

Focus on Marine Animal Safety and Marine Bioresources in Response to the SARS-CoV-2 Crisis

1
Fisheries College, Jimei University, Xiamen 361021, China
2
Key Laboratory of Healthy Mariculture for the East China Sea, Ministry of Agriculture and Rural, Xiamen 361021, China
3
Affairs Fujian Provincial Key Laboratory of Marine Fishery Resources and Eco-Environment, Xiamen 361021, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(23), 15136; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315136
Submission received: 18 October 2022
/
Revised: 18 November 2022
/
Accepted: 29 November 2022
/
Published: 1 December 2022
(This article belongs to the Special Issue Virus–Host Interaction and Cell Restriction Mechanisms)
Abstract
:SARS-CoV-2 as a zoonotic virus has significantly affected daily life and social behavior since its outbreak in late 2019. The concerns over its transmission through different media directly or indirectly have evoked great attention about the survival of SARS-CoV-2 virions in the environment and its potential infection of other animals. To evaluate the risk of infection by SARS-CoV-2 and to counteract the COVID-19 disease, extensive studies have been performed to understand SARS-CoV-2 biogenesis and its pathogenesis. This review mainly focuses on the molecular architecture of SARS-CoV-2, its potential for infecting marine animals, and the prospect of drug discovery using marine natural products to combat SARS-CoV-2. The main purposes of this review are to piece together progress in SARS-CoV-2 functional genomic studies and antiviral drug development, and to raise our awareness of marine animal safety on exposure to SARS-CoV-2.
1. Introduction
There are some common human coronaviruses which people normally get infected with, such as 229E, NL63, OC43, and HKU1 according to a report of the Center for Disease Control and Prevention (CDC) of the US (cdc.gov). Recently, some zoonotic coronaviruses have transmitted to, and attacked, humans regionally and globally, developing into human coronaviruses exemplified by the three viruses MERS-CoV (the β-coronavirus causing Middle East respiratory syndrome; MERS), SARS-CoV (the β-coronavirus causing severe acute respiratory syndrome; SARS), and SARS-CoV-2 (the novel coronavirus causing coronavirus disease; COVID-19). The burst of the infection by the latest coronavirus in humans was first reported in Wuhan, Hubei, China in December 2019, with the interim name of coronavirus 2019-nCoV proposed by the World Health Organization (WHO). Shortly after, a Public Health Emergency of International Concern (PHEIC) was declared by WHO on 30 January 2020 [1]. Later, the novel pathogenic coronavirus was officially renamed SARS-CoV-2 by the International Committee on Taxonomy of Virus (ICTV), and the disease was named COVID-19 (coronavirus disease 2019) in the system of International Classification of Disease (ICD) by WHO on 11 February 2020 [2]. The rapid spread of SARS-CoV-2, and the sharp increase in COVID-19 cases since its onset, drew international attention and constituted a global pandemic that was announced by WHO on 11 March 2020 [3].
To mitigate the transmission of SARS-CoV-2, strict restrictions and preventive measures have been implemented, causing tremendous impact on the economics, social behavior, and many aspects of daily life. Although diverse countermeasures were imposed, the COVID-19 pandemic has still caused more than 6 million deaths globally as of the preparation of this manuscript. In addition, the SARS-CoV-2 virus has been reported to have the potential to infect animals with close contact with humans, and some wild animals as well. Tremendous efforts and funds have been invested on clinical diagnostics, research, and medicine and vaccine development to understand and control COVID-19 disease. In this review, our focuses are centered on the genomic structure of SARS-CoV-2 and their functions, its potential targeting of marine animals, and marine bioactive compounds with therapeutic effect against COVID-19.
2. Genomic Structure of SARS-CoV-2 and Their Functions
Coronaviruses are enveloped viruses containing a positive-sense single-stranded RNA (+ssRNA) genome, belonging to the subfamily Coronavirinae in Coronaviridae family [4]. There are four genera of coronaviruses including α, β, γ, and δ coronavirus with some specific mutations and recombination. Currently, only the α- and β- coronaviruses have been reported to infect humans, such as 229E and NL63 of α-coronavirus and OC43 and HKU1 of β-coronavirus, while the γ- and δ- coronaviruses are known to infect only birds or birds and non-human mammals, respectively [5]. The SARS-CoV-2 virus is a member of β-coronaviruses that include at least four lineages of A, B, C, and D. The SARS-CoV-2 and SARS-CoV (or SARS-CoV-1) belong to lineage B, while OC43 and HKU1 belong to lineage A and MERS-CoV belongs to lineage C [6].
2.1. Full Genetic Makeup of SARS-CoV-2
After the initial clinical diagnosis of the novel coronavirus-caused disease, scientists are striving to dissect the genetic elements of SARS-CoV-2. On 3 February 2020, the full genome sequence of SARS-CoV-2 was published, which contains 29,903 nucleotides consisting of at least 14 ORFs (1ab, S, 3a, 3b, E, M, 6, 7a, 7b, 8, 9a, 9b, N, 10) with some overlaps encoding some structural, nonstructural, and accessory proteins [7]. Transcriptome data support the transcription of ORF1a, 1b, S, 3a, E, M, 6, 7a, 7b, 8, N [8]. Comparative genomic analysis further concluded that the protein-coding ORFs in the genome of SARS-CoV-2 include ORF1a, 1ab, S, 3a, 3c, E, M, 6, 7a, 7b, 8, N, and 9b [9]. Some disagreement and ambiguity exist for the numbers and names of the ORFs, such as ORF9b and 9c being also called ORF9a and 9b or ORF13 and ORF14, respectively [7,9,10,11]. Collectively, the primary genome structure of SARS-CoV-2 (Figure 1A) and its proposed protein-coding ORFs (Figure 1B) in the linear structure have been elucidated.
(+) Genomic RNA can function as mRNA directing translation into viral proteins, and can also be used as a template for the synthesis of (−) genomic RNA, which then guides new (+) genomic RNA synthesis by the action of RNA-dependent RNA polymerase (RdRp) during replication (Figure 1B). It is a common strategy for the (+) ssRNA virus to synthesize a set of subgenomic RNAs (sgRNAs) to direct the translation of its 3ʹ-proximal genes [12]. The (+) genomic RNA of SARS-CoV-2 can guide the synthesis of a polyprotein, namely pp1ab, by translating the ORF1ab located in the 5ʹ-end covering about two-thirds of the genome. The ORF1ab can also be referred to as ORF1a and ORF1b, in which ORF1a can be translated into pp1a by canonical translation, while a programmed −1 ribosomal frameshift (−1 PRF) at the end of ORF1a may happen leading to the synthesis of pp1ab by non-canonical translation (Figure 1B). The rest, approximately one-third, of the genome located at the 3′-end is transcribed into sgRNAs with varying lengths for their translation into the structural proteins S (spike), E (envelop), M (membrane), and N (nucleocapsid), and the at least six accessory proteins including 3a, 6, 7a, 7b, 8, and 9b [13]. Some ORFs, including ORF3b, 9c, and 10, obtained by computational analysis, may not encode a protein in the virus, although some studies on their ectopic expression have shown their biological functions [9,13]. One of the mechanisms proposed for the production of sgRNAs is a template switch, or jumping through the long-range base paring between the transcription regulatory sequence (ACGAAC; TRS) located in the 5′-leader (TRSL) and the one in the genome body (TRSB) before the ORFs during the synthesis of (−) gRNA. sgRNAs appear polycistronic, while it is assumed that only the first ORF after the junction part of the TRS sites is translated, and the sgRNA is named with the first ORF accordingly [14]. The transcription and translation of the gRNA and sgRNA are achieved through the action of replication and transcription complex (RTC) consisting of RdRp and other RNA processing enzymes in the double membrane vesicles (DMV) formed through transmembrane proteins nsp3/nsp4/nsp6-induced ER membrane remodeling [13,15,16].
2.2. ORF1ab Cleavage
ORF1ab encodes 16 non-structural proteins (nsps) with pp1a being cleaved into nsp1-11 and pp1ab into nsp1-10 and nsp12-16 [9] for the initial hijacking and colonization of host cells [17]. The cleavage of pp1a and pp1ab are mediated by two proteases, nsp3 (papain-like protease, PLpro) and nsp5 (chymotrypsin-like protease, 3CLpro, also called main protease, Mpro) [18]. The recognition sites for PLpro and Mpro are LXGG↓XX and X-(L/F/M)-Q↓(G/A/S)-X, respectively, and PLpro is responsible for the first three cuts to release nsp1/nsp2/nsp3, while Mpro cleaves the rest of the sites in pp1a/pp1ab after nsp4 [19] (Figure 2).
ORF1ab encodes components to facilitate SARS-CoV-2 immediate infection (Table 1). nsp1 binds to the small subunit of the host cell ribosome to block the translation of host transcripts while initiating the translation of the viral genome [20,21]. nsp2 is a zinc-finger protein that can interact with the human 4EHP-GIGYF2 complex to impact the functioning of post-transcriptional silencing machinery to suppress the host defense response [22]. nsp3, the longest nsp in the genome, can interconnect with the host proteins involved in the immune response, RNA metabolism, and some fundamental cellular functions, suggesting the attack of host cell functioning by nsp3 [23]. The nsp7-nsp8 tetramer (dimer of dimer) acts as the primase to initiate RNA replication and interact with nsp12 to form a replication/transcription complex [24]. nsp12 encodes RNA-dependent RNA polymerase responsible for viral replication and transcription with the participation of helicase encoded by nsp13 [25]. nsp14 is a bifunctional protein which possesses an N7-methyltransferase activity at its C-terminus and 3ʹ-5ʹ exonuclease (ExoN) activity at its N-terminus. The nsp14 interacts with nsp10 cofactor to form a complex to excise its proofreading during the RNA synthesis mediated by the RdRp, which lacks extension fidelity [26]. In addition, the nsp14-nsp10 complex catalyzes the formation of cap-0 structure (m7GpppA) of the newly synthesized viral RNA. Furthermore, the nsp16 methyltransferase and the activation cofactor nsp10 work as a complex to methylate the cap-0 structure at 2′-O position to form the cap-1 structure (m7GpppAm) [27]. Accumulating studies have shown that the nsps processed from ORF1ab can suppress the innate immunity of host cells to evade the host defense response, such as the suppression of type I interferon production [28].
2.3. Structural Proteins
The structural proteins of SARS-CoV-2 include S, E, M, and N proteins, and their structural domains are shown in Figure 3. The SARS-CoV-2 virion contains the gRNA bound with N protein packed within a capsid and the M, E, and S proteins incorporated in the outside membrane.
