
Non-Coding RNAs in Pulmonary Diseases: Comparison of Different Airway-Derived Biosamples

Abstract
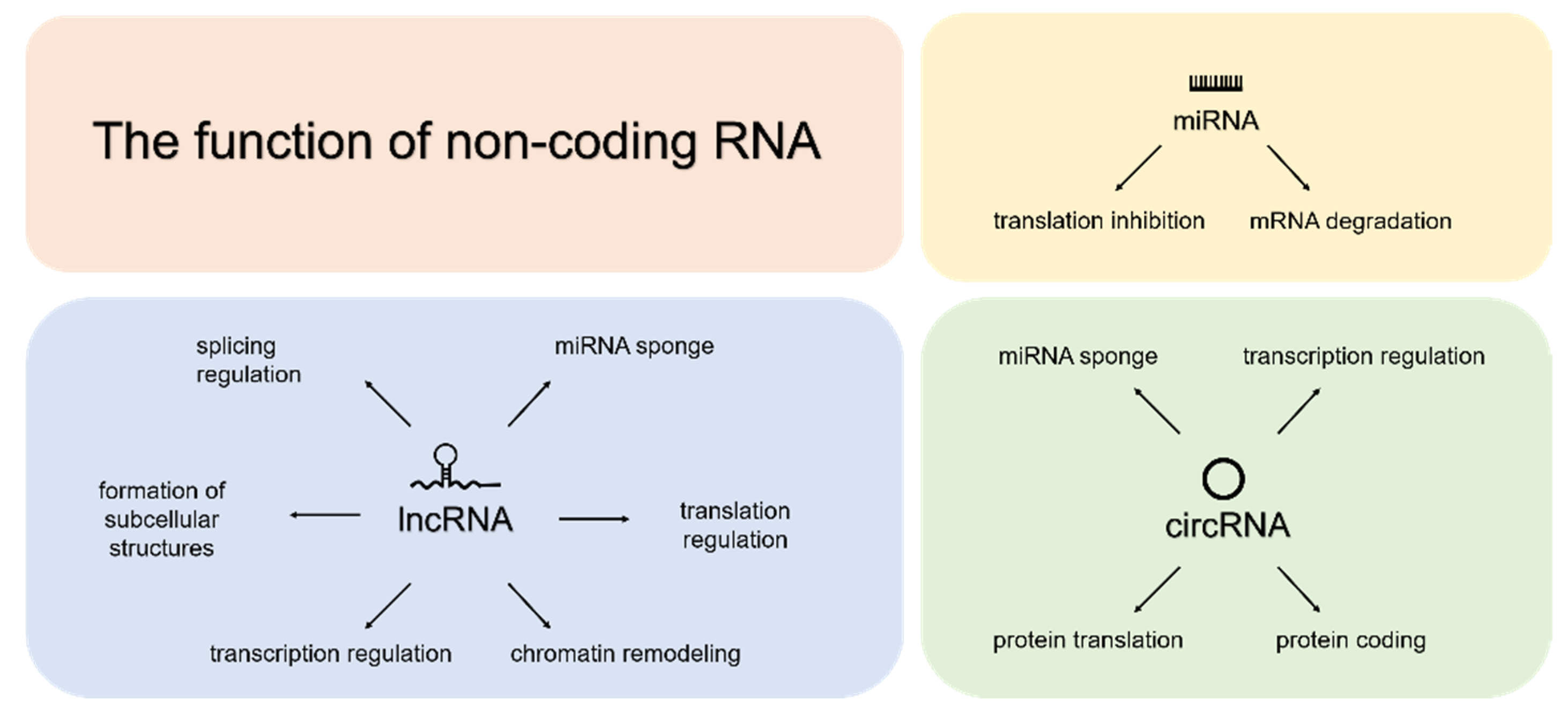
:1. Introduction
2. Exhaled Breath Condensate
2.1. Cystic Fibrosis

{kind=link}
{kind=link}
{kind=link}
| Disease | miRNA | lncRNA | circRNA | Sources | ||
|---|---|---|---|---|---|---|
| Differentiating | Clinically Correlated/Associated | |||||
| (↑) | (↓) | |||||
| Cystic fibrosis | No reports | No reports | miRNA-1247 miRNA-1276 miRNA-449c miRNA-3170 miRNA-432-5p miRNA-548 miR-27b-3p | No reports | No reports | miRNA: [18,19] |
| Asthma | No reports | miR-1248 ### miR-1291 ### let-7a ### miR-133a miR-155 miR-570-3p # | No reports | No reports | No reports | miRNA: [21,22,23,24] |
| Interstitial lung diseases | No reports | miR-29a | No reports | No reports | No reports | miRNA: [25,26] |
2.2. Asthma
2.3. Interstitial Lung Diseases
3. Sputum
3.1. Cystic Fibrosis
3.2. Asthma
3.3. Interstitial Lung Diseases (including ChILD, Sarcoidosis and IPF)
| Disease | miRNA | lncRNA | circRNA | Sources | ||
|---|---|---|---|---|---|---|
| Differentiating | Clinically Correlated/Associated | |||||
| (↑) | (↓) | |||||
| Cystic fibrosis | miR-17 # miR-18a # miR-19a # miR-20a # miR-19b-1 # miR-92a # | No reports | miR-223 miR-451a miR-27b-3p miR-486-5p | No reports | No reports | miRNA: [19,36] |
| Asthma | miR-629-3p miR-223-3p miR-142-3p miR-9 ## miR-145 # miR-338 # | miR-221-3p miR-155 | Nely module: (let-7a-5p let-7d-5p let-7g-5p let-7i-5p miR-15b-5p miR-23a-3p miR-25-3p miR-150-5p miR-181a-5p miR-191-5p miR-223-3p miR-342) miR-199a-5p miR-3935 | lncRNA-NEAT1 | No reports | miRNA: [45,46,47,48,49,50,51,52] lncRNA: [53] |
