
miR-383-5p, miR-181a-5p, and miR-181b-5p as Predictors of Response to First-Generation Somatostatin Receptor Ligands in Acromegaly
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
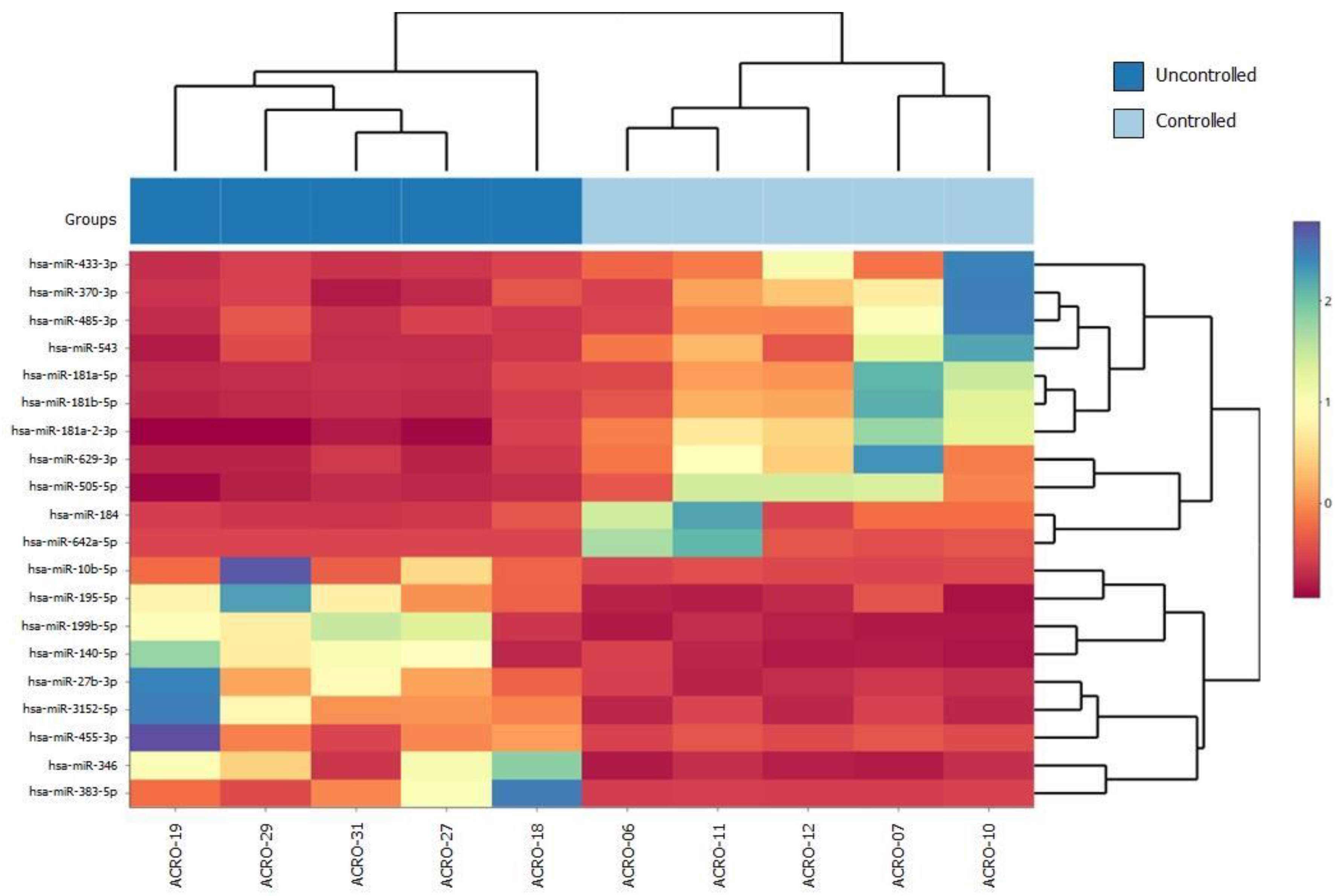
2.1. Cohort Selection and Differentially Expressed miRNAs in Uncontrolled Patients with fg-SRL Treatment
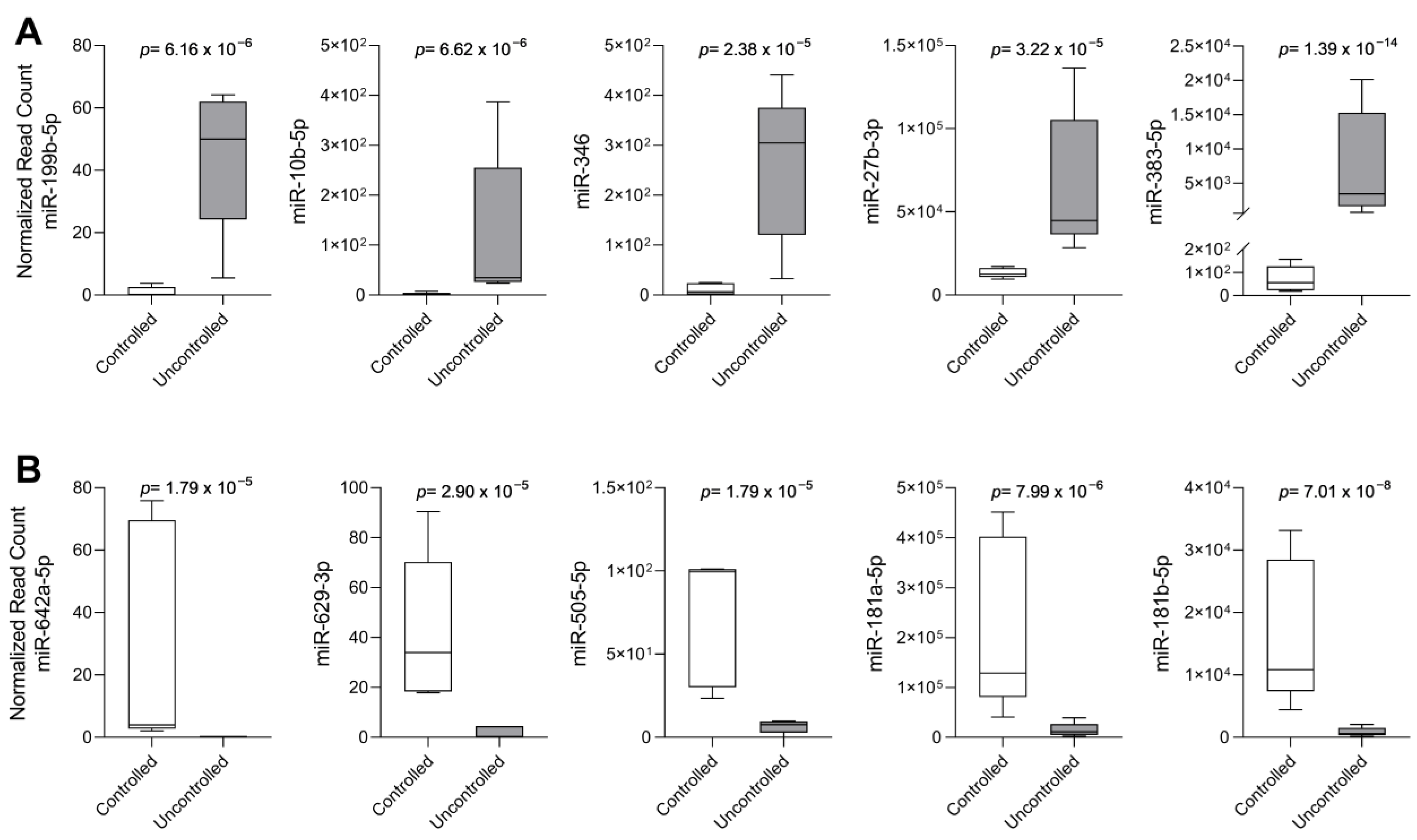
2.2. Analysis of the Top 10 Differentially Expressed miRNAs by qPCR
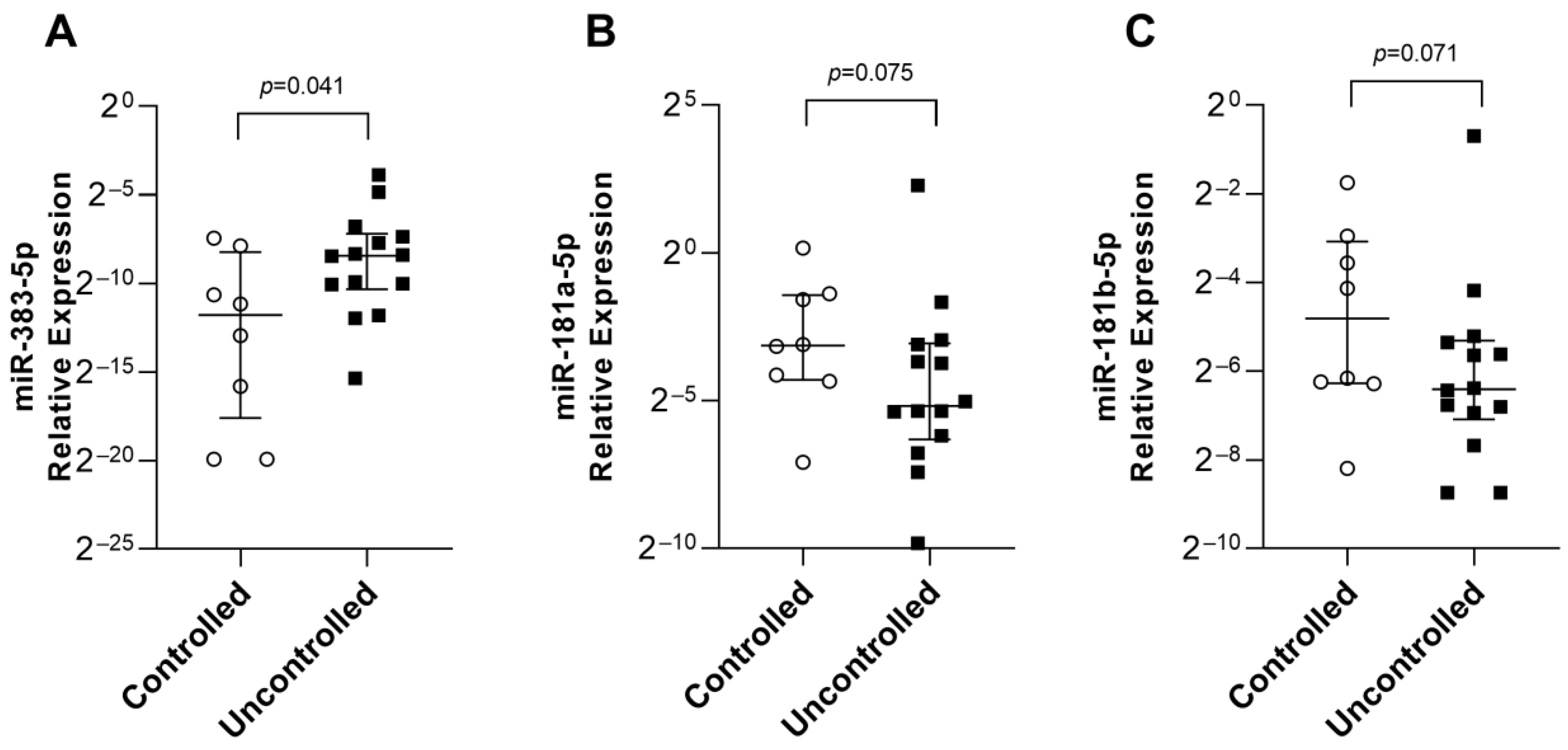
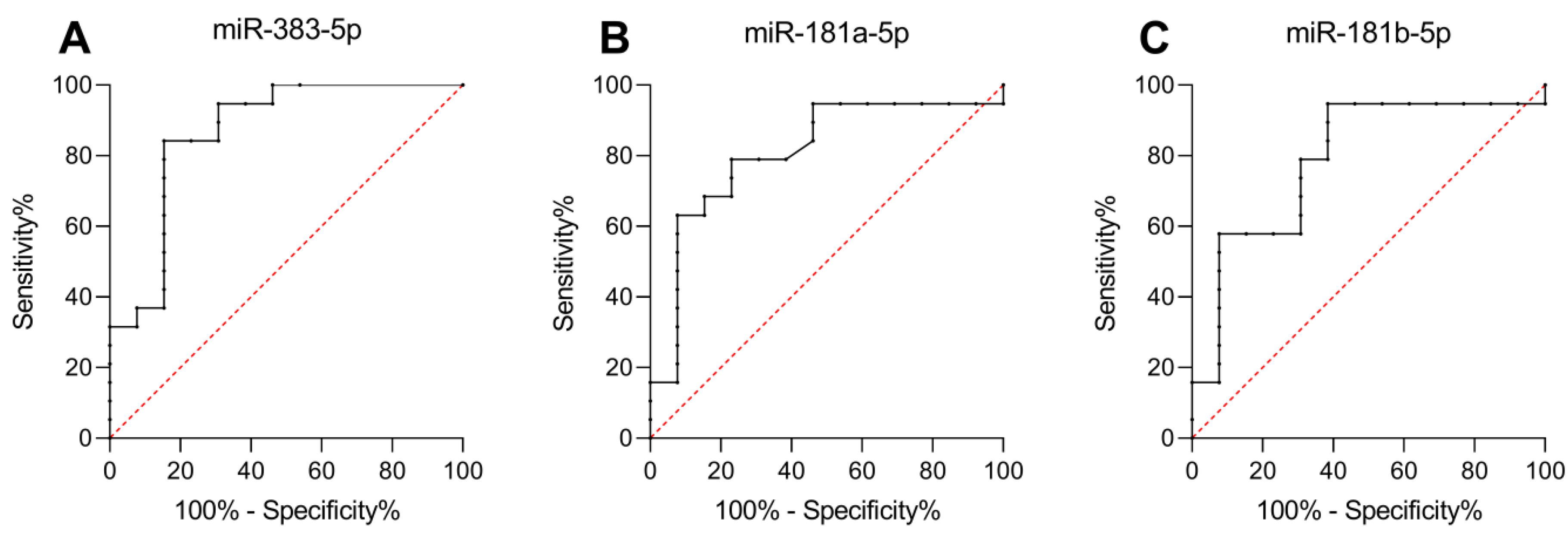
2.3. miR-383-5p, miR-181a-5p, and miR-181b-5p as Predictors of Response to fg-SRL Treatment
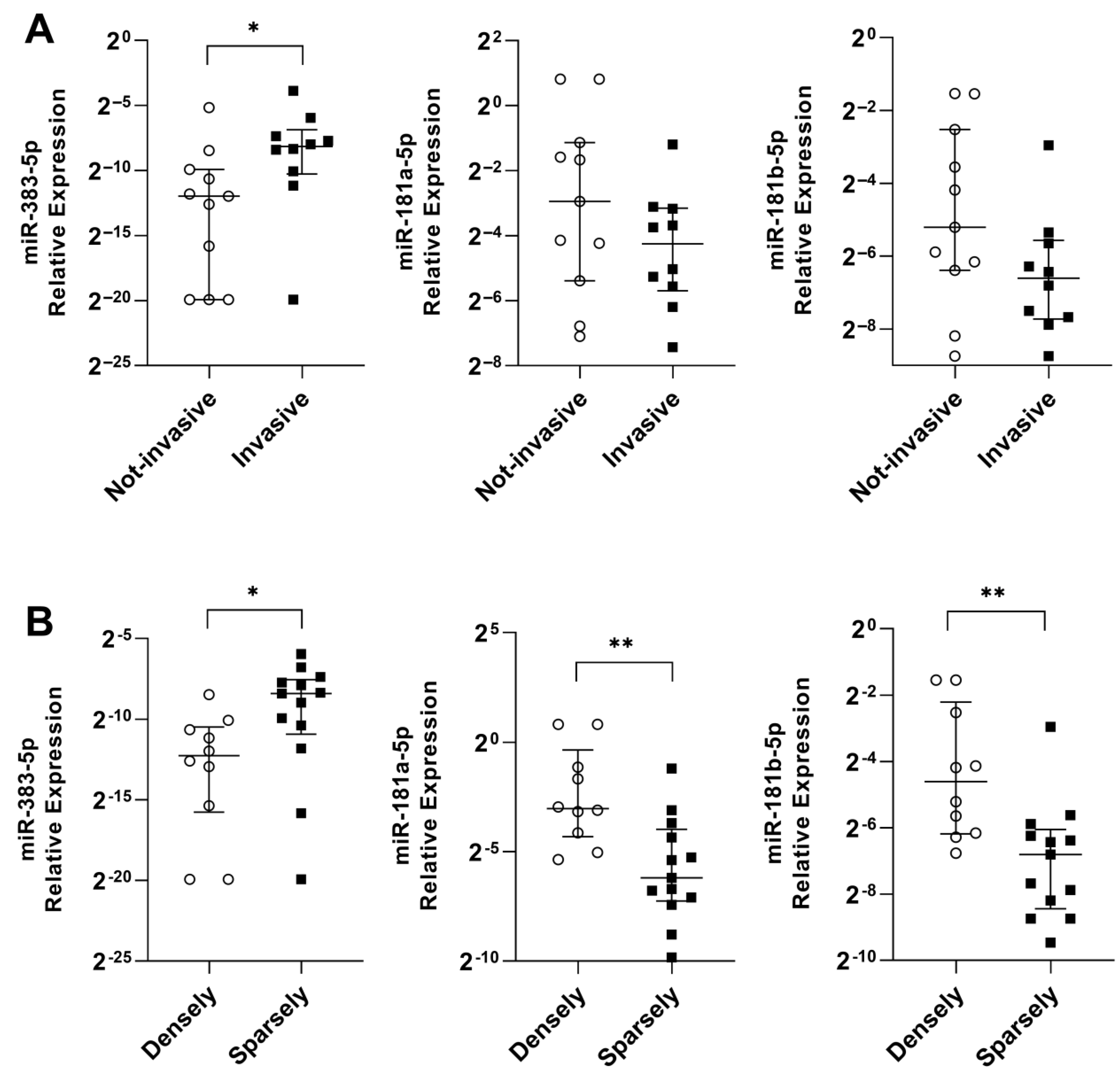
2.4. miR-383-5p, miR-181a-5p, and miR-181b-5p Are Associated with Tumor and Patient Characteristics
