
Circulating Extracellular Vesicles Impair Mesenchymal Stromal Cell Differentiation Favoring Adipogenic Rather than Osteogenic Differentiation in Adolescents with Obesity
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
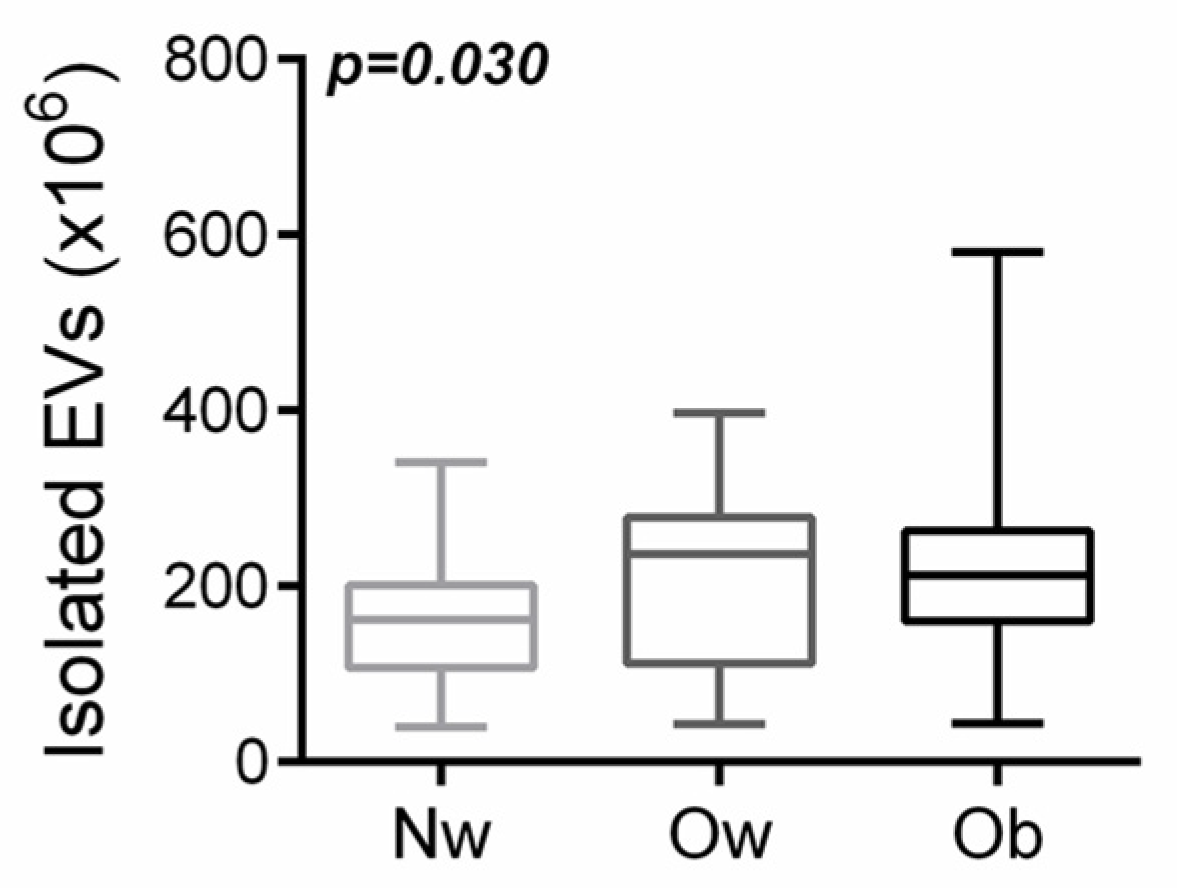
2.1. Participants with Overweight/Obesity Had a Higher Amount of Circulating Evs Than Normal-Weight Peers
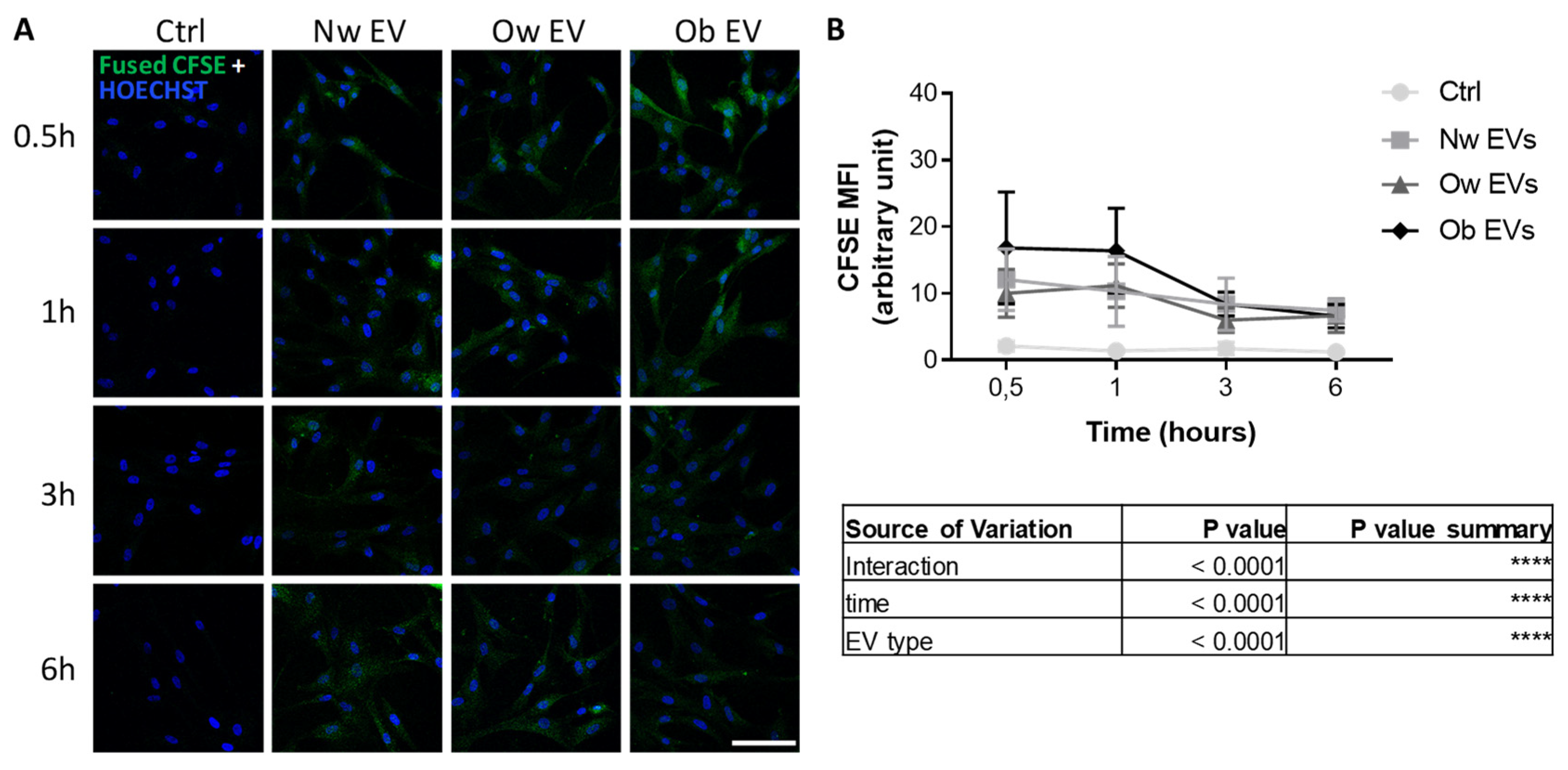
2.2. Uptake of Circulating EVs by Mesenchymal Stromal Cells
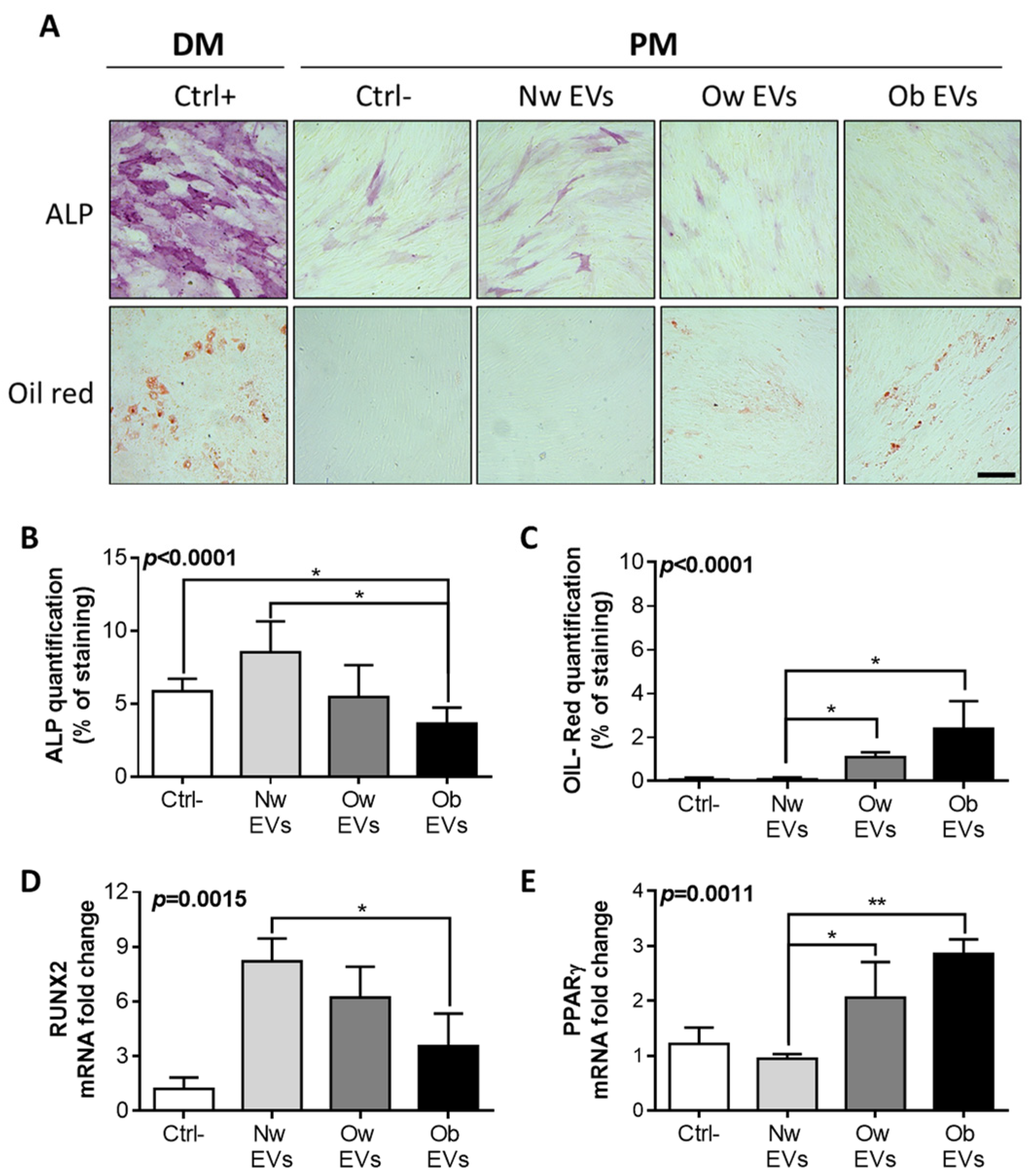
2.3. Treatment of EV from Obese Patients on Mesenchymal Stromal Cells Affects Both Osteogenic and Adipogenic Differentiation
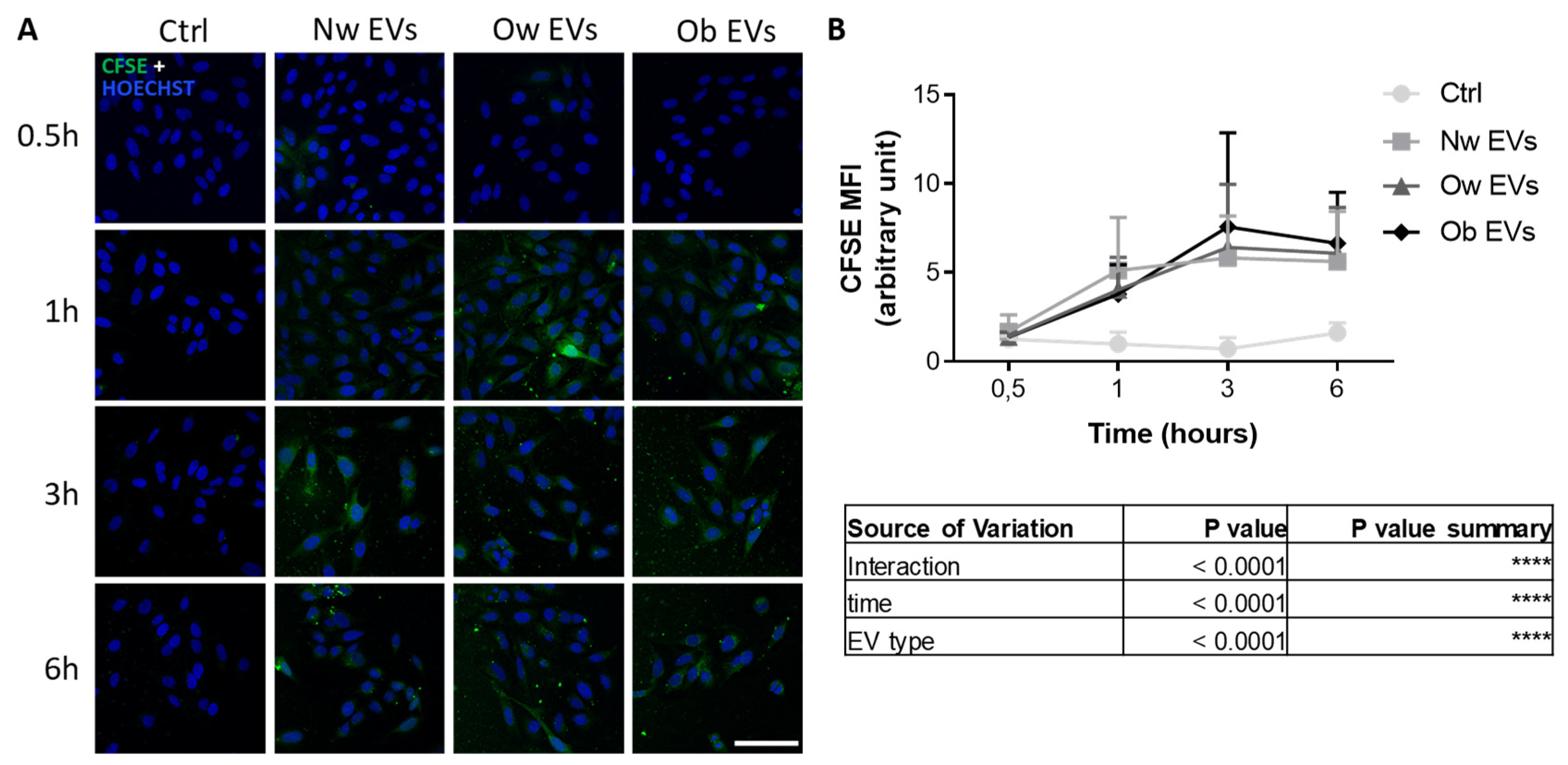
2.4. Uptake of Circulating EVs by Mature Osteoblasts
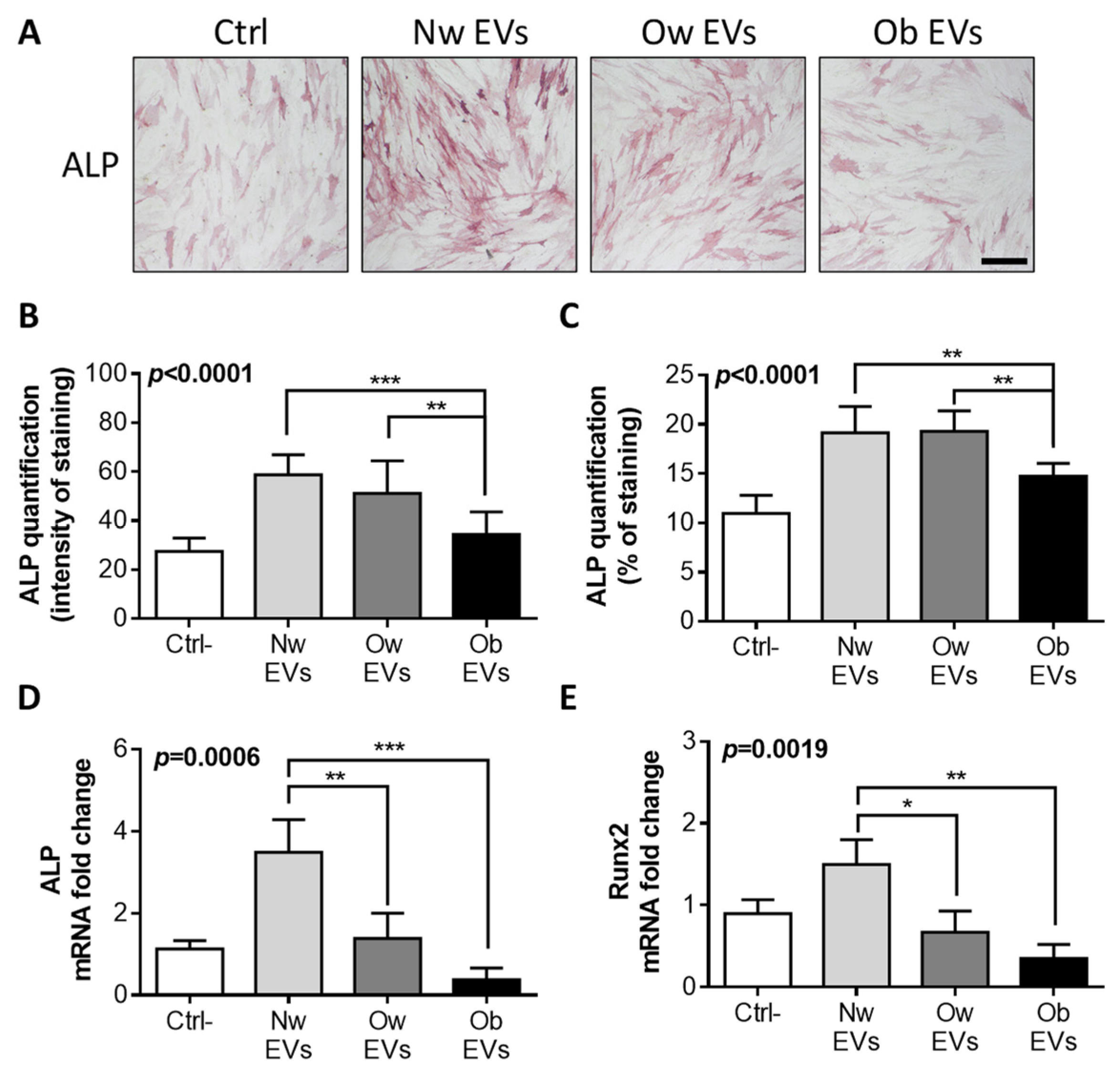
2.5. Obese EV Treatment Impairs Osteoblast Differentiation
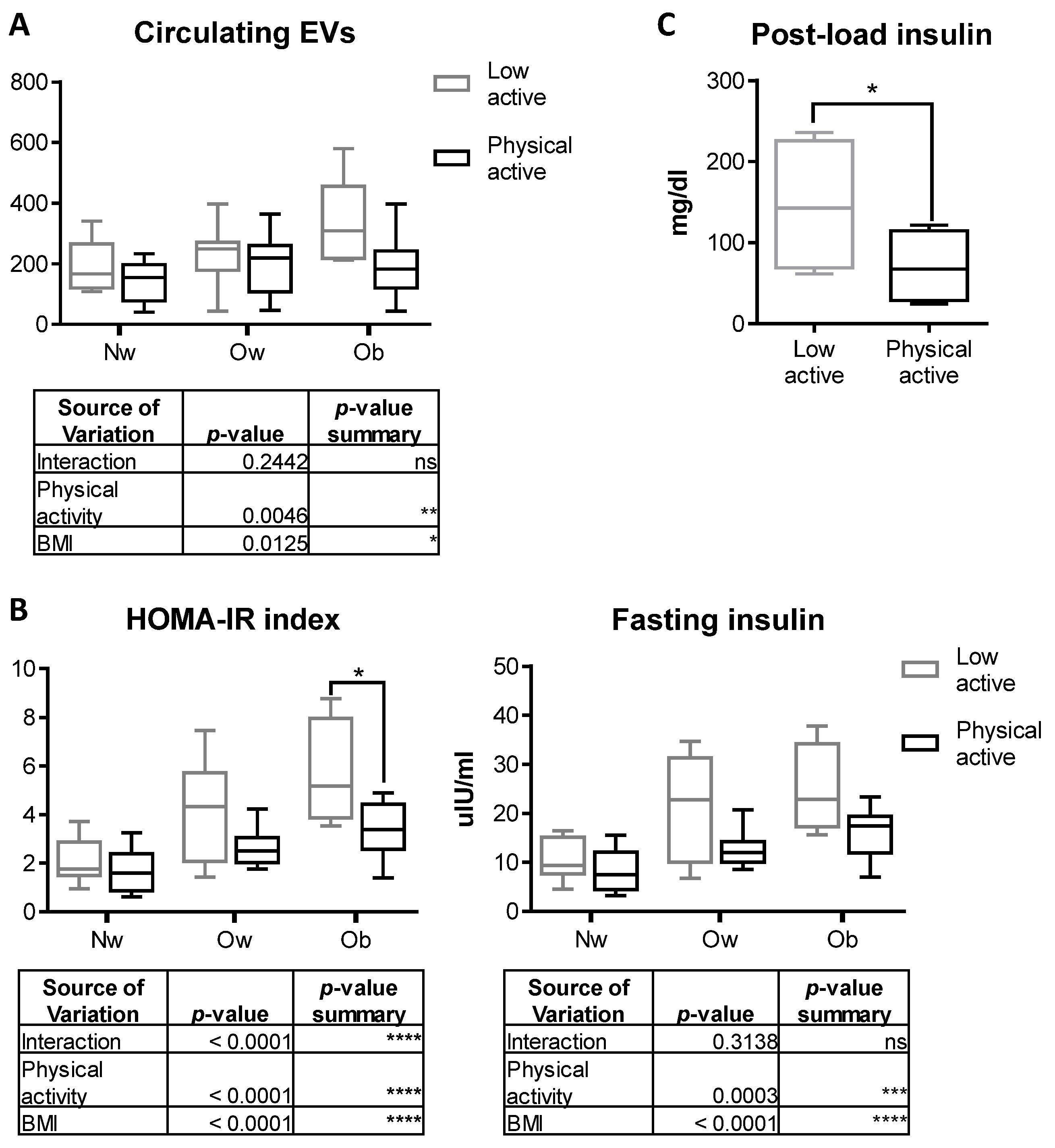
2.6. Physical Activity Counteracts the Increase in Circulating EVs in Obese Children
3. Discussion
4. Materials and Methods
4.1. Study Sample
4.2. Lifestyle Habits and Physical Activity
4.3. Anthropometric Measurements and Biochemical Assays
4.4. Isolation and Staining of Extracellular Vesicles from Patients’ Plasma
4.5. Flow Cytometry Analysis
4.6. Cell Cultures
4.7. EV Uptake
4.8. Confocal Microscopy Analysis
4.9. MSCs Treatment
4.10. HOBs Treatment
4.11. RT-qPCR
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jebeile, H.; Kelly, A.S.; O’Malley, G.; Baur, L.A. Obesity in children and adolescents: Epidemiology, causes, assessment, and management. Lancet Diabetes Endocrinol. 2022, 10, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellar, E.R.; Hill, C.; Melling, G.E.; Carter, D.R.; Baena-Lopez, L.A. Unpacking extracellular vesicles: RNA cargo loading and function. J. Extracell. Biol. 2022, 1, e40. [Google Scholar] [CrossRef]
- Anand, S.; Samuel, M.; Kumar, S.; Mathivanan, S. Ticket to a bubble ride: Cargo sorting into exosomes and extracellular vesicles. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140203. [Google Scholar] [CrossRef]