| Interstitial lung diseases | miR-142-3p miR-33a-5p | let-7d-5p | No reports | No reports | No reports | miRNA: [56,57] |
4. Bronchoalveolar Lavage and Bronchial Brushings
4.1. Cystic Fibrosis
4.2. Asthma
| Disease | miRNA | lncRNA | circRNA | Sources | ||
|---|---|---|---|---|---|---|
| Differentiating | Clinically Correlated/Associated | |||||
| (↑) | (↓) | |||||
| Cystic fibrosis | miR-145 miR-155 miR-101 miR-223 miR-494 miR-221 miR-143-5p | miR-31 miR-126 miR-331-3p miR-362-5p let-7b miR-17 miR-203 | miR-451a miR-663 | XIST TLR8-as1 MALAT1 HOTAIR | No reports | miRNA: [64,65,66,67,68,69,70,71,72,73,74] lncRNA: [75] |
| Asthma | miR-1268 miR-658 let-7a ### let-7b ### let-7c ### miR-26a ### miR146a ### miR-31 ### miR-150 miR-152 miR-375 let-7f miR-487b miR-181c miR-1246 miR-663a miR-1248 miR-1915-3p miR-665 miR-1268a miR-1207-5p miR-221 miR-206 ## | let-7a miR-21 miR-26a miR-99a miR-200c miR-200b miR-203 miR-107 miR-7c miR-31-5p miR-99a-5p miR-29a-3p miR-449a miR-106b-5p miR-19b-3p miR-27b-3p miR-449b-5p miR-200a-3p miR-27a-3p miR-29c-3p miR-34b-5p miR-34c-5p miR-181b-5p miR-218-5p miR-30a-3p miR-146a miR-206 | miR-625-3p miR-202-5p miR-202-3p miR-568 miR-151a-5p miR-615-3p miR-10b-5p miR-151a-3p miR-224-5p miR-581 miR-9-5p miR-148b | ZNF667-AS1 (lncRNA MORT) | circZNF652 hsa_circ_0001585 hsa_circ_0078031 hsa_circ_0000552 | miRNA: [49,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90] lncRNA: [92] circRNA: [62,91] |
