
Apicobasal Surfaceome Architecture Encodes for Polarized Epithelial Functionality and Depends on Tumor Suppressor PTEN
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
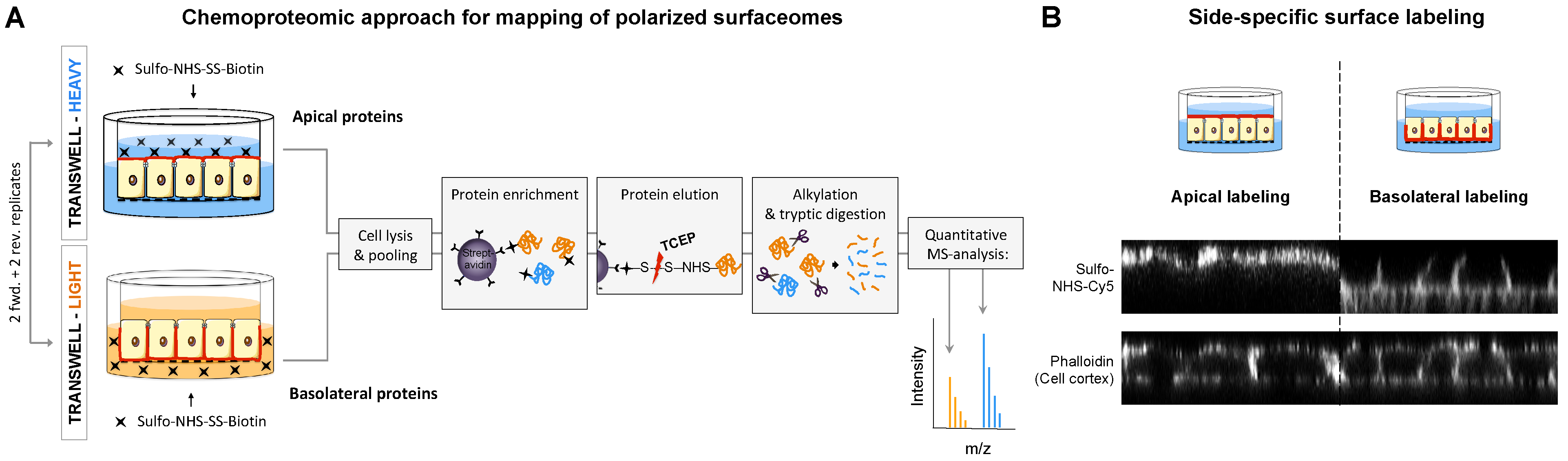
2.1. Chemoproteomic Approach for Mapping of Polarized Surfaceomes
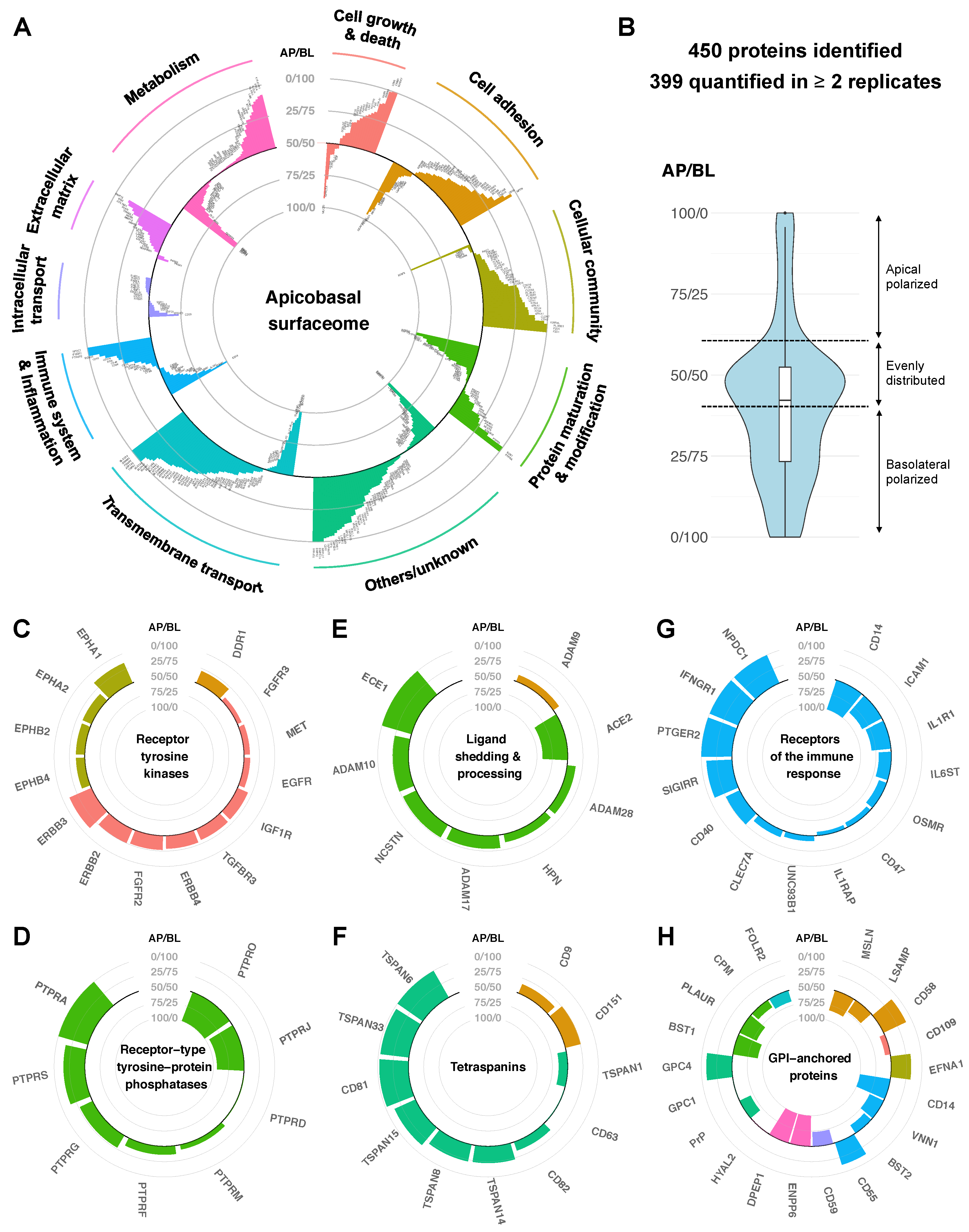
2.2. Quantitative Protein Distribution Is the Key Feature of the Apicobasal Surfaceome
2.2.1. Protein Abundances Indicate Functional Capacities of Apical versus Basolateral Membranes
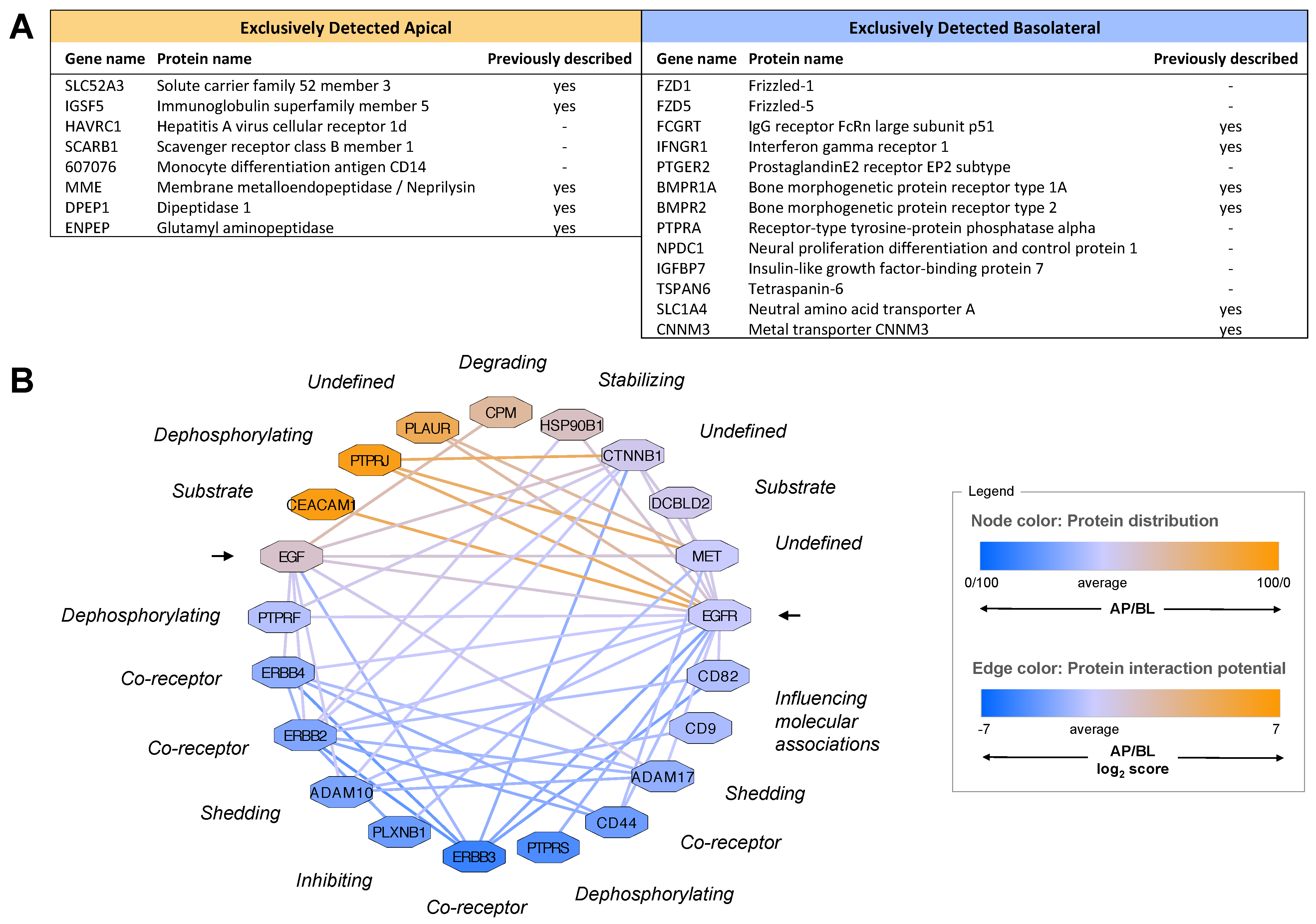
2.2.2. Polarized Distribution of Modulatory Proteins
2.2.3. Quantitative Protein Distributions Give Rise to Polarized Functional Networks
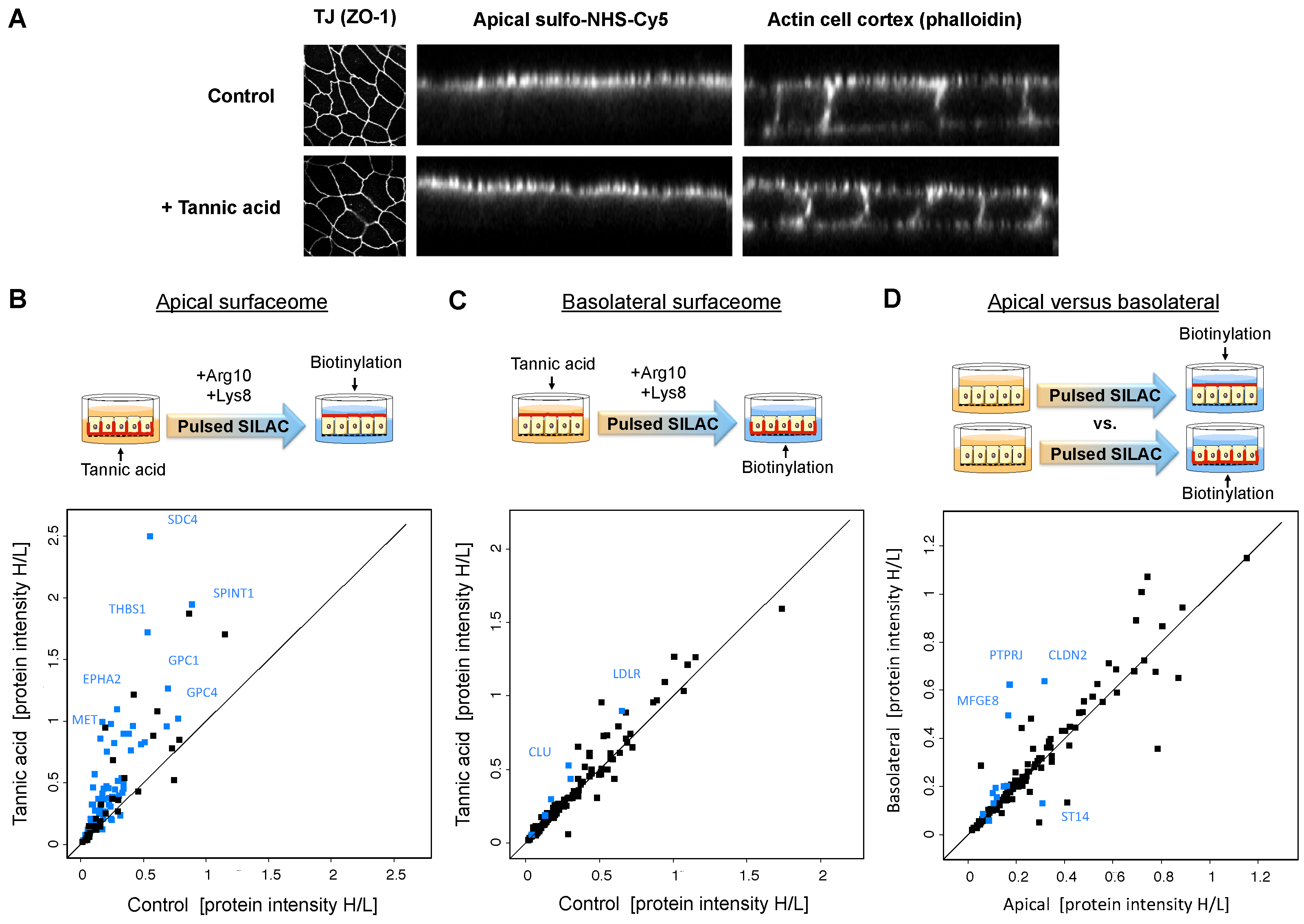
2.3. Tracking of Polarized Protein Trafficking Suggests Intertwining of Sorting Routes
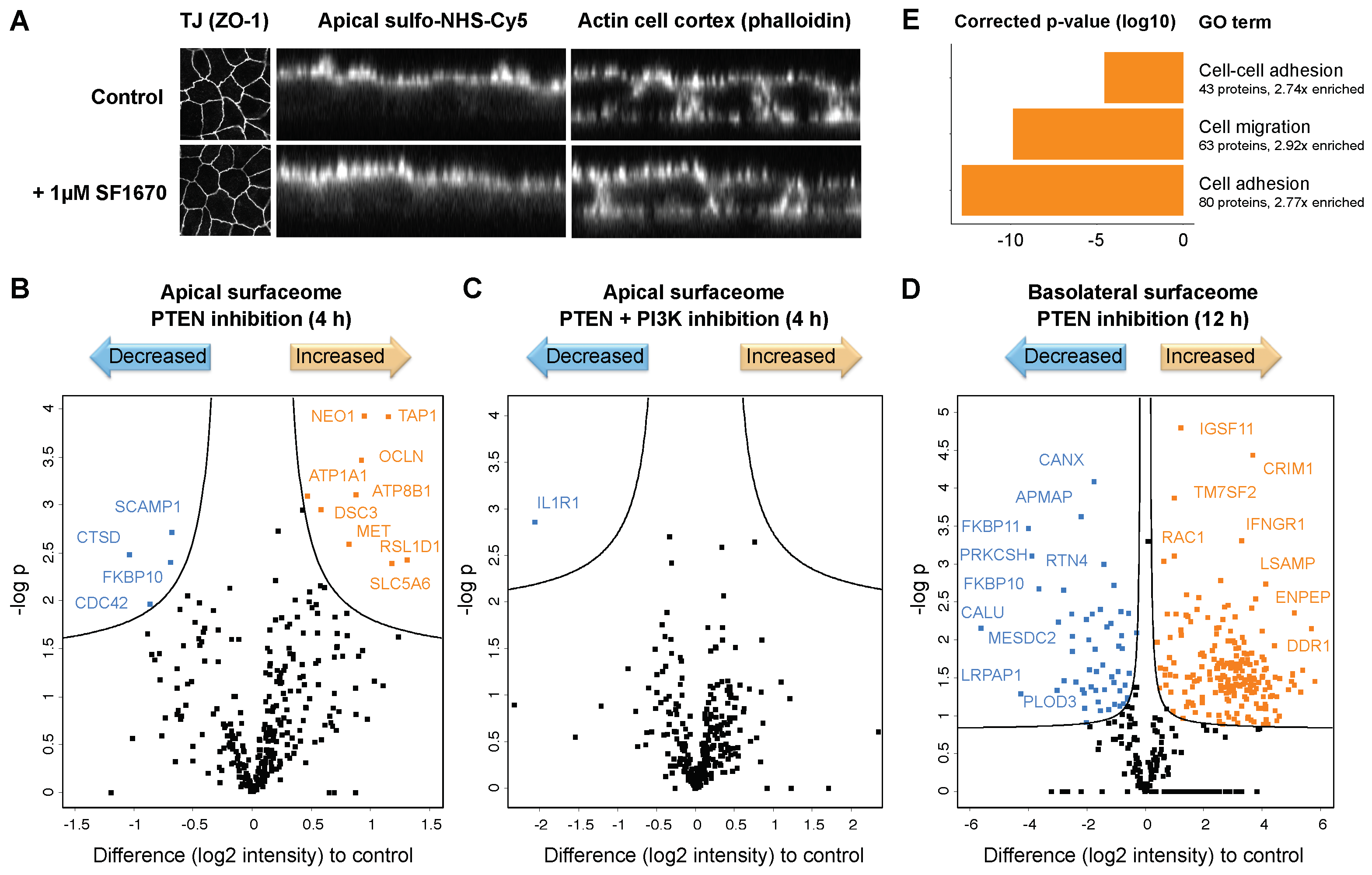
2.3.1. Impaired PTEN Function Causes Alterations in the Polarized Surfaceome
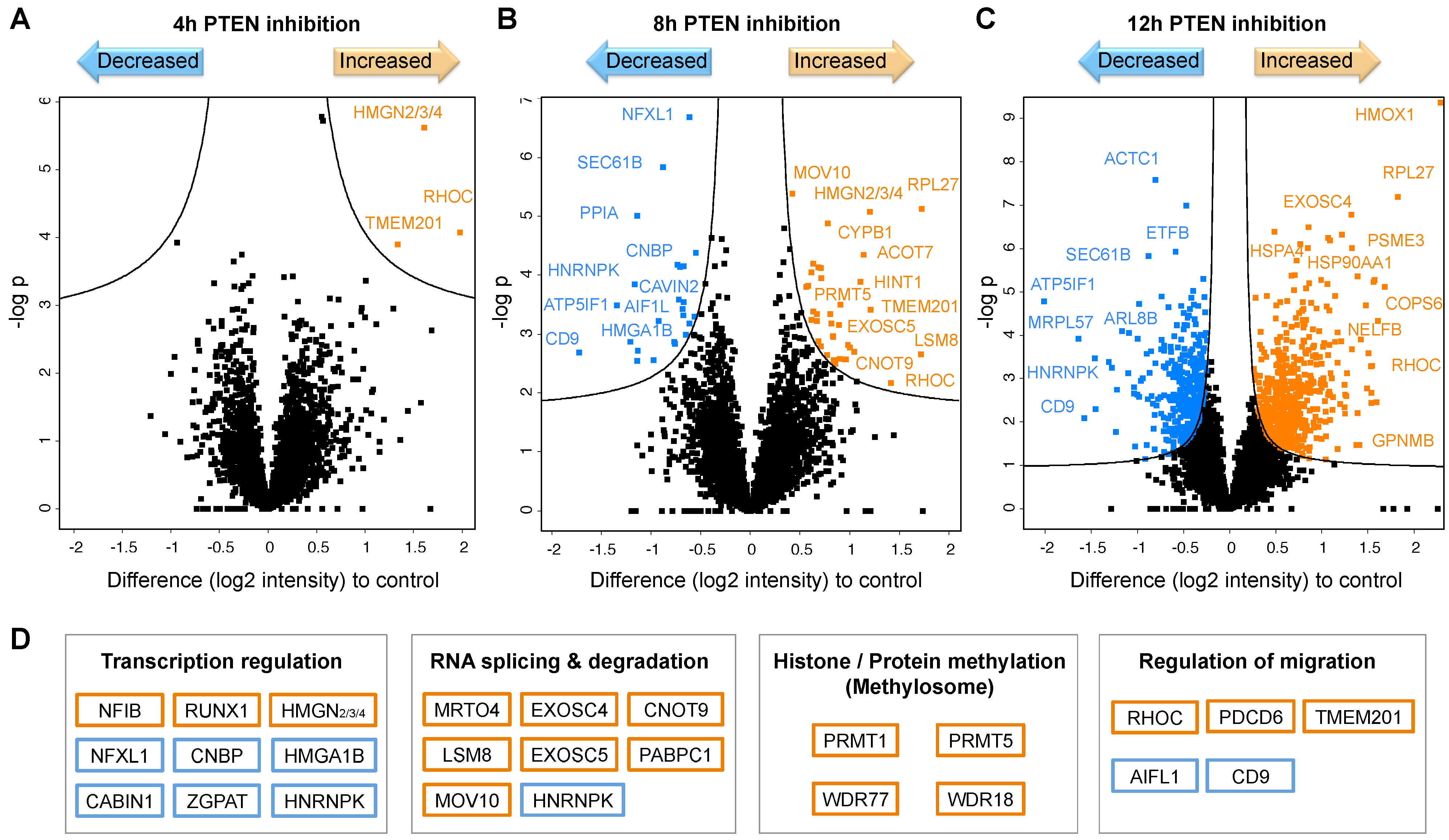
2.3.2. Impaired PTEN Function Causes Massive Proteotype Reorganization
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Lines
4.3. Mammalian Cell Culture, SILAC Labeling, and Polarized Culture
4.4. Confocal Microscopy Imaging
4.5. Tannic Acid Membrane Fixation and Pulsed SILAC
4.6. Inhibitor Treatments
4.7. Cell Surface Protein Enrichment
4.8. Sample Preparation for MS Analysis
4.9. LC-MS/MS Analysis
4.10. Data Analysis and Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amsler, K.; Kuwada, S.K. Membrane receptor location defines receptor interaction with signaling proteins in a polarized epithelium. Am. J. Physiol. Physiol. 1999, 276, C91–C101. [Google Scholar] [CrossRef] [PubMed]