2.3.1. N Protein
The N protein consists of two structural domains and three intrinsically disordered regions (IDRs). The N-terminal domain (NTD) and the C-terminal domain (CTD) are flanked with IDRs at the N-terminus and C-terminus, respectively, with the central IDR in between [30]. The two structure domains are also called the RNA-binding domain (RBD) and dimerization domain (DD), and the three IDRs are named NTD, a central linker (LINK), and CTD, respectively (Figure 3A) [31]. The structural signatures of N protein suggest that the N protein can physically interact with the gRNA of SARS-CoV-2 for packaging through liquid-liquid phase separation (LLPS) to form a condensate, and this process is proposed to be driven by the recognition and binding of a specific element in the 5ʹ-end of ORF1ab as virion assembly excludes the sgRNAs [32]. Functional domain analysis suggests that the L/Q-rich subdomain of the central IDR or central linker of N protein is essential for its interaction with gRNA [30], while the other domains are also important for their interaction, as their presence can compensate for the missing of SR-rich or L/Q-rich region of the central IDR [30,33]. Further probing into the cis-acting element in the ORF1ab region for N protein-mediated gRNA packaging shows that the core element at 20080-21171 nt (nsp15-nsp16 region) is sufficient for viral packaging [34]. The packaging process proposed is that the N protein binds to the packaging signal in the gRNA to initiate the condensation and recruit more N protein along the gRNA. The recruited N proteins can interact through the dimerization domain to form large oligomers, eventually forming the mature and stable condensate of viral RNA-protein complex [30,31].
2.3.2. E Protein
The E protein is the smallest protein of SARS-CoV-2 structural proteins, with only one transmembrane domain (TMD) (Figure 3B) [35]. During viral infection, a small portion of the expressed E protein is incorporated into the viral envelope while the majority is in the ER-Golgi intermediate compartment (ERGIC) of the host cell, assembling into a pentamer imbedded in the membranes as ion channels (ICs). The C-terminal domain of the E protein faces the cytoplasmic side, and the N-terminal domain faces the ER-lumen [36]. The insertion of the E protein in the ER, Golgi, and ERGIC membranes can induce membrane curvature towards the cytoplasm, potentially facilitating the budding of the SARS-CoV-2 virions. In addition, the channel-like structure formation in the ER/Golgi membrane system can stimulate viral entry and trafficking in the host cell and affect the normal physiological activity of the host cell by altering membrane permeability [35].
2.3.3. M Protein
The M protein is the most abundant structural protein of SARS-CoV-2, consisting of three transmembrane domains (TMD) flanked with a short N-terminal domain (NTD) and a β-sheet sandwich domain (BD or CTD) at the C-terminus (Figure 3C). The M protein is also located in the ERGIC, where it can form a dimer with some structural plasticity, while it is unlikely to function as a channel for ion conduction [37]. The M protein is considered the major driver for virial assembly, which is supported by the observation that it can interact with the N protein and ribonucleoprotein complex, further recruiting S and E proteins, potentially through physical interactions to form virion particles [37,38].
2.3.4. S Protein
The S protein is the most extensively studied structural protein. It is a type I membrane protein with only a single-span transmembrane domain, while it is assembled as a trimer anchored in the surface of the SARS-CoV-2 virion [39]. Basically, the S protein contains a short signal peptide located at the N-terminus, followed by S1 and S2 subunits in the middle and at the C-terminus, respectively (Figure 3D). The S1 and S2 subunits can be further divided into several different domains [40]. The S1 subunit contains an N-terminal domain (NTD), a receptor-binding domain (RBD), and two C-terminal domains (CTD1 and CTD2). The RBD harbors the receptor-binding motif (RBM) interacting with the ACE2 receptor on the host cell [41]. The S2 subunit contains a fusion peptide (FP), transmembrane anchor (TM), cytoplasmic tail (CT), and other domains [40,42]. During SARS-CoV-2 virus maturation in the infected host cells, the connection between S1 and S2 subunits is cleaved by furin protease, and then the S1 and S2 subunits are linked non-covalently. The cleavage of S1 and S2 by furin definitely promotes viral infection, but other protease may also function for the cleavage with less efficiency as the knock-out of furin does not totally abolish SARS-CoV-2 infection and replication [43]. The S1 subunit, especially the RBD, exhibits high structural flexibility, shaping an active “up” or inactive “down” states by conformational change to modulate the accessibility to the ACE2 receptor [44]. Once the S1 domain of the active state interacts with the ACE2 receptor, another cleavage essential for the activation of SARS-CoV-2 in post fusion stage is the cleavage at the S2′ site, which can be achieved by the action of transmembrane serine protease 2 (TMPRSS2) on the cell surface or by cathepsin L in the endosomal compartment [45,46]. The cleavage at the S2ʹ site exposes the fusion peptide and the fusion machinery of the S2 subunit to the host cell membrane and further drives their fusion to form a fusion pore in the host cell membrane, facilitating the entry of viral gRNA into the host cell [44,47]. It has been shown that ACE2 undergoes SUMOylation (conjugation with small ubiquitin-like modifier 3) at the lysin (K) 187 residue that compromises its K48-ubiquitination, in turn, suppressing ACE2 protein degradation mediated by the TOLLIP (Toll interacting protein) cargo receptor in the autophagic degradation system. As such, inhibition of ACE2 SUMOylation to destabilize ACE2 can be an attractive approach to combat SARS-CoV-2 [48].
2.4. Accessory Proteins
Accessory proteins are considered to be dispensable for SARS-CoV-2 replication. The overall functions of the accessory proteins include dysregulation of host defense response and physiological activities and induction of apoptosis of the host cell, thus contributing to the pathogenicity of SARS-CoV-2 virus [49]. ORF3a and ORF7a demonstrate potent antagonist activity against autophagy [50], and ORF3a shows proapoptotic activity by activation of caspase-3 [51]. The contribution of ORF3a to the pathogenicity of SARS-CoV-2 is also supported by the observation that the deletion of ORF3a can reduce the cytokine storm in the host cell, reflected by the decreased ratio of IL6/IL10 [52]. ORF6 and ORF8 can suppress the host defense response by inhibiting the promoter activity of NF-κB transcription factor and its responsive gene of type I interferon IFN-α [53]. Similarly, ORF7b can induce the expression of type I interferon IFN-β, tumor necrosis factor TNF-α, and interleukin IL-6, and stimulate caspase-mediated apoptosis [54]. ORF9b is localized to mitochondria in the SARS-CoV-2 infected cell and can interact with mitochondrial importer receptor Tom70 of the translocase of mitochondrial outer membrane (TOM) complex, potentially interacting with its preprotein substrate binding site [55,56]. In addition, the overexpression of ORF9b and SARS-CoV-2 infection lead to reduced expression of Tom70, thus affecting the functions of Tom70 in mediating translocation of preprotein from cytosol to mitochondria and in recruiting antiviral proteins in mitochondria antiviral signaling (MAVS) [57].
2.5. Variants and Mutations
Although there is a proofreading mechanism in SARS-CoV-2 during its genome replication for the sake of genome conservation, mutations still occurred during the pandemic and led to the generation of lineages and variants. Especially, mutations in ExoN of nsp14 apparently can cause a higher mutation load [58], and deletion mutations can escape the correction by proofreading activity [59]. The World Health Organization has assigned simple labels for the SARS-CoV-2 variants using letters of the Greek alphabet (www.who.int). Since its initial burst in late 2019, the variants of SARS-CoV-2 considered variants of concern (VOC) include Alpha, Beta, Gamma, Delta, and Omicron, and each contains multiple PANGO lineages (cov-lineages.org). Omicron is the current variant circulating around the world, and includes BA1, BA2, BA3, BA4, and BA5 sub-lineages. The mutations can occur in the spike protein and other proteins, and the mutations in the spike protein may enhance its cleavage by furin, thus potentially facilitating viral cell entry, increasing viral transmissibility, and resulting in waning antibody efficacy [60]. The mutations not including the insertions and deletions in the spike protein of the Omicron variant with at least 75% prevalence include 20 substitutions, with K417N, S477N, N501Y, P681H being the mutations of interest [61]. The K417N, S477N, N501Y mutations in the receptor binding domain of the S protein can strengthen its interaction with the ACE2 receptor [62], and the P681H mutation resides in the spike S1/S2 cleavage site that potentially increases its cleavage [63]. However, these mutations are not unique to the Omicron variant.
3. Threat to Marine Animals
It has been reported that SARS-CoV-2 can be transmitted from humans to pet animals, zoo and farm animals, and wild animals [64]. According to the CDC report, companion animals such as cats, dogs, hamsters, and ferrets, as well as wildlife such as mink, white-tailed deer, and mule deer, and animals in zoos and sanctuaries, such as lions and tigers, can catch SARS-CoV-2 virus, with more than 400 confirmed cases in total in the USA to date. The report from the World Organization for Animal Health (WOAH) released on 31 July 2022 indicates that a total of 679 outbreaks have been reported worldwide [65]. The transmission potential has raised the concern that SARS-CoV-2 might be transmitted to more wild animals with possible susceptibility, and thus precautionary measures should be in practice for outdoor activities such as wildlife research [66]. Genomic sequencing analysis of SARS-CoV-2 isolated from mink and white-tailed deer suggests that no significant mutations or accelerated mutation rate has occurred during the spillover from human to animal for viral adaption to the animal host, implying the nature of a general mammalian virus of SARS-CoV-2 [67].
Under the pandemic circumstances, human body fluids, feces, and contaminants, through close contact containing SARS-CoV-2 particles, may enter the sewage system, and identification of the viral particles provides an efficient early surveillance of the emergence of new SARS-CoV-2 variants and an earlier indicator of its rising incidence in the community [68,69]. The contaminated wastewater may eventually flow into the sea without sufficient treatment, or spill over to the saltwater environment. With an attempt to remove SARS-CoV-2 viral particles from wastewater, an algae-based microrobot was designed using Chlamydomonas reinhardtii as the self-driven matrix and fusing ACE2 protein to the algal cell surface, which can effectively adsorb the spike protein and SARS-CoV-2 pseudovirus in the tested aqueous media [70].
Shellfish are used as sentinels to monitor the potential contamination of SARS-CoV-2 in marine coastal areas; no apparent contamination of SARS-Co-2 was detected in the French shores in the summer of 2020 [71]. However, in another similar study, the SARS-CoV-2 RNA was detected in estuarine sediments and in bivalve molluscan species from a natural clam bank in Spain [72]. By using PMAxxx DNA modifier dye, which can only permeate dead cells and covalently binds to nucleic acid after photoactivation, to differentiate free RNA or damaged virion from intact encapsidated viral RNA, the PMAxxx-triton viability RT-PCR assay demonstrated that no infectious viral particles were detected even though the SARS-CoV-2 RNA was detected [72]. The absence or under the detection limit of the viable SARS-CoV-2 virion might be partially explained by the fact the marine high pH and salinity can affect the infectivity and integrity of SARS-CoV-2 virus [73].
Meanwhile, the possibility of marine animals harboring or getting infected with SARS-CoV-2 may exist. A novel nidovirus, Pacific salmon nidovirus (PsNV), which is distantly related to SARS-CoV-2, has been detected in wild keystone salmon of the northeastern Pacific with high abundance in the gill tissue, potentially causing declines in their population [74]. Additionally, air-breathing marine animals such otters and dolphins may exposure to SARS-CoV-2 or act as intermediate hosts of SARS-CoV-2, posing the risk of zoonotic COVID-19 disease [75]. Concerns over the spread of SARS-CoV-2 to marine wildlife in Antarctica have also been discussed, exploring the transmission potentials by human activities, human-to-animal-to-animal route, wastewater, and marine animal migration [76]. Generally, the binding affinity of SARS-CoV-2 spike protein to the ACE2 receptor of animals is a factor in the success of viral infection and the determination of susceptibility of the host to SARS-CoV-2, while some other factors play a role as well. The cold temperature of the Antarctic ocean may favor the viability of SARS-CoV-2 for an extended period of time, and the predicted high binding affinity of ACE2 of Antarctic minke whales and killer whales, and the medium affinity of the ACE2 of sperm whales, suggest the infection potentials of these Antarctic mammals by SARS-CoV-2 [76,77]. In addition, other cetacean species including the bottlenose dolphin, Pacific white-sided dolphin, baiji, beluga whale, long-finned pilot whale, and vaquita, are predicted to be highly susceptible to SARS-CoV-2 infection after computational analysis of their ACE2 binding affinity to the viral spike protein: baiji and vaquita are on the brink of extinction [77,78]. Additionally, sea otters of Fissipedia and Hawaiian monk seal of Pinnipedia are endangered species with high susceptibility to SARS-CoV-2 [78]. In a similar study by analysis of the 25 amino acids of ACE2 interacting with SARS-CoV-2 spike protein, the marine mammals living in Italian coastal waters are supposed to have medium to high susceptibility to SARS-CoV-2 infection, and the immunohistochemistry for ACE2 protein distribution in lung tissues of cetacean species suggests that ACE2 is expressed in alveolar and bronchial epithelium, supporting viral infection potentials [79]. On the other hand, fish such as zebrafish, Nile tilapia, large yellow croaker, and rainbow trout all are predicted to have very low susceptibility to SARS-CoV-2 [77], and no fish cell lines or HeLa cells transfected with ACE2 orthologs of some fish species have shown any infection possibility by SARS-CoV-2 [80]. A comparison of the computed three-dimensional structures of hACE2 and ACE2 proteins of Nibea albiflora (yellow drum) and Larimichthys crocea (large yellow croaker) marine fishes shows the differential spatial distribution of the spike-interacting residues (Figure 4), which may account for the low affinity of marine fish ACE2 proteins to the spike protein of SARS-CoV-2.
4. Antiviral Activity against SARS-CoV-2 in Marine Resources
Since the outbreak of COVID-19, tremendous endeavors have been undertaken to find effective antiviral strategies against SARS-CoV-2 infection. The development and distribution of COVID-19 (SARS-CoV-2) vaccines have greatly helped people alleviate the risk of getting seriously ill if contracting SARS-CoV-2. To date, there are four COVID-19 vaccines that have been authorized by the Food and Drug Administration (FDA) in the US, including Pfizer-BioNTech, Moderna, Johnson & Johnson’s Janssen, and the most recently approved Novavax, with notable effectiveness [81]. Some new vaccines are also under development, such as the plant-based virus-like particle (CoVLP) vaccine by adjuvanting the purified modified SARS-CoV-2 spike protein expressed in Nicotiana benthamiana with AS03 adjuvant, which has demonstrated notable cross-reactivity against different SARS-CoV-2 variants [82]. However, with the everchanging genomic information of SARS-CoV-2 and the waning of immunity, breakthrough infections of SARS-CoV-2 have occurred in many cases in vaccinated people [83]. In addition, some antiviral drugs, monoclonal antibodies, and immune-modulators have been developed as COVID-19 therapeutics for emergency use. Veklury (remdesivir) and Olumiant (baricitinib) are so far the two FDA-approved drugs for treatment of COVID-19 in the US. Remdesivir is a prodrug, an adenosine nucleotide analogue, which undergoes intracellular phosphorylation to be accommodated by the RdRp of SARS-CoV-2 to inhibit the viral RNA synthesis process [84]. Baricitinib is a Janus kinase (JAK) inhibitor with anti-inflammatory property, demonstrating activities in reducing receptor-mediated viral endocytosis and suppressing cytokine storm, and has been repurposed as a medication to treat hospitalized adults with moderate to severe COVID-19 [85,86]. In the meantime, scientists have devoted tremendous effort to identifying new compounds targeting SARS-CoV-2 and the post-infection immune system to combat viral infection and the disease through in silico, in vitro, and in vivo approaches [87,88]. Natural products with antimicrobial and antiviral properties are of great interest to scientists to explore their efficiency in combating COVID-19 [89]. It has been known for many decades that marine products serve as a seemingly limitless bio-resource for combating pathogenic microbes and cancers [90]. Studies on marine-derived antiviral compounds have discovered a wide collection of bioactive molecules, with different targets of components shaping SARS-CoV-2 successful infection and proliferation (Table 2), aiming to find alternative pharmaceuticals with enhanced specificity to SARS-CoV-2 and reduced side effects on the human body. Here some extensively studied compounds are further discussed with more details; for brevity, not all the functions of the compounds are mentioned.
4.1. Targeting Viral Recognition and Interaction
4.1.1. Sulfated Polysaccharides
The entry of SARS-CoV-2 is initialized largely by the interaction between the viral spike protein and the host ACE2 receptor through attraction by electrostatic forces, in which the RBD of spike protein is dominantly positively charged while the ACE2 has a negatively charged surface [112]. Sulfated polysaccharides are highly diverse and abundant in the ocean, especially in macroalgae and some marine animals harboring polyanion of sulfate ions, which have been demonstrated to be effective against SARS-CoV-2 entry into host cells [91]. Three types of marine sulfated polysaccharides including sea cucumber sulfated polysaccharide (SCSP), fucoidan from brown algae, and ι-carrageenan from red algae have been shown to be capable of binding the spike protein of SARS-CoV-2, with SCSP exhibiting the strongest inhibitory effect [113]. A nasal spray containing ι- and κ-carrageenan, and an oral spray containing ι-carrageenan, showed anti-SARS-CoV-2 activity by preventing the attachment of viral particles to, and its entry into, TMPRSS2-expressing Vero E6 cells, while it did not apparently affect the host cell viability [114]. In addition, the λ-carrageenan from marine red algae was shown to be able to inhibit the entry of SARS-CoV-2 spike-pseudotyped virus and the infectious SARS-CoV-2 into Vero E6 cells, interfering with the spike protein-associated entry step [115].
4.1.2. Inorganic Polyphosphates
Inorganic polyphosphate (polyP) is another negatively charged polymer that has shown promising antiviral effect against SARS-CoV-2. polyPs with varying lengths are ubiquitously distributed in all living organisms, and involved in many physiological functions [116]. The enriched accumulation of polyP has been found in some marine bacteria and marine sponges [117,118]. A polyP with as short as 3 phosphate (Pi) units (polyP3) can significantly inhibit the binding of RBD of SARS-CoV-2 to ACE2, potentially through negatively charged Pi units of polyP interacting with the positively charged residues of Arg, Lys, and His of RBD through electrostatic interaction, especially since chemical modification of Arg residues with increased reactivity can enhance the inhibitory efficiency of polyP on RBD binding to ACE2 [92,93]. Besides its ability to bind RBD of SARS-CoV-2, long-chain polyP (polyP120) can also bind ACE2 through the interaction with its positively charged residues His378, His401, Arg393, and Arg514, leading to the proteasome-mediated degradation of ACE2 [107]. Furthermore, the polymer polyP40 showed activating effects on the expression of MUC1 and MUC5AC, which are membrane-tethered mucin and the gel-forming secreted mucin, respectively, after the polyP in a collagen hydrogel-mucin environment was attached to human alveolar basal epithelial A549 cells, preventing the invasion of SARS-CoV-2 in the epithelium of the airway and lung [109]. The secretion of mucin can benefit from the generation of ATP through the hydrolysis of polyP by ALP (alkaline phosphatase) and the phosphorylation of ADP by ADK (adenylate kinase) [109,119], and quercetin antioxidant and the synthetic anti-inflammatory dexamethasone in caged nanoparticles with polyP can enhance the effect of polyP on mucin production [119].
Additionally, polyP is found in dense granules of platelets, which release polyP after activation, subsequently triggering the initiation of blood clotting and the liberation of inflammatory mediators [120]. Later studies indicated the activity of polyP in triggering blood clotting varies depending on the length of the polymer [121], and a conflicting result was also reported in which the synthetic polyP can inhibit blood clotting by reducing the levels of Ca2+ and thromboxane affecting platelet aggregation [122]. It has been shown that severe symptomatic COVID-19 patients have hyperactivated platelets and a noted drop in the count of platelets, which induce local thrombus formation and a systemic coagulation defect resulting in serious and even fatal consequence [123,124]. These observations suggest that exogenous supplementation of polyP might be an option, as the high consumption of platelets in COVID-19 patients with thrombocytopenia, and a unit length of P50 was suggested, while further clinical studies are required [125].
4.1.3. Cyanobacteria Molecules
As well as their richness in sulfated polysaccharides, cyanobacteria possess other bioactive compounds such as pigments and amino acids. Four bioactive molecules (phycoerythrobilin, phycocyanobilin, phycourobilin, and folic acid) were identified, by molecular docking assays, in the microalgae Arthrospira with high binding affinity to the RBD region of SARS-CoV-2 spike protein and good bioavailability [94]. Additionally, through molecular docking, ADME (absorption, distribution, metabolism, excretion), and cell toxicity analysis for isolation of natural inhibitors against ACE2 from cyanobacteria bioactive compounds, mycosporine-glycine-valine and shinorine demonstrated low binding energy to ACE2, high solubility, and free of toxicity, providing strong potentials for antiviral drug development [108]. Another important group of antiviral molecules is the mannose-specific lectins, which are found in cyanobacteria and red and green algae [95]. SARS-CoV-2 can use the C-type lectin receptors such as DC-SIGN, L-SIGN and the sialic acid-binding immunoglobulin-like lectin 1 (SIGLEC1) as attachment receptors to facilitate its presentation to ACE2 receptor for viral trans infection [126]. The mannose-specific lectins, such as cyanovirin from cyanobacteria Nostoc ellipsosporum and griffithsin from red algae Griffithsia sp. can recognize the N-glycosylated spike protein of SARS-CoV-2 with high-mannose glycans, inhibiting SARS-CoV-2 infection [95].
4.2. Targeting Viral Replication
4.2.1. Polyphenols against Mpro
Phlorotannins are secondary metabolites in brown algae, and are oligomers of phloroglucinol. Molecular docking analysis predicted that eckol and trifucol with negative higher binding energy (lower binding energy) to 3CLpro could be good inhibitors of Mpro [96]. By computational modeling to virtually screen a Marine Natural Product (MNP) library for the interaction with Mpro, 17 compounds out of 180 selected molecular after initial pharmacophore filtering stood out as the best candidates showing interaction with Mpro, and phlorotannins represent a major group among these compounds [97]. Further docking studies suggested that 8,8′-bieckol, 6,6′-bieckol, and dieckol of phlorotannins identified in the brown algae Ecklonia cava were the most active inhibitors against Mpro. In addition, some flavonoids, including apigenin-7-O-neohesperidoside, luteolin-7-rutinoside, and resinoside were also listed as promising inhibitors of Mpro [97].