| Interstitial lung diseases | miR-150 miR-146a miR-192 miR-221 miR-24 # miR-27a # miR-34a # miR-125b # miR-146b # miR-155 # miR-22-3p miR-320a-3p miR-320b miR-24-3p | miR-202 miR-204 miR-222 miR-15b miR-130b # miR-150 # miR-30a-5p miR-375-3p ### miR-200a-3p miR-200b-3p miR-141-3p miR-423-5p miR-29a miR-185 | miR-155 let-7c miR-146a miR-150 | No reports | No reports | miRNA: [93,94,95,96,97,98,99,100] |
4.3. Interstitial Lung Diseases
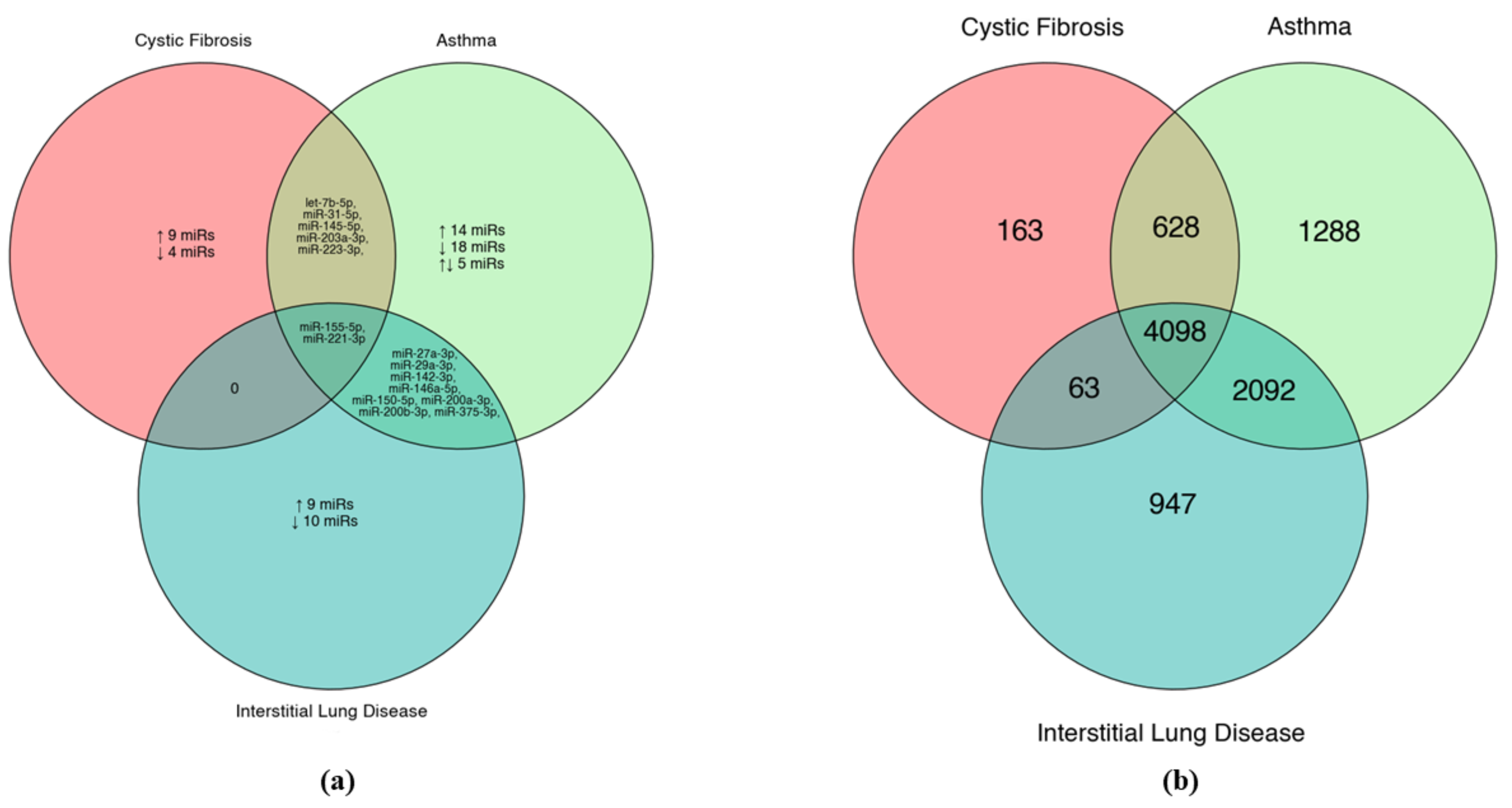
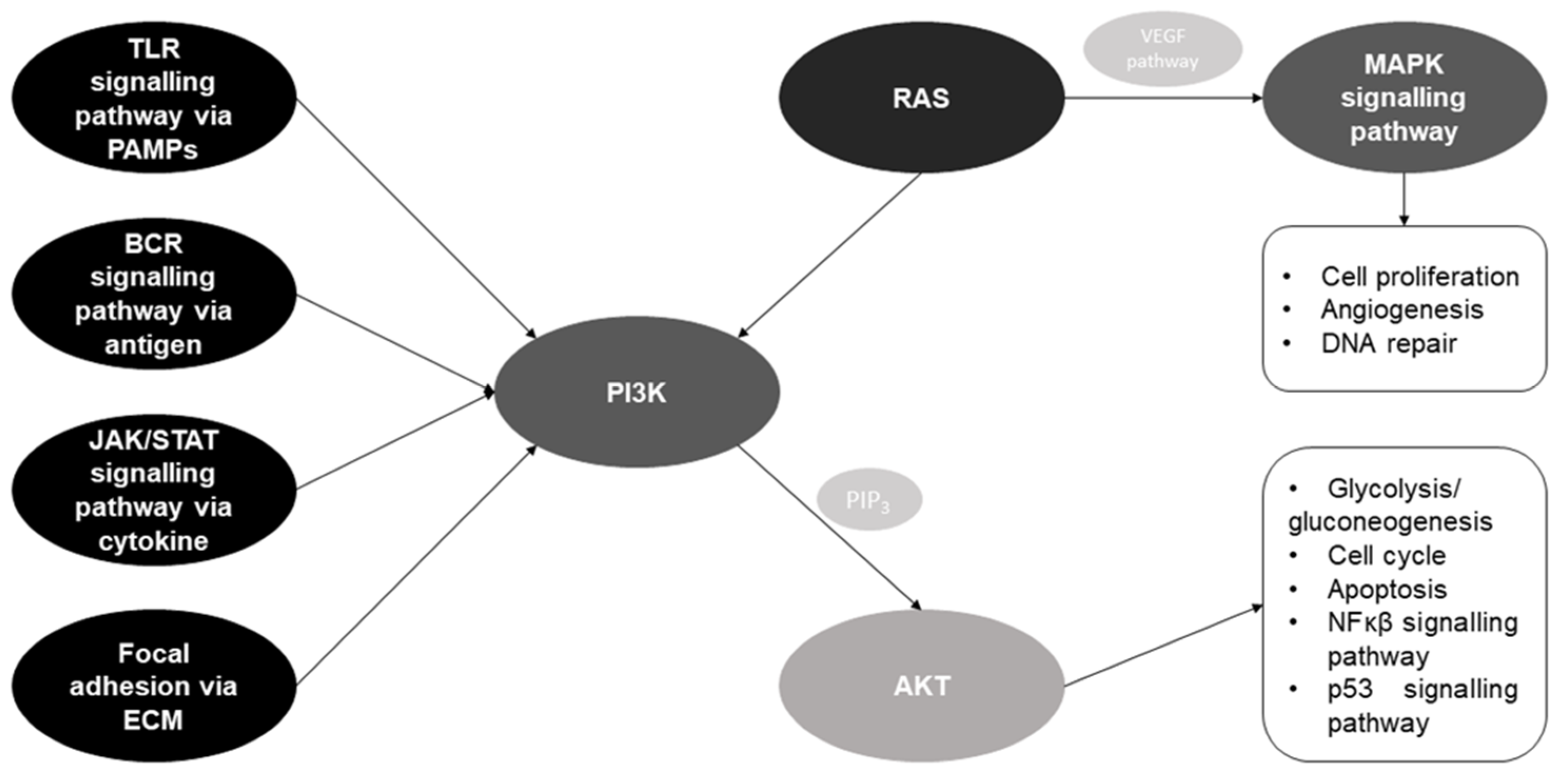
5. In Silico Potential of Non-Coding RNAs in Different Lung Diseases
6. Conclusions and Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Tsagakis, I.; Douka, K.; Birds, I.; Aspden, J.L. Long non-coding RNAs in development and disease: Conservation to mechanisms. J. Pathol. 2020, 250, 480–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef] [Green Version]
- Cantin, A.M.; Hartl, D.; Konstan, M.W.; Chmiel, J.F. Inflammation in cystic fibrosis lung disease: Pathogenesis and therapy. J. Cyst. Fibros. 2015, 14, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, J.M. Treatment of pulmonary exacerbations in cystic fibrosis. Eur. Respir. Rev. 2013, 22, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, M.F.; Tariq, M.; Cataletto, M.E. Asthma. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK430901/ (accessed on 18 August 2022).
- Antoine, M.; Mlika, M. Interstitial Lung Disease. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK541084/ (accessed on 1 August 2022).