- Kuwada, S.K.; Lund, K.A.; Li, X.F.; Cliften, P.; Amsler, K.; Opresko, L.K.; Wiley, H.S. Differential signaling and regulation of apical vs. basolateral EGFR in polarized epithelial cells. Am. J. Physiol. Physiol. 1998, 275, C1419–C1428. [Google Scholar] [CrossRef] [PubMed]
- Hobert, M.E.; Friend, L.A.; Carlin, C.R. Regulation of EGF signaling by cell polarity in MDCK kidney epithelial cells. J. Cell. Physiol. 1999, 181, 330–341. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Martín-Belmonte, F.; Rodríguez-Fraticelli, A.E. Chapter 3 Acquisition of Membrane Polarity in Epithelial Tube Formation: Patterns, Signaling Pathways, Molecular Mechanisms, and Disease. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2009; Volume 274, pp. 129–182. [Google Scholar]
- Muthuswamy, S.K.; Xue, B. Cell Polarity as a Regulator of Cancer Cell Behavior Plasticity. Annu. Rev. Cell Dev. Biol. 2012, 28, 599–625. [Google Scholar] [CrossRef] [Green Version]
- Polishchuk, R.; Di Pentima, A.; Lippincott-Schwartz, J. Delivery of raft-associated, GPI-anchored proteins to the apical surface of polarized MDCK cells by a transcytotic pathway. Nature 2004, 6, 297–307. [Google Scholar] [CrossRef]
