

Coupling VIGS with Short- and Long-Term Stress Exposure to Understand the Fiskeby III Iron Deficiency Stress Response

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
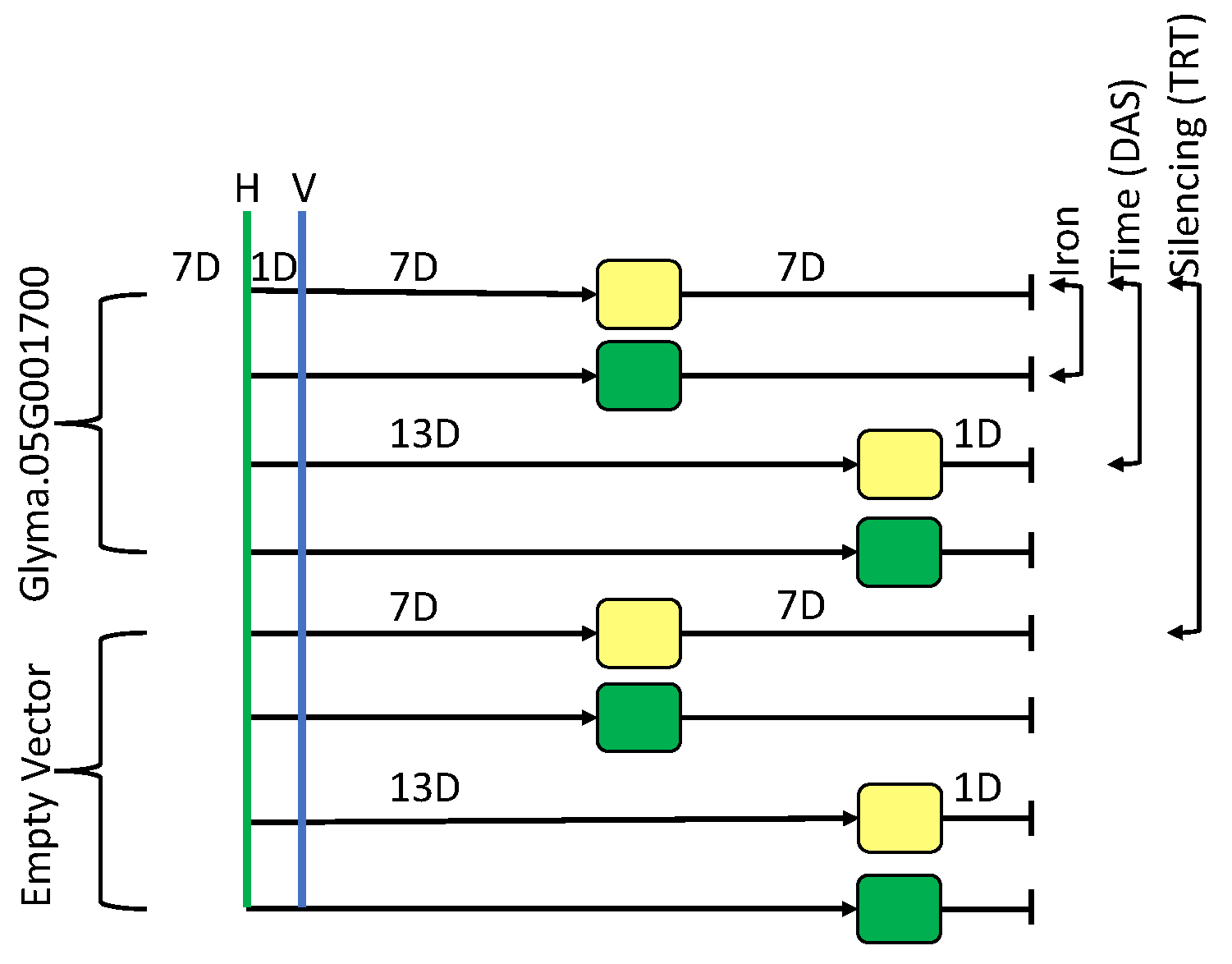
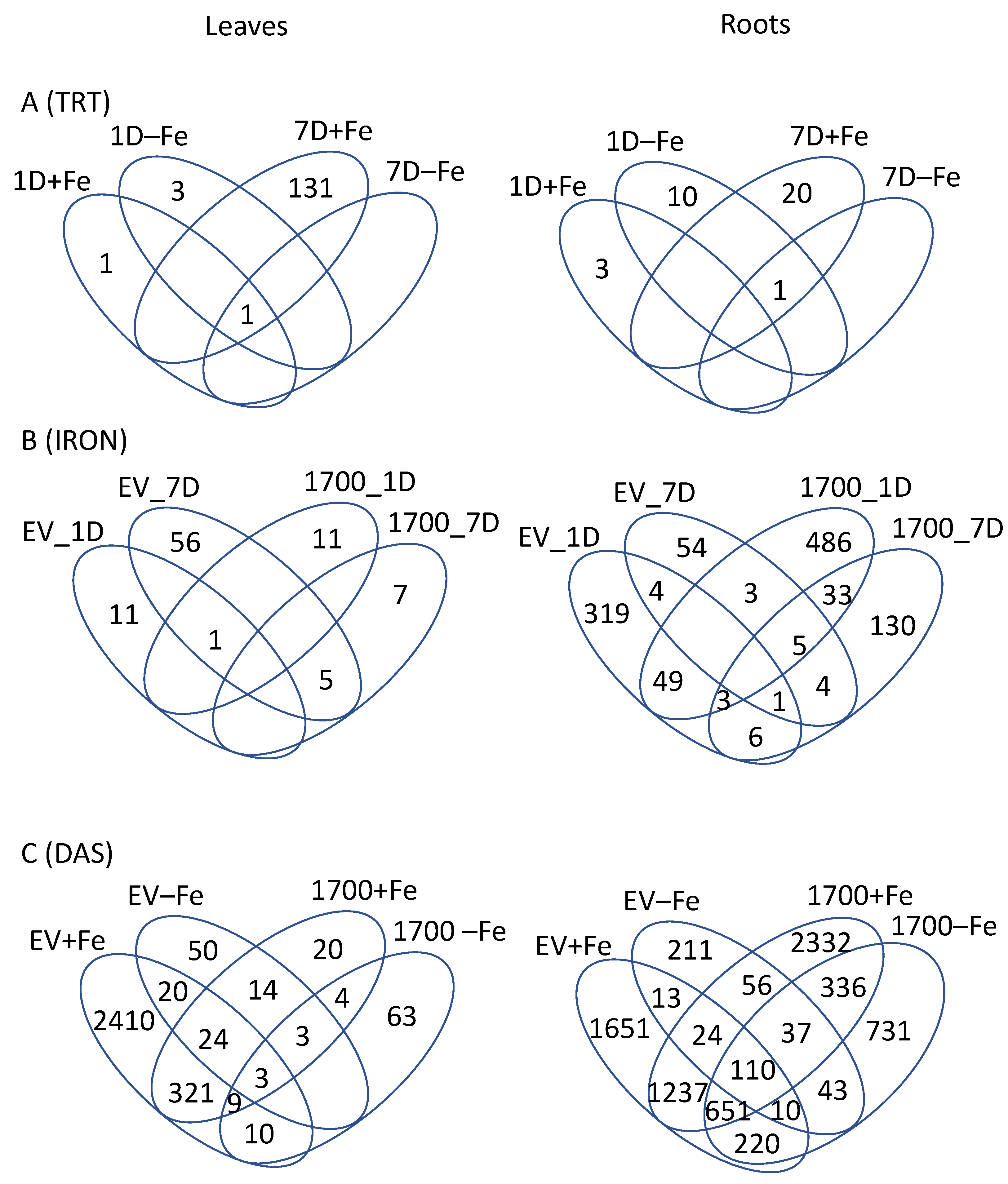
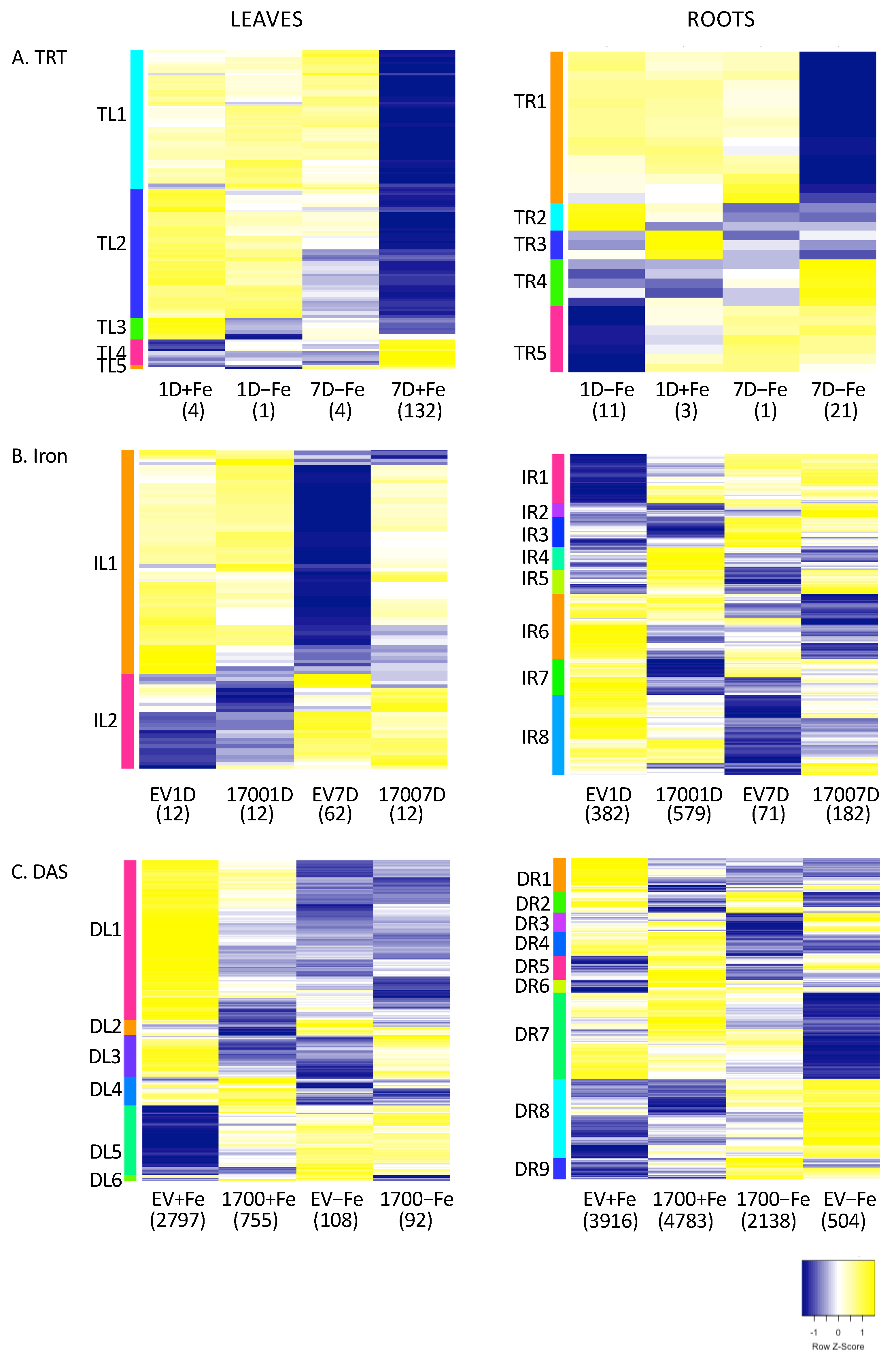
2.1. Effect of Silencing (VIGS_Glyma.05G001700 vs. VIGS_EV)
2.2. Effect of Iron Treatment (+Fe vs. −Fe)
2.3. Effect of Time (1D vs. 7D)
3. Discussion
3.1. Silencing Glyma.05G001700
3.2. Fiskeby III Induces Phosphate Deficiency Responses under −Fe Stress Conditions at 1 Day
3.3. Days after Stress (DAS)
3.4. Conclusions
4. Materials and Methods
4.1. Plant Growth and VIGS Infection
4.2. RNA Extraction and Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- USDA NASS. Soybeans County Maps. Available online: www.nass.usda.gov (accessed on 5 July 2022).