4.2.2. Alkaloids against PLpro and Mpro
Alkaloids are a large group of structurally diverse natural compounds, that contain at least one nitrogen atom, with great potential for drug development to treat diseases and mental disorders [127]. Marine sponges provide an abundant resource of bioactive alkaloid compounds [128]. Through molecular docking and molecular dynamics simulation studies to screen polycyclic guanidine alkaloid compounds in marine sponge Monanchora n. sp. targeting SARS-CoV-2 Mpro and other proteins, two compounds crambescidin 786 and crambescidin 826, showed high binding affinity to the enzyme pockets of Mpro with very low toxicity and high bioavailability, rendering them promising candidates as anti-SARS-CoV-2 drugs [98]. By employing a similar approach to probe marine compounds targeting Mpro of SARS-CoV-2 as potential inhibitors, an alkaloid compound fistularin 3 (also known as isofistularin-3; PubChem CID 159041) isolated from marine sponges of the Aplysinidae family was identified as the most potent candidate with strong bonding with the amino acid residues in the active site of Mpro [99]. Another virtual screen for anti-SARS-CoV-2 secondary metabolites from marine and terrestrial fungi identified three fumiquinazoline marine alkaloids scedapin, norquinadoline A, and quinadoline B that showed high affinity to the putative binding site of PLpro [100]. Another marine algae-derived alkaloid caulerpin was isolated after virtual screen of 10 bioactive natural compounds for the anti-SARS-CoV-2 potentials targeting Mpro, demonstrating the highest binding affinity to Mpro among the tested compounds [101]. The high negative free binding energy between caulerpin molecule and the modelled Mpro suggests caulerpin could be an effective antiviral drug against SARS-CoV-2.
4.2.3. Plitidepsin against Host Factor eEF1A
Plitidepsin, also known as dehydrodidemnin B, is a cyclic depsipeptide. It is a marine-derived compound extracted from Mediterranean tunicate Aplidium albicans, which has shown anticancer activity potentially targeting the eukaryotic elongation factor 1A2 (eEF1A2) [110]. Evaluation of the antiviral effect of plitidepsin as a repurposed drug against SARS-CoV-2 showed that plitidepsin can significantly inhibit SARS-CoV-2 replication post viral entry in Vero E6 cells and hACE2-293T cells with a substantially lower IC90 compared to remdesivir, while cytostatic impact on cell proliferation was observed [111]. The antiviral and antiproliferative actions of plitidepsin are mediated by its inhibitory activity on eEF1A by introduction of the mutated version eEF1A-A399V into hACE2-293T imparts the cells resistance to plitidepsin treatment, and the refractory effects can be rescued by transfection with the wild-type eEF1A [111]. The antiviral activity of plitidepsin was shown by its marked inhibition of SARS-CoV-2 viral genomic RNA replication and sub-genomic N RNA synthesis and the N protein expression after initial viral infection, with a stronger effect than remdesivir. In vivo data obtained from the infection of hACE2-sensitized or hACE2-transgenic mouse further supported antiviral effect of plitidepsin against SARS-CoV-2 [111]. Further investigation of the antiviral activity of plitidepsin on SARS-CoV-2 by transmission electron microscopy and immunohistochemistry was performed to examine its effect on viral replication [129]. It was evident that plitidepsin treatment induced the disappearance of DMVs structure for viral genome replication, and the absence of viral particle distribution in single-membrane vesicles, in the large vacuole, and on the extracellular side of the plasma membrane in Vero E6 cells at 24–D48 h post infection. In addition, the viral N protein and dsRNA were not detected by immunostaining in plitidepsin-treated Vero E6 cells at 48 h post infection [129]. A more recent study showed that plitidepsin can virtually bind to the main protease of SARS-CoV-2 and inhibit its activity [130].
5. Conclusions
This review aimed to provide an update on our understanding of SARS-CoV-2 genome composition and viral components for virion replication and assembly, and to present an overview of SARS-CoV-2 infection mechanism in host cells. The viral genome largely encodes structural proteins of spike, envelop, membrane, and nucleocapsid proteins, and some nonstructural and accessory proteins facilitating SARS-CoV-2 proliferation and infection. Given the complexity of SARS-CoV-2 genome composition, the translation and function of the molecular fragments are not fully understood yet and more in vivo studies are required to explore the unknowns. In addition, the potential of spreading SARS-CoV-2 to marine mammals has been reviewed largely based on the compatibility of host ACE2 receptor to the spike of SARS-CoV-2 while other entry routes may exist, at least in an auxiliary manner. Precaution and monitoring are necessary to avoid spillover to marine animals, although there has been no report about infection of marine mammals by SARS-CoV-2 yet. Antiviral drug development has driven enormous efforts in discovering natural products such as marine metabolites with anti-SARS-CoV-2 properties through in silico, in vitro, and in vivo approaches. One compound may target more than one protein component of SARS-CoV-2 to mitigate its infection. The most promising bioactive compounds, such as inorganic polyphosphates and plitidepsin, are to undergo comprehensive and thorough evaluation for their eventual application in clinical treatment.
Author Contributions
Conceptualization, F.H. and Y.Y.; formal analysis, F.H.; data curation, F.H., Y.Y. and J.L.; writing-original draft preparation, F.H., Y.Y. and J.L.; writing-review and editing, F.H.; visualization, J.L.; project administration, F.H.; funding acquisition, F.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 32072969 and 31872562; Natural Science Foundation of Fujian Province (2022J01325); Open Research Fund Program of Fujian Provincial Key Laboratory of Marine Fishery Resources and Eco-environment (Z822280); Open Research Fund Program of Key Laboratory of Healthy Mariculture for the East China Sea (2022ESHML10); National Marine Fisheries Industrial Technology System Post Scientist Project (CARS-47-G04).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest pertaining to the research report in this manuscript.
References
- Sohrabi, C.; Alsafi, Z.; O’Neill, N.; Khan, M.; Kerwan, A.; Al-Jabir, A.; Iosifidis, C.; Agha, R. World Health Organization declares global emergency: A review of the 2019 novel coronavirus (COVID-19). Int. J. Surg. 2020, 76, 71–76. [Google Scholar] [CrossRef]
- Dhama, K.; Khan, S.; Tiwari, R.; Sircar, S.; Bhat, S.; Malik, Y.S.; Singh, K.P.; Chaicumpa, W.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. Coronavirus Disease 2019-COVID-19. Clin. Microbiol. Rev. 2020, 33, e00028-20. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar]
- Paim, F.C.; Bowman, A.S.; Miller, L.; Feehan, B.J.; Marthaler, D.; Saif, L.J.; Vlasova, A.N. Epidemiology of deltacoronaviruses (δ-CoV) and gammacoronaviruses (γ-CoV) in wild birds in the United States. Viruses 2019, 11, 897. [Google Scholar] [CrossRef] [Green Version]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Lee, J.Y.; Yang, J.S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef]
- Jungreis, I.; Sealfon, R.; Kellis, M. SARS-CoV-2 gene content and COVID-19 mutation impact by comparing 44 Sarbecovirus genomes. Nat. Commun. 2021, 12, 2642. [Google Scholar] [CrossRef]
- Jungreis, I.; Nelson, C.W.; Ardern, Z.; Finkel, Y.; Krogan, N.J.; Sato, K.; Ziebuhr, J.; Stern-Ginossar, N.; Pavesi, A.; Firth, A.E.; et al. Conflicting and ambiguous names of overlapping ORFs in the SARS-CoV-2 genome: A homology-based resolution. Virology 2021, 558, 145–151. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koev, G.; Miller, W.A. A positive-strand RNA virus with three very different subgenomic RNA promoters. J. Virol. 2000, 74, 5988–5996. [Google Scholar] [CrossRef]
- Brant, A.C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z.M. SARS-CoV-2: From its discovery to genome structure, transcription, and replication. Cell Biosci. 2021, 11, 136. [Google Scholar] [CrossRef]
- Long, S. SARS-CoV-2 Subgenomic RNAs: Characterization, Utility, and Perspectives. Viruses 2021, 13, 1923. [Google Scholar] [CrossRef]
- Slanina, H.; Madhugiri, R.; Bylapudi, G.; Schultheiß, K.; Karl, N.; Gulyaeva, A.; Gorbalenya, A.E.; Linne, U.; Ziebuhr, J. Coronavirus replication–transcription complex: Vital and selective NMPylation of a conserved site in nsp9 by the NiRAN-RdRp subunit. Proc. Natl. Acad. Sci. USA 2021, 118, e2022310118. [Google Scholar] [CrossRef] [PubMed]
- Mariano, G.; Farthing, R.J.; Lale-Farjat, S.L.M.; Bergeron, J.R.C. Structural characterization of SARS-CoV-2: Where we are, and where we need to be. Front. Mol. Biosci. 2020, 7, 605236. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.A.; Woodside, M.T.; Dinman, J.D. Programmed-1 Ribosomal Frameshifting in coronaviruses: A therapeutic target. Virology 2021, 554, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Kok, K.H.; Zhu, Z.; Chu, H.; To, K.K.; Yuan, S.; Yuen, K.Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Osipiuk, J.; Azizi, S.-A.; Dvorkin, S.; Endres, M.; Jedrzejczak, R.; Jones, K.A.; Kang, S.; Kathayat, R.S.; Kim, Y.; Lisnyak, V.G.; et al. Structure of papain-like protease from SARS-CoV-2 and its complexes with non-covalent inhibitors. Nat. Commun. 2021, 12, 743. [Google Scholar] [CrossRef]
- Schubert, K.; Karousis, E.D.; Jomaa, A.; Scaiola, A.; Echeverria, B.; Gurzeler, L.A.; Leibundgut, M.; Thiel, V.; Mühlemann, O.; Ban, N. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat. Struct. Mol. Biol. 2020, 27, 959–966. [Google Scholar] [CrossRef]