- Loo, C.-S.; Chen, C.-W.; Wang, P.-J.; Chen, P.-Y.; Lin, S.-Y.; Khoo, K.-H.; Fenton, R.A.; Knepper, M.A.; Yu, M.-J. Quantitative apical membrane proteomics reveals vasopressin-induced actin dynamics in collecting duct cells. Proc. Natl. Acad. Sci. USA 2013, 110, 17119–17124. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Ohtsuki, S.; Katsukura, Y.; Ikeda, C.; Suzuki, T.; Kamiie, J.; Terasaki, T. Quantitative targeted absolute proteomics of human blood-brain barrier transporters and receptors. J. Neurochem. 2011, 117, 333–345. [Google Scholar] [CrossRef]
- Yu, M.-J.; Pisitkun, T.; Wang, G.; Shen, R.-F.; Knepper, M.A. LC-MS/MS Analysis of Apical and Basolateral Plasma Membranes of Rat Renal Collecting Duct Cells. Mol. Cell. Proteom. 2006, 5, 2131–2145. [Google Scholar] [CrossRef] [Green Version]
- Hartwell, L.H.; Hopfield, J.J.; Leibler, S.; Murray, A.W. From molecular to modular cell biology. Nature 1999, 402, C47–C52. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef]
- Bausch-Fluck, D.; Milani, E.S.; Wollscheid, B. Surfaceome nanoscale organization and extracellular interaction networks. Curr. Opin. Chem. Biol. 2018, 48, 26–33. [Google Scholar] [CrossRef]
- Elschenbroich, S.; Kim, Y.; Medin, J.A.; Kislinger, T. Isolation of cell surface proteins for mass spectrometry-based proteomics. Expert Rev. Proteom. 2010, 7, 141–154. [Google Scholar] [CrossRef]
- Wollscheid, B.; Bausch-Fluck, D.; Henderson, C.; O’Brien, R.; Bibel, M.; Schiess, R.; Aebersold, R.; Watts, J.D. Mass-spectrometric identification and relative quantification of N-linked cell surface glycoproteins. Nat. Biotechnol. 2009, 27, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Boulan, E.; Kreitzer, G.; Müsch, A. Organization of vesicular trafficking in epithelia. Nat. Rev. Mol. Cell Biol. 2005, 6, 233–247. [Google Scholar] [CrossRef]