3. Materials and Methods
3.1. Patients
3.2. Immunohistochemistry
3.3. RNA Extraction
3.4. MicroRNA Sequencing (miRNA-seq)
3.5. Quantitative Real-Time Polymerase Chain Reaction
3.6. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gadelha, M.R.; Kasuki, L.; Lim, D.S.T.; Fleseriu, M. Systemic Complications of Acromegaly and the Impact of the Current Treatment Landscape: An Update. Endocr. Rev. 2019, 40, 268–332. [Google Scholar] [CrossRef]
- Kasuki, L.; Rocha, P.D.S.; Lamback, E.B.; Gadelha, M.R. Determinants of morbidities and mortality in acromegaly. Arch. Endocrinol. Metab. 2019, 63, 630–637. [Google Scholar] [CrossRef]
- Kasuki, L.; Antunes, X.; Lamback, E.B.; Gadelha, M.R. Acromegaly: Update on Management and Long-Term Morbidities. Endocrinol Metab Clin North Am. 2020, 49, 475–486. [Google Scholar] [CrossRef]
- Kasuki, L.; Maia, B.; Gadelha, M.R. Acromegaly and Colorectal Neoplasm: An Update. Front. Endocrinol. 2022, 13, 924952. [Google Scholar] [CrossRef]
- Fleseriu, M.; Biller, B.M.K.; Freda, P.U.; Gadelha, M.R.; Giustina, A.; Katznelson, L.; Molitch, M.E.; Samson, S.L.; Strasburger, C.J.; van der Lely, A.J.; et al. A Pituitary Society update to acromegaly management guidelines. Pituitary 2021, 24, 1–13. [Google Scholar] [CrossRef]
- Melmed, S.; Colao, A.; Barkan, A.; Molitch, M.; Grossman, A.B.; Kleinberg, D.; Clemmons, D.; Chanson, P.; Laws, E.; Schlechte, J.; et al. Guidelines for acromegaly management: An update. J. Clin. Endocrinol. Metab. 2009, 94, 1509–1517. [Google Scholar] [CrossRef]