- Ates, A.M.; Bukowski, M. Oil Crops Outlook: May 2022, OCS-22e; U.S. Department of Agriculture, Economic Research Service: Washington, DC, USA, 2022. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press Limited: San Diego, CA, USA, 1995. [Google Scholar]
- Clemens, S.; Weber, M. The essential role of courmarin secretion for Fe acquisition from alkaline soil. Plant Signal. Behav. 2016, 11, e1114197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liesch, A.M.; Diaz, D.R.; Roozeboom, K.L. Interpreting relationships between soil variables adn soybean iron deficiency using factor analysis. Soil Sci. Soc. Am. J. 2012, 76, 1311–1318. [Google Scholar] [CrossRef]
- Kaiser, D.E.; Lamb, J.; Hernandez, J.A. Comparison of field management strategies for preventing iron deficiency chlorosis in soybean. Agron. J. 2014, 106, 1963–1974. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.L.; Burkey, K.O.; Carter, T.E., Jr.; Orf, J.; Cregan, P.B. Phenotypic variation and identification of quantitative trait loci for ozone tolerance in a Fiskeby III x Mandarin (Ottawa) soybean population. Theor. Appl. Genet. 2016, 129, 1113–1125. [Google Scholar] [CrossRef]
- Do, T.D.; Vuong, T.D.; Dunn, D.; Smothers, S.; Patil, G.; Yungbluth, D.C.; Chen, P.; Scaboo, A.; Xu, D.; Carter, T.E.; et al. Mapping and confirmation of loci for salt tolerance in a novel soybean germplasm, Fiskeby III. Theor. Appl. Genet. 2018, 131, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Whaley, A.; Sheridan, J.; Safari, S.; Burton, A.; Burkey, K.; Schlueter, J. RNA-seq analysis reveals genetic response and tolerance mechanisms to ozone exposure in soybean. BMC Genom. 2015, 16, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rod, K.S.; Walker, D.; Bradley, C.A. Evaluation of major ancestors of North American soybean cultivars for resistance to three Pythium species that cause seedling blight. Plant Dis. 2018, 102, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.; Burkey, K.; Taggart, M.; Rufty, T. Leaf traits that contribute to differential ozone response in ozone-tolerant and sensitive soybean genotypes. Plants 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Merry, R.; Butenhoff, K.; Campbell, B.; Michno, J.-M.; Wang, D.; Orf, J.; Lorenz, A.; Stupar, R. Identification and fine-mapping of a soybean quantitative trait locus on chromosome 5 confering tolerance to iron deficiency chlorosis. Plant Genome 2019, 12, 190007. [Google Scholar] [CrossRef] [Green Version]
- Atwood, S.E.; O'Rourke, J.A.; Peiffer, G.A.; Yin, T.; Majumder, M.; Zhang, C.; Cianzio, S.R.; Hill, J.H.; Cook, D.; Whitham, S.A.; et al. Replication protein A subunit 3 and the iron efficiency response in soybean. Plant Cell Environ. 2014, 37, 213–234. [Google Scholar] [CrossRef]
- Assefa, T.; Zhang, J.; Chowda-Reddy, R.V.; Moran Lauter, A.N.; Singh, A.; O’Rourke, J.A.; Graham, M.A.; Singh, A.K. Deconstrcting the genetic architecture of iron deficiency chlorosis in soybean using genome-wide approaches. BMC Plant Biol. 2020, 20, 42. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, H.; Moriyama, S.; Kusakizako, T.; Kumazaki, K.; Nakane, T.; Yamashita, K.; Hirata, K.; Dohmae, N.; Nishizawa, T.; Ito, K.; et al. Structural basis for xenobiotic extrusion by eukaryotic MATE transporter. Nat. Commun. 2017, 8, 1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, Y.; Wang, W.; Gai, J.; Li, Y. Genome-wide analysis of MATE transporters and expression patterns of a subgroup of MATE genes in response to aluminum toxicity in soybean. BMC Genom. 2016, 17, 223. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, N.; Kar, D.; Mahajan, B.D.; Nanda, S.; Rahiman, R.; Panchakshari, N.; Bhagavatula, L.; Datta, S. The multitasking abilities of MATE transporters in plants. J. Exp. Bot. 2019, 70, 4643–4656. [Google Scholar] [CrossRef] [PubMed]