- Mendez, A.S.; Ly, M.; González-Sánchez, A.M.; Hartenian, E.; Ingolia, N.T.; Cate, J.H.; Glaunsinger, B.A. The N-terminal domain of SARS-CoV-2 nsp1 plays key roles in suppression of cellular gene expression and preservation of viral gene expression. Cell Rep. 2021, 37, 109841. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Moch, C.; Graille, M.; Chapat, C. The SARS-CoV-2 protein NSP2 impairs the silencing capacity of the human 4EHP-GIGYF2 complex. iScience 2022, 25, 104646. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Feng, Z.; Zhang, X. Integrative multi-omics landscape of non-structural protein 3 of severe acute respiratory syndrome coronaviruses. Genom. Proteom. Bioinform. 2021, 19, 707–726. [Google Scholar] [CrossRef] [PubMed]
- Konkolova, E.; Klima, M.; Nencka, R.; Boura, E. Structural analysis of the putative SARS-CoV-2 primase complex. J. Struct. Biol. 2020, 211, 107548. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Malone, B.; Llewellyn, E.; Grasso, M.; Shelton, P.M.M.; Olinares, P.D.B.; Maruthi, K.; Eng, E.T.; Vatandaslar, H.; Chait, B.T.; et al. Structural basis for helicase-polymerase coupling in the SARS-CoV-2 replication-transcription complex. Cell 2020, 182, 1560–1573.e13. [Google Scholar] [CrossRef] [PubMed]
- Moeller, N.H.; Shi, K.; Demir, Ö.; Belica, C.; Banerjee, S.; Yin, L.; Durfee, C.; Amaro, R.E.; Aihara, H. Structure and dynamics of SARS-CoV-2 proofreading exoribonuclease ExoN. Proc. Natl. Acad. Sci. USA 2022, 119, e2106379119. [Google Scholar] [CrossRef]
- Benoni, R.; Krafcikova, P.; Baranowski, M.R.; Kowalska, J.; Boura, E.; Cahová, H. Substrate specificity of SARS-CoV-2 nsp10-nsp16 methyltransferase. Viruses 2021, 13, 1722. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Gao, M.; Gao, X.; Zhu, B.; Huang, J.; Luo, K.; Zhang, Y.; Sun, J.; Deng, M.; Lou, Z. SARS-CoV-2 non-structural protein 13 (nsp13) hijacks host deubiquitinase USP13 and counteracts host antiviral immune response. Signal Transduct. Target. Ther. 2021, 6, 119. [Google Scholar] [CrossRef]
- Jahirul Islam, M.; Nawal Islam, N.; Siddik Alom, M.; Kabir, M.; Halim, M.A. A Review on structural, non-structural, and accessory proteins of SARS-CoV-2: Highlighting drug target sites. Immunobiology 2022, 228, 152302. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Ye, Q.; Singh, D.; Villa, E.; Cleveland, D.W.; Corbett, K.D. The SARS-CoV-2 Nucleocapsid phosphoprotein forms mutually exclusive condensates with RNA and the membrane-associated M protein. Nat. Commun. 2020, 12, 502. [Google Scholar] [CrossRef]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Iserman, C.; Roden, C.A.; Boerneke, M.A.; Sealfon, R.S.G.; McLaughlin, G.A.; Jungreis, I.; Fritch, E.J.; Hou, Y.J.; Ekena, J.; Weidmann, C.A.; et al. Genomic RNA elements drive phase separation of the SARS-CoV-2 nucleocapsid. Mol. Cell 2020, 80, 1078–1091.e6. [Google Scholar] [CrossRef] [PubMed]
- Perdikari, T.M.; Murthy, A.C.; Ryan, V.H.; Watters, S.; Naik, M.T.; Fawzi, N.L. SARS-CoV-2 nucleocapsid protein phase-separates with RNA and with human hnRNPs. EMBO J. 2020, 39, e106478. [Google Scholar] [CrossRef]
- Syed, A.M.; Taha, T.Y.; Tabata, T.; Chen, I.P.; Ciling, A.; Khalid, M.M.; Sreekumar, B.; Chen, P.Y.; Hayashi, J.M.; Soczek, K.M.; et al. Rapid assessment of SARS-CoV-2-evolved variants using virus-like particles. Science 2021, 374, 1626–1632. [Google Scholar] [CrossRef]
- Cao, Y.; Yang, R.; Lee, I.; Zhang, W.; Sun, J.; Wang, W.; Meng, X. Characterization of the SARS-CoV-2 E protein: Sequence, structure, viroporin, and inhibitors. Protein Sci. 2021, 30, 1114–1130. [Google Scholar] [CrossRef]
- Kuzmin, A.; Orekhov, P.; Astashkin, R.; Gordeliy, V.; Gushchin, I. Structure and dynamics of the SARS-CoV-2 envelope protein monomer. Proteins 2022, 90, 1102–1114. [Google Scholar] [CrossRef]
- Zhang, Z.; Nomura, N.; Muramoto, Y.; Ekimoto, T.; Uemura, T.; Liu, K.; Yui, M.; Kono, N.; Aoki, J.; Ikeguchi, M.; et al. Structure of SARS-CoV-2 membrane protein essential for virus assembly. Nat. Commun. 2022, 13, 4399. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, A.; Garg, N.; Giri, R. An insight into SARS-CoV-2 membrane protein interaction with spike, envelope, and nucleocapsid proteins. J. BioMol. Struct. Dyn. 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiao, T.; Cai, Y.; Chen, B. Structure of SARS-CoV-2 spike protein. Curr. Opin. Virol. 2021, 50, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Papa, G.; Mallery, D.L.; Albecka, A.; Welch, L.G.; Cattin-Ortolá, J.; Luptak, J.; Paul, D.; McMahon, H.T.; Goodfellow, I.G.; Carter, A.; et al. Furin cleavage of SARS-CoV-2 Spike promotes but is not essential for infection and cell-cell fusion. PLoS Pathog. 2021, 17, e1009246. [Google Scholar] [CrossRef]
- Henderson, R.; Edwards, R.J.; Mansouri, K.; Janowska, K.; Stalls, V.; Gobeil, S.M.C.; Kopp, M.; Li, D.; Parks, R.; Hsu, A.L.; et al. Controlling the SARS-CoV-2 spike glycoprotein conformation. Nat. Struct. Mol. Biol. 2020, 27, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Icho, S.; Rujas, E.; Muthuraman, K.; Tam, J.; Liang, H.; Landreth, S.; Liao, M.; Falzarano, D.; Julien, J.P.; Melnyk, R.A. Dual inhibition of vacuolar-ATPase and TMPRSS2 is required for complete blockade of SARS-CoV-2 entry into cells. Antimicrob. Agents Chemother. 2022, 66, e0043922. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Canziani, G.A.; Carter, E.P.; Chaiken, I. The case for S2: The potential benefits of the S2 subunit of the SARS-CoV-2 spike protein as an immunogen in fighting the COVID-19 pandemic. Front. Immunol. 2021, 12, 637651. [Google Scholar] [CrossRef]
- Jin, S.; He, X.; Ma, L.; Zhuang, Z.; Wang, Y.; Lin, M.; Cai, S.; Wei, L.; Wang, Z.; Zhao, Z.; et al. Suppression of ACE2 SUMOylation protects against SARS-CoV-2 infection through TOLLIP-mediated selective autophagy. Nat. Commun. 2022, 13, 5204. [Google Scholar] [CrossRef]
- Redondo, N.; Zaldívar-López, S.; Garrido, J.J.; Montoya, M. SARS-CoV-2 accessory proteins in viral pathogenesis: Knowns and unknowns. Front. Immunol. 2021, 12, 708264. [Google Scholar] [CrossRef]
- Hayn, M.; Hirschenberger, M.; Koepke, L.; Nchioua, R.; Straub, J.H.; Klute, S.; Hunszinger, V.; Zech, F.; Prelli Bozzo, C.; Aftab, W.; et al. Systematic functional analysis of SARS-CoV-2 proteins uncovers viral innate immune antagonists and remaining vulnerabilities. Cell Rep. 2021, 35, 109126. [Google Scholar] [CrossRef]
- Ren, Y.; Shu, T.; Wu, D.; Mu, J.; Wang, C.; Huang, M.; Han, Y.; Zhang, X.Y.; Zhou, W.; Qiu, Y.; et al. The ORF3a protein of SARS-CoV-2 induces apoptosis in cells. Cell Mol. Immunol. 2020, 17, 881–883. [Google Scholar] [CrossRef] [PubMed]
- Silvas, J.A.; Vasquez, D.M.; Park, J.G.; Chiem, K.; Allué-Guardia, A.; Garcia-Vilanova, A.; Platt, R.N.; Miorin, L.; Kehrer, T.; Cupic, A.; et al. Contribution of SARS-CoV-2 accessory proteins to viral pathogenicity in K18 human ACE2 transgenic mice. J. Virol. 2021, 95, e0040221. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Liao, C.H.; Wang, Q.; Tan, Y.J.; Luo, R.; Qiu, Y.; Ge, X.Y. The ORF6, ORF8 and nucleocapsid proteins of SARS-CoV-2 inhibit type I interferon signaling pathway. Virus Res. 2020, 286, 198074. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zhao, Q.; Rao, J.; Zeng, F.; Yuan, S.; Ji, M.; Sun, X.; Li, J.; Yang, J.; Cui, J.; et al. SARS-CoV-2 Accessory protein ORF7b mediates tumor necrosis factor-α-induced apoptosis in cells. Front. Microbiol. 2021, 12, 654709. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.E.; Hiatt, J.; Bouhaddou, M.; Rezelj, V.V.; Ulferts, S.; Braberg, H.; Jureka, A.S.; Obernier, K.; Guo, J.Z.; Batra, J.; et al. Comparative host-coronavirus protein interaction networks reveal pan-viral disease mechanisms. Science 2020, 370, eabe9403. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhu, K.; Qin, B.; Olieric, V.; Wang, M.; Cui, S. Crystal structure of SARS-CoV-2 Orf9b in complex with human TOM70 suggests unusual virus-host interactions. Nat. Commun. 2021, 12, 2843. [Google Scholar] [CrossRef]
- Lin, R.; Paz, S.; Hiscott, J. Tom70 imports antiviral immunity to the mitochondria. Cell Res. 2010, 20, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Eskier, D.; Suner, A.; Oktay, Y.; Karakülah, G. Mutations of SARS-CoV-2 nsp14 exhibit strong association with increased genome-wide mutation load. PeerJ 2020, 8, e10181. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.R.; Rennick, L.J.; Nambulli, S.; Robinson-McCarthy, L.R.; Bain, W.G.; Haidar, G.; Duprex, W.P. Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape. Science 2021, 371, 1139–1142. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; De Lorenzo, G.; Furnon, W.; Cantoni, D.; Scott, S.; Logan, N.; Ashraf, S.; et al. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nat. Microbiol. 2022, 7, 1161–1179. [Google Scholar] [CrossRef] [PubMed]
- SARS-CoV-2 (hCoV-19) Mutation Reports. Available online: https://outbreak.info/situation-reports/omicron (accessed on 15 November 2022).
- Barton, M.I.; MacGowan, S.A.; Kutuzov, M.A.; Dushek, O.; Barton, G.J.; van der Merwe, P.A. Effects of common mutations in the SARS-CoV-2 Spike RBD and its ligand, the human ACE2 receptor on binding affinity and kinetics. eLife 2021, 10, e70658. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, B.; Fernandes, M.H.V.; Frazier, L.; Tang, T.; Daniel, S.; Diel, D.G.; Jaimes, J.A.; Whittaker, G.R. Functional evaluation of the P681H mutation on the proteolytic activation of the SARS-CoV-2 variant B.1.1.7 (Alpha) spike. iScience 2022, 25, 103589. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 Infection of Animal Hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef]
- SARS-CoV-2 in Animals—Situation Report 15. Available online: https://www.woah.org/en/document/86934/ (accessed on 6 November 2022).