- Ong, S.-E.; Blagoev, B.; Kratchmarova, I.; Kristensen, D.B.; Steen, H.; Pandey, A.; Mann, M. Stable Isotope Labeling by Amino Acids in Cell Culture, SILAC, as a Simple and Accurate Approach to Expression Proteomics. Mol. Cell. Proteom. 2002, 1, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Mann, M. Fifteen Years of Stable Isotope Labeling by Amino Acids in Cell Culture (SILAC). In Stable Isotope Labeling by Amino Acids in Cell Culture (SILAC): Methods and Protocols; Warscheid, B., Ed.; Springer: New York, NY, USA, 2014; pp. 1–7. ISBN 9781493911424. [Google Scholar]
- Bausch-Fluck, D.; Goldmann, U.; Müller, S.; van Oostrum, M.; Müller, M.; Schubert, O.T.; Wollscheid, B. The in silico human surfaceome. Proc. Natl. Acad. Sci. USA 2018, 115, E10988–E10997. [Google Scholar] [CrossRef] [Green Version]
- Stoops, E.H.; Caplan, M.J. Trafficking to the Apical and Basolateral Membranes in Polarized Epithelial Cells. J. Am. Soc. Nephrol. 2014, 25, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Boulan, E.; Macara, I.G. Organization and execution of the epithelial polarity programme. Nat. Rev. Mol. Cell Biol. 2014, 15, 225–242. [Google Scholar] [CrossRef]
- Lisanti, M.P.; Caras, I.W.; Davitz, M.A.; Rodriguez-Boulan, E. A glycophospholipid membrane anchor acts as an apical targeting signal in polarized epithelial cells. J. Cell Biol. 1989, 109, 2145–2156. [Google Scholar] [CrossRef]