- Barratt, S.; Creamer, A.; Hayton, C.; Chaudhuri, N. Idiopathic Pulmonary Fibrosis (IPF): An Overview. J. Clin. Med. 2018, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Horvath, I.; Hunt, J.; Barnes, P.J.; Alving, K.; Antczak, A.; Baraldi, E.; Becher, G.; van Beurden, W.J.; Corradi, M.; Dekhuijzen, R.; et al. Exhaled breath condensate: Methodological recommendations and unresolved questions. Eur. Respir. J. 2005, 26, 523–548. [Google Scholar] [CrossRef]
- Hunt, J. Exhaled Breath Condensate: An Overview. Immunol. Allergy Clin. N. Am. 2007, 27, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, P.; Ishmael, F.T. Exhaled Breath Condensates as a Source for Biomarkers for Characterization of Inflammatory Lung Diseases. J. Anal. Sci. Methods Instrum. 2013, 3, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sánchez, C.; Barbarroja, N.; Pantaleão, L.; López-Sánchez, L.; Ozanne, S.; Jurado-Gámez, B.; Aranda, E.; Lopez-Pedrera, C.; Rodríguez-Ariza, A. Clinical Utility of microRNAs in Exhaled Breath Condensate as Biomarkers for Lung Cancer. J. Pers. Med. 2021, 11, 111. [Google Scholar] [CrossRef]
- Dobhal, G.; Datta, A.; Ayupova, D.; Teesdale-Spittle, P.; Goreham, R.V. Isolation, characterisation and detection of breath-derived extracellular vesicles. Sci. Rep. 2020, 10, 17381. [Google Scholar] [CrossRef] [PubMed]
- Soyer, T.; Birben, E.; Türer, B.; Kahveci, M.; Tuğcu, G.D.; Soyer, U.; Yalçın, E.; Doğru, D.; Özçelik, U.; Kiper, N.; et al. MicroRNA Levels in Exhaled Breath Condensate of Patients with Esophageal Atresia. Dis. Esophagus 2020, 34, doaa082. [Google Scholar] [CrossRef] [PubMed]
- Fesen, K.; Silveyra, P.; Fuentes, N.; Nicoleau, M.; Rivera, L.; Kitch, D.; Graff, G.R.; Siddaiah, R. The role of microRNAs in chronic pseudomonas lung infection in Cystic fibrosis. Respir. Med. 2019, 151, 133–138. [Google Scholar] [CrossRef]
- Stachowiak, Z.; Wojsyk-Banaszak, I.; Jończyk-Potoczna, K.; Narożna, B.; Langwiński, W.; Kycler, Z.; Sobkowiak, P.; Bręborowicz, A.; Szczepankiewicz, A. MiRNA Expression Profile in the Airways Is Altered during Pulmonary Exacerbation in Children with Cystic Fibrosis—A Preliminary Report. J. Clin. Med. 2020, 9, 1887. [Google Scholar] [CrossRef]
- Zeng, X.; Huang, C.; Senavirathna, L.; Wang, P.; Liu, L. miR-27b inhibits fibroblast activation via targeting TGFβ signaling pathway. BMC Cell Biol. 2017, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Pinkerton, M.; Chinchilli, V.; Banta, E.; Craig, T.; August, A.; Bascom, R.; Cantorna, M.; Harvill, E.; Ishmael, F.T. Differential expression of microRNAs in exhaled breath condensates of patients with asthma, patients with chronic obstructive pulmonary disease, and healthy adults. J. Allergy Clin. Immunol. 2013, 132, 217–219.e2. [Google Scholar] [CrossRef]
- Mendes, F.C.; Paciência, I.; Ferreira, A.C.; Martins, C.; Rufo, J.C.; Silva, D.; Cunha, P.; Farraia, M.; Moreira, P.; Delgado, L.; et al. Development and validation of exhaled breath condensate microRNAs to identify and endotype asthma in children. PLoS ONE 2019, 14, e0224983. [Google Scholar] [CrossRef]
- Roff, A.N.; Craig, T.J.; August, A.; Stellato, C.; Ishmael, F.T. MicroRNA-570-3p regulates HuR and cytokine expression in airway epithelial cells. Am. J. Clin. Exp. Immunol. 2014, 3, 68–83. [Google Scholar] [PubMed]
- Mendes, F.C.; Paciência, I.; Rufo, J.C.; Silva, D.; Delgado, L.; Moreira, A.; Moreira, P. Dietary Acid Load Modulation of Asthma-Related miRNAs in the Exhaled Breath Condensate of Children. Nutrients 2022, 14, 1147. [Google Scholar] [CrossRef] [PubMed]
- Ahmadzai, H.; Loke, W.S.J.; Huang, S.; Herbert, C.; Wakefield, D.; Thomas, P. Biomarkers in sarcoidosis: A review. Clin. Med. Insights Pediatr. 2014, 4, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Pattnaik, B.; Sryma, P.; Mittal, S.; Agrawal, A.; Guleria, R.; Madan, K. MicroRNAs in pulmonary sarcoidosis: A systematic review. Respir. Investig. 2020, 58, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Farzan, S. Cough and Sputum Production. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- Lu, J.; Carmody, L.A.; Opron, K.; Simon, R.H.; Kalikin, L.M.; Caverly, L.J.; LiPuma, J.J. Parallel Analysis of Cystic Fibrosis Sputum and Saliva Reveals Overlapping Communities and an Opportunity for Sample Decontamination. Msystems 2020, 5, e00296-20. [Google Scholar] [CrossRef]
- Shen, F.; Sergi, C. Sputum Analysis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK563195/ (accessed on 28 February 2022).
- Jain, V.; Vashisht, R.; Yilmaz, G.; Bhardwaj, A. Pneumonia Pathology. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK526116/ (accessed on 1 August 2022).
- Ho, J.; Marks, G.B.; Fox, G.J. The impact of sputum quality on tuberculosis diagnosis: A systematic review. Int. J. Tuberc. Lung Dis. 2015, 19, 537–544. [Google Scholar] [CrossRef]
- Ronchetti, K.; Tame, J.-D.; Paisey, C.; Thia, L.P.; Doull, I.; Howe, R.; Mahenthiralingam, E.; Forton, J. The CF-Sputum Induction Trial (CF-SpIT) to assess lower airway bacterial sampling in young children with cystic fibrosis: A prospective internally controlled interventional trial. Lancet Respir. Med. 2018, 6, 461–471. [Google Scholar] [CrossRef]
- Grant, L.R.; Hammitt, L.L.; Murdoch, D.R.; O’Brien, K.; Scott, J.A. Procedures for Collection of Induced Sputum Specimens from Children. Clin. Infect. Dis. 2012, 54, S140–S145. [Google Scholar] [CrossRef]
- Pattarayan, D.; Thimmulappa, R.K.; Ravikumar, V.; Rajasekaran, S. Diagnostic Potential of Extracellular MicroRNA in Respiratory Diseases. Clin. Rev. Allergy Immunol. 2016, 54, 480–492. [Google Scholar] [CrossRef]
- Sastre, B.; Cañas, J.A.; Rodrigo-Muñoz, J.M.; Del Pozo, V. Novel Modulators of Asthma and Allergy: Exosomes and MicroRNAs. Front. Immunol. 2017, 8, 826. [Google Scholar] [CrossRef]