- Gryseels, S.; De Bruyn, L.; Gyselings, R.; Calvignac-Spencer, S.; Leendertz, F.H.; Leirs, H. Risk of human-to-wildlife transmission of SARS-CoV-2. Mamm. Rev. 2021, 51, 272–292. [Google Scholar] [CrossRef]
- Tan, C.C.S.; Lam, S.D.; Richard, D.; Owen, C.J.; Berchtold, D.; Orengo, C.; Nair, M.S.; Kuchipudi, S.V.; Kapur, V.; van Dorp, L.; et al. Transmission of SARS-CoV-2 from humans to animals and potential host adaptation. Nat. Commun. 2022, 13, 2988. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Levy, J.I.; De Hoff, P.; Humphrey, G.; Birmingham, A.; Jepsen, K.; Farmer, S.; Tubb, H.M.; Valles, T.; Tribelhorn, C.E.; et al. Wastewater sequencing reveals early cryptic SARS-CoV-2 variant transmission. Nature 2022, 609, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.N.; Le, G.T.; Nguyen, D.T.; Juang, R.S.; Rinklebe, J.; Bhatnagar, A.; Lima, E.C.; Iqbal, H.M.N.; Sarmah, A.K.; Chao, H.P. SARS-CoV-2 coronavirus in water and wastewater: A critical review about presence and concern. Environ. Res. 2021, 193, 110265. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Z.; Yin, L.; Zhang, Q.; Askarinam, N.; Mundaca-Uribe, R.; Tehrani, F.; Karshalev, E.; Gao, W.; Zhang, L.; et al. ACE2 receptor-modified algae-based microrobot for removal of SARS-CoV-2 in wastewater. J. Am. Chem. Soc. 2021, 143, 12194–12201. [Google Scholar] [CrossRef] [PubMed]
- Desdouits, M.; Piquet, J.C.; Wacrenier, C.; Le Mennec, C.; Parnaudeau, S.; Jousse, S.; Rocq, S.; Bigault, L.; Contrant, M.; Garry, P.; et al. Can shellfish be used to monitor SARS-CoV-2 in the coastal environment? Sci. Total Environ. 2021, 778, 146270. [Google Scholar] [CrossRef] [PubMed]
- Polo, D.; Lois, M.; Fernández-Núñez, M.T.; Romalde, J.L. Detection of SARS-CoV-2 RNA in bivalve mollusks and marine sediments. Sci. Total Environ. 2021, 786, 147534. [Google Scholar] [CrossRef]
- Seyer, A. The fate of SARS-CoV-2 in the marine environments: Are marine environments safe of COVID-19? Erciyes Med. J. 2021, 43, 606–607. [Google Scholar]
- Mordecai, G.J.; Miller, K.M.; Di Cicco, E.; Schulze, A.D.; Kaukinen, K.H.; Ming, T.J.; Li, S.; Tabata, A.; Teffer, A.; Patterson, D.A.; et al. Endangered wild salmon infected by newly discovered viruses. eLife 2019, 8, e47615. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, C.; Báez, J.C. Placing the COVID-19 Pandemic in a Marine Ecological Context: Potential risks for conservation of marine air-breathing animals and future zoonotic outbreaks. Front. Mar. Sci. 2021, 8, 691682. [Google Scholar] [CrossRef]
- Barbosa, A.; Varsani, A.; Morandini, V.; Grimaldi, W.; Vanstreels, R.E.T.; Diaz, J.I.; Boulinier, T.; Dewar, M.; González-Acuña, D.; Gray, R.; et al. Risk assessment of SARS-CoV-2 in Antarctic wildlife. Sci. Total Environ. 2021, 755, 143352. [Google Scholar] [CrossRef]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.-P.; Pfenning, A.R.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Mathavarajah, S.; Stoddart, A.K.; Gagnon, G.A.; Dellaire, G. Pandemic danger to the deep: The risk of marine mammals contracting SARS-CoV-2 from wastewater. Sci. Total Environ. 2021, 760, 143346. [Google Scholar] [CrossRef] [PubMed]
- Audino, T.; Grattarola, C.; Centelleghe, C.; Peletto, S.; Giorda, F.; Florio, C.L.; Caramelli, M.; Bozzetta, E.; Mazzariol, S.; Di Guardo, G.; et al. SARS-CoV-2, a threat to marine mammals? A study from Italian seawaters. Animals 2021, 11, 1663. [Google Scholar] [CrossRef]
- Xie, S.Z.; Liu, M.Q.; Jiang, R.D.; Lin, H.F.; Zhang, W.; Li, B.; Su, J.; Ke, F.; Zhang, Q.Y.; Shi, Z.L.; et al. Fish ACE2 is not susceptible to SARS-CoV-2. Virol. Sin. 2022, 37, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Kaki, M.; Potluri, V.S.; Kahar, P.; Khanna, D. A comprehensive review of SARS-CoV-2 vaccines: Pfizer, Moderna & Johnson & Johnson. Hum. Vaccines Immunother. 2022, 18, 2002083. [Google Scholar]
- Gobeil, P.; Pillet, S.; Boulay, I.; Charland, N.; Lorin, A.; Cheng, M.P.; Vinh, D.C.; Boutet, P.; Van Der Most, R.; Roman, F.; et al. Durability and cross-reactivity of immune responses induced by a plant-based virus-like particle vaccine for COVID-19. Nat. Commun. 2022, 13, 6905. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M.; Krammer, F.; Regev-Yochay, G.; Lustig, Y.; Balicer, R.D. SARS-CoV-2 breakthrough infections in vaccinated individuals: Measurement, causes and impact. Nat. Rev. Immunol. 2022, 22, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Götte, M. Remdesivir for the treatment of COVID-19: The value of biochemical studies. Curr. Opin. Virol. 2021, 49, 81–85. [Google Scholar] [CrossRef]
- Iglesias Gómez, R.; Méndez, R.; Palanques-Pastor, T.; Ballesta-López, O.; Borrás Almenar, C.; Megías Vericat, J.E.; López-Briz, E.; Font-Noguera, I.; Menéndez Villanueva, R.; Román Iborra, J.A.; et al. Baricitinib against severe COVID-19: Effectiveness and safety in hospitalised pretreated patients. Eur. J. Hosp. Pharm. 2022, 29, e41–e45. [Google Scholar] [CrossRef] [PubMed]
- Titanji, B.K.; Farley, M.M.; Mehta, A.; Connor-Schuler, R.; Moanna, A.; Cribbs, S.K.; O’Shea, J.; DeSilva, K.; Chan, B.; Edwards, A.; et al. Use of baricitinib in patients with moderate to severe coronavirus disease 2019. Clin. Infect. Dis. 2021, 72, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gan, J.; Wang, R.; Yang, X.; Xiao, Z.; Cao, Y. DrugDevCovid19: An atlas of anti-COVID-19 compounds derived by computer-aided drug design. Molecules 2022, 27, 683. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Ding, C.; Jiang, L.; Tang, W.; Liu, Y.; Zhao, L.; Yi, Z.; Ren, H.; Li, C.; He, Y.; et al. Discovery of potential anti-SARS-CoV-2 drugs based on large-scale screening in vitro and effect evaluation in vivo. Sci. China Life Sci. 2022, 65, 1181–1197. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.-H. Anti–SARS-CoV-2 natural products as potentially therapeutic agents. Front. Pharmacol. 2021, 12, 590509. [Google Scholar] [CrossRef]
- Rahman, M.M.; Islam, M.R.; Shohag, S.; Hossain, M.E.; Shah, M.; Shuvo, S.K.; Khan, H.; Chowdhury, M.A.R.; Bulbul, I.J.; Hossain, M.S.; et al. Multifaceted role of natural sources for COVID-19 pandemic as marine drugs. Environ. Sci. Pollut. Res. Int. 2022, 29, 46527–46550. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.; Jayaraman, G. Marine sulfated polysaccharides as potential antiviral drug candidates to treat Corona Virus disease (COVID-19). Carbohydr. Res. 2021, 505, 108326. [Google Scholar] [CrossRef] [PubMed]
- Neufurth, M.; Wang, X.; Tolba, E.; Lieberwirth, I.; Wang, S.; Schröder, H.C.; Müller, W.E.G. The inorganic polymer, polyphosphate, blocks binding of SARS-CoV-2 spike protein to ACE2 receptor at physiological concentrations. Biochem. Pharmacol. 2020, 182, 114215. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Neufurth, M.; Schepler, H.; Wang, S.; Tolba, E.; Schröder, H.C.; Wang, X. The biomaterial polyphosphate blocks stoichiometric binding of the SARS-CoV-2 S-protein to the cellular ACE2 receptor. Biomater. Sci. 2020, 8, 6603–6610. [Google Scholar] [CrossRef]
- Petit, L.; Vernès, L.; Cadoret, J.P. Docking and in silico toxicity assessment of Arthrospira compounds as potential antiviral agents against SARS-CoV-2. J. Appl. Phycol. 2021, 33, 1579–1602. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Van Damme, E.J.M.; Simplicien, M.; Le Poder, S.; Klonjkowski, B.; Benoist, H.; Peyrade, D.; Rougé, P. Man-specific lectins from plants, fungi, algae and cyanobacteria, as potential blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) coronaviruses: Biomedical perspectives. Cells 2021, 10, 1619. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, M.; Gunaseelan, S.; Kubendran Aravind, M.; Mohankumar, V.; Anupam, P.; Harikrishnan, M.; Siva, A.; Ashokkumar, B.; Varalakshmi, P. Marine algal antagonists targeting 3CL protease and spike glycoprotein of SARS-CoV-2: A computational approach for anti-COVID-19 drug discovery. J. BioMol. Struct. Dyn. 2021, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Patamia, V.; Scala, A.; Sciortino, M.T.; Piperno, A.; Rescifina, A. Putative inhibitors of SARS-CoV-2 main protease from a library of marine natural products: A virtual screening and molecular modeling study. Mar. Drugs 2020, 18, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Demerdash, A.; Metwaly, A.M.; Hassan, A.; Abd El-Aziz, T.M.; Elkaeed, E.B.; Eissa, I.H.; Arafa, R.K.; Stockand, J.D. Comprehensive virtual screening of the antiviral potentialities of marine polycyclic guanidine alkaloids against SARS-CoV-2 (COVID-19). Biomolecules 2021, 11, 460. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Ali, A.; Wang, Q.; Irfan, M.; Khan, A.; Zeb, M.T.; Zhang, Y.J.; Chinnasamy, S.; Wei, D.Q. Marine natural compounds as potents inhibitors against the main protease of SARS-CoV-2-a molecular dynamic study. J. BioMol. Struct. Dyn. 2021, 39, 3627–3637. [Google Scholar] [CrossRef]
- Quimque, M.T.J.; Notarte, K.I.R.; Fernandez, R.A.T.; Mendoza, M.A.O.; Liman, R.A.D.; Lim, J.A.K.; Pilapil, L.A.E.; Ong, J.K.H.; Pastrana, A.M.; Khan, A.; et al. Virtual screening-driven drug discovery of SARS-CoV2 enzyme inhibitors targeting viral attachment, replication, post-translational modification and host immunity evasion infection mechanisms. J. BioMol. Struct. Dyn. 2021, 39, 4316–4333. [Google Scholar] [CrossRef] [PubMed]
- Abdelrheem, D.A.; Ahmed, S.A.; Abd El-Mageed, H.R.; Mohamed, H.S.; Rahman, A.A.; Elsayed, K.N.M.; Ahmed, S.A. The inhibitory effect of some natural bioactive compounds against SARS-CoV-2 main protease: Insights from molecular docking analysis and molecular dynamic simulation. J. Environ. Sci. Health A Toxic Hazard. Subst. Environ. Eng. 2020, 55, 1373–1386. [Google Scholar] [CrossRef]