- Cortes, L.K.; Vainauskas, S.; Dai, N.; McClung, C.M.; Shah, M.; Benner, J.S.; Corrêa, I.R.; VerBerkmoes, N.C.; Taron, C.H. Proteomic identification of mammalian cell surface derived glycosylphosphatidylinositol-anchored proteins through selective glycan enrichment. PROTEOMICS 2014, 14, 2471–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeusen, B.; Janssens, V. Tumor suppressive protein phosphatases in human cancer: Emerging targets for therapeutic intervention and tumor stratification. Int. J. Biochem. Cell Biol. 2018, 96, 98–134. [Google Scholar] [CrossRef] [PubMed]
- Stanoev, A.; Mhamane, A.; Schuermann, K.C.; Grecco, H.E.; Stallaert, W.; Baumdick, M.; Brüggemann, Y.; Joshi, M.S.; Roda-Navarro, P.; Fengler, S.; et al. Interdependence between EGFR and Phosphatases Spatially Established by Vesicular Dynamics Generates a Growth Factor Sensing and Responding Network. Cell Syst. 2018, 7, 295–309.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Termini, C.M.; Gillette, J.M. Tetraspanins Function as Regulators of Cellular Signaling. Front. Cell Dev. Biol. 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Pankow, S.; Martínez-Bartolomé, S.; Bamberger, C.; Yates, J.R. Understanding molecular mechanisms of disease through spatial proteomics. Curr. Opin. Chem. Biol. 2018, 48, 19–25. [Google Scholar] [CrossRef]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Hu, Q.; Li, C.; Wang, S.; Li, Y.; Wen, B.; Zhang, Y.; Liang, K.; Yao, J.; Ye, Y.; Hsiao, H.; et al. LncRNAs-directed PTEN enzymatic switch governs epithelial–mesenchymal transition. Cell Res. 2019, 29, 286–304. [Google Scholar] [CrossRef]