- Tetta, C.; Ghigo, E.; Silengo, L.; Deregibus, M.C.; Camussi, G. Extracellular vesicles as an emerging mechanism of cell-to-cell communication. Endocrine 2013, 44, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Ipinmoroti, A.O.; Matthews, Q.L. Extracellular Vesicles: Roles in Human Viral Infections, Immune-Diagnostic, and Therapeutic Applications. Pathogens 2020, 9, 1056. [Google Scholar] [CrossRef]
- Urciuoli, E.; Peruzzi, B. Inhibiting Extracellular Vesicle Trafficking as Antiviral Approach to Corona Virus Disease 2019 Infection. Front. Pharmacol. 2020, 11, 580505. [Google Scholar] [CrossRef]
- Hill, A.F. Extracellular Vesicles and Neurodegenerative Diseases. J. Neurosci. 2019, 39, 9269–9273. [Google Scholar] [CrossRef]
- Jansen, F.; Nickenig, G.; Werner, N. Extracellular Vesicles in Cardiovascular Disease. Circ. Res. 2017, 120, 1649–1657. [Google Scholar] [CrossRef]
- Chang, W.-H.; Cerione, R.A.; Antonyak, M.A. Extracellular Vesicles and Their Roles in Cancer Progression. In Cancer Cell Signaling; Methods in Molecular Biology; Robles-Flores, M., Ed.; Humana: New York, NY, USA, 2021; Volume 2174, pp. 143–170. [Google Scholar] [CrossRef]
- van Niel, G.; Carter, D.R.F.; Clayton, A.; Lambert, D.W.; Raposo, G.; Vader, P. Challenges and directions in studying cell–cell communication by extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2022, 23, 369–382. [Google Scholar] [CrossRef]
- Pardo, F.; Villalobos-Labra, R.; Sobrevia, B.; Toledo, F.; Sobrevia, L. Extracellular vesicles in obesity and diabetes mellitus. Mol. Asp. Med. 2018, 60, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kiran, S.; Kumar, S.; Singh, U.P. Extracellular vesicles in obesity and its associated inflammation. Int. Rev. Immunol. 2022, 41, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Bond, S.T.; Calkin, A.C.; Drew, B.G. Adipose-Derived Extracellular Vesicles: Systemic Messengers and Metabolic Regulators in Health and Disease. Front. Physiol. 2022, 13, 837001. [Google Scholar] [CrossRef] [PubMed]
- Iwaniec, U.T.; Turner, R.T. Influence of body weight on bone mass, architecture and turnover. J. Endocrinol. 2016, 230, R115–R130. [Google Scholar] [CrossRef] [PubMed]