- Pendyala, B.; Patras, A.; Dash, C. Phycobilins as potent food bioactive broad-spectrum inhibitors against proteases of SARS-CoV-2 and other coronaviruses: A preliminary study. Front. Microbiol. 2021, 12, 645713. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraj, R.; Altaff, K.; Rosita, A.S.; Ramadevi, S.; Revathy, J. Bioactive compounds from marine resources against novel corona virus (2019-nCoV): In silico study for corona viral drug. Nat. Prod. Res. 2021, 35, 5525–5529. [Google Scholar] [CrossRef] [PubMed]
- Bhati, S. Structure-based drug designing of naphthalene based SARS-CoV PLpro inhibitors for the treatment of COVID-19. Heliyon 2020, 6, e05558. [Google Scholar] [CrossRef]
- Hamoda, A.M.; Fayed, B.; Ashmawy, N.S.; El-Shorbagi, A.A.; Hamdy, R.; Soliman, S.S.M. Marine sponge is a promising natural source of anti-SARS-CoV-2 scaffold. Front. Pharmacol. 2021, 12, 666664. [Google Scholar] [CrossRef] [PubMed]
- Mahmudpour, M.; Nabipour, I.; Keshavarz, M.; Farrokhnia, M. Virtual screening on marine natural products for discovering TMPRSS2 inhibitors. Front. Chem. 2021, 9, 722633. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, V.; Kong, D.Y.; Asadzadeh, F.; Marrone, L.; Boccia, A.; Siciliano, R.; Criscuolo, G.; Anastasio, C.; Quarantelli, F.; Comegna, M.; et al. Long-chain polyphosphates impair SARS-CoV-2 infection and replication. Sci. Signal 2021, 14, eabe5040. [Google Scholar] [CrossRef] [PubMed]
- Sahu, N.; Mishra, S.; Kesheri, M.; Kanchan, S.; Sinha, R.P. Identification of cyanobacteria-based natural inhibitors against SARS-CoV-2 druggable target ACE2 using molecular docking study, ADME and toxicity analysis. Indian J. Clin. Biochem. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Neufurth, M.; Wang, S.; Tan, R.; Schröder, H.C.; Wang, X. Morphogenetic (mucin expression) as well as potential anti-corona viral activity of the marine secondary metabolite polyphosphate on A549 cells. Mar. Drugs 2020, 18, 639. [Google Scholar] [CrossRef] [PubMed]
- Leisch, M.; Egle, A.; Greil, R. Plitidepsin: A potential new treatment for relapsed/refractory multiple myeloma. Future Oncol. 2019, 15, 109–120. [Google Scholar] [CrossRef]
- White, K.M.; Rosales, R.; Yildiz, S.; Kehrer, T.; Miorin, L.; Moreno, E.; Jangra, S.; Uccellini, M.B.; Rathnasinghe, R.; Coughlan, L.; et al. Plitidepsin has potent preclinical efficacy against SARS-CoV-2 by targeting the host protein eEF1A. Science 2021, 371, 926–931. [Google Scholar] [CrossRef]
- Xie, Y.; Karki, C.B.; Du, D.; Li, H.; Wang, J.; Sobitan, A.; Teng, S.; Tang, Q.; Li, L. Spike proteins of SARS-CoV and SARS-CoV-2 utilize different mechanisms to bind with human ACE2. Front. Mol. Biosci. 2020, 7, 392. [Google Scholar] [CrossRef]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Schütz, D.; Conzelmann, C.; Fois, G.; Groß, R.; Weil, T.; Wettstein, L.; Stenger, S.; Zelikin, A.; Hoffmann, T.K.; Frick, M.; et al. Carrageenan-containing over-the-counter nasal and oral sprays inhibit SARS-CoV-2 infection of airway epithelial cultures. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L750–L756. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Shin, H.; Lee, M.K.; Kwon, O.S.; Shin, J.S.; Kim, Y.I.; Kim, C.W.; Lee, H.R.; Kim, M. Antiviral activity of lambda-carrageenan against influenza viruses and severe acute respiratory syndrome coronavirus 2. Sci. Rep. 2021, 11, 821. [Google Scholar] [CrossRef]
- Kornberg, A.; Rao, N.N.; Ault-Riché, D. Inorganic polyphosphate: A molecule of many functions. Annu. Rev. Biochem. 1999, 68, 89–125. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.; Ingall, E.; Benitez-Nelson, C.; Paterson, D.; de Jonge, M.D.; McNulty, I.; Brandes, J.A. Marine polyphosphate: A key player in geologic phosphorus sequestration. Science 2008, 320, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Li, M.; Wu, S.; Jia, L.; Hill, R.T.; Zhao, J. Characteristic microbiomes correlate with polyphosphate accumulation of marine sponges in South China Sea areas. Microorganisms 2020, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufurth, M.; Wang, X.; Wang, S.; Schröder, H.C.; Müller, W.E.G. Caged dexamethasone/quercetin nanoparticles, formed of the morphogenetic active inorganic polyphosphate, are strong inducers of MUC5AC. Mar. Drugs 2021, 19, 64. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Mutch, N.J.; Schenk, W.A.; Smith, S.A.; Esterl, L.; Spronk, H.M.; Schmidbauer, S.; Gahl, W.A.; Morrissey, J.H.; Renné, T. Platelet polyphosphates are proinflammatory and procoagulant mediators in vivo. Cell 2009, 139, 1143–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.A.; Choi, S.H.; Davis-Harrison, R.; Huyck, J.; Boettcher, J.; Rienstra, C.M.; Morrissey, J.H. Polyphosphate exerts differential effects on blood clotting, depending on polymer size. Blood 2010, 116, 4353–4359. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wan, M.; Liang, T.; Peng, M.; Chen, F. Synthetic polyphosphate inhibits endogenous coagulation and platelet aggregation in vitro. Biomed. Rep. 2017, 6, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, T.; Levy, J.H.; Levi, M.; Connors, J.M.; Thachil, J. Coagulopathy of coronavirus disease 2019. Crit. Care Med. 2020, 48, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Freedman, J.E. Platelets and COVID-19: Inflammation, hyperactivation and additional questions. Circ. Res. 2020, 127, 1419–1421. [Google Scholar] [CrossRef] [PubMed]
- Schepler, H.; Wang, X.; Neufurth, M.; Wang, S.; Schröder, H.C.; Müller, W.E.G. The therapeutic potential of inorganic polyphosphate: A versatile physiological polymer to control coronavirus disease (COVID-19). Theranostics 2021, 11, 6193–6213. [Google Scholar] [CrossRef]
- Lempp, F.A.; Soriaga, L.B.; Montiel-Ruiz, M.; Benigni, F.; Noack, J.; Park, Y.-J.; Bianchi, S.; Walls, A.C.; Bowen, J.E.; Zhou, J.; et al. Lectins enhance SARS-CoV-2 infection and influence neutralizing antibodies. Nature 2021, 598, 342–347. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Conte-Junior, C.A.; Domínguez, R.; Nawaz, A.; Walayat, N.; Movilla Fierro, E.; Lorenzo, J.M. Marine alkaloids: Compounds with in vivo activity and chemical synthesis. Mar. Drugs 2021, 19, 374. [Google Scholar] [CrossRef] [PubMed]
- Elissawy, A.M.; Soleiman Dehkordi, E.; Mehdinezhad, N.; Ashour, M.L.; Mohammadi Pour, P. Cytotoxic alkaloids derived from marine sponges: A comprehensive review. Biomolecules 2021, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Sachse, M.; Tenorio, R.; Fernández de Castro, I.; Muñoz-Basagoiti, J.; Perez-Zsolt, D.; Raïch-Regué, D.; Rodon, J.; Losada, A.; Avilés, P.; Cuevas, C.; et al. Unraveling the antiviral activity of plitidepsin against SARS-CoV-2 by subcellular and morphological analysis. Antivir. Res. 2022, 200, 105270. [Google Scholar] [CrossRef] [PubMed]
- Vishvakarma, V.K.; Singh, M.B.; Jain, P.; Kumari, K.; Singh, P. Hunting the main protease of SARS-CoV-2 by plitidepsin: Molecular docking and temperature-dependent molecular dynamics simulations. Amino Acids 2022, 54, 205–213. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic view of SARS-CoV-2 genome and its replication and transcription. (A) Genome structure of SARS-CoV-2. (B) SARS-CoV-2 genome replication and expression. The positive-sense single stranded RNA of SARS-CoV-2 can act as template guiding the synthesis of negative-sense gRNA for genome replication to (+) gRNA. The (+) gRNA can serve as mRNA for translation of the long ORF1a and ORF1ab at its 5′-terminus of genome, and the transcription of the 3′-terminus of its genome is achieved by the discontinuous synthesis of a set of subgenomic mRNAs (sgRNA) through a template switch mechanism by jumping from the transcription regulatory sequence in the genome body (TRSB; indicated by arrows) to the leader TRS (TRSL). The sgRNAs guide the synthesis of structural proteins and some accessory proteins.
Figure 1.
Schematic view of SARS-CoV-2 genome and its replication and transcription. (A) Genome structure of SARS-CoV-2. (B) SARS-CoV-2 genome replication and expression. The positive-sense single stranded RNA of SARS-CoV-2 can act as template guiding the synthesis of negative-sense gRNA for genome replication to (+) gRNA. The (+) gRNA can serve as mRNA for translation of the long ORF1a and ORF1ab at its 5′-terminus of genome, and the transcription of the 3′-terminus of its genome is achieved by the discontinuous synthesis of a set of subgenomic mRNAs (sgRNA) through a template switch mechanism by jumping from the transcription regulatory sequence in the genome body (TRSB; indicated by arrows) to the leader TRS (TRSL). The sgRNAs guide the synthesis of structural proteins and some accessory proteins.

Figure 2.
Cleavage of pp1a and pp1ab by PLpro and Mpro. (A) Composition and cleavage of pp1a. (B) Composition and cleavage of pp1ab. Arrows represent the cleavages by PLpro, and triangles represent the cleavages by Mpro. The polyprotein pp1a is cleaved by PLpro and Mpro into 11 non-structural proteins (nsps) after its synthesis at their respective sites. The pp1ab is synthesized by translating the ORF1ab with the programmed −1 ribosomal frameshift (−1 PRF) on nsp11 and cleaved by PLpro and Mpro into 15 nsps.
Figure 2.
Cleavage of pp1a and pp1ab by PLpro and Mpro. (A) Composition and cleavage of pp1a. (B) Composition and cleavage of pp1ab. Arrows represent the cleavages by PLpro, and triangles represent the cleavages by Mpro. The polyprotein pp1a is cleaved by PLpro and Mpro into 11 non-structural proteins (nsps) after its synthesis at their respective sites. The pp1ab is synthesized by translating the ORF1ab with the programmed −1 ribosomal frameshift (−1 PRF) on nsp11 and cleaved by PLpro and Mpro into 15 nsps.

Figure 3.
