
An Escherichia coli Expressed Multi-Disulfide Bonded SARS-CoV-2 RBD Shows Native-like Biophysical Properties and Elicits Neutralizing Antisera in a Mouse Model
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
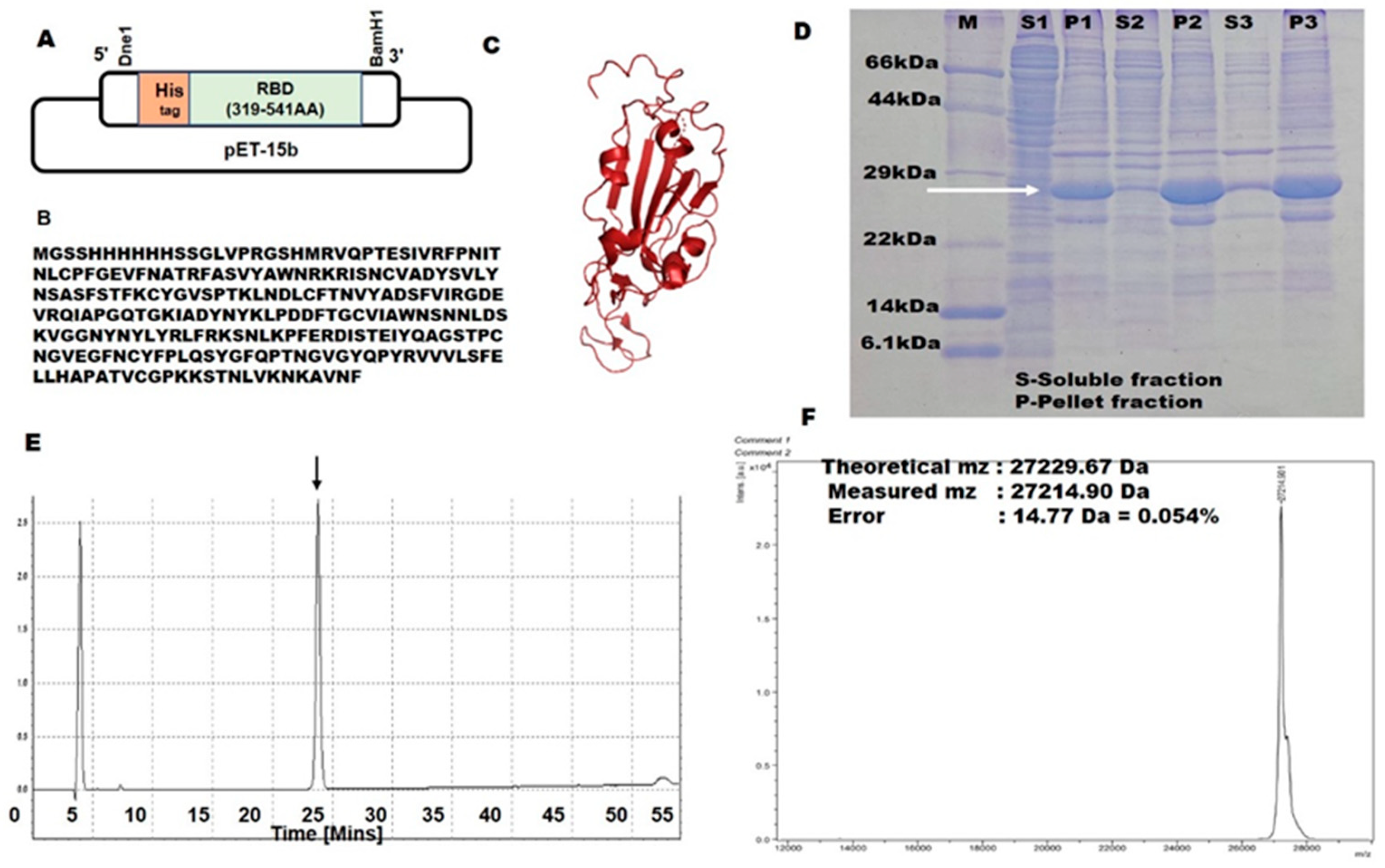
2.1. SARS-CoV-2 RBD Expression Using a Bacterial Expression System
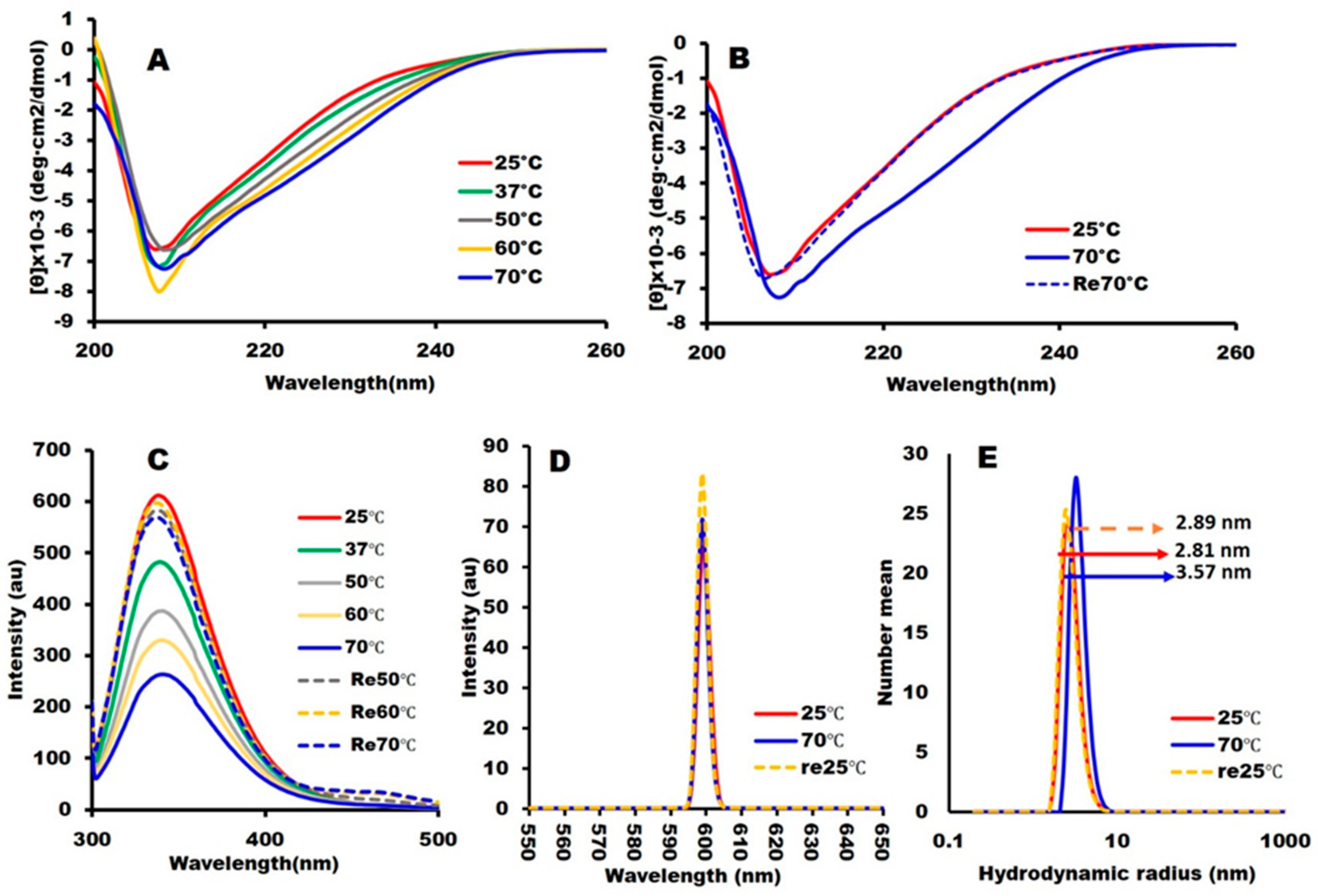
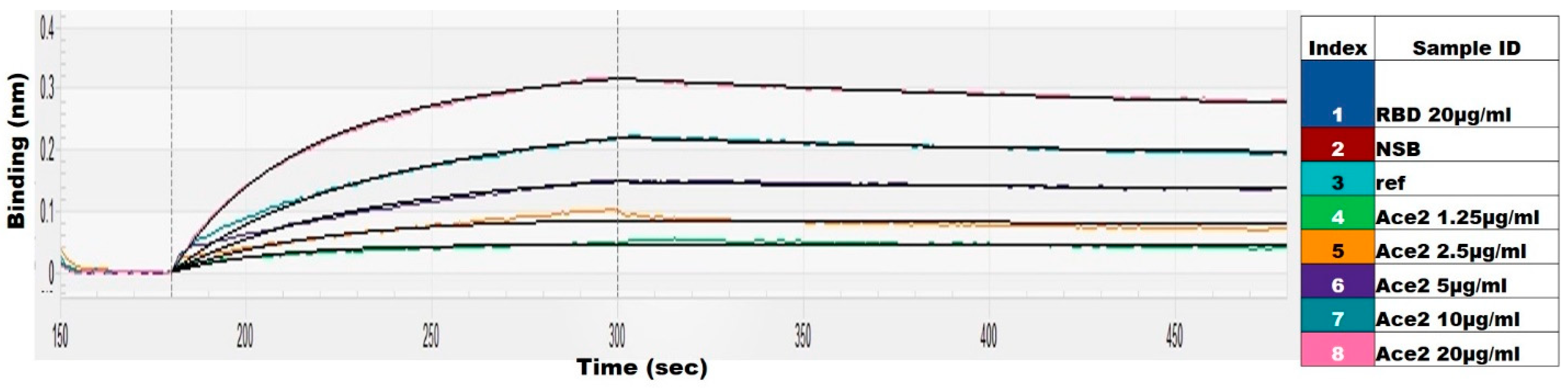
2.2. Structural Integrity and Binding Activity
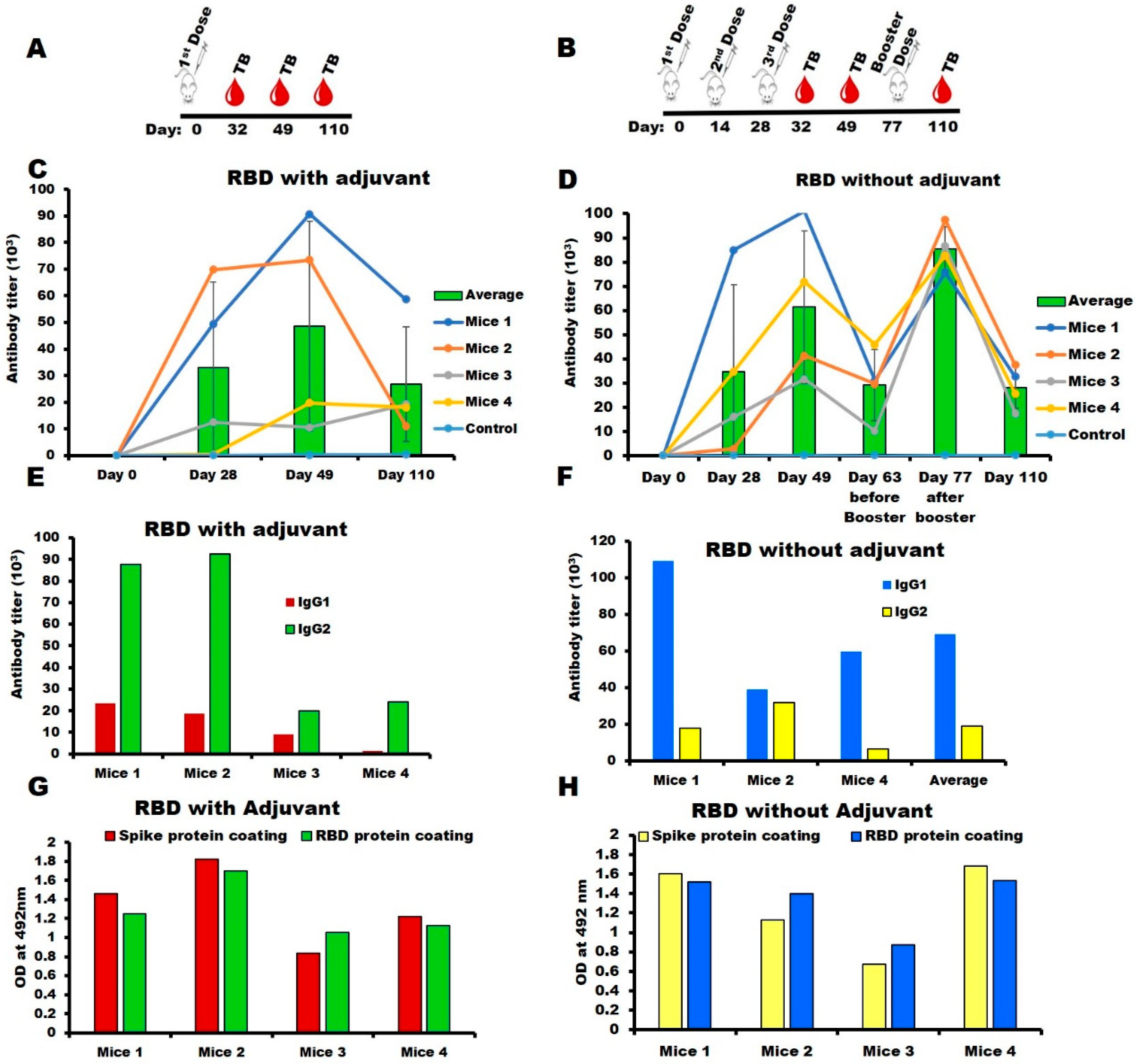
2.3. Immunogenicity of SARS-CoV-2-RBD in the Mouse Model
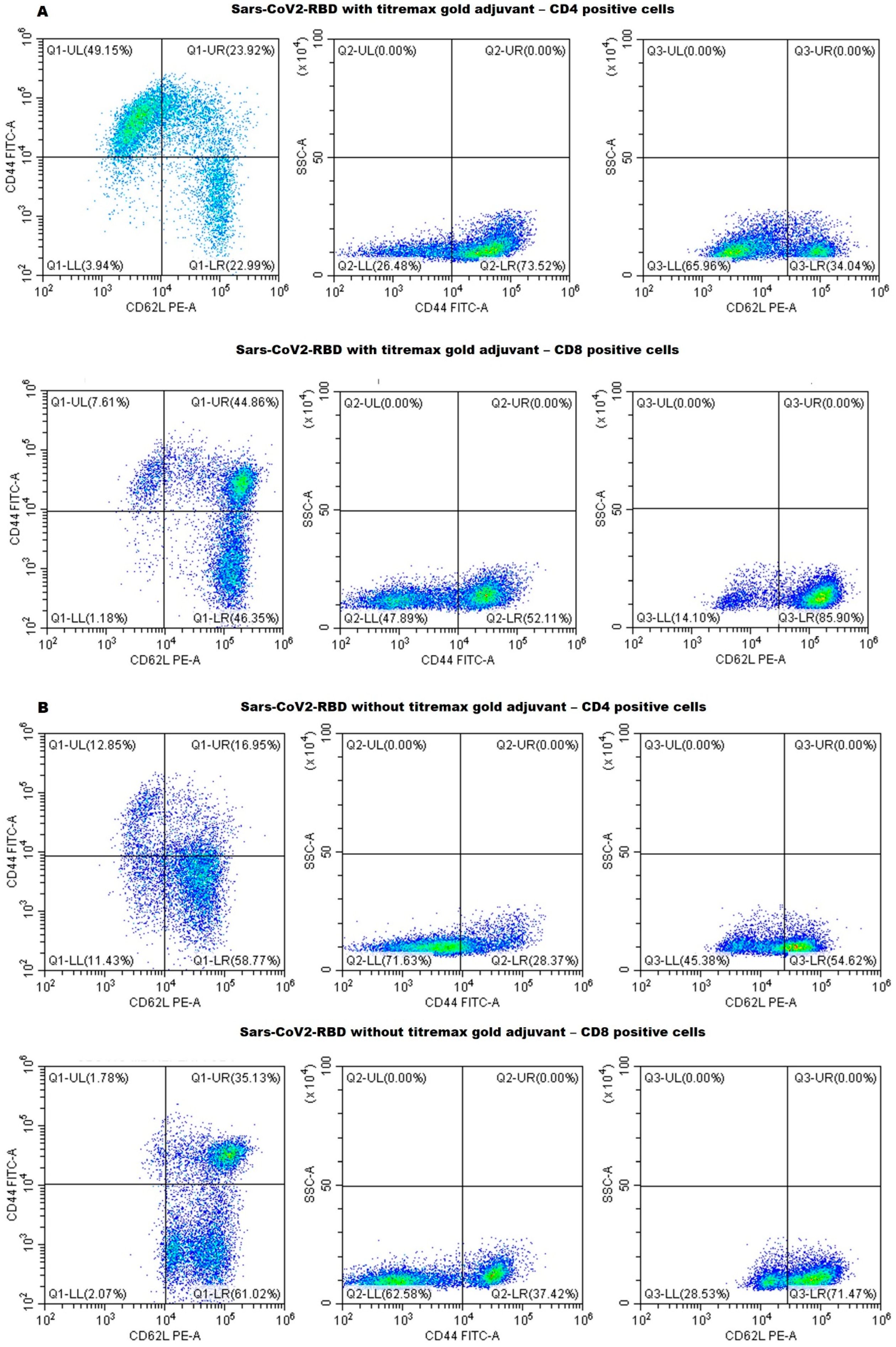
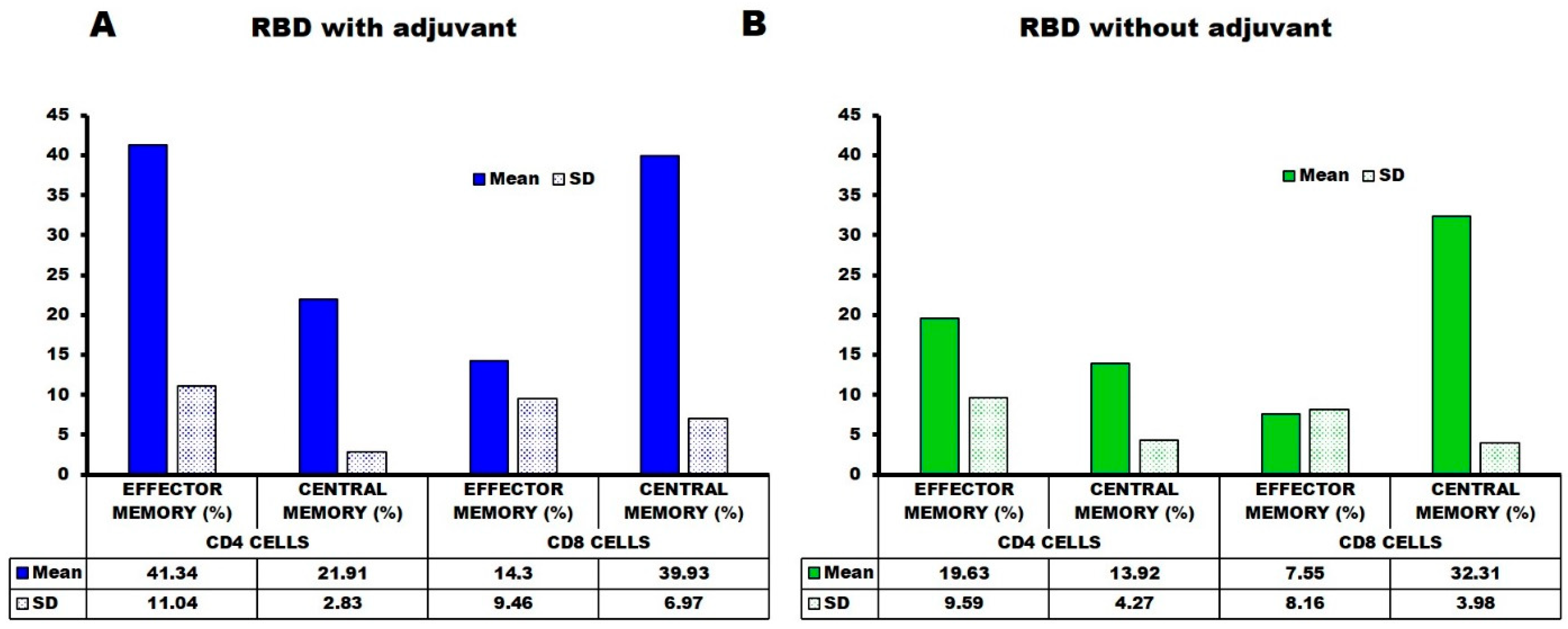
2.4. Flow Cytometry—Cell Surface Marker Analysis
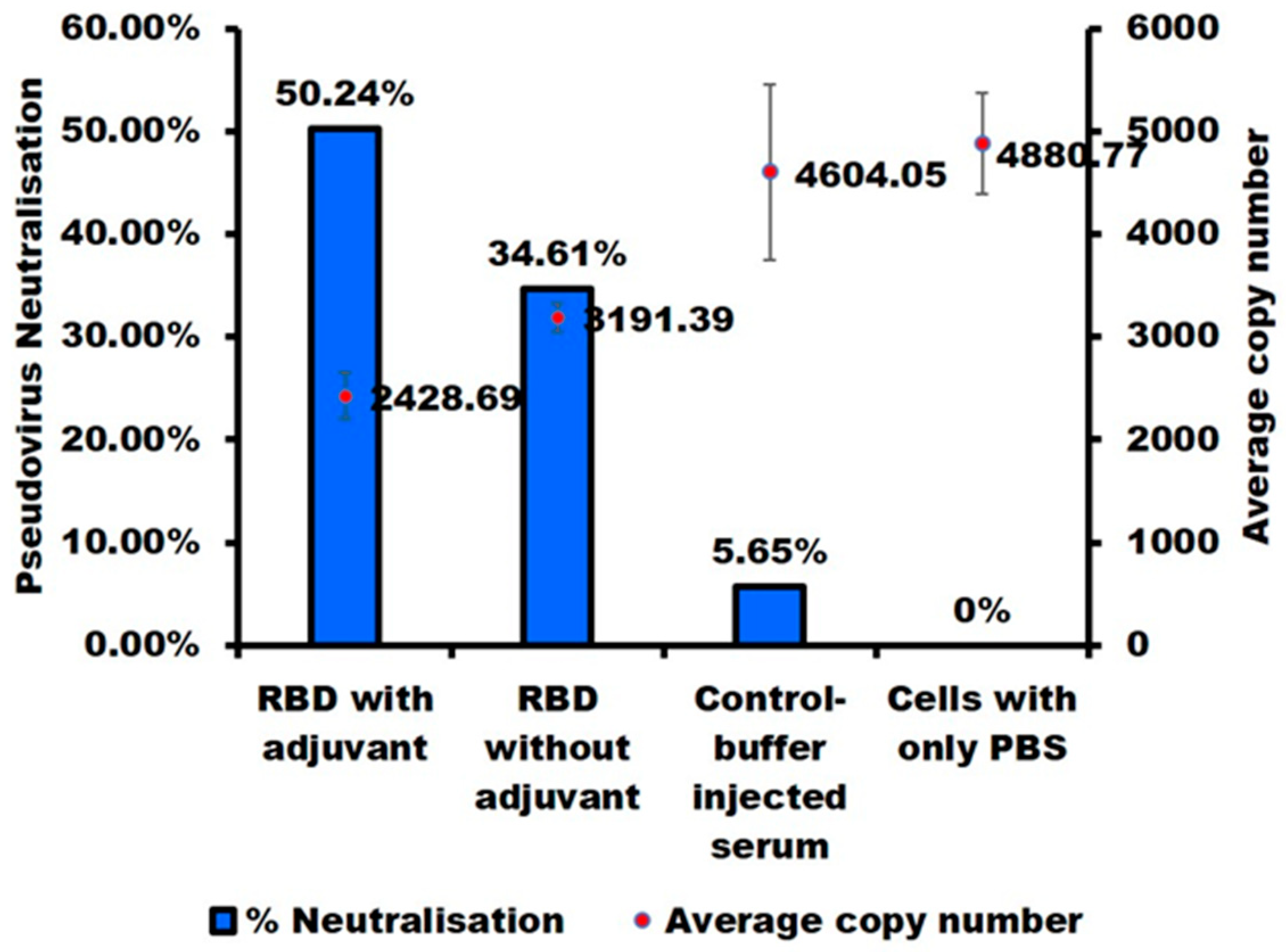
2.5. E. coli-Produced SARS-CoV-2-RBD Can Induce Specific and Neutralizing Antibodies
3. Materials and Methods
3.1. Protein Expression and Purification
3.2. Physicochemical Measurements
3.3. Mice Immunization
3.4. Cell Surface CD Marker Analysis
3.5. Pseudovirus-Based Neutralization Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahase, E. COVID-19: What do we know about the late stage vaccine candidates? Brit. Med. J. 2020, 371, 4576. [Google Scholar] [CrossRef]