- Shapses, S.A.; Sukumar, D. Bone Metabolism in Obesity and Weight Loss. Annu. Rev. Nutr. 2012, 32, 287–309. [Google Scholar] [CrossRef] [Green Version]
- Fintini, D.; Cianfarani, S.; Cofini, M.; Andreoletti, A.; Ubertini, G.M.; Cappa, M.; Manco, M. The Bones of Children with Obesity. Front. Endocrinol. 2020, 11, 200. [Google Scholar] [CrossRef]
- da Silva, S.V.; Renovato-Martins, M.; Ribeiro-Pereira, C.; Citelli, M.; Barja-Fidalgo, C. Obesity modifies bone marrow microenvironment and directs bone marrow mesenchymal cells to adipogenesis. Obesity 2016, 24, 2522–2532. [Google Scholar] [CrossRef]
- Tencerova, M.; Figeac, F.; Ditzel, N.; Taipaleenmäki, H.; Nielsen, T.K.; Kassem, M. High-Fat Diet-Induced Obesity Promotes Expansion of Bone Marrow Adipose Tissue and Impairs Skeletal Stem Cell Functions in Mice. J. Bone Miner. Res. 2018, 33, 1154–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Peralta, S.; Romero-Velarde, E.; Vásquez-Garibay, E.M.; González-Hita, M.; Robles-Robles, L.C.; Ruiz-González, F.J.; Pérez-Romero, M.A. Bone mineral density and body composition in normal weight, overweight and obese children. BMC Pediatr. 2022, 22, 249. [Google Scholar] [CrossRef]
- Frühbeis, C.; Helmig, S.; Tug, S.; Simon, P.; Krämer-Albers, E.-M. Physical exercise induces rapid release of small extracellular vesicles into the circulation. J. Extracell. Vesicles 2015, 4, 28239. [Google Scholar] [CrossRef] [Green Version]
- Mera, P.; Laue, K.; Ferron, M.; Confavreux, C.; Wei, J.; Galán-Díez, M.; Lacampagne, A.; Mitchell, S.J.; Mattison, J.A.; Chen, Y.; et al. Osteocalcin Signaling in Myofibers Is Necessary and Sufficient for Optimum Adaptation to Exercise. Cell Metab. 2016, 23, 1078–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chahla, S.E.; Frohnert, B.I.; Thomas, W.; Kelly, A.S.; Nathan, B.M.; Polgreen, L.E. Higher daily physical activity is associated with higher osteocalcin levels in adolescents. Prev. Med. Rep. 2015, 2, 568–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Ferron, M.; Clarke, C.J.; Hannun, Y.A.; Jiang, H.; Blaner, W.S.; Karsenty, G. Bone-specific insulin resistance disrupts whole-body glucose homeostasis via decreased osteocalcin activation. J. Clin. Investig. 2014, 124, 1781–1793. [Google Scholar] [CrossRef] [Green Version]
