
Aggression Results in the Phosphorylation of ERK1/2 in the Nucleus Accumbens and the Dephosphorylation of mTOR in the Medial Prefrontal Cortex in Female Syrian Hamsters
 ,
,
Abstract
:1. Introduction
2. Results
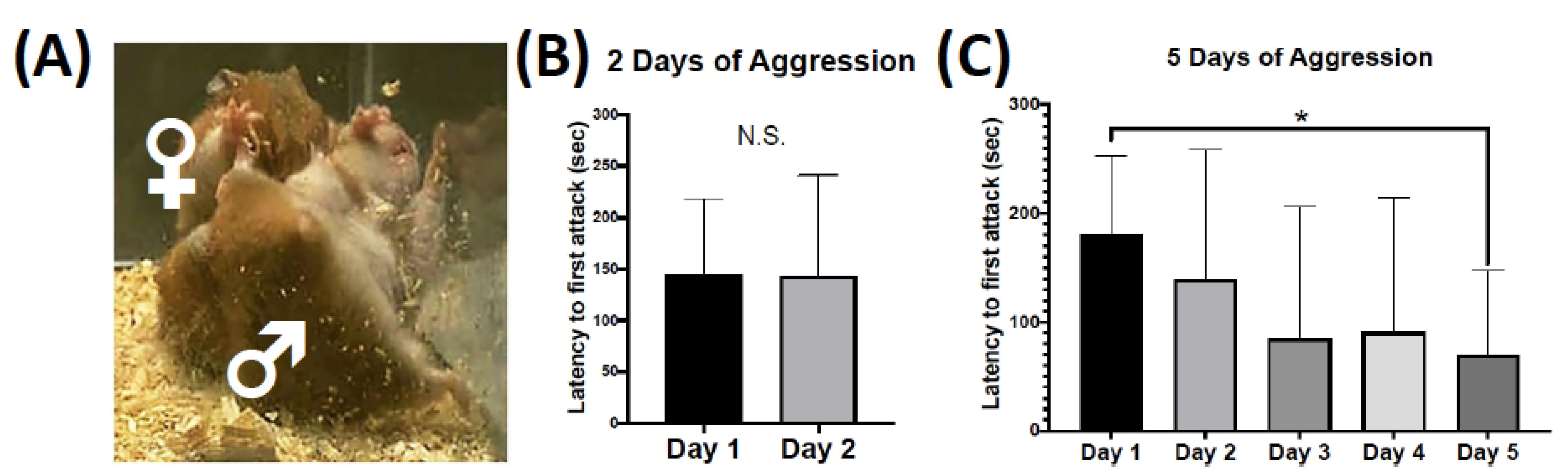
2.1. Behavioral Plasticity for Two versus Five Consecutive Days of Aggressive Behavior
2.1.1. Effect of Aggressive Experience on the Phosphorylation and Expression of Kinases and Phosphatases of FMRP in the Nucleus Accumbens
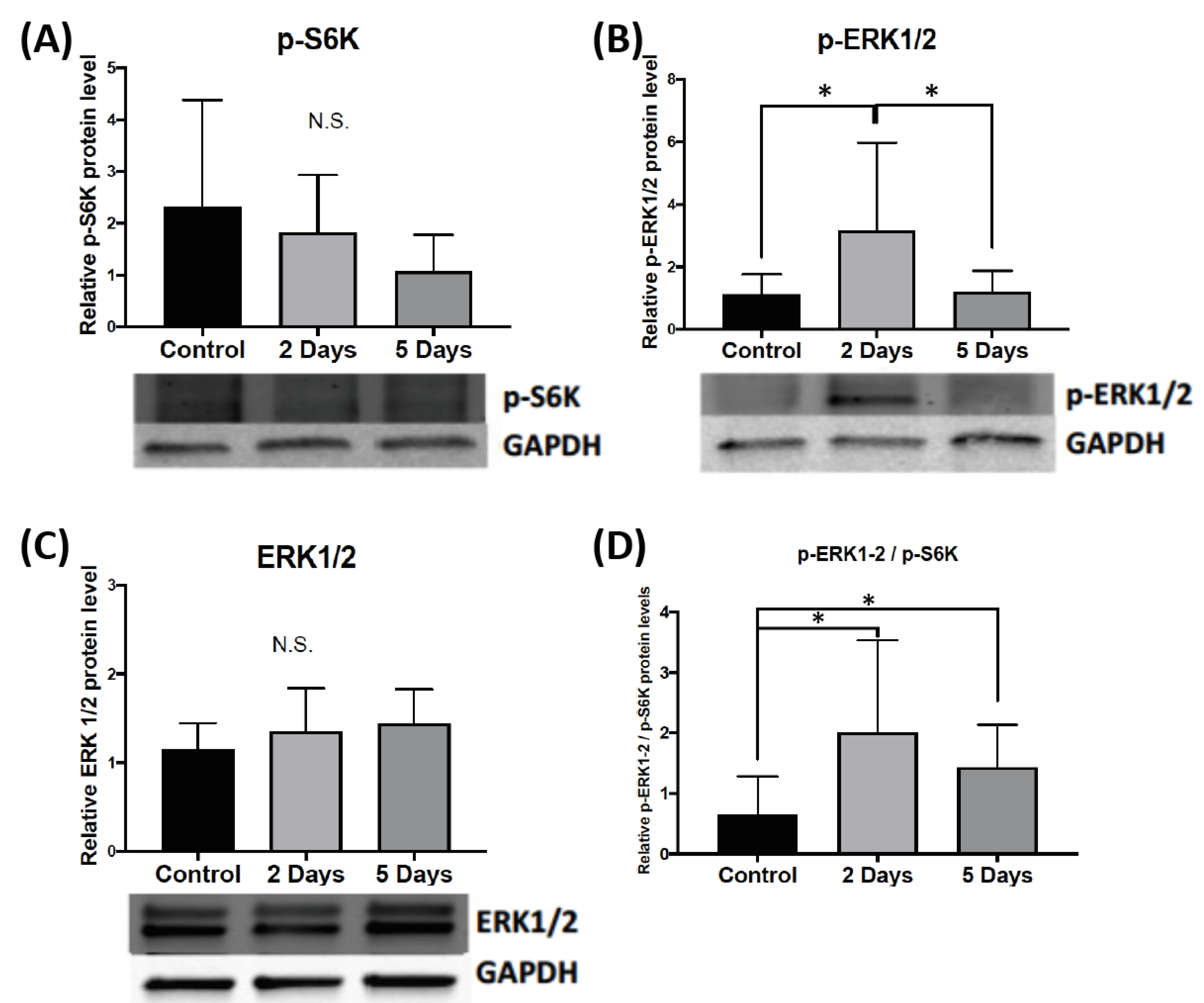
2.1.2. Effect of Aggressive Experience on the Phosphorylation and Expression of Upstream Signaling Molecules that Regulate FMRP Kinases and Phosphatases in the Nucleus Accumbens
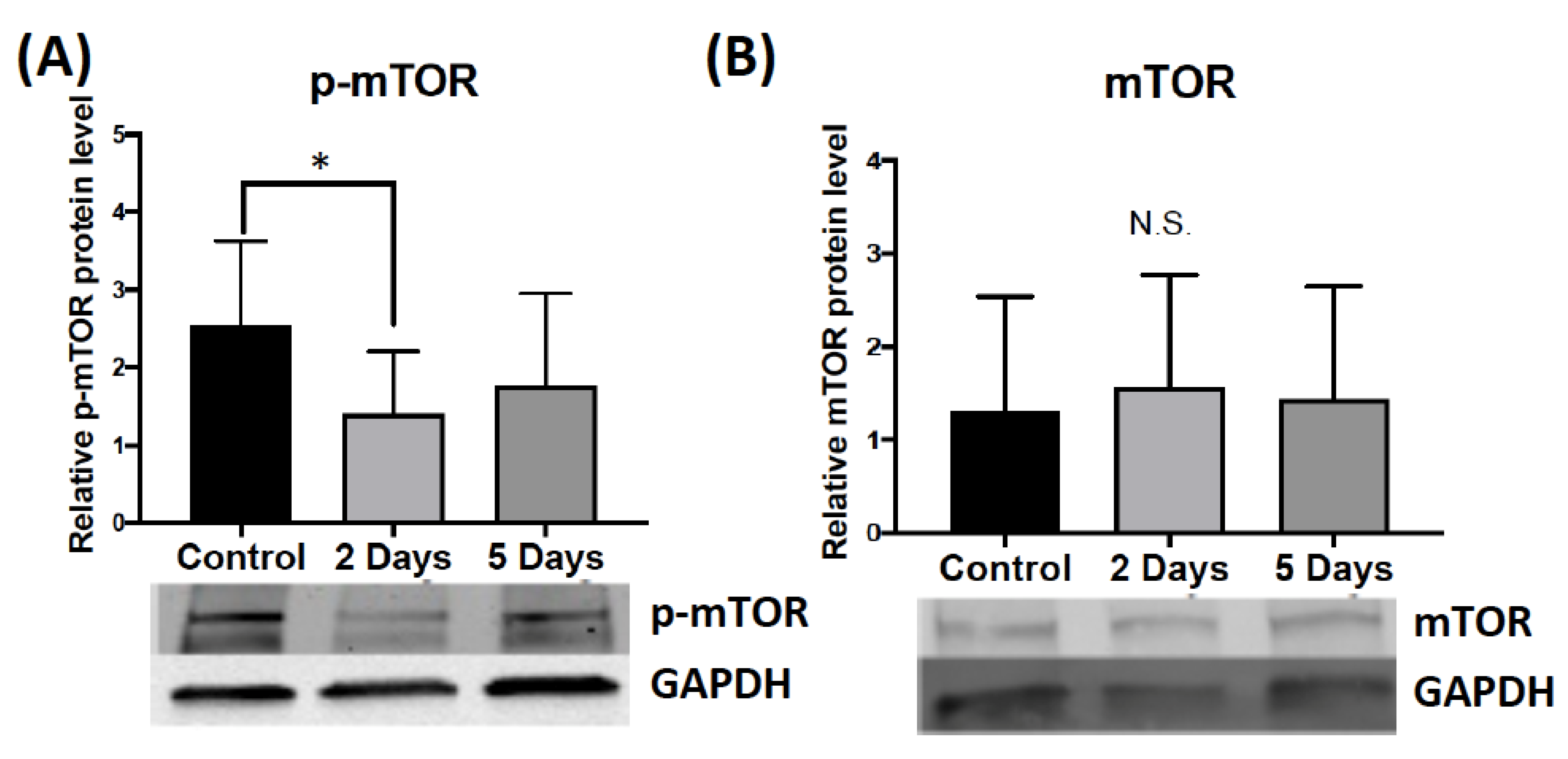
2.1.3. Effect of Aggressive Experience on the Phosphorylation and Expression of Secondary Signaling Molecules that Regulate FMRP Kinases and Phosphatases in the Medial Prefrontal Cortex
2.2. Effect of Aggressive Experience on the Phosphorylation and Expression of the Translational Regulator FMRP in the Forebrain
3. Discussion
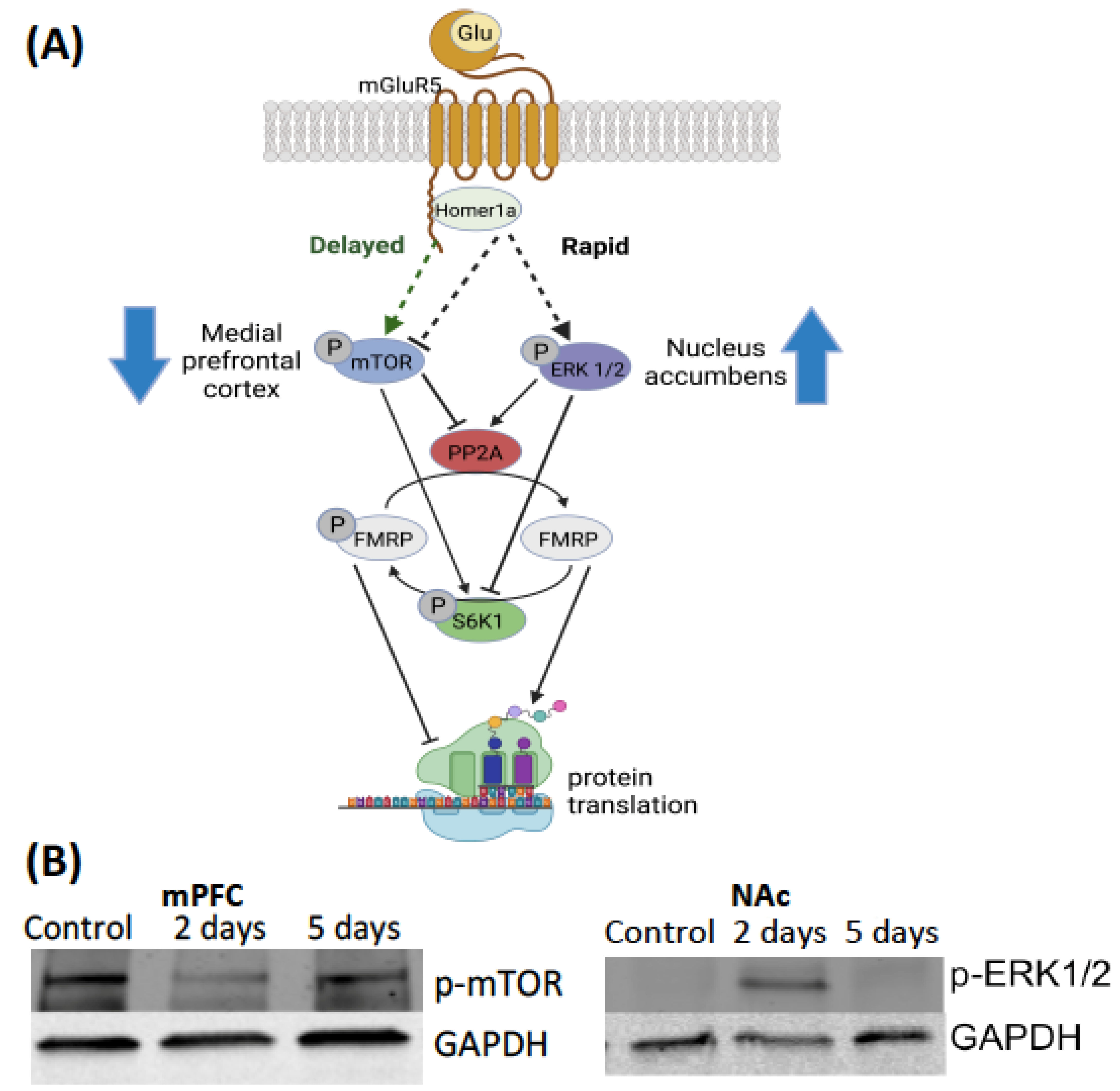
3.1. Intracellular Mechanisms of Translation
3.2. Heterogeneity of Intracellular Signaling
3.3. Challenges of Interpretation
4. Materials and Methods
4.1. Animals
4.2. Ovariectomy
4.3. Behavioral Testing and Scoring
4.4. Tissue Collection
4.5. Tissue Preparation
4.6. Western Blot
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pettinger, A.M.; Steiger, S.; Muller, J.K.; Sakaluk, S.K.; Eggert, A.K. Dominance status and carcass availability affect the outcome of sperm competition in burying beetles. Behav. Ecol. 2011, 22, 1079–1087. [Google Scholar] [CrossRef]
- Stockley, P.; Bro-Jorgensen, J. Female competition and its evolutionary consequences in mammals. Biol. Rev. Camb. Philos. Soc. 2011, 86, 341–366. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Makinodan, M.; Kishimoto, T. Social status and modern-type depression: A review. Brain Behav. 2019, 9, e01464. [Google Scholar] [CrossRef] [PubMed]
- Snyder-Mackler, N.; Burger, J.R.; Gaydosh, L.; Belsky, D.W.; Noppert, G.A.; Campos, F.A.; Bartolomucci, A.; Yang, Y.C.; Aiello, A.E.; O’Rand, A.; et al. Social determinants of health and survival in humans and other animals. Science 2020, 368, eaax9553. [Google Scholar] [CrossRef] [PubMed]
- Been, L.E.; Gibbons, A.B.; Meisel, R.L. Towards a neurobiology of female aggression. Neuropharmacology 2019, 156, 107451. [Google Scholar] [CrossRef]
- Terranova, J.I.; Song, Z.; Larkin, T.E.; Hardcastle, N.; Norvelle, A.; Riaz, A.; Albers, H.E. Serotonin and arginine-vasopressin mediate sex differences in the regulation of dominance and aggression by the social brain. Proc. Natl. Acad. Sci. USA 2016, 113, 13233–13238. [Google Scholar] [CrossRef] [Green Version]
- Borland, J.M.; Aiani, L.M.; Norvelle, A.; Grantham, K.N.; O’Laughlin, K.; Terranova, J.; Frantz, K.J.; Albers, H.E. Sex-dependent regulation of social reward by oxytocin receptors in the ventral tegmental area. Neuropsychopharmacology 2019, 44, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Meisel, R.L.; Joppa, M.A. Conditioned place preference in female hamsters following aggressive or sexual encounters. Physiol. Behav. 1994, 56, 1115–1118. [Google Scholar] [CrossRef]
- Borland, J.M.; Kim, E.; Swanson, S.P.; Rothwell, P.E.; Mermelstein, P.G.; Meisel, R.L. Effect of aggressive experience in female Syrian hamsters on glutamate receptor expression in the nucleus accumbens. Front. Behav. Neurosci. 2020, 14, 583395. [Google Scholar] [CrossRef]
- Been, L.E.; Moore, K.M.; Kennedy, B.C.; Meisel, R.L. Metabotropic glutamate receptor and fragile X signaling in a female model of escalated aggression. Biol. Psychiatry 2016, 79, 685–692. [Google Scholar] [CrossRef]
- Staffend, N.A.; Meisel, R.L. Aggressive experience increases dendritic spine density within the nucleus accumbens core in female Syrian hamsters. Neuroscience 2012, 227, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Cory, S. The developing synapse: Construction and modulation of synaptic structures and circuits. Science 2002, 298, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Luchkina, N.V.; Coleman, S.K.; Huupponen, J.; Cai, C.; Kivistö, A.; Taira, T.; Keinänen, K.; Lauri, S.E. Molecular mechanisms controlling synaptic recruitment of GluA4 subunit-containing AMPA-receptors critical for functional maturation of CA1 glutamatergic synapses. Neuropharmacology 2017, 112, 46–56. [Google Scholar] [CrossRef]
- Scheyer, A.F.; Wolf, M.E.; Tseng, K.Y. A protein synthesis-dependent mechanism sustains calcium-permeable AMPA receptor transmission in nucleus accumbens synapses during withdrawal from cocaine self-administration. J. Neurosci. 2014, 34, 3095–3100. [Google Scholar] [CrossRef] [Green Version]
- Muddashetty, R.S.; Kelic, S.; Gross, C.; Xu, M.; Bassell, G.J. Dysregulated metabotropic glutamate receptor-dependent translation of AMPA receptor and postsynaptic density-95 mRNAs at synapses in a mouse model of fragile X syndrome. J. Neurosci. 2007, 27, 5338–5348. [Google Scholar] [CrossRef] [Green Version]
- Kute, P.M.; Ramakrishna, S.; Neelagandan, N.; Chattarji, S.; Muddashetty, R.S. NMDAR mediated translation at the synapse is regulated by MOV10 and FMRP. Mol. Brain 2019, 12, 65. [Google Scholar] [CrossRef]
- Clifton, N.E.; Thomas, K.L.; Wilkinson, L.S.; Hall, J.; Trent, S. FMRP and CYFIP1 at the synapse and their role in psychiatric vulnerability. Complex Psychiatry 2020, 6, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Sidorov, M.S.; Auerbach, B.D.; Bear, M.F. Fragile X mental retardation protein and synaptic plasticity. Mol. Brain 2013, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhao, H.; Lu, C.; Li, X.; Xie, Y.; Fu, H.; Xu, H. Synaptic plasticity, a prominent contributor to the anxiety in fragile X syndrome. Neural Plast. 2016, 2016, 9353929. [Google Scholar] [CrossRef] [Green Version]
- D’Incal, C.; Broos, J.; Torfs, T.; Kooy, R.F.; Vanden Berghe, W. Towards kinase inhibitor therapies for fragile X syndrome: Tweaking twists in the autism spectrum kinase signaling network. Cells 2022, 11, 1325. [Google Scholar] [CrossRef]
- Narayanan, U.; Nalavadi, V.; Nakamoto, M.; Pallas, D.C.; Ceman, S.; Bassell, G.J.; Warren, S.T. FMRP phosphorylation reveals an immediate-early signaling pathway triggered by group I mGluR and mediated by PP2A. J. Neurosci. 2007, 27, 14349–14357. [Google Scholar] [CrossRef] [Green Version]
- Sunahori, K.; Nagpal, K.; Hedrich, C.M.; Mizui, M.; Fitzgerald, L.M.; Tsokos, G.C. The catalytic subunit of protein phosphatase 2A (PP2Ac) promotes DNA hypomethylation by suppressing the phosphorylated mitogen-activated protein kinase/extracellular signal-regulated kinase (ERK) kinase (MEK)/phosphorylated ERK/DNMT1 protein pathway in T-cells from controls and systemic lupus erythematosus patients. J. Biol. Chem. 2013, 288, 21936–21944. [Google Scholar] [CrossRef] [Green Version]
- Osterweil, E.K.; Krueger, D.D.; Reinhold, K.; Bear, M.F. Hypersensitivity to mGluR5 and ERK1/2 leads to excessive protein synthesis in the hippocampus of a mouse model of fragile X syndrome. J. Neurosci. 2010, 30, 15616–15627. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, U.; Nalavadi, V.; Nakamoto, M.; Thomas, G.; Ceman, S.; Bassell, G.J.; Warren, S.T. S6K1 phosphorylates and regulates fragile X mental retardation protein (FMRP) with the neuronal protein synthesis-dependent mammalian target of rapamycin (mTOR) signaling cascade. J. Biol. Chem. 2008, 283, 18478–18482. [Google Scholar] [CrossRef] [Green Version]
- Parkington, J.D.; LeBrasseur, N.K.; Siebert, A.P.; Fielding, R.A. Contraction-mediated mTOR, p70S6k, and ERK1/2 phosphorylation in aged skeletal muscle. J. Appl. Physiol. 2004, 97, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Xu, S. ERK1/2 MAP kinases in cell survival and apoptosis. IUBMB Life 2006, 58, 621–631. [Google Scholar] [CrossRef]
- Kalous, J.; Tetkova, A.; Kubelka, M.; Susor, A. Importance of ERK1/2 in regulation of protein translation during oocyte meiosis. Int. J. Mol. Sci. 2018, 19, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolch, W. Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat. Rev. Mol. Cell Biol. 2005, 6, 827–837. [Google Scholar] [CrossRef]
- Nishida, E.; Gotoh, Y. The MAP kinase cascade is essential for diverse signal transduction pathways. Trends Biochem. Sci. 1993, 18, 128–131. [Google Scholar] [CrossRef]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef]
- Contesse, T.; Broussot, L.; Fofo, H.; Vanhoutte, P.; Fernandez, S.P.; Barik, J. Dopamine and glutamate receptors control social stress-induced striatal ERK1/2 activation. Neuropharmacology 2021, 190, 108534. [Google Scholar] [CrossRef]
- Amaral, I.M.; Hofer, A.; El Rawas, R. Implication of extracellular signal-regulated kinase in the expression of natural reward: Evidence not found. Front. Behav. Neurosci. 2022, 16, 856675. [Google Scholar] [CrossRef]
- Sharma, A.; Hoeffer, C.A.; Takayasu, Y.; Miyawaki, T.; McBride, S.M.; Klann, E.; Zukin, R.S. Dysregulation of mTOR signaling in fragile X syndrome. J. Neurosci. 2010, 30, 694–702. [Google Scholar] [CrossRef] [Green Version]
- Rehnitz, J.; Messmer, B.; Bender, U.; Nguyen, X.P.; Germeyer, A.; Hinderhofer, K.; Strowitzki, T.; Capp, E. Activation of AKT/mammalian target of rapamycin signaling in the peripheral blood of women with premature ovarian insufficiency and its correlation with FMR1 expression. Reprod. Biol. Endocrinol. 2022, 20, 44. [Google Scholar] [CrossRef]
- Koehl, M.; Ladevèze, E.; Catania, C.; Cota, D.; Abrous, D.N. Inhibition of mTOR signaling by genetic removal of p70 S6 kinase 1 increases anxiety-like behavior in mice. Transl. Psychiatry 2021, 11, 165. [Google Scholar] [CrossRef]
- Urbano-Gámez, J.D.; Casañas, J.J.; Benito, I.; Montesinos, M.L. Prenatal treatment with rapamycin restores enhanced hippocampal mGluR-LTD and mushroom spine size in a Down’s syndrome mouse model. Mol. Brain 2021, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, D.; Popic, J.; Enns, J.P.; Inserra, A.; Skalecka, A.; Markopoulos, A.; Posa, L.; Lopez-Canul, M.; Qianzi, H.; Lafferty, C.K.; et al. Lysergic acid diethylamide (LSD) promotes social behavior through mTORC1 in the excitatory neurotransmission. Proc. Natl. Acad. Sci. USA 2021, 118, e2020705118. [Google Scholar] [CrossRef]
- Maruska, K.P.; Sohn, Y.C.; Fernald, R.D. Mechanistic target of rapamycin (mTOR) implicated in plasticity of the reproductive axis during social status transitions. Gen. Comp. Endocrinol. 2019, 282, 113209. [Google Scholar] [CrossRef]
- Sawicka, K.; Pyronneau, A.; Chao, M.; Bennett, M.V.L.; Zukin, R.S. Elevated ERK/p90 ribosomal S6 kinase activity underlies audiogenic seizure susceptibility in fragile X mice. Proc. Natl. Acad. Sci. USA 2016, 113, E6290–E6297. [Google Scholar] [CrossRef] [Green Version]
- Utami, K.; Yusof, A.; Kwa, J.; Peteri, U.-K.; Castren, M.; Pouladi, M. Elevated de novo protein synthesis in FMRP-deficient human neurons and its correction by metformin treatment. Mol. Autism 2020, 11, 41. [Google Scholar] [CrossRef]
- Brain, P.F. Effects of isolation/grouping on endocrine function and fighting behavior in male and female golden hamsters. (Mesocricetus auratus Waterhouse). Behav. Biol. 1972, 7, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Grelk, D.F.; Papson, B.A.; Cole, J.E.; Rowe, F.A. The influence of caging conditions and hormone treatments on fighting in male and female hamsters. Horm. Behav. 1974, 5, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.P.; Norvelle, A.; Choi, D.C.; Walton, J.C.; Albers, H.E.; Huhman, K.L. Social housing and social isolation: Impact on stress indices and energy balance in male and female Syrian hamsters (Mesocricetus auratus). Physiol. Behav. 2017, 177, 264–269. [Google Scholar] [CrossRef]
- Meisel, R.L.; Sterner, M.R.; Diekman, M.A. Differential hormonal control of aggression and sexual behavior in female Syrian hamsters. Horm. Behav. 1988, 22, 453–466. [Google Scholar] [CrossRef]
- Huhman, K.L.; Solomon, M.B.; Janicki, M.; Harmon, A.C.; Lin, S.M.; E Israel, J.; Jasnow, A.M. Conditioned defeat in male and female Syrian hamsters. Horm. Behav. 2003, 44, 293–299. [Google Scholar] [CrossRef]
- Morin, L.P.; Wood, R.I. A Stereotaxic Atlas of the Golden Hamster Brain; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Patino, C.M.; Ferreira, J.C. Test for trend: Evaluating dose-response effects in association studies. J. Bras. Pneumol. 2016, 42, 240. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Lottes, E.N.; Nanda, S.; Golshir, A.; Patel, A.A.; Ascoli, G.A.; Cox, D.N. PP2A phosphatase regulates cell-type specific cytoskeletal organization to drive dendrite diversity. Front. Mol. Neurosci. 2022, 15, 926567. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Chen, X.C.; Yang, G.Y.; Zhou, L.F. U0126 prevents ERK pathway phosphorylation and interleukin-1beta mRNA production after cerebral ischemia. Chin. Med. Sci. J. 2004, 19, 270–275. [Google Scholar]
- You, Y.; Niu, Y.; Zhang, J.; Huang, S.; Ding, P.; Sun, F.; Wang, X. U0126: Not only a MAPK kinase inhibitor. Front. Pharmacol. 2022, 13, 927083. [Google Scholar] [CrossRef]
- Trepanier, D.J.; Gallant, H.; Legatt, D.F.; Yatscoff, R.W. Rapamycin: Distribution, pharmacokinetics and therapeutic range investigations: An update. Clin. Biochem. 1998, 31, 345–351. [Google Scholar] [CrossRef]
- Hagan, M.J.; Shenkar, R.; Srinath, A.; Romanos, S.G.; Stadnik, A.; Kahn, M.L.; Marchuk, D.A.; Girard, R.; Awad, I.A. Rapamycin in cerebral cavernous malformations: What doses to test in mice and humans. ACS Pharmacol. Transl. Sci. 2022, 5, 266–277. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Concentration | Company | Cat. # |
|---|---|---|---|
| rabbit anti-phospho-p70 S6 Kinase (Thr421/Ser424) | 1:667 | Cell Signaling | 9204S |
| rabbit anti-PP2A C Subunit | 1:10,000 | Cell Signaling | 2038S |
| rabbit anti-phosphop44/42 –MAPK (Erk1/2) (Thr202/Tyr204) (D13.14.4E) XP | 1:667 | Cell Signaling | 4370S |
| rabbit anti-phospho-mTOR (Ser2448) | 1:1000 | Cell Signaling | 2971S |
| rabbit polyclonal to FMRP (phospho S499) | 1:500 | Abcam | ab183319 |
| rabbit anti-FMRP (Ser500) Polyclonal | 1:500 | Bioss Antibodies | BS013188R |
| rabbit anti-p70 S6 Kinase | 1:1000 | Cell Signaling | 9202S |
| rabbit anti-p44/42 MAPK (Erk1/2) | 1:20,000 | Cell Signaling | 9102S |
| rabbit anti-mTOR | 1:10,000 | Cell Signaling | 2983S |
| rabbit anti-FMRP | 1:750 | Cell Signaling | 4317S |
| mouse anti-GAPDH | 1:10,000 | Novus Biologicals LLC | b8245 |
| mouse anti-GAPDH | 1:20,000 | Abcam | ab8245 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borland, J.M.; Dempsey, D.A.; Peyla, A.C.; Hall, M.A.L.; Kohut-Jackson, A.L.; Mermelstein, P.G.; Meisel, R.L. Aggression Results in the Phosphorylation of ERK1/2 in the Nucleus Accumbens and the Dephosphorylation of mTOR in the Medial Prefrontal Cortex in Female Syrian Hamsters. Int. J. Mol. Sci. 2023, 24, 1379. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021379
Borland JM, Dempsey DA, Peyla AC, Hall MAL, Kohut-Jackson AL, Mermelstein PG, Meisel RL. Aggression Results in the Phosphorylation of ERK1/2 in the Nucleus Accumbens and the Dephosphorylation of mTOR in the Medial Prefrontal Cortex in Female Syrian Hamsters. International Journal of Molecular Sciences. 2023; 24(2):1379. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021379
Chicago/Turabian StyleBorland, Johnathan M., Desarae A. Dempsey, Anna C. Peyla, Megan A. L. Hall, Abigail L. Kohut-Jackson, Paul G. Mermelstein, and Robert L. Meisel. 2023. "Aggression Results in the Phosphorylation of ERK1/2 in the Nucleus Accumbens and the Dephosphorylation of mTOR in the Medial Prefrontal Cortex in Female Syrian Hamsters" International Journal of Molecular Sciences 24, no. 2: 1379. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021379