- Zhang, N.; Jiang, S.; Du, L. Current advancements and potential strategies in the development of MERS-CoV vaccines. Expert Rev. Vaccines 2014, 13, 761–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Lateef, H.M.; El-Dabea, T.; Khalaf, M.M.; Abu-Dief, A.M. Development of Metal Complexes for Treatment of Coronaviruses. Int. J. Mol. Sci. 2022, 23, 6418. [Google Scholar] [CrossRef] [PubMed]
- Al-Radadi, N.S.; Abu-Dief, A.M. Silver nanoparticles (AgNPs) as a metal nano-therapy: Possible mechanisms of antiviral action against COVID-19. Inorg. Nano-Met. Chem. 2022, 22, 1–19. [Google Scholar] [CrossRef]
- Luo, F.; Liao, F.L.; Wang, H.; Tang, H.B.; Yang, Z.Q.; Hou, W. Evaluation of antibody-dependent enhancement of SARS-CoV infection in rhesus macaques immunized with an inactivated SARS-CoV vaccine. Virol. Sin. 2018, 33, 201–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzer, R.; Clark, J.J.; Vaysburd, M.; Chang, V.T.; Abecka, A.A.; Kiss, L.; Sharma, P.; Llamazares, A.G.; Kipar, A.; Hiscox, J.A.; et