Domain structures of SARS-CoV-2 N, E, M, and S structural proteins. (A) N protein. IDR, intrinsically disordered region. RBD, RNA binding domain. DD, dimerization domain. (B) E protein. NTD, N-terminal domain (NTD). TMD, transmembrane domain. CTD, C-terminal domain. (C) M protein. (D) S protein. RBD, receptor-binding domain. FP, fusion peptide. TM, transmembrane anchor. CT, cytoplasmic tail. The aa denotes the amino acids. The size of the diagram of S protein is not proportional to those of N, E, and M proteins.
Figure 3.
Domain structures of SARS-CoV-2 N, E, M, and S structural proteins. (A) N protein. IDR, intrinsically disordered region. RBD, RNA binding domain. DD, dimerization domain. (B) E protein. NTD, N-terminal domain (NTD). TMD, transmembrane domain. CTD, C-terminal domain. (C) M protein. (D) S protein. RBD, receptor-binding domain. FP, fusion peptide. TM, transmembrane anchor. CT, cytoplasmic tail. The aa denotes the amino acids. The size of the diagram of S protein is not proportional to those of N, E, and M proteins.

Figure 4.
Three-dimensional structures of ACE2 protein in huamn (GenBank No. NP_S.1) and marine yellow drum (GenBank No. KAG8001285.1) and large yellow croaker fish (GenBank No. XP_010730146.1). The tertiary structure oSSf ACE2 protein were predicted with SWISS-MODEL workspace (swissmodel.expasy.org) and visualized by Visual Molecular Dynamics 1.9.4a51 and Pymol. Blue represents the active sites of ACE2 (346–517aa residues), and red represents the 25 spike-interacting key residues and their substitutes.
Figure 4.
Three-dimensional structures of ACE2 protein in huamn (GenBank No. NP_S.1) and marine yellow drum (GenBank No. KAG8001285.1) and large yellow croaker fish (GenBank No. XP_010730146.1). The tertiary structure oSSf ACE2 protein were predicted with SWISS-MODEL workspace (swissmodel.expasy.org) and visualized by Visual Molecular Dynamics 1.9.4a51 and Pymol. Blue represents the active sites of ACE2 (346–517aa residues), and red represents the 25 spike-interacting key residues and their substitutes.


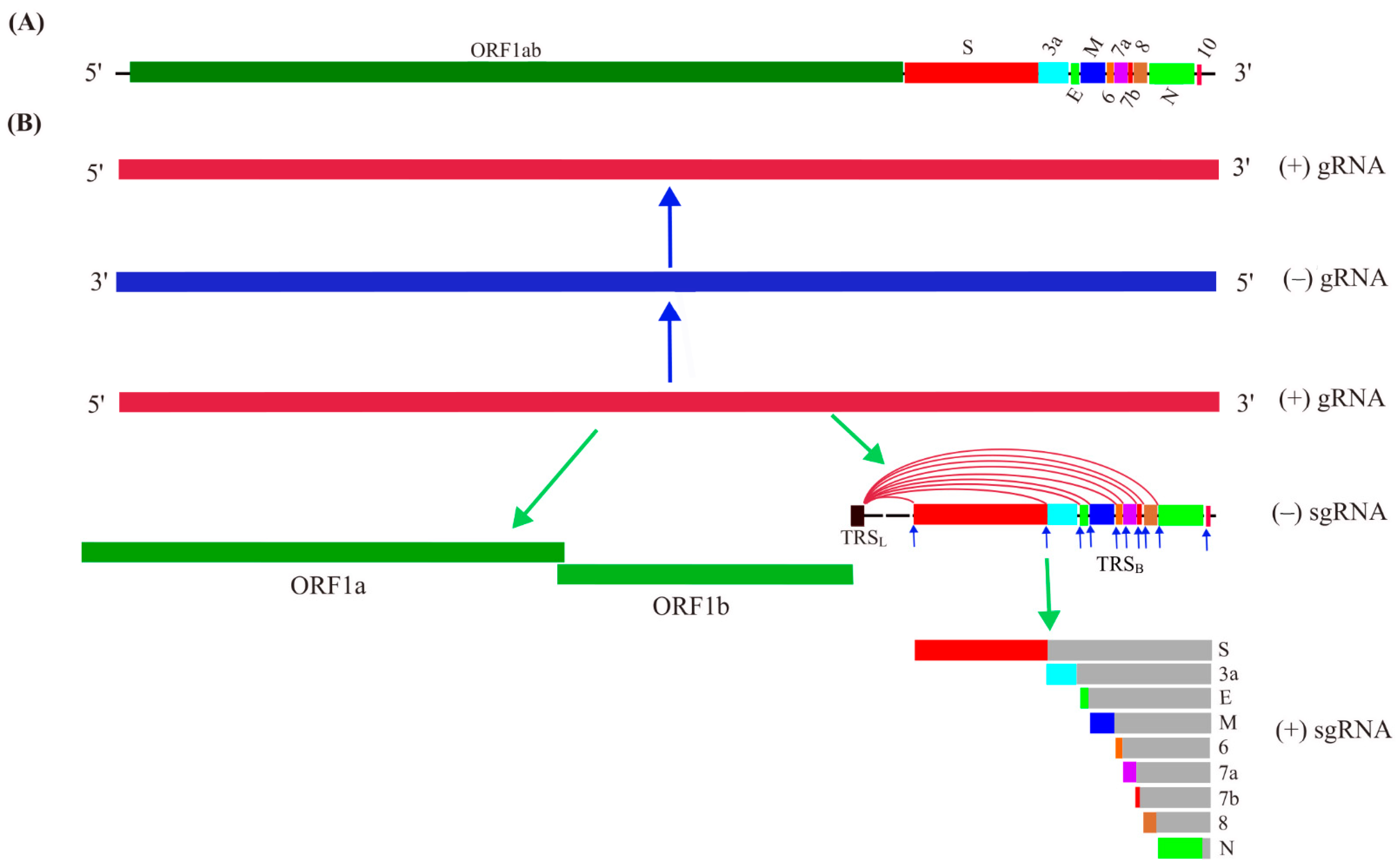{kind=link}
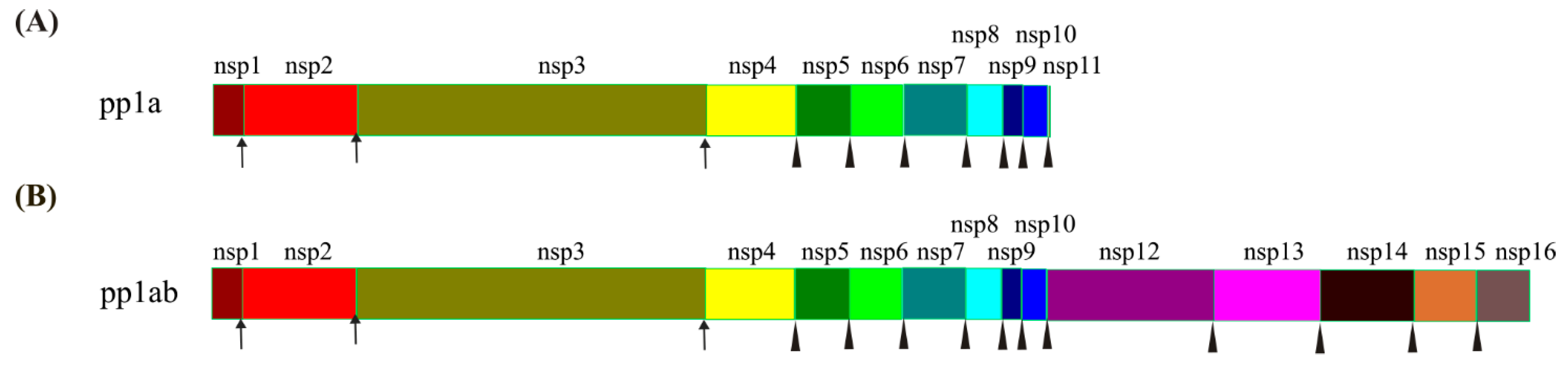{kind=link}
{kind=link}
{kind=link}
Table 1.
The basic description of the nonstructural proteins based on annotation of the GenBank ID NC_045512.2. and with reference to a recent review on SARS-CoV-2 genomic structure [29].
Table 1.
The basic description of the nonstructural proteins based on annotation of the GenBank ID NC_045512.2. and with reference to a recent review on SARS-CoV-2 genomic structure [29].
| Protein | Amino Acids | Function |
|---|---|---|
| nsp1 | 180 | Leader protein |
| nsp2 | 638 | Zinc-finger protein |
| nsp3 | 1945 | Papain-like proteinase (PLpro) |
| nsp4 | 500 | Tetra spanning transmembrane protein |
| nsp5 | 306 | 3C-like proteinase (3CLpro) or main protease (Mpro) |
| nsp6 | 290 | Transmembrane domain-containing protein |
| nsp7 | 353 | A component of primase complex with nsp8 and nsp12 |
| nsp8 | 198 | A component of primase complex with nsp7 and nsp12 |
| nsp9 | 113 | ssRNA-binding protein |
| nsp10 | 139 | Interacting with nsp14 and nsp16 |
| nsp11 | 13 | n.d.(not defined) |
| nsp12 | 932 | RNA-dependent RNA polymerase (RdRp) |
| nsp13 | 601 | Helicase |
| nsp14 | 527 | N7-Methyltransferase and 3′-5′ exonuclease (ExoN) |
| nsp15 | 346 | Uridine-specific endoribonuclease |
| nsp16 | 298 | 2′-O-ribose Methyltransferase |
Table 2.
Marine-derived compounds as potent inhibitors of SARS-CoV-2.
| Target | Marine Compound | Reference |
|---|---|---|
| Viral spike protein | Sulfated polysaccharides | [91] |
| Inorganic polyphosphates | [92,93] | |
| Phycobilins | [94] | |
| Mannose-specific lectins | [95] | |
| Mpro and/or PLpro | Polyphenols | [96,97] |
| Alkaloids | [98,99,100,101] | |
| Phycobilins | [102] | |
| Coumarin derivatives | [103] | |
| Naphthalene derivatives | [104] | |
| RdRp | Nucleoside analogues | [105] |
| TMPRSS2 | Watasenia preluciferyl β-D- glucopyranosiduronic acid | [106] |
| hACE2 | Inorganic polyphosphates | [107] |
| Mycosporin-like amino acids | [108] | |
| Immune system | Inorganic polyphosphates | [107,109] |
| Host eEF1A | Plitidepsin | [110,111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, Y.; Li, J.; Han, F. Focus on Marine Animal Safety and Marine Bioresources in Response to the SARS-CoV-2 Crisis. Int. J. Mol. Sci. 2022, 23, 15136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315136
AMA Style
Yang Y, Li J, Han F. Focus on Marine Animal Safety and Marine Bioresources in Response to the SARS-CoV-2 Crisis. International Journal of Molecular Sciences. 2022; 23(23):15136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315136
Chicago/Turabian StyleYang, Yao, Jiacheng Li, and Fang Han. 2022. "Focus on Marine Animal Safety and Marine Bioresources in Response to the SARS-CoV-2 Crisis" International Journal of Molecular Sciences 23, no. 23: 15136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315136
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.