- Krause, K.; Kopp, B.T.; Tazi, M.F.; Caution, K.; Hamilton, K.; Badr, A.; Shrestha, C.; Tumin, D.; Hayes, D.; Robledo-Avila, F.; et al. The expression of Mirc1/Mir17–92 cluster in sputum samples correlates with pulmonary exacerbations in cystic fibrosis patients. J. Cyst. Fibros. 2017, 17, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Tazi, M.F.; Dakhlallah, D.A.; Caution, K.; Gerber, M.M.; Chang, S.-W.; Khalil, H.; Kopp, B.T.; Ahmed, A.E.; Krause, K.; Davis, I.; et al. Elevated Mirc1/Mir17-92 cluster expression negatively regulates autophagy and CFTR (cystic fibrosis transmembrane conductance regulator) function in CF macrophages. Autophagy 2016, 12, 2026–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, A.C.; Guo, S.; Huang, L.; Kerem, B.; Oren, Y.S.; Walker, A.J.; Hart, S.L. New approaches to genetic therapies for cystic fibrosis. J. Cyst. Fibros. 2020, 19, S54–S59. [Google Scholar] [CrossRef] [Green Version]
- Tagalakis, A.D.; Munye, M.M.; Ivanova, R.; Chen, H.; Smith, C.M.; Aldossary, A.; Rosa, L.Z.; Moulding, D.; Barnes, J.L.; Kafetzis, K.N.; et al. Effective silencing of ENaC by siRNA delivered with epithelial-targeted nanocomplexes in human cystic fibrosis cells and in mouse lung. Thorax 2018, 73, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Wine, J.J.; Hansson, G.C.; König, P.; Joo, N.S.; Ermund, A.; Pieper, M. Progress in understanding mucus abnormalities in cystic fibrosis airways. J. Cyst. Fibros. 2018, 17, S35–S39. [Google Scholar] [CrossRef] [Green Version]
- Conte, G.; Costabile, G.; Baldassi, D.; Rondelli, V.; Bassi, R.; Colombo, D.; Linardos, G.; Fiscarelli, E.V.; Sorrentino, R.; Miro, A.; et al. Hybrid Lipid/Polymer Nanoparticles to Tackle the Cystic Fibrosis Mucus Barrier in siRNA Delivery to the Lungs: Does PEGylation Make the Difference? ACS Appl. Mater. Interfaces 2022, 14, 7565–7578. [Google Scholar] [CrossRef]
- Ogulur, I.; Pat, Y.; Ardicli, O.; Barletta, E.; Cevhertas, L.; Fernandez-Santamaria, R.; Huang, M.; Imam, M.B.; Koch, J.; Ma, S.; et al. Advances and highlights in biomarkers of allergic diseases. Allergy 2021, 76, 3659–3686. [Google Scholar] [CrossRef]
- Seys, S.F. Role of sputum biomarkers in the management of asthma. Curr. Opin. Pulm. Med. 2017, 23, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez, C.L.; Shaughnessy, T.E.; Matthay, M.A.; Fahy, J.V. Increased Neutrophil Numbers and IL-8 Levels in Airway Secretions in Acute Severe Asthma: Clinical and biologic significance. Am. J. Respir. Crit. Care Med. 2000, 161, 1185–1190. [Google Scholar] [CrossRef]
- Maes, T.; Cobos, F.A.; Schleich, F.; Sorbello, V.; Henket, M.; De Preter, K.; Bracke, K.R.; Conickx, G.; Mesnil, C.; Vandesompele, J.; et al. Asthma inflammatory phenotypes show differential microRNA expression in sputum. J. Allergy Clin. Immunol. 2016, 137, 1433–1446. [Google Scholar] [CrossRef]
- Gomez, J.L.; Chen, A.; Diaz, M.P.; Zirn, N.; Gupta, A.; Britto, C.; Sauler, M.; Yan, X.; Stewart, E.; Santerian, K.; et al. A Network of Sputum MicroRNAs Is Associated with Neutrophilic Airway Inflammation in Asthma. Am. J. Respir. Crit. Care Med. 2020, 202, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, S.; Fang, X.; Qin, L.; Fan, Y.; Ding, D.; Liu, X.; Xie, M. Plasma miR-199a-5p is increased in neutrophilic phenotype asthma patients and negatively correlated with pulmonary function. PLoS ONE 2018, 13, e0193502. [Google Scholar] [CrossRef]
- Li, J.J.; Tay, H.L.; Maltby, S.; Xiang, Y.; Eyers, F.; Hatchwell, L.; Zhou, H.; Toop, H.D.; Morris, J.C.; Nair, P.; et al. MicroRNA-9 regulates steroid-resistant airway hyperresponsiveness by reducing protein phosphatase 2A activity. J. Allergy Clin. Immunol. 2015, 136, 462–473. [Google Scholar] [CrossRef]
- Zhang, K.; Liang, Y.; Feng, Y.; Wu, W.; Zhang, H.; He, J.; Hu, Q.; Zhao, J.; Xu, Y.; Liu, Z.; et al. Decreased epithelial and sputum miR-221-3p associates with airway eosinophilic inflammation and CXCL17 expression in asthma. Am. J. Physiol. Cell. Mol. Physiol. 2018, 315, L253–L264. [Google Scholar] [CrossRef] [PubMed]
- Lacedonia, D.; Palladino, G.P.; Barbaro, M.P.F.; Scioscia, G.; Carpagnano, G.E. Expression profiling of miRNA-145 and miRNA-338 in serum and sputum of patients with COPD, asthma, and asthma–COPD overlap syndrome phenotype. Int. J. Chronic Obstr. Pulm. Dis. 2017, 12, 1811–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmhäll, C.; Johansson, K.; Winkler, C.; Alawieh, S.; Ekerljung, L.; Rådinger, M. Altered miR-155 Expression in Allergic Asthmatic Airways. Scand. J. Immunol. 2017, 85, 300–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakwerth, C.A.; Chaker, A.M.; Guerth, F.; Oelsner, M.; Pechtold, L.; Bonsen, L.S.Z.; Ullmann, J.T.; Krauss-Etschmann, S.; Erb, A.; Kau, J.; et al. Sputum microRNA-screening reveals Prostaglandin EP3 receptor as selective target in allergen-specific immunotherapy. Clin. Exp. Allergy 2021, 51, 1577–1591. [Google Scholar] [CrossRef]
- Song, D.; Jiang, Y.; Zhao, Q.; Li, J.; Zhao, Y. lncRNA-NEAT1 Sponges miR-128 to Promote Inflammatory Reaction and Phenotypic Transformation of Airway Smooth Muscle Cells. Comput. Math. Methods Med. 2022, 2022, 7499911. [Google Scholar] [CrossRef]
- Li, X.; Ye, S.; Lu, Y. Long non-coding RNA NEAT1 overexpression associates with increased exacerbation risk, severity, and inflammation, as well as decreased lung function through the interaction with microRNA-124 in asthma. J. Clin. Lab. Anal. 2019, 34, e23023. [Google Scholar] [CrossRef] [Green Version]
- Antoniou, K.M.; Alexandrakis, M.; Tzanakis, N.; Tsiligianni, I.; Tzortzaki, E.G.; Siafakas, N.M.; Bouros, D.E. Induced Sputum versus Bronchoalveolar Lavage Fluid in the Evaluation of Patients with Idiopathic Pulmonary Fibrosis. Respiration 2005, 72, 32–38. [Google Scholar] [CrossRef]
- Njock, M.-S.; Guiot, J.; Henket, M.A.; Nivelles, O.; Thiry, M.; Dequiedt, F.; Corhay, J.-L.; Louis, R.E.; Struman, I. Sputum exosomes: Promising biomarkers for idiopathic pulmonary fibrosis. Thorax 2018, 74, 309–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiot, J.; Cambier, M.; Boeckx, A.; Henket, M.; Nivelles, O.; Gester, F.; Louis, E.; Malaise, M.; Dequiedt, F.; Louis, R.; et al. Macrophage-derived exosomes attenuate fibrosis in airway epithelial cells through delivery of antifibrotic miR-142-3p. Thorax 2020, 75, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Connett, G. Bronchoalveolar lavage. Paediatr. Respir. Rev. 2000, 1, 52–56. [Google Scholar] [CrossRef]
- Patel, P.H.; Antoine, M.; Ullah, S. Bronchoalveolar Lavage. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK430762/ (accessed on 28 August 2022).