- Mizokami, A.; Kawakubo-Yasukochi, T.; Hirata, M. Osteocalcin and its endocrine functions. Biochem. Pharmacol. 2017, 132, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, I. Osteocalcin as a hormone regulating glucose metabolism. World J. Diabetes 2015, 6, 1345. [Google Scholar] [CrossRef]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Eguchi, A.; Tempaku, M.; Honda, T.; Togashi, K.; Iwasa, M.; Hasegawa, H.; Takei, Y.; Sumida, Y.; Taguchi, O. Circulating extracellular vesicles are associated with lipid and insulin metabolism. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E574–E582. [Google Scholar] [CrossRef]
- Dimitri, P. The Impact of Childhood Obesity on Skeletal Health and Development. J. Obes. Metab. Syndr. 2019, 28, 4–17. [Google Scholar] [CrossRef]
- He, J.; Ren, W.; Wang, W.; Han, W.; Jiang, L.; Zhang, D.; Guo, M. Exosomal targeting and its potential clinical application. Drug Deliv. Transl. Res. 2022, 12, 2385–2402. [Google Scholar] [CrossRef]
- Sancho-Albero, M.; Navascués, N.; Mendoza, G.; Sebastián, V.; Arruebo, M.; Martín-Duque, P.; Santamaría, J. Exosome origin determines cell targeting and the transfer of therapeutic nanoparticles towards target cells. J. Nanobiotechnol. 2019, 17, 16. [Google Scholar] [CrossRef] [Green Version]
- Lai, R.C.; Yeo, R.W.Y.; Tan, K.H.; Lim, S.K. Exosomes for drug delivery—A novel application for the mesenchymal stem cell. Biotechnol. Adv. 2013, 31, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Faienza, M.F.; D’Amato, G.; Chiarito, M.; Colaianni, G.; Colucci, S.; Grano, M.; Corbo, F.; Brunetti, G. Mechanisms Involved in Childhood Obesity-Related Bone Fragility. Front. Endocrinol. 2019, 10, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanan, R.; Huang, C.-C.; Ravindran, S. Hijacking the Cellular Mail: Exosome Mediated Differentiation of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 3808674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Li, X.; Wang, F.; Wang, S.; Wang, X.; Tian, X.; Bai, S.; Miao, D.; Fan, J. Emerging Potential of Exosomes on Adipogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 1614. [Google Scholar] [CrossRef] [PubMed]