- Gassama-Diagne, A.; Yu, W.; ter Beest, M.; Martin-Belmonte, F.; Kierbel, A.; Engel, J.; Mostov, K. Phosphatidylinositol-3,4,5-trisphosphate regulates the formation of the basolateral plasma membrane in epithelial cells. Nature 2006, 8, 963–970. [Google Scholar] [CrossRef]
- Martin-Belmonte, F.; Gassama, A.; Datta, A.; Yu, W.; Rescher, U.; Gerke, V.; Mostov, K. PTEN-Mediated Apical Segregation of Phosphoinositides Controls Epithelial Morphogenesis through Cdc42. Cell 2007, 128, 383–397. [Google Scholar] [CrossRef]
- Chen, H.; Lorton, B.; Gupta, V.; Shechter, D. A TGFβ-PRMT5-MEP50 axis regulates cancer cell invasion through histone H3 and H4 arginine methylation coupled transcriptional activation and repression. Oncogene 2016, 36, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Avasarala, S.; Van Scoyk, M.; Rathinam, M.K.K.; Zerayesus, S.; Zhao, X.; Zhang, W.; Pergande, M.R.; Borgia, J.A.; DeGregori, J.; Port, J.D.; et al. PRMT1 Is a Novel Regulator of Epithelial-Mesenchymal-Transition in Non-small Cell Lung Cancer. J. Biol. Chem. 2015, 290, 13479–13489. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Bedford, M.T. Protein arginine methyltransferases and cancer. Nat. Rev. Cancer 2013, 13, 37–50. [Google Scholar] [CrossRef]