al. Single-dose immunisation with a multimerised SARS-CoV-2 receptor binding domain (RBD) induces an enhanced and protective response in mice. FEBS Lett. 2021, 595, 2323–2340. [Google Scholar] [CrossRef]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 55–170. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.J.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.; Yao, G.; Yihang, P.; Zhizhuang, J.Z. Structure analysis of the receptor binding of 2019-nCoV. Biochem. Biophys. Res. Commun. 2020, 525, 135–140. [Google Scholar]
- Wanbo, T.; Lei, H.; Xiujuan, Z.; Jing, P.; Denis, V.; Shibo, J.; Yusen, Z.; Lanying, D. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar]
- Jaimes, J.A.; André, N.M.; Chappie, J.S.; Millet, J.K.; Whittaker, G.R. Phylogenetic Analysis and Structural Modeling of SARS-CoV-2 Spike Protein Reveals an Evolutionary Distinct and Proteolytically Sensitive Activation Loop. J. Mol. Biol. 2020, 432, 3309–3325. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.O.; Lee, E.; Parks, R.; Martinez, D.R.; Li, D.; Chen, H.; Edwards, R.J.; Gobeil, S.; Barr, M.; Mansouri, K.; et al. Neutralizing antibody vaccine for pandemic and pre-emergent coronaviruses. Nature 2021, 594, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Arbeitman, C.R.; Auge, G.; Blaustein, M.; Bredeston, L.; Corapi, E.S.; Craig, P.O.; Cossio, L.A.; Dain, L.; D’Alessio, C.; Elias, F.; et al. Argentinian AntiCovid Consortium Structural and functional comparison of SARS-CoV-2-spike receptor binding domain produced in Pichia pastoris and mammalian cells. Sci. Rep. 2020, 10, 21779. [Google Scholar]
- Müller, D.; Bayer, K.; Mattanovich, D. Potentials and limitations of prokaryotic and eukaryotic expression systems for recombinant protein production—A comparative view. Microb. Cell Fact. 2006, 5, P61. [Google Scholar] [CrossRef]
- Proffitt, A. First HEV vaccine approved. Nat. Biotechnol. 2012, 30, 300. [Google Scholar] [CrossRef]
- Qiao, Y.L.; Wu, T.; Li, R.C.; Hu, Y.M.; Wei, L.H.; Li, C.G.; Chen, W.; Huang, S.; Zhao, F.; Li, M.; et al. Efficacy, safety, and immunogenicity of an Escherichia coli-produced bivalent human papillomavirus vaccine: An interim analysis of a randomized clinical trial. J. Natl. Cancer Inst. 2020, 112, 145–153. [Google Scholar] [CrossRef]