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.E.; Egan, C.L.; Cron, L.; Watt, K.I.; Kuchel, R.P.; et al. Extracellular Vesicles Provide a Means for Tissue Crosstalk during Exercise. Cell Metab. 2018, 27, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, I.R.; Palazzo, R.P.; Cechinel, L.R. Circulating extracellular vesicles delivering beneficial cargo as key players in exercise effects. Free. Radic. Biol. Med. 2021, 172, 273–285. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Bollati, V.; Pergoli, L.; Iodice, S.; De Col, A.; Tamini, S.; Cicolini, S.; Tringali, G.; De Micheli, R.; Cella, S.G.; et al. Effects of an acute bout of exercise on circulating extracellular vesicles: Tissue-, sex-, and BMI-related differences. Int. J. Obes. 2020, 44, 1108–1118. [Google Scholar] [CrossRef]
- McIlvenna, L.C.; Whitham, M. Exercise, healthy ageing, and the potential role of small extracellular vesicles. J. Physiol 2022, 1–15. [Google Scholar] [CrossRef]
- Bertoldi, K.; Cechinel, L.R.; Schallenberger, B.; Corssac, G.B.; Davies, S.; Guerreiro, I.C.K.; Belló-Klein, A.; Araujo, A.S.R.; Siqueira, I.R. Circulating extracellular vesicles in the aging process: Impact of aerobic exercise. Mol. Cell. Biochem. 2018, 440, 115–125. [Google Scholar] [CrossRef]
- Polgreen, L.E.; Jacobsr, D.R., Jr.; Nathan, B.M.; Steinberger, J.; Moran, A.; Sinaiko, A.R. Association of Osteocalcin with Obesity, Insulin Resistance, and Cardiovascular Risk Factors in Young Adults. Obesity 2012, 20, 2194–2201. [Google Scholar] [CrossRef] [Green Version]
- Moser, S.C.; van der Eerden, B.C.J. Osteocalcin—A Versatile Bone-Derived Hormone. Front. Endocrinol. 2019, 9, 794. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, O.M.; Gabre, A.A.; Sallam, S.F.; El-Alameey, I.R.; Sabry, R.N.; Galal, E.M.; Tawfik, S.M.; Zarouk, W.A.; Mosaad, R.M.; Ramadan, A. Influence of Interleukin-6 (174G/C) Gene Polymorphism on Obesity in Egyptian Children. Open Access Maced. J. Med. Sci. 2017, 5, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.P. Interleukin-6 in Acute Exercise and Training: What Is the Biological Relevance? Exerc. Immunol. Rev 2006, 12, 41. [Google Scholar]