- Rogers, E.E.; Wu, X.; Stacey, G.; Nguyen, H.T. Two MATE proteins play a role in iron efficiency in soybean. J. Plant Physiol. 2009, 166, 1453–1459. [Google Scholar] [CrossRef]
- Seo, P.J.; Park, J.; Park, M.-J.; Kim, Y.-S.; Kim, S.-G.; Jung, J.-H.; Park, C.-M. A golgi-localized MATE transporter mediates iron homeostasis under osmotic stress in Arabidopsis. Biochem. J. 2012, 442, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Durrett, T.P.; Gassmann, W.; Rogers, E.E. The FRD3-mediated efflux of citrate into the root vasculature is necessary for efficient iron translocation. Plant Physiol. 2007, 144, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.E.; Guerinot, M.L. FRD3, a member of the multidrug and toxin efflux family, controls iron deficiency responses in Arabidopsis. Plant Cell 2002, 14, 1787–1799. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Qian, C.; Guo, X.; Liu, E.; Mao, K.; Mu, C.; Chen, N.; Zhang, W.; Liu, H. ELS1, a novel MATE transporter related to leaf senesence and iron homeostasis in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2016, 476, 319–325. [Google Scholar] [CrossRef]
- Moran Lauter, A.N.; Rutter, L.; Cook, D.; O’Rourke, J.A.; Graham, M.A. Examining short-term responses to a long-term problem: RNA-seq analyses of iron deficiency chlorosis tolerant soybean. Int. J. Mol. Sci. 2020, 21, 3591. [Google Scholar] [CrossRef]
- Atencio, L.; Salazar, J.; Moran Lauter, A.N.; Gonzales, M.D.; O’Rourke, J.A.; Graham, M.A. Characterizing short and long term iron stress responses in iron deficiency tolerant and susceptible soybean (Glycine max L. Merr). Plant Stress 2021, 2, 100012. [Google Scholar] [CrossRef]
- Kohlhase, D.R.; McCabe, C.E.; Singh, A.K.; O’Rourke, J.A.; Graham, M.A. Comparing Early Transcriptomic Responses of 18 Soybean (Glycine max) Genotypes to Iron Stress. Int. J. Mol. Sci. 2021, 22, 11643. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, J.A.; McCabe, C.E.; Graham, M.A. Dynamic gene expression changes in response to micronutrient, macronutrient, and multiple stress exposure in soybean. Funct. Integr. Genom. 2020, 20, 321–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckhout, T.J.; Yang, T.J.W.; Schmidt, W. Early iron-deficiency-induced transcriptional changes in Arabidopsis roots as revealed by microarray analyses. BMC Genom. 2009, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Castro-Guerrero, N.A.; McInturf, S.A.; Nguyen, N.T.; Dame, A.N.; Wang, J.; Bindbeutel, R.K.; Joshi, T.; Jurisson, S.S.; Nusinow, D.A.; et al. Changes in iron availability in Arabidopsis are rapidly sensed in the leaf vasculature and impaired sensing leads to opposite transcriptional programs in leaves and roots. Plant Cell Environ. 2018, 41, 2263–2276. [Google Scholar] [CrossRef]
- Dolan, W.L.; Chapple, C. Transcriptome analysis of four Arabidopsis thaliana mediator tail mutants reveals overlapping and unique functions in gene regulation. Genes Genomes Genet. 2018, 8, 3093–3108. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Du, X.; Mou, Z. The mediator complex subunits MED14, MED15, and MED16 are involved in defense signaling crosstalk in Arabidopsis. Front. Plant Sci. 2016, 7, 1947. [Google Scholar] [CrossRef] [Green Version]
- Bonawitz, N.D.; Kim, J.I.; Tobimatus, Y.; Cleslelski, P.N.; Anderson, N.A.; Ximenes, E.; Maeda, J.; Ralph, J.; Donohoe, B.S.; Ladisch, M.; et al. Disruption of Mediator rescues the stunted growth of a lignin-deficient Arabidopsis mutant. Nature 2014, 509, 376–380. [Google Scholar] [CrossRef]