- Caceres, P.S.; Gravotta, D.; Zager, P.J.; Dephoure, N.; Rodriguez-Boulan, E. Quantitative proteomics of MDCK cells identify unrecognized roles of clathrin adaptor AP-1 in polarized distribution of surface proteins. Proc. Natl. Acad. Sci. USA 2019, 116, 11796–11805. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-W.; Rees, J.S.; Xue, P.; Zhang, H.; Hamaia, S.W.; Sanderson, B.; Funk, P.E.; Farndale, R.W.; Lilley, K.S.; Perrett, S.; et al. New Insights into the DT40 B Cell Receptor Cluster Using a Proteomic Proximity Labeling Assay. J. Biol. Chem. 2014, 289, 14434–14447. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.; Fan, L.; Alarcón, B. Multiple Sorting Signals Determine Apical Localization of a Nonglycosylated Integral Membrane Protein. J. Biol. Chem. 1997, 272, 30748–30752. [Google Scholar] [CrossRef] [Green Version]
- Youker, R.T.; Bruns, J.R.; Costa, S.A.; Rbaibi, Y.; Lanni, F.; Kashlan, O.B.; Teng, H.; Weisz, O.A. Multiple motifs regulate apical sorting of p75 via a mechanism that involves dimerization and higher-order oligomerization. Mol. Biol. Cell 2013, 24, 1996–2007. [Google Scholar] [CrossRef]
- Mayor, R.; Etienne-Manneville, S. The front and rear of collective cell migration. Nat. Rev. Mol. Cell Biol. 2016, 17, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Lacalle, R.A.; Gómez-Moutón, C.; Barber, D.F.; Jiménez-Baranda, S.; Mira, E.; Martínez-A, C.; Carrera, A.C.; Mañes, S. PTEN regulates motility but not directionality during leukocyte chemotaxis. J. Cell Sci. 2004, 117, 6207–6215. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Graue-Hernandez, E.O.; Tran, V.; Reid, B.; Pu, J.; Mannis, M.J.; Zhao, M. Downregulation of PTEN at Corneal Wound Sites Accelerates Wound Healing through Increased Cell Migration. Investig. Opthalmol. Vis. Sci. 2011, 52, 2272–2278. [Google Scholar] [CrossRef]