- Peruzzi, B.; Cappariello, A.; Del Fattore, A.; Rucci, N.; De Benedetti, F.; Teti, A. c-Src and IL-6 inhibit osteoblast differentiation and integrate IGFBP5 signalling. Nat. Commun. 2012, 3, 630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetti, F.; Rucci, N.; Del Fattore, A.; Peruzzi, B.; Paro, R.; Longo, M.; Vivarelli, M.; Muratori, F.; Berni, S.; Ballanti, P.; et al. Impaired skeletal development in interleukin-6–transgenic mice: A model for the impact of chronic inflammation on the growing skeletal system. Arthritis Rheum. 2006, 54, 3551–3563. [Google Scholar] [CrossRef]
- Balkhiyarova, Z.; Luciano, R.; Kaakinen, M.; Ulrich, A.; Shmeliov, A.; Bianchi, M.; Chioma, L.; Dallapiccola, B.; Prokopenko, I.; Manco, M. Relationship between glucose homeostasis and obesity in early life—A study of Italian children and adolescents. Hum. Mol. Genet. 2022, 31, 816–826. [Google Scholar] [CrossRef]
- Pedicelli, S.; Fintini, D.; Ravà, L.; Inzaghi, E.; Deodati, A.; Spreghini, M.R.; Bizzarri, C.; Mariani, M.; Cianfarani, S.; Cappa, M.; et al. Prevalence of prediabetes in children and adolescents by class of obesity. Pediatr. Obes. 2022, 17, e12900. [Google Scholar] [CrossRef]
- Shashaj, B.; Luciano, R.; Contoli, B.; Morino, G.S.; Spreghini, M.R.; Rustico, C.; Sforza, R.W.; Dallapiccola, B.; Manco, M. Reference ranges of HOMA-IR in normal-weight and obese young Caucasians. Acta Diabetol. 2016, 53, 251–260. [Google Scholar] [CrossRef]
- Shashaj, B.; Bedogni, G.; Graziani, M.P.; Tozzi, A.E.; DiCorpo, M.L.; Morano, D.; Tacconi, L.; Veronelli, P.; Contoli, B.; Manco, M. Origin of Cardiovascular Risk in Overweight Preschool Children: A Cohort Study of Cardiometabolic Risk Factors at the Onset of Obesity. JAMA Pediatr. 2014, 168, 917–924. [Google Scholar] [CrossRef] [Green Version]