- Shirley, M.; Taha, M.K. MenB-FHbp meningococcal group B vaccine (Trumenba): A review in active immunization in individuals aged ≥ 10 years. Drugs 2018, 78, 257–268. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front Microbiol. 2014, 17, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantiwiwat, T.; Thaiprayoon, A.; Siriatcharanon, A.K.; Tachaapaikoon, C.; Plongthongkum, N.; Waraho-Zhmayev, D. Utilization of Receptor-Binding Domain of SARS-CoV-2 Spike Protein Expressed in Escherichia coli for the Development of Neutralizing Antibody Assay. Mol. Biotechnol. 2022, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Salem, R.; El-Kholy, A.A.; Waly, F.R.; Ayman, D.; Sakr, A.; Hussein, M. Generation and utility of a single-chain fragment variable monoclonal antibody platform against a baculovirus expressed recombinant receptor binding domain of SARS-CoV-2 spike protein. Mol. Immunol. 2021, 141, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Missiakas, D.; Georgopoulos, C.; Raina, S. The Escherichia coli dsbC (xprA) gene encodes a periplasmic protein involved in disulfide bond formation. EMBO J. 1994, 13, 2013–2020. [Google Scholar] [CrossRef]
- Lobstein, J.; Emrich, C.A.; Jeans, C.; Faulkner, M.; Riggs, P.; Berkmen, M. SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide bonded proteins in its cytoplasm. Microb. Cell Factories 2012, 11, 56. [Google Scholar] [CrossRef] [Green Version]
- Hartl, F.U.; Hartl, M.M. Converging concepts of protein folding in vitro and in vivo. Nat. Struct. Mol. Biol. 2009, 16, 574–581. [Google Scholar] [CrossRef]
- Chen, X.; Li, C.; Liu, H. Enhanced recombinant protein production under special environmental stress. Front. Microbiol. 2021, 12, 630814. [Google Scholar] [CrossRef]
- Veldkamp, C.T.; Koplinski, C.A.; Jensen, D.R.; Peterson, F.C.; Smits, K.M.; Smith, B.L.; Johnson, S.K.; Lettieri, C.; Buchholz, W.G.; Solheim, J.C.; et al. Production of Recombinant Chemokines and Validation of Refolding. Methods Enzymol. 2016, 570, 539–565. [Google Scholar]