- Guo, J.; Wei, L.; Chen, S.-S.; Cai, X.-W.; Su, Y.-N.; Li, L.; Chen, S.; He, X.-J. The CBP/p300 histone acetyltransferases function as plant-specific MEDIATOR subunits in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 755–771. [Google Scholar] [CrossRef]
- Hemsley, P.A.; Hurst, C.H.; Kaliyadasa, E.; Lamb, R.; Knight, M.R.; De Cothi, E.A.; Steele, J.F.; Knight, H. The Arabidopsis mediator complex subunits MED16, MED14, and MED2 regulate mediator and RNA polymerase II recruitment to CBF-responsive cold-regulated genes. Plant Cell 2014, 26, 465–484. [Google Scholar] [CrossRef]
- Weis, C.; Huckelhoven, R.; Eichmann, R. LIFEGUARD proteins support plant colonization by biotrophic powdery mildew fungi. J. Exp. Bot. 2013, 64, 3855–3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, N.; Caruana, J.; Erdem, I.; Raina, R. An Arabidopsis DISEASE RELATED NONSPECIFIC LIPID TRANSFER PROTEIN 1 is required for resistance against various phytopathogens and tolerance to salt stress. Gene 2020, 753, 144802. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wei, Z.; Yang, Z.; Li, F.; Wang, Z. Updated mechanisms of GCN5- the monkey king of the plant kingdom in plant development and resistance to abiotic stresses. Cells 2021, 10, 979. [Google Scholar]
- Dwyer, M.E.; Hangarter, R.P. Light-dependent phosphorylation of THRUMIN1 regulates its association with actin filaments and 14-3-3 proteins. Plant Physiol. 2021, 187, 1445–1461. [Google Scholar] [CrossRef] [PubMed]
- Chutia, R.; Abel, S.; Ziegler, J. Iron and phosphate deficiency regulators concertedly control coumarin profiles in Arabidopsis thaliana roots during iron, phosphate, and combined deficiencies. Front. Plant Sci. 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, J.A.; Morrisey, M.J.; Merry, R.; Espina, M.J.; Lorenz, A.J.; Stupar, R.M.; Graham, M.A. Mining Fiskeby III and Mandarin (Ottawa) Expression Profiles to Understand Iron Stress Tolerant Responses in Soybean. Int. J. Mol. Sci. 2021, 22, 11032. [Google Scholar] [CrossRef]
- Zientara, K.; Wawrzyńska, A.; Łukomska, J.; López-Moya, J.R.; Liszewska, F.; Assunção, A.G.; Aarts, M.G.; Sirko, A. Activity of the AtMRP3 promoter in transgenic Arabidopsis thaliana and Nicotiana tabacum plants is increased by cadmium, nickel, arsenic, cobalt and lead but not by zinc and iron. J. Biotechnol. 2009, 139, 258–263. [Google Scholar] [CrossRef]
- Rodriguez-Celma, J.; Chou, H.; Kobayashi, T.; Long, T.A.; Balk, J. Hemerythrin E3 ubiquitin ligases as negative regulators of iron homeostasis in plants. Front. Plant Sci. 2019, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yang, S.; Fan, B.; Zhu, C.; Chen, Z. The Mediator Complex: A central coordinator of plant adaptive responses to environmental stresses. Int. J. Mol. Sci. 2022, 23, 6170. [Google Scholar] [CrossRef]
- Dolan, W.L.; Dilkes, B.P.; Stout, J.M.; Bonawitz, N.D.; Chapple, C. Mediator complex subunits MED2, MED5, MED16, and MED23 genetically interact in the regulation of phenylpropanoid biosynthesis. Plant Cell 2017, 29, 3269–3285. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.-M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Waters, B.M.; Amundsen, K.; Graef, G. Gene expression profiling of iron deficiency chlorosis sensitive and tolerant soybean indicates key roles for phenylpropanoids under alkalinity stress. Front. Plant Sci. 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penninckx, I.; Thomma, B.P.H.J.; Buchala, A.; Metraux, J.P.; Broekert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 19, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, P.-Y.; Cheng, C.-P.; Koh, K.W.; Chan, M.-T. The Arabidopsis defensin gene, AtPDF1.1, mediates defence against Pectobacterium carotovorum subsp. carotovorum via an iron-witholding defence system. Sci. Rep. 2017, 7, 9175. [Google Scholar] [CrossRef] [Green Version]