- Liu, Z.; Yan, J.; Tong, L.; Liu, S.; Zhang, Y. The role of exosomes from BALF in lung disease. J. Cell. Physiol. 2021, 237, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Pavel, A.B.; Campbell, J.D.; Liu, G.; Elashoff, D.; Dubinett, S.; Smith, K.; Whitney, D.; Lenburg, M.E.; Spira, A. Alterations in Bronchial Airway miRNA Expression for Lung Cancer Detection. Cancer Prev. Res. 2017, 10, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wu, W.; Yi, L.; Feng, Y.; Chang, C.; Chen, S.; Gao, J.; Chen, G.; Zhen, G. A Potential circRNA-miRNA-mRNA Regulatory Network in Asthmatic Airway Epithelial Cells Identified by Integrated Analysis of Microarray Datasets. Front. Mol. Biosci. 2021, 8, 703307. [Google Scholar] [CrossRef]
- Guthrie, J. General Considerations when Dealing with Biological Fluid Samples. In Comprehensive Sampling and Sample Preparation; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1–19. [Google Scholar]
- Kabir, F.L.; Ambalavanan, N.; Liu, G.; Li, P.; Solomon, G.M.; Lal, C.V.; Mazur, M.; Halloran, B.; Szul, T.; Gerthoffer, W.T.; et al. MicroRNA-145 Antagonism Reverses TGF-β Inhibition of F508del CFTR Correction in Airway Epithelia. Am. J. Respir. Crit. Care Med. 2018, 197, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Harris, W.T.; Kortyka, S.; Kotha, K.; Ostmann, A.J.; Rezayat, A.; Sridharan, A.; Sanders, Y.; Naren, A.P.; Clancy, J.P. TGF-Beta Downregulation of Distinct Chloride Channels in Cystic Fibrosis-Affected Epithelia. PLoS ONE 2014, 9, e106842. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Nymon, A.B.; Ringelberg, C.S.; Lesseur, C.; Hazlett, H.F.; Howard, L.; Marsit, C.J.; Ashare, A. Pulmonary microRNA profiling: Implications in upper lobe predominant lung disease. Clin. Epigenetics 2017, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Oglesby, I.K.; Vencken, S.F.; Agrawal, R.; Gaughan, K.; Molloy, K.; Higgins, G.; McNally, P.; McElvaney, N.G.; Mall, M.A.; Greene, C.M. miR-17 overexpression in cystic fibrosis airway epithelial cells decreases interleukin-8 production. Eur. Respir. J. 2015, 46, 1350–1360. [Google Scholar] [CrossRef]
- De Santi, C.; Gadi, S.; Swiatecka-Urban, A.; Greene, C.M. Identification of a novel functional miR-143-5p recognition element in the Cystic Fibrosis Transmembrane Conductance Regulator 3’UTR. AIMS Genet. 2018, 05, 053–062. [Google Scholar] [CrossRef]
- Weldon, S.; McNally, P.; McAuley, D.F.; Oglesby, I.K.; Wohlford-Lenane, C.L.; Bartlett, J.A.; Scott, C.J.; McElvaney, N.G.; Greene, C.M.; McCray, P.B.; et al. miR-31 Dysregulation in Cystic Fibrosis Airways Contributes to Increased Pulmonary Cathepsin S Production. Am. J. Respir. Crit. Care Med. 2014, 190, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Oglesby, I.K.; Bray, I.M.; Chotirmall, S.H.; Stallings, R.L.; O’Neill, S.J.; McElvaney, N.G.; Greene, C.M. miR-126 Is Downregulated in Cystic Fibrosis Airway Epithelial Cells and Regulates TOM1 Expression. J. Immunol. 2010, 184, 1702–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, S.; Balakathiresan, N.S.; Dalgard, C.; Gutti, U.; Armistead, D.; Jozwik, C.; Srivastava, M.; Pollard, H.B.; Biswas, R. Elevated miR-155 Promotes Inflammation in Cystic Fibrosis by Driving Hyperexpression of Interleukin-8. J. Biol. Chem. 2011, 286, 11604–11615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oglesby, I.K.; Chotirmall, S.H.; McElvaney, N.G.; Greene, C.M. Regulation of Cystic Fibrosis Transmembrane Conductance Regulator by MicroRNA-145, -223, and -494 Is Altered in ΔF508 Cystic Fibrosis Airway Epithelium. J. Immunol. 2013, 190, 3354–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oglesby, I.K.; Agrawal, R.; A Mall, M.; McElvaney, N.G.; Greene, C.M. miRNA-221 is elevated in cystic fibrosis airway epithelial cells and regulates expression of ATF6. Mol. Cell. Pediatr. 2015, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- De Santi, C.; Fernández, E.F.; Gaul, R.; Vencken, S.; Glasgow, A.; Oglesby, I.K.; Hurley, K.; Hawkins, F.; Mitash, N.; Mu, F.; et al. Precise Targeting of miRNA Sites Restores CFTR Activity in CF Bronchial Epithelial Cells. Mol. Ther. 2020, 28, 1190–1199. [Google Scholar] [CrossRef]
- McKiernan, P.J.; Molloy, K.; Cryan, S.A.; McElvaney, N.G.; Greene, C.M. Long noncoding RNA are aberrantly expressed in vivo in the cystic fibrosis bronchial epithelium. Int. J. Biochem. Cell Biol. 2014, 52, 184–191. [Google Scholar] [CrossRef]