- Brindha, S.; Kibria, M.G.; Saotome, T.; Unzai, S.; Kuroda, Y. EGFR extracellular domain III expressed in Escherichia coli with SEP tag shows improved biophysical and functional properties and generate anti-sera inhibiting cancer cell growth. Biochem. Biophys. Res. Commun. 2021, 555, 121–127. [Google Scholar] [CrossRef]
- Brindha, S.; Kuroda, Y. A Multi-Disulfide Receptor-Binding Domain (RBD) of the SARS-CoV-2 Spike Protein Expressed in E. coli Using a SEP-Tag Produces Antisera Interacting with the Mammalian Cell Expressed Spike (S1) Protein. Int. J. Mol. Sci. 2022, 23, 1703. [Google Scholar]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.S. Solute perturbation of protein fluorescence. Quenching of the tryptophyl fluorescence of model compounds and of lysozyme by iodide ion. Biochemistry 1971, 10, 3254–3263. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiaka, S.; Masuda, T.; Sugimura, S.; Kobayashi, S.; Komatsu, N.; Nagai, M.; Omatsu, T.; Furuya, T.; Oba, M.; Katayama, Y.; et al. Development of a novel detection system for microbes from bovine diarrhea by real-time PCR. J. Vet. Med. Sci. 2016, 78, 383–389. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brindha, S.; Yoshizue, T.; Wongnak, R.; Takemae, H.; Oba, M.; Mizutani, T.; Kuroda, Y. An Escherichia coli Expressed Multi-Disulfide Bonded SARS-CoV-2 RBD Shows Native-like Biophysical Properties and Elicits Neutralizing Antisera in a Mouse Model. Int. J. Mol. Sci. 2022, 23, 15744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415744
Brindha S, Yoshizue T, Wongnak R, Takemae H, Oba M, Mizutani T, Kuroda Y. An Escherichia coli Expressed Multi-Disulfide Bonded SARS-CoV-2 RBD Shows Native-like Biophysical Properties and Elicits Neutralizing Antisera in a Mouse Model. International Journal of Molecular Sciences. 2022; 23(24):15744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415744
Chicago/Turabian StyleBrindha, Subbaian, Takahiro Yoshizue, Rawiwan Wongnak, Hitoshi Takemae, Mami Oba, Tetsuya Mizutani, and Yutaka Kuroda. 2022. "An Escherichia coli Expressed Multi-Disulfide Bonded SARS-CoV-2 RBD Shows Native-like Biophysical Properties and Elicits Neutralizing Antisera in a Mouse Model" International Journal of Molecular Sciences 23, no. 24: 15744. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415744