- Moran Lauter, A.N.; Peiffer, G.A.; Yin, T.; Whitham, S.A.; Cook, D.; Shoemaker, R.C.; Graham, M.A. Identification of candidate genes involved in early iron deficiency chlorosis signaling in soybean (Glycine max) roots and leaves. BMC Genom. 2014, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Richter, A.S.; Tohge, T.; Fernie, A.R.; Grimm, B. The genomes uncoupled-dependent signaling pathway coordinates plastid biogenesis with the synthesis of anthocyanins. Phil. Trans. R. Soc. B 2020, 375, 20190403. [Google Scholar] [CrossRef]
- Huang, W.-L.; Wu, F.-L.; Huang, H.-Y.; Huang, W.T.; Deng, C.-L.; Yang, L.-T.; Huang, Z.-R.; Chen, L.-S. Excess copper-induced alterations of protein profiles and related physiological parameters in Citrus leaves. Plants 2020, 9, 291. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.-T.; Wang, F.; Ma, Y.-P.; Jia, L.-J.; Liu, N.; Wang, H.-Y.; Zhao, P.; Xia, G.-X.; Zhong, N.-Q. Ectopic expression of SsPETE2, a plastocyanin from Suaeda salsa, improves plant tolerance to abiotic stress. Plant Sci. 2018, 268, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chodasiewicz, M.; Sokolowska, E.M.; Nelson-Dittrich, A.C.; Masiuk, A.; Beltran, J.C.M.; Nelson, A.D.L.; Skirycz, A. Identification and characterization of the heat induced plastidial stress granules reveal new insight into the Arabidopsis stress response. Front. Plant Sci. 2020, 11, 595792. [Google Scholar] [CrossRef]
- Zhao, P.; Cui, R.; Xu, P.; Wu, J.; Mao, J.-L.; Chen, Y.; Zhou, C.-Z.; Yu, L.-H.; Xiang, C.-B. ATHB17 enhances stress tolerance by coordinating photosynthesis associated nuclear gene and ATSIG6 expression in response to abioitc stress. Sci. Rep. 2017, 7, 45492. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.J.; Zhu, X.F.; Wang, Z.W.; Dong, F.; Dong, N.Y.; Zheng, S.J. Abscisic acid alleviates iron deficiency by promoting root iron reutilization and transport from root to shoot in Arabidopsis. Plant Cell Environ. 2013, 37, 852–863. [Google Scholar] [CrossRef]
- Zhang, J.-C.; Wang, X.-F.; Wang, X.-N.; Wang, F.-P.; Ji, X.-L.; An, J.-P.; Yang, K.; Zhao, Q.; You, C.-X.; Hao, Y.-J. Abscisic acid alleviates iron deficiency by regulating iron distribution in roots and shoots of apple. Sci. Hortic. 2020, 262, 109018. [Google Scholar] [CrossRef]
- Chong, L.; Guo, P.; Zhu, Y. Mediator Complex: A pivotal regulator of ABA signaling pathway and abiotic stress response in plants. Int. J. Mol. Sci. 2020, 21, 7755. [Google Scholar] [CrossRef]
- Long, T.A.; Tsukagoshi, H.; Busch, W.; Lahner, B.; Salt, D.E.; Benfey, P.N. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell 2010, 22, 2219–2236. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Fabre, R.; Gullien, G.; Loredo, M.; Arellano, J.; Valdes-Lopez, O.; Ramirez, M.; Iniguez, L.P.; Panzeri, D.; Castiglioni, B.; Cremonesi, P. Common bean (Phaseolus vulgaris L.) PvTIFY orchestrates global changes in transcript profile response to jasmonate and phosphorus deficiency. BMC Plant Biol. 2013, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.X.; Stridh, M.H.; Larsson, K.E.; Liljenberg, C.; Sandelius, A.S. Phosphate-deficient oat replaces a major portion of the plasma membrane phospholipids with the galactolipid digalactosyldiacylglycerol. FEBS Lett. 2003, 537, 128–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehara, K.; Yu, C.-Y.; Cho, Y.; Cheong, W.-F.; Torta, F.; Shui, G.; Wenk, M.R.; Nakamura, Y. Arabidopsis AtPLC2 is a primary phosphoinositide-specific phospholipase C in phosphoinositide metabolism and the endoplasmic reticulum stress response. PLoS Genet. 2015, 11, e1005511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, J.; Graham, M.A. Gene expression responses to sequential nutrient deficiency stresses in soybean. Int. J. Mol. Sci. 2021, 22, 1252. [Google Scholar] [CrossRef] [PubMed]