- Antunes, X.; Ventura, N.; Camilo, G.B.; Wildemberg, L.E.; Guasti, A.; Pereira, P.J.M.; Camacho, A.H.S.; Chimelli, L.; Niemeyer, P.; Gadelha, M.R.; et al. Predictors of surgical outcome and early criteria of remission in acromegaly. Endocrine 2018, 60, 415–422. [Google Scholar] [CrossRef]
- Theodoropoulou, M.; Stalla, G.K. Somatostatin receptors: From signaling to clinical practice. Front. Neuroendocrinol. 2013, 34, 228–252. [Google Scholar] [CrossRef]
- Gatto, F.; Barbieri, F.; Arvigo, M.; Thellung, S.; Amarù, J.; Albertelli, M.; Ferone, D.; Florio, T. Biological and Biochemical Basis of the Differential Efficacy of First and Second Generation Somatostatin Receptor Ligands in Neuroendocrine Neoplasms. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Gadelha, M.R.; Wildemberg, L.E.; Bronstein, M.D.; Gatto, F.; Ferone, D. Somatostatin receptor ligands in the treatment of acromegaly. Pituitary 2017, 20, 100–108. [Google Scholar] [CrossRef]
- Wildemberg, L.E.; da Silva Camacho, A.H.; Miranda, R.L.; Elias, P.C.L.; de Castro Musolino, N.R.; Nazato, D.; Jallad, R.; Huayllas, M.K.P.; Mota, J.I.S.; Almeida, T.; et al. Machine Learning-based Prediction Model for Treatment of Acromegaly With First-generation Somatostatin Receptor Ligands. J. Clin. Endocrinol. Metab. 2021, 106, 2047–2056. [Google Scholar] [CrossRef]
- Wildemberg, L.E.; Neto, L.V.; Costa, D.F.; Nasciuti, L.E.; Takiya, C.M.; Alves, L.M.; Rebora, A.; Minuto, F.; Ferone, D.; Gadelha, M.R. Low somatostatin receptor subtype 2, but not dopamine receptor subtype 2 expression predicts the lack of biochemical response of somatotropinomas to treatment with somatostatin analogs. J. Endocrinol. Investig. 2013, 36, 38–43. [Google Scholar] [CrossRef]
- Puig-Domingo, M.; Gil, J.; Sampedro-Nuñez, M.; Jordà, M.; Webb, S.M.; Serra, G.; Pons, L.; Salinas, I.; Blanco, A.; Marques-Pamies, M.; et al. Molecular profiling for acromegaly treatment: A validation study. Endocr. Relat. Cancer 2020, 27, 375–389. [Google Scholar] [CrossRef]
- Gadelha, M.R.; Wildemberg, L.E.; Kasuki, L. The Future of Somatostatin Receptor Ligands in Acromegaly. