

DBP7 and YRF1-6 Are Involved in Cell Sensitivity to LiCl by Regulating the Translation of PGM2 mRNA
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. DBP7 and YRF1-6 Gene Deletions Diminish Yeast Tolerance to Lithium Chloride
2.2. DBP7 and YRF1-6 Regulate PGM2 Expression at the Translational Level
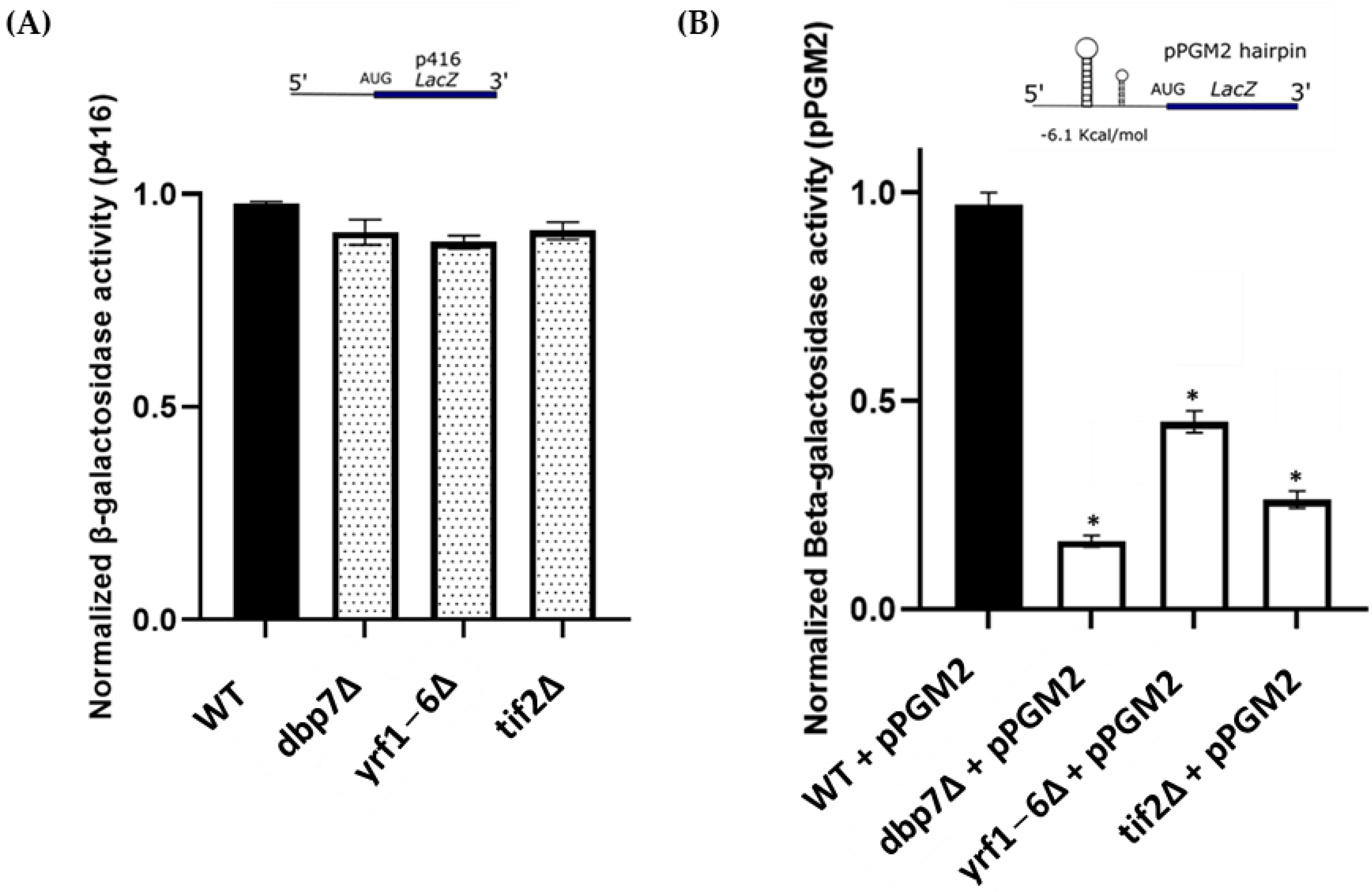
2.3. DBP7 and YRF1-6 Deletion Directly Impact β-Galactosidase Reporter mRNAs with Secondary Structures at 5′-UTR
2.4. Deletion Mutants for DBP7 and YRF1-6 Show Increased Sensitivity to Phenethyl Isothiocyanate
2.5. Genetic Interaction Analysis Connects the Activity of DBP7 and YRF1-6 to Protein Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Drug Sensitivity Analysis
4.3. mRNA Quantification Analysis (qRT-PCR)
4.4. Western Blot Analysis
4.5. Quantitative β-Galactosidase Assay
4.6. Genetic Interaction Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grande, I.; Berk, M.; Birmaher, B.; Vieta, E. Bipolar Disorder. Lancet 2016, 387, 1561–1572. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, A.; Koukopoulos, A.E.; Kotzalidis, G.D.; Janiri, D.; de Chiara, L.; Janiri, L.; Sani, G. Stabilization Beyond Mood: Stabilizing Patients With Bipolar Disorder in the Various Phases of Life. Front. Psychiatry 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Machado-Vieira, R.; Manji, H.K.; Zarate, C.A., Jr. The Role of Lithium in the Treatment of Bipolar Disorder: Convergent Evidence for Neurotrophic Effects as a Unifying Hypothesis. Bipolar Disord. 2009, 11 (Suppl. 2), 92–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.W.; Baek, J.H.; Yang, S.-Y.; Kim, Y.; Cho, Y.; Choi, Y.; Lee, K.; Park, T.; Hong, K.S. Long-Term Response to Mood Stabilizer Treatment and Its Clinical Correlates in Patients with Bipolar Disorders: A Retrospective Observational Study. Int. J Bipolar Disord. 2017, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, P.; Xu, D.; Ji, Y.; Yin, F.; Cui, J.; Su, R.; Wang, Y.; Zhu, Y.; Wei, S.; Lai, J. LiCl Pretreatment Ameliorates Adolescent Methamphetamine Exposure-Induced Long-Term Alterations in Behavior and Hippocampal Ultrastructure in Adulthood in Mice. Int. J. Neuropsychopharmacol. 2019, 22, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Stambolic, V.; Ruel, L.; Woodgett, J.R. Lithium Inhibits Glycogen Synthase Kinase-3 Activity and Mimics Wingless Signalling in Intact Cells. Curr. Biol. 1996, 6, 1664–1669. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Zhang, G.C.; Kong, I.I.; Yun, E.J.; Zheng, J.Q.; Kweon, D.H.; Jin, Y.S. A Mutation in PGM2 Causing Inefficient Galactose Metabolism in the Probiotic Yeast Saccharomyces Boulardii. Appl. Environ. Microbiol. 2021, 84, e02858-17. [Google Scholar] [CrossRef] [Green Version]
- Montero-Lomelí, M.; Morais, B.L.B.; Figueiredo, D.L.; Neto, D.C.S.; Martins, J.R.P.; Masuda, C.A. The Initiation Factor EIF4A Is Involved in the Response to Lithium Stress in Saccharomyces cerevisiae*. J. Biol. Chem. 2002, 277, 21542–21548. [Google Scholar] [CrossRef] [Green Version]
- Masuda, C.A.; Xavier, M.A.; Mattos, K.A.; Galina, A.; Montero-Lomelí, M. Phosphoglucomutase Is an In Vivo Lithium Target in Yeast*. J. Biol. Chem. 2001, 276, 37794–37801. [Google Scholar] [CrossRef]
- Dichtl, B.; Stevens, A.; Tollervey, D. Lithium Toxicity in Yeast Is Due to the Inhibition of RNA Processing Enzymes. EMBO J. 1997, 16, 7184–7195. [Google Scholar] [CrossRef]
- Bergkessel, M.; Whitworth, G.B.; Guthrie, C. Diverse Environmental Stresses Elicit Distinct Responses at the Level of Pre-MRNA Processing in Yeast. RNA 2011, 17, 1461–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, J.A.; Tack, D.C.; Ritchey, L.E.; Gillen, S.L.; Wilczynska, A.; Turro, E.; Bevilacqua, P.C.; Assmann, S.M.; Bushell, M.; le Quesne, J. MRNA Structural Elements Immediately Upstream of the Start Codon Dictate Dependence upon EIF4A Helicase Activity. Genome Biol. 2019, 20, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajikarimlou, M.; Hunt, K.; Kirby, G.; Takallou, S.; Jagadeesan, S.K.; Omidi, K.; Hooshyar, M.; Burnside, D.; Moteshareie, H.; Babu, M.; et al. Lithium Chloride Sensitivity in Yeast and Regulation of Translation. Int. J. Mol. Sci. 2020, 21, 5730. [Google Scholar] [CrossRef] [PubMed]
- Hajikarimlou, M.; Moteshareie, H.; Omidi, K.; Hooshyar, M.; Shaikho, S.; Kazmirchuk, T.; Burnside, D.; Takallou, S.; Zare, N.; Jagadeesan, S.K.; et al. Sensitivity of Yeast to Lithium Chloride Connects the Activity of YTA6 and YPR096C to Translation of Structured MRNAs. PLoS ONE 2020, 15, e0235033. [Google Scholar] [CrossRef]
- Hinnebusch, A.G.; Ivanov, I.P.; Sonenberg, N. Translational Control by 5′-Untranslated Regions of Eukaryotic MRNAs. Science 2016, 352, 1413–1416. [Google Scholar] [CrossRef]
- de Smit, M.H.; van Duin, J. Control of Translation by MRNA Secondary Structure in Escherichia Coli: A Quantitative Analysis of Literature Data. J. Mol. Biol. 1994, 244, 144–150. [Google Scholar] [CrossRef]
- Kozak, M. Downstream Secondary Structure Facilitates Recognition of Initiator Codons by Eukaryotic Ribosomes. Proc. Natl. Acad. Sci. USA 1990, 87, 8301–8305. [Google Scholar] [CrossRef] [Green Version]
- Jagadeesan, S.K.; Al-gafari, M.; Hajikarimlou, M.; Takallou, S.; Moteshareie, H.; Tayabali, A.; Samanfar, B.; Smith, M.; Golshani, A. Lithium Chloride Sensitivity Connects the Activity of PEX11 and RIM20 to the Translation of PGM2 and Other MRNAs with Structured 5′-UTRs. Mol. Cell Biochem. 2022, 477, 2643–2656. [Google Scholar] [CrossRef]
- Kuzmin, E.; VanderSluis, B.; Wang, W.; Tan, G.; Deshpande, R.; Chen, Y.; Usaj, M.; Balint, A.; Mattiazzi Usaj, M.; van Leeuwen, J.; et al. Systematic Analysis of Complex Genetic Interactions. Science (1979) 2018, 360, eaao1729. [Google Scholar] [CrossRef] [Green Version]
- Samanfar, B.; Omidi, K.; Hooshyar, M.; Laliberte, B.; Alamgir, M.D.; Seal, A.J.; Ahmed-Muhsin, E.; Viteri, D.F.; Said, K.; Chalabian, F.; et al. Large-Scale Investigation of Oxygen Response Mutants in Saccharomyces cerevisiae. Mol. Biosyst. 2013, 9, 1351–1359. [Google Scholar] [CrossRef]