- Squarize, C.; Castilho, R.M.; Bugge, T.H.; Gutkind, J.S. Accelerated Wound Healing by mTOR Activation in Genetically Defined Mouse Models. PLoS ONE 2010, 5, e10643. [Google Scholar] [CrossRef] [PubMed]
- Labernadie, A.; Kato, T.; Brugués, A.; Serra-Picamal, X.; Derzsi, S.; Arwert, E.; Weston, A.; González-Tarragó, V.; Elosegui-Artola, A.; Albertazzi, L.; et al. A mechanically active heterotypic E-cadherin/N-cadherin adhesion enables fibroblasts to drive cancer cell invasion. Nature 2017, 19, 224–237. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koetemann, A.; Wollscheid, B. Apicobasal Surfaceome Architecture Encodes for Polarized Epithelial Functionality and Depends on Tumor Suppressor PTEN. Int. J. Mol. Sci. 2022, 23, 16193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232416193
Koetemann A, Wollscheid B. Apicobasal Surfaceome Architecture Encodes for Polarized Epithelial Functionality and Depends on Tumor Suppressor PTEN. International Journal of Molecular Sciences. 2022; 23(24):16193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232416193
Chicago/Turabian StyleKoetemann, Anika, and Bernd Wollscheid. 2022. "Apicobasal Surfaceome Architecture Encodes for Polarized Epithelial Functionality and Depends on Tumor Suppressor PTEN" International Journal of Molecular Sciences 23, no. 24: 16193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232416193