- Levänen, B.; Bhakta, N.R.; Paredes, P.T.; Barbeau, R.; Hiltbrunner, S.; Pollack, J.L.; Sköld, C.M.; Svartengren, M.; Grunewald, J.; Gabrielsson, S.; et al. Altered microRNA profiles in bronchoalveolar lavage fluid exosomes in asthmatic patients. J. Allergy Clin. Immunol. 2013, 131, 894–903.e8. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Wu, F.; Fan, J.; Jin, Y.; Wang, J.; Yang, G. Posttranscriptional Regulation of Interleukin-33 Expression by MicroRNA-200 in Bronchial Asthma. Mol. Ther. 2018, 26, 1808–1817. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Li, C.-X.; Tang, L.-F. The Differential Expression Profiles of miRNA-let 7a, 7b, and 7c in Bronchoalveolar Lavage Fluid from Infants with Asthma and Airway Foreign Bodies. J. Evid. Based Integr. Med. 2019, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.G.; Sun, Y.; Wang, K.S.; Jia, J.D.; Yang, J.; Li, Y.N. Effects of mir-26a/mir-146a/miR-31 on airway inflammation of asthma mice and asthma children. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5432–5440. [Google Scholar] [CrossRef] [PubMed]
- Francisco-Garcia, A.S.; Garrido-Martín, E.M.; Rupani, H.; Lau, L.C.K.; Martinez-Nunez, R.T.; Howarth, P.H.; Sanchez-Elsner, T. Small RNA Species and microRNA Profiles are Altered in Severe Asthma Nanovesicles from Broncho Alveolar Lavage and Associate with Impaired Lung Function and Inflammation. Non-Coding RNA 2019, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupani, H.; Martinez-Nunez, R.T.; Dennison, P.; Lau, L.C.K.; Jayasekera, N.; Havelock, T.; Francisco-Garcia, A.S.; Grainge, C.; Howarth, P.H.; Sanchez-Elsner, T. Toll-like Receptor 7 Is Reduced in Severe Asthma and Linked to an Altered MicroRNA Profile. Am. J. Respir. Crit. Care Med. 2016, 194, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jardim, M.J.; Dailey, L.; Silbajoris, R.; Diaz-Sanchez, D. Distinct MicroRNA Expression in Human Airway Cells of Asthmatic Donors Identifies a Novel Asthma-Associated Gene. Am. J. Respir. Cell Mol. Biol. 2012, 47, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Solberg, O.D.; Ostrin, E.J.; Love, M.I.; Peng, J.C.; Bhakta, N.R.; Hou, L.; Nguyen, C.; Solon, M.; Nguyen, C.; Barczak, A.J.; et al. Airway Epithelial miRNA Expression Is Altered in Asthma. Am. J. Respir. Crit. Care Med. 2012, 186, 965–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, X.; Zhang, K.; Yi, L.; Mo, Y.; Liang, Y.; Zhao, J.; Zhang, Z.; Xu, Y.; Zhen, G. Decreased epithelial and plasma miR-181b-5p expression associates with airway eosinophilic inflammation in asthma. Clin. Exp. Allergy 2016, 46, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Feng, Y.; Wu, W.; Chang, C.; Chen, D.; Chen, S.; Zhen, G. microRNA-218-5p plays a protective role in eosinophilic airway inflammation via targeting δ-catenin, a novel catenin in asthma. Clin. Exp. Allergy 2019, 50, 29–40. [Google Scholar] [CrossRef]
- Wu, W.; Gao, J.; Chen, D.; Chen, G.; Feng, Y.; Chang, C.; Chen, S.; Yi, L.; Zhen, G. Epithelial microRNA-30a-3p targets RUNX2/HMGB1 axis to suppress airway eosinophilic inflammation in asthma. Respir. Res. 2022, 23, 17. [Google Scholar] [CrossRef]
- Kivihall, A.; Aab, A.; Soja, J.; Sładek, K.; Sanak, M.; Altraja, A.; Jakiela, B.; Bochenek, G.; Rebane, A. Reduced expression of miR-146a in human bronchial epithelial cells alters neutrophil migration. Clin. Transl. Allergy 2019, 9, 62. [Google Scholar] [CrossRef]
- Zhang, H.; Song, Y.; Rong, W.; Fan, L.; Cai, Y.; Qu, Q.; Gao, Y.; Zhao, H. miR-221 participates in the airway epithelial cells injury in asthma via targeting SIRT1. Exp. Lung Res. 2018, 44, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Nicodemus-Johnson, J.; Laxman, B.; Stern, R.K.; Sudi, J.; Tierney, C.N.; Norwick, L.; Hogarth, D.K.; McConville, J.F.; Naureckas, E.T.; Sperling, A.I.; et al. Maternal asthma and microRNA regulation of soluble HLA-G in the airway. J. Allergy Clin. Immunol. 2013, 131, 1496–1503.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Feng, Y.; Liang, Y.; Wu, W.; Chang, C.; Chen, D.; Chen, S.; Gao, J.; Chen, G.; Yi, L.; et al. Epithelial microRNA-206 targets CD39/extracellular ATP to upregulate airway IL-25 and TSLP in type 2-high asthma. JCI Insight. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, C.; Cai, Y.; Zou, X.; Chao, Y.; Yan, Z.; Zou, C.; Wu, X.; Tang, L. CircZNF652 promotes the goblet cell metaplasia by targeting the miR-452-5p/JAK2 signaling pathway in allergic airway epithelia. J. Allergy Clin. Immunol. 2022, 150, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Jiang, H.; Jiang, N.; Gao, J. Integrative analysis of the contribution of mRNAs and long non-coding RNAs to the pathogenesis of asthma. Mol. Med. Rep. 2019, 20, 2617–2624. [Google Scholar] [CrossRef]
- Dyskova, T.; Fillerova, R.; Novosad, T.; Kudelka, M.; Zurkova, M.; Gajdos, P.; Kolek, V.; Kriegova, E. Correlation Network Analysis Reveals Relationships between MicroRNAs, Transcription FactorT-bet, and Deregulated Cytokine/Chemokine-Receptor Network in Pulmonary Sarcoidosis. Mediat. Inflamm. 2015, 2015, 121378. [Google Scholar] [CrossRef] [Green Version]
- Kiszałkiewicz, J.; Piotrowski, W.J.; Pastuszak-Lewandoska, D.; Górski, P.; Antczak, A.; Górski, W.; Domańska-Senderowska, D.; Migdalska-Sęk, M.; Czarnecka, K.H.; Nawrot, E.; et al. Altered miRNA expression in pulmonary sarcoidosis. BMC Med. Genet. 2016, 17, 2. [Google Scholar] [CrossRef] [Green Version]