- Del Chierico, F.; Manco, M.; Gardini, S.; Guarrasi, V.; Russo, A.; Bianchi, M.; Tortosa, V.; Quagliariello, A.; Shashaj, B.; Fintini, D.; et al. Fecal microbiota signatures of insulin resistance, inflammation, and metabolic syndrome in youth with obesity: A pilot study. Acta Diabetol. 2021, 58, 1009–1022. [Google Scholar] [CrossRef]
- Manco, M.; Bedogni, G.; Marcellini, M.; Devito, R.; Ciampalini, P.; Sartorelli, M.R.; Comparcola, D.; Piemonte, F.; Nobili, V. Waist circumference correlates with liver fibrosis in children with non-alcoholic steatohepatitis. Gut 2008, 57, 1283–1287. [Google Scholar] [CrossRef]
- Manco, M.; Marcellini, M.; De Vito, R.; Comparcola, D.; Sartorelli, M.; Nobili, V. Metabolic syndrome and liver histology in paediatric non-alcoholic steatohepatitis. Int. J. Obes. 2007, 32, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, K.C.; Crocker, P.R.E.; Donen, R.M.; Honours, B. The Physical Activity Questionnaire for Older Children (PAQ-C) and Adolescents (PAQ-A) Manual. Coll. Kinesiol. Univ. Sask. 2004, 87, 1–38. [Google Scholar]
- Mannocci, A.; Masala, D.; Mei, D.; Tribuzio, A.M.; Villari, P.; LA Torre, G. International Physical Activity Questionnaire for Adolescents (IPAQ A): Reliability of an Italian version. Minerva Pediatr. 2021, 73, 383–390. [Google Scholar] [CrossRef]
- Flegal, K.M.; Wei, R.; Ogden, C.L.; Freedman, D.S.; Johnson, C.L.; Curtin, L.R. Characterizing extreme values of body mass index–for-age by using the 2000 Centers for Disease Control and Prevention growth charts. Am. J. Clin. Nutr. 2009, 90, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Freedman, D.S.; Berenson, G.S. Tracking of BMI z Scores for Severe Obesity. Pediatrics 2017, 140, e20171072. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.G. Using body mass index Z-score among severely obese adolescents: A cautionary note. Int. J. Pediatr. Obes. 2009, 4, 405–410. [Google Scholar] [CrossRef]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Urciuoli, E.; Giorda, E.; Scarsella, M.; Petrini, S.; Peruzzi, B. Osteosarcoma-derived extracellular vesicles induce a tumor-like phenotype in normal recipient cells. J. Cell. Physiol. 2018, 233, 6158–6172. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Weight Cohort (n = 32) | Overweight Cohort (n = 28) | Obese Cohort (n = 22) | p1 | |