- Jagadeesan, S.K.; Potter, T.; Al-gafari, M.; Hooshyar, M.; Hewapathirana, C.M.; Takallou, S.; Hajikarimlou, M.; Burnside, D.; Samanfar, B.; Moteshareie, H.; et al. Discovery and Identification of Genes Involved in DNA Damage Repair in Yeast. Gene 2022, 831, 146549. [Google Scholar] [CrossRef] [PubMed]
- Moteshareie, H.; Hajikarimlou, M.; Mulet Indrayanti, A.; Burnside, D.; Paula Dias, A.; Lettl, C.; Ahmed, D.; Omidi, K.; Kazmirchuk, T.; Puchacz, N.; et al. Heavy Metal Sensitivities of Gene Deletion Strains for ITT1 and RPS1A Connect Their Activities to the Expression of URE2, a Key Gene Involved in Metal Detoxification in Yeast. PLoS ONE 2018, 13, e0198704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Straub, J.; Xiao, D.; Singh, S.v; Yang, H.-S.; Sonenberg, N.; Vatsyayan, J. Phenethyl Isothiocyanate, a Cancer Chemopreventive Constituent of Cruciferous Vegetables, Inhibits Cap-Dependent Translation by Regulating the Level and Phosphorylation of 4E-BP1. Cancer Res. 2007, 67, 3569–3573. [Google Scholar] [CrossRef] [Green Version]
- Syed Alwi, S.S.; Cavell, B.E.; Telang, U.; Morris, M.E.; Parry, B.M.; Packham, G. In Vivo Modulation of 4E Binding Protein 1 (4E-BP1) Phosphorylation by Watercress: A Pilot Study. Br. J. Nutr. 2010, 104, 1288–1296. [Google Scholar] [CrossRef] [Green Version]
- Tong, A.H.Y.; Evangelista, M.; Parsons, A.B.; Xu, H.; Bader, G.D.; Pagé, N.; Robinson, M.; Raghibizadeh, S.; Hogue, C.W.V.; Bussey, H.; et al. Systematic Genetic Analysis with Ordered Arrays of Yeast Deletion Mutants. Science (1979) 2001, 294, 2364–2368. [Google Scholar] [CrossRef] [PubMed]
- Schnier, J.; Schwelberger, H.G.; Smit-McBride, Z.; Kang, H.A.; Hershey, J.W. Translation Initiation Factor 5A and Its Hypusine Modification Are Essential for Cell Viability in the Yeast Saccharomyces cerevisiae. Mol. Cell Biol. 1991, 11, 3105–3114. [Google Scholar] [CrossRef]
- Schuller, A.P.; Wu, C.C.-C.; Dever, T.E.; Buskirk, A.R.; Green, R. EIF5A Functions Globally in Translation Elongation and Termination. Mol. Cell 2017, 66, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Bushman, J.L.; Foiani, M.; Cigan, A.M.; Paddon, C.J.; Hinnebusch, A.G. Guanine Nucleotide Exchange Factor for Eukaryotic Translation Initiation Factor 2 in Saccharomyces cerevisiae: Interactions between the Essential Subunits GCD2, GCD6, and GCD7 and the Regulatory Subunit GCN3. Mol. Cell Biol. 1993, 13, 4618–4631. [Google Scholar] [CrossRef]
- Pavitt, G.D.; Ramaiah, K.v; Kimball, S.R.; Hinnebusch, A.G. EIF2 Independently Binds Two Distinct EIF2B Subcomplexes That Catalyze and Regulate Guanine-Nucleotide Exchange. Genes Dev. 1998, 12, 514–526. [Google Scholar] [CrossRef] [Green Version]
- Luke, B.; Azzalin, C.M.; Hug, N.; Deplazes, A.; Peter, M.; Lingner, J. Saccharomyces cerevisiae Ebs1p Is a Putative Ortholog of Human Smg7 and Promotes Nonsense-Mediated MRNA Decay. Nucleic Acids Res. 2007, 35, 7688–7697. [Google Scholar] [CrossRef]