- Chaney, R.L.; Coulombe, B.A.; Bell, P.F.; Angle, J.S. Detailed method to screen dicot cultivars for resistance to Fe-chlorosis using FeDTPA and bicarbonate in nutrient solutions. J. Plant Nutr. 1992, 15, 2063–2083. [Google Scholar] [CrossRef]
- O'Rourke, J.A.; Graham, M.A.; Vodkin, L.; Gonzalez, D.O.; Cianzio, S.R.; Shoemaker, R.C. Recovering from iron deficiency chlorosis in near-isogenic soybeans: A microarray study. Plant Physiol. Biochem. 2007, 45, 287–292. [Google Scholar] [CrossRef]
- Lin, S.-F.; Baumer, J.S.; Ivers, D.; de Cianzo, S.R.; Shoemaker, R.C. Field and nutrient solution tests measure similar mechanisms controlling iron deficiency chlorosis in soybean. Crop Sci. 1998, 38, 254–259. [Google Scholar] [CrossRef]
- Lin, S.F.; Baumer, J.; Ivers, D.; Cianzio, S.R.; Shoemaker, R.C. Nutrient solution screening of Fe chlorosis resistance in soybean evaluated by molecular characterization. J. Plant Nutr. 2000, 23, 1915–1928. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Babraham Bioinformatics. Online [Mar 2016] 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 December 2021).
- Wingett, S.W.; Andrews, S. FastQ Screen: A tool for multi-genome mapping nd quality control (version 0.14.1). F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef]
- Daehwan, K.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2015; Available online: http://www.rstudio.com (accessed on 4 January 2022).
- Lawrence, M.; Gentleman, R.; Carey, V. rtracklayer: An R package for interfacing with genome browsers. Bioinformatics 2009, 25, 1841–1842. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.; Huber, W.; Pagès, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. software for computing and annotating genomic ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lun, A.A.T.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [PubMed] [Green Version]
- Morales, A.M.A.P.; O'Rourke, J.A.; van de Mortel, M.; Scheider, K.T.; Bancroft, T.J.; Borem, A.; Nelson, R.T.; Nettleton, D.; Baum, T.J.; Shoemaker, R.C.; et al. Transcriptome analyses and virus induced gene silencing identify genes in the Rpp4-mediated Asian soybean rust resistance pathway. Funct. Plant Biol. 2013, 40, 1029–1047. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R. The Design of Experiments, 8th ed.; London Oliver and Boyd: Edinburgh, UK, 1966. [Google Scholar]
- Bonferroni, C. III Calcolo Delle Assicurazioni Su Gruppi Di Teste; Studi Onore Del Profr. Salvatore Ortu Carboni: Rome, Italy, 1935; pp. 13–60. [Google Scholar]
- Wang, Z.; Libault, M.; Joshi, T.; Valliyodan, B.; Nguyen, H.T.; Xu, D.; Stacey, G.; Cheng, J. SoyDB: A knowledge database of soybean transcription factors. BMC Plant Biol. 2010, 10, 14. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Rourke, J.A.; Graham, M.A. Coupling VIGS with Short- and Long-Term Stress Exposure to Understand the Fiskeby III Iron Deficiency Stress Response. Int. J. Mol. Sci. 2023, 24, 647. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010647
O’Rourke JA, Graham MA. Coupling VIGS with Short- and Long-Term Stress Exposure to Understand the Fiskeby III Iron Deficiency Stress Response. International Journal of Molecular Sciences. 2023; 24(1):647. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010647
Chicago/Turabian StyleO’Rourke, Jamie A., and Michelle A. Graham. 2023. "Coupling VIGS with Short- and Long-Term Stress Exposure to Understand the Fiskeby III Iron Deficiency Stress Response" International Journal of Molecular Sciences 24, no. 1: 647. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010647