|---|---|---|---|---|
| Sex (M/F) | 13/19 (41/59%) | 13/15 (46/54%) | 14/8 (64/36%) | 0.2 |
| Age (years) | 12.35 (10.29–18.77) | 13.08 (10.73–16.17) | 12.52 (9.08–16.21) | 0.9 |
| BMI 2 z-score (SDS) | −0.04 (−3.14–1.02) | 1.43 (1.06–1.62) | 2.05 (1.67–2.64) | <0.0001 |
| BMI (kg/m2) | 18.88 (12.9–25.0) | 24.09 (21.3–28.7) | 29.21 (22.4–38.6) | <0.0001 |
| FG 3 (mmol/l) | 83.03 (70–92) | 83.29 (69–104) | 89.68 (79–100) | 0.0005 |
| FI 4 (ulU/mL) | 9.281 (3.2–26.0) | 15.64 (6.7–34.7) | 19.62 (7.0–41.7) | <0.0001 |
| HOMA 5 -IR | 1.911 (0.6–5.8) | 3.192 (1.4–7.5) | 4.378 (1.4–9.1) | <0.0001 |
| HOMA-B | 186.3 (54.0–699.4) | 353.9 (100.5–2058) | 267.7 (128.0–600.5) | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peruzzi, B.; Urciuoli, E.; Mariani, M.; Chioma, L.; Tomao, L.; Montano, I.; Algeri, M.; Luciano, R.; Fintini, D.; Manco, M. Circulating Extracellular Vesicles Impair Mesenchymal Stromal Cell Differentiation Favoring Adipogenic Rather than Osteogenic Differentiation in Adolescents with Obesity. Int. J. Mol. Sci. 2023, 24, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010447
Peruzzi B, Urciuoli E, Mariani M, Chioma L, Tomao L, Montano I, Algeri M, Luciano R, Fintini D, Manco M. Circulating Extracellular Vesicles Impair Mesenchymal Stromal Cell Differentiation Favoring Adipogenic Rather than Osteogenic Differentiation in Adolescents with Obesity. International Journal of Molecular Sciences. 2023; 24(1):447. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010447
Chicago/Turabian StylePeruzzi, Barbara, Enrica Urciuoli, Michela Mariani, Laura Chioma, Luigi Tomao, Ilaria Montano, Mattia Algeri, Rosa Luciano, Danilo Fintini, and Melania Manco. 2023. "Circulating Extracellular Vesicles Impair Mesenchymal Stromal Cell Differentiation Favoring Adipogenic Rather than Osteogenic Differentiation in Adolescents with Obesity" International Journal of Molecular Sciences 24, no. 1: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010447