with three very different subgenomic RNA promoters. J. Virol. 2000, 74, 5988–5996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galiakparov, N.; Goszczynski, D.E.; Che, X.; Batuman, O.; Bar-Joseph, M.; Mawassi, M. Two classes of subgenomic RNA of grapevine virus A produced by internal controller elements. Virology 2003, 312, 434–448. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S.; Tennant, P. Replication and expression strategies of viruses. Viruses 2018, 55–82. [Google Scholar] [CrossRef]
- Olschewski, S.; Cusack, S.; Rosenthal, M. The Cap-Snatching Mechanism of Bunyaviruses. Trends Microbiol. 2020, 28, 293–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerts-Dimitriadou, C.; Goldbach, R.; Kormelink, R. Preferential use of RNA leader sequences during influenza A transcription initiation in vivo. Virology 2011, 409, 27–32. [Google Scholar] [CrossRef]
- van Knippenberg, I.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus transcriptase in vitro displays a preference for cap donor with multiple base complementarity to the viral template. Virology 2005, 335, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yu, C.; Wang, G.; Shi, K.; Li, F.; Yuan, X. Phylogenetic and recombination analysis of tobacco bushy top virus in China. Virol. J. 2015, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yang, C.; Deng, Y.; Cao, X.; Xu, W.; Han, Z.; Li, Q.; Yang, Y.; Yuan, X. Conserved RNA secondary structure in cherry virus A 5′-UTR associated with translation regulation. Virol. J. 2022, 19, 91. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Yu, C.; Zhang, Z.; Wang, D.; Yuan, X. Molecular Characteristics of Subgenomic RNAs and the Cap-Dependent Translational Advantage Relative to Corresponding Genomic RNAs of Tomato spotted wilt virus. Int. J. Mol. Sci. 2022, 23, 15074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315074
Yang C, Yu C, Zhang Z, Wang D, Yuan X. Molecular Characteristics of Subgenomic RNAs and the Cap-Dependent Translational Advantage Relative to Corresponding Genomic RNAs of Tomato spotted wilt virus. International Journal of Molecular Sciences. 2022; 23(23):15074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315074
Chicago/Turabian StyleYang, Chen, Chengming Yu, Zhenjia Zhang, Deya Wang, and Xuefeng Yuan. 2022. "Molecular Characteristics of Subgenomic RNAs and the Cap-Dependent Translational Advantage Relative to Corresponding Genomic RNAs of Tomato spotted wilt virus" International Journal of Molecular Sciences 23, no. 23: 15074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315074