- Ford, A.S.; Guan, Q.; Neeno-Eckwall, E.; Culbertson, M.R. Ebs1p, a Negative Regulator of Gene Expression Controlled by the Upf Proteins in the Yeast Saccharomyces cerevisiae. Eukaryot. Cell 2006, 5, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischer, T.C.; Weaver, C.M.; McAfee, K.J.; Jennings, J.L.; Link, A.J. Systematic Identification and Functional Screens of Uncharacterized Proteins Associated with Eukaryotic Ribosomal Complexes. Genes Dev. 2006, 20, 1294–1307. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, S.E.; Terenin, I.M.; Andreev, D.E.; Ivanov, P.A.; Dunaevsky, J.E.; Merrick, W.C.; Shatsky, I.N. GTP-Independent TRNA Delivery to the Ribosomal P-Site by a Novel Eukaryotic Translation Factor. J. Biol. Chem. 2010, 285, 26779–26787. [Google Scholar] [CrossRef] [Green Version]
- Mirón-García, M.C.; Garrido-Godino, A.I.; García-Molinero, V.; Hernández-Torres, F.; Rodríguez-Navarro, S.; Navarro, F. The Prefoldin Bud27 Mediates the Assembly of the Eukaryotic RNA Polymerases in an Rpb5-Dependent Manner. PLoS Genet. 2013, 9, e1003297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoll, W.L.; Horton, L.E.; Komar, A.A.; Hensold, J.O.; Merrick, W.C. Characterization of Mammalian EIF2A and Identification of the Yeast Homolog *. J. Biol. Chem. 2002, 277, 37079–37087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougherty, J.D.; Reineke, L.C.; Lloyd, R.E. MRNA Decapping Enzyme 1a (Dcp1a)-Induced Translational Arrest through Protein Kinase R (PKR) Activation Requires the N-Terminal Enabled Vasodilator-Stimulated Protein Homology 1 (EVH1) Domain. J. Biol. Chem. 2014, 289, 3936–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searfoss, A.M.; Wickner, R.B. 3′ Poly(A) Is Dispensable for Translation. Proc. Natl. Acad. Sci. USA 2000, 97, 9133–9137. [Google Scholar] [CrossRef] [Green Version]
- de la Cruz, J.; Kressler, D.; Linder, P. Unwinding RNA in Saccharomyces cerevisiae: DEAD-Box Proteins and Related Families. Trends Biochem. Sci. 1999, 24, 192–198. [Google Scholar] [CrossRef]
- Takizawa, P.A.; Vale, R.D. The Myosin Motor, Myo4p, Binds Ash1 MRNA via the Adapter Protein, She3p. Proc. Natl. Acad. Sci. USA 2000, 97, 5273–5278. [Google Scholar] [CrossRef] [Green Version]
- Long, R.M.; Gu, W.; Lorimer, E.; Singer, R.H.; Chartrand, P. She2p Is a Novel RNA-Binding Protein That Recruits the Myo4p-She3p Complex to ASH1 MRNA. EMBO J. 2000, 19, 6592–6601. [Google Scholar] [CrossRef]
- Li, Z.; Lee, I.; Moradi, E.; Hung, N.-J.; Johnson, A.W.; Marcotte, E.M. Rational Extension of the Ribosome Biogenesis Pathway Using Network-Guided Genetics. PLoS Biol. 2009, 7, e1000213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, W.; Deng, Y.; Zenklusen, D.; Singer, R.H. A New Yeast PUF Family Protein, Puf6p, Represses ASH1 MRNA Translation and Is Required for Its Localization. Genes Dev. 2004, 18, 1452–1465. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, M.; Baryshnikova, A.; Bellay, J.; Kim, Y.; Spear, E.D.; Sevier, C.S.; Ding, H.; Koh, J.L.Y.; Toufighi, K.; Mostafavi, S.; et al. The Genetic Landscape of a Cell. Science 2010, 327, 425–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, E.P.; Kerscher, O. Budding Yeast Protein Extraction and Purification for the Study of Function, Interactions, and Post-Translational Modifications. JoVE J. Vis. Exp. 2013, 80, e50921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omidi, K.; Jessulat, M.; Hooshyar, M.; Burnside, D.; Schoenrock, A.; Kazmirchuk, T.; Hajikarimlou, M.; Daniel, M.; Moteshareie, H.; Bhojoo, U.; et al. Uncharacterized ORF HUR1 Influences the Efficiency of Non-Homologous End-Joining Repair in Saccharomyces cerevisiae. Gene 2018, 639, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Benko, A.L.; Vaduva, G.; Martin, N.C.; Hopper, A.K. Competition between a Sterol Biosynthetic Enzyme and TRNA Modification in Addition to Changes in the Protein Synthesis Machinery Causes Altered Nonsense Suppression. Proc. Natl. Acad. Sci. USA 2000, 97, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Skaar, D.A.; Greenleaf, A.L. The RNA Polymerase II CTD Kinase CTDK-I Affects Pre-MRNA 3′ Cleavage/Polyadenylation through the Processing Component Pti1p. Mol. Cell 2002, 10, 1429–1439. [Google Scholar] [CrossRef]
- Coordes, B.; Brünger, K.M.; Burger, K.; Soufi, B.; Horenk, J.; Eick, D.; Olsen, J.v; Sträßer, K. Ctk1 Function Is Necessary for Full Translation Initiation Activity in Saccharomyces cerevisiae. Eukaryot. Cell 2015, 14, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Röther, S.; Strässer, K. The RNA Polymerase II CTD Kinase Ctk1 Functions in Translation Elongation. Genes Dev. 2007, 21, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, G.P.; Schmelzle, T.; Haghighat, A.; Helliwell, S.B.; Hall, M.N.; Sonenberg, N. Eap1p, a Novel Eukaryotic Translation Initiation Factor 4E-Associated Protein in Saccharomyces cerevisiae. Mol. Cell Biol. 2000, 20, 4604–4613. [Google Scholar] [CrossRef]
- Chial, H.J.; Stemm-Wolf, A.J.; McBratney, S.; Winey, M. Yeast Eap1p, an EIF4E-Associated Protein, Has a Separate Function Involving Genetic Stability. Curr. Biol. 2000, 10, 1519–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewett, N.H.; Goldstrohm, A.C. A Eukaryotic Translation Initiation Factor 4E-Binding Protein Promotes MRNA Decapping and Is Required for PUF Repression. Mol. Cell Biol. 2012, 32, 4181–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neef, D.W.; Thiele, D.J. Enhancer of Decapping Proteins 1 and 2 Are Important for Translation during Heat Stress in Saccharomyces cerevisiae. Mol. Microbiol. 2009, 73, 1032–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunckley, T.; Tucker, M.; Parker, R. Two Related Proteins, Edc1p and Edc2p, Stimulate MRNA Decapping in Saccharomyces cerevisiae. Genetics 2001, 157, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Rutter, J.; Probst, B.L.; McKnight, S.L. Coordinate Regulation of Sugar Flux and Translation by PAS Kinase. Cell 2002, 111, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Röther, S.; Burkert, C.; Brünger, K.M.; Mayer, A.; Kieser, A.; Strässer, K. Nucleocytoplasmic Shuttling of the La Motif-Containing Protein Sro9 Might Link Its Nuclear and Cytoplasmic Functions. RNA 2010, 16, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Sobel, S.G.; Wolin, S.L. Two Yeast La Motif-Containing Proteins Are RNA-Binding Proteins That Associate with Polyribosomes. Mol. Biol. Cell 1999, 10, 3849–3862. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, D.J.; Beggs, J.D. A Suppressor of Yeast Spp81/Ded1 Mutations Encodes a Very Similar Putative ATP-Dependent RNA Helicase. Mol. Microbiol. 1991, 5, 805–812. [Google Scholar] [CrossRef]
- Sen, N.D.; Gupta, N.; Archer, S.K.; Preiss, T.; Lorsch, J.R.; Hinnebusch, A.G. Functional Interplay between DEAD-Box RNA Helicases Ded1 and Dbp1 in Preinitiation Complex Attachment and Scanning on Structured MRNAs In Vivo. Nucleic Acids Res. 2019, 47, 8785–8806. [Google Scholar] [CrossRef]
- Sammons, M.A.; Samir, P.; Link, A.J. Saccharomyces cerevisiae Gis2 Interacts with the Translation Machinery and Is Orthogonal to Myotonic Dystrophy Type 2 Protein ZNF9. Biochem. Biophys. Res. Commun. 2011, 406, 13–19. [Google Scholar] [CrossRef]
- Samanfar, B.; Shostak, K.; Moteshareie, H.; Hajikarimlou, M.; Shaikho, S.; Omidi, K.; Hooshyar, M.; Burnside, D.; Márquez, I.G.; Kazmirchuk, T.; et al. The Sensitivity of the Yeast, Saccharomyces cerevisiae, to Acetic Acid Is Influenced by DOM34 and RPL36A. PeerJ 2017, 5, e4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boone, C.; Bussey, H.; Andrews, B.J. Exploring Genetic Interactions and Networks with Yeast. Nat. Rev. Genet. 2007, 8, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.C.; Smith, A.M.; Sharifpoor, S.; Yan, Z.; Durbic, T.; Heisler, L.E.; Lee, A.Y.; Ryan, O.; Göttert, H.; Surendra, A.; et al. Functional Analysis With a Barcoder Yeast Gene Overexpression System. G3 Genes|Genomes|Genet. 2012, 2, 1279–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Düring, L.; Thorsen, M.; Petersen, D.S.N.; Køster, B.; Jensen, T.H.; Holmberg, S. MRN1 Implicates Chromatin Remodeling Complexes and Architectural Factors in MRNA Maturation. PLoS ONE 2012, 7, e44373. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.J.; Riordan, D.P.; Gerber, A.P.; Herschlag, D.; Brown, P.O. Diverse RNA-Binding Proteins Interact with Functionally Related Sets of RNAs, Suggesting an Extensive Regulatory System. PLoS Biol. 2008, 6, e255. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Ota, K.; Sakaki, Y.; Ito, T. Budding Yeast GCN1 Binds the GI Domain to Activate the EIF2α Kinase GCN2 *. J. Biol. Chem. 2001, 276, 17591–17596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinisch, J.J.; Lorberg, A.; Schmitz, H.-P.; Jacoby, J.J. The Protein Kinase C-Mediated MAP Kinase Pathway Involved in the Maintenance of Cellular Integrity in Saccharomyces cerevisiae. Mol. Microbiol. 1999, 32, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.S.; Munchel, S.E.; Weis, K. The DExD/H Box ATPase Dhh1 Functions in Translational Repression, MRNA Decay, and Processing Body Dynamics. J. Cell Biol. 2011, 194, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Daugeron, M.C.; Linder, P. Dbp7p, a Putative ATP-Dependent RNA Helicase from Saccharomyces cerevisiae, Is Required for 60S Ribosomal Subunit Assembly. RNA 1998, 4, 566–581. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Hayatsu, N.; Matsuura, A.; Ishikawa, F. Y′-Help1, a DNA Helicase Encoded by the Yeast Subtelomeric Y′ Element, Is Induced in Survivors Defective for Telomerase *. J. Biol. Chem. 1998, 273, 33360–33366. [Google Scholar] [CrossRef]
- Niederer, R.O.; Rojas-Duran, M.F.; Zinshteyn, B.; Gilbert, W.v. Direct Analysis of Ribosome Targeting Illuminates Thousand-Fold Regulation of Translation Initiation. Cell Syst. 2022, 13, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Winzeler, E.A.; Shoemaker, D.D.; Astromoff, A.; Liang, H.; Anderson, K.; Andre, B.; Bangham, R.; Benito, R.; Boeke, J.D.; Bussey, H.; et al. Functional Characterization of the S. cerevisiae Genome by Gene Deletion and Parallel Analysis. Science (1979) 1999, 285, 901–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson-Rosenthal, C.; Millar, J.B.A. Cdc25: Mechanisms of Checkpoint Inhibition and Recovery. Trends Cell Biol. 2006, 16, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Krogan, N.J.; Kim, M.; Tong, A.; Golshani, A.; Cagney, G.; Canadien, V.; Richards, D.P.; Beattie, B.K.; Emili, A.; Boone, C.; et al. Methylation of Histone H3 by Set2 in Saccharomyces cerevisiae Is Linked to Transcriptional Elongation by RNA Polymerase II. Mol. Cell Biol. 2003, 23, 4207–4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagih, O.; Usaj, M.; Baryshnikova, A.; VanderSluis, B.; Kuzmin, E.; Costanzo, M.; Myers, C.L.; Andrews, B.J.; Boone, C.M.; Parts, L. SGAtools: One-Stop Analysis and Visualization of Array-Based Genetic Interaction Screens. Nucleic Acids Res. 2013, 41, W591–W596. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jagadeesan, S.K.; Al-gafari, M.; Wang, J.; Takallou, S.; Allard, D.; Hajikarimlou, M.; Kazmirchuk, T.D.D.; Moteshareie, H.; Said, K.B.; Nokhbeh, R.; et al. DBP7 and YRF1-6 Are Involved in Cell Sensitivity to LiCl by Regulating the Translation of PGM2 mRNA. Int. J. Mol. Sci. 2023, 24, 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021785
Jagadeesan SK, Al-gafari M, Wang J, Takallou S, Allard D, Hajikarimlou M, Kazmirchuk TDD, Moteshareie H, Said KB, Nokhbeh R, et al. DBP7 and YRF1-6 Are Involved in Cell Sensitivity to LiCl by Regulating the Translation of PGM2 mRNA. International Journal of Molecular Sciences. 2023; 24(2):1785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021785
Chicago/Turabian StyleJagadeesan, Sasi Kumar, Mustafa Al-gafari, Jiashu Wang, Sarah Takallou, Danielle Allard, Maryam Hajikarimlou, Thomas David Daniel Kazmirchuk, Houman Moteshareie, Kamaledin B. Said, Reza Nokhbeh, and et al. 2023. "DBP7 and YRF1-6 Are Involved in Cell Sensitivity to LiCl by Regulating the Translation of PGM2 mRNA" International Journal of Molecular Sciences 24, no. 2: 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021785