- Kishore, A.; Navratilova, Z.; Kolek, V.; Novosadova, E.; Čépe, K.; Du Bois, R.M.; Petrek, M. Expression analysis of extracellular microRNA in bronchoalveolar lavage fluid from patients with pulmonary sarcoidosis. Respirology 2018, 23, 1166–1172. [Google Scholar] [CrossRef]
- Kachamakova-Trojanowska, N.; Jazwa-Kusior, A.; Szade, K.; Kasper, L.; Soja, J.; Andrychiewicz, A.; Jakiela, B.; Plutecka, H.; Sanak, M.; Jozkowicz, A.; et al. Molecular profiling of regulatory T cells in pulmonary sarcoidosis. J. Autoimmun. 2018, 94, 56–69. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, T.; Hu, X.; Liu, Z.; Zhao, L.; Liu, H.; Liu, Z.; Ma, L. Downregulation of microRNA-30a in bronchoalveolar lavage fluid from idiopathic pulmonary fibrosis patients. Mol. Med. Rep. 2018, 18, 5799–5806. [Google Scholar] [CrossRef]
- Kaur, G.; Maremanda, K.P.; Campos, M.; Chand, H.S.; Li, F.; Hirani, N.; Haseeb, M.A.; Li, D.; Rahman, I. Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients. Int. J. Mol. Sci. 2021, 22, 11830. [Google Scholar] [CrossRef] [PubMed]
- Tsitoura, E.; Wells, A.U.; Karagiannis, K.; Lasithiotaki, I.; Vasarmidi, E.; Bibaki, E.; Koutoulaki, C.; Sato, H.; Spandidos, D.A.; Siafakas, N.M.; et al. MiR-185/AKT and miR-29a/Collagen 1a pathways are activated in IPF BAL cells. Oncotarget 2016, 7, 74569–74581. [Google Scholar] [CrossRef] [Green Version]
- Bibaki, E.; Tsitoura, E.; Vasarmidi, E.; Margaritopoulos, G.; Trachalaki, A.; Koutoulaki, C.; Georgopoulou, T.; Spandidos, D.A.; Tzanakis, N.; Antoniou, K.M. miR-185 and miR-29a are similarly expressed in the bronchoalveolar lavage cells in IPF and lung cancer but common targets DNMT1 and COL1A1 show disease specific patterns. Mol. Med. Rep. 2018, 17, 7105–7112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Hu, K.; Cai, X.; Yang, B.; He, Q.; Wang, J.; Weng, Q. Targeting PI3K/AKT signaling for treatment of idiopathic pulmonary fibrosis. Acta Pharm. Sin. B 2021, 12, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Gutti, U.; Mercado, J.; Moore, C.; Pollard, H.B.; Biswas, R. MAPK signaling pathways regulate IL-8 mRNA stability and IL-8 protein expression in cystic fibrosis lung epithelial cell lines. Am. J. Physiol. Cell. Mol. Physiol. 2011, 300, L81–L87. [Google Scholar] [CrossRef]
- Tezuka, T.; Azuma, M.; Ogawa, H.; Kondo, M.; Uehara, H.; Aono, Y.; Hanibuchi, M.; Nishioka, Y. A RAS inhibitor reduces allergic airway remodeling via regulating IL-33-derived type 2 innate lymphoid cells. Exp. Lung Res. 2021, 47, 451–463. [Google Scholar] [CrossRef]
- Xiang, S. Endocytosis and human innate immunity. J. Immunol. Sci. 2018, 2, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Barratt, S.L.; Flower, V.A.; Pauling, J.D.; Millar, A.B. VEGF (Vascular Endothelial Growth Factor) and Fibrotic Lung Disease. Int. J. Mol. Sci. 2018, 19, 1269. [Google Scholar] [CrossRef] [Green Version]
- Amano, H.; Matsui, Y.; Hatanaka, K.; Hosono, K.; Ito, Y. VEGFR1-tyrosine kinase signaling in pulmonary fibrosis. Inflamm. Regen. 2021, 41, 16. [Google Scholar] [CrossRef]
- Muzanyi, G.; Peace, A.; Wamuntu, B.; Joseph, A.; Nassali, J. The threat of persistent bacteria and fungi contamination in tuberculosis sputum cultures. Afr. Health Sci. 2021, 21, 628–632. [Google Scholar] [CrossRef]
- Paska, C.; Barta, I.; Drozdovszky, O.; Antus, B. Elimination of bacterial DNA during RNA isolation from sputum: Bashing bead vortexing is preferable over prolonged DNase treatment. PLoS ONE 2019, 14, e0214609. [Google Scholar] [CrossRef] [PubMed]



| EBC | SPUTUM | BALF/BB | |
|---|---|---|---|
| Invasiveness | None | Low (higher with induction) | High |
| Risk of bacterial contamination | Medium | High | Medium/high (BALF) |
| RNA abundance | Very low | Low/medium | High |
| Equipment and staff | Dedicated collection, devices and trained personnel | Collection pan and trained personnel (induction) | Specialised equipment (bronchoscope) and highly trained personnel |
| Time of sample collection | 10–30 min | Depending on the expectoration capacity | Around 2 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stachowiak, Z.; Narożna, B.; Szczepankiewicz, A. Non-Coding RNAs in Pulmonary Diseases: Comparison of Different Airway-Derived Biosamples. Int. J. Mol. Sci. 2023, 24, 2006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032006
Stachowiak Z, Narożna B, Szczepankiewicz A. Non-Coding RNAs in Pulmonary Diseases: Comparison of Different Airway-Derived Biosamples. International Journal of Molecular Sciences. 2023; 24(3):2006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032006
Chicago/Turabian StyleStachowiak, Zuzanna, Beata Narożna, and Aleksandra Szczepankiewicz. 2023. "Non-Coding RNAs in Pulmonary Diseases: Comparison of Different Airway-Derived Biosamples" International Journal of Molecular Sciences 24, no. 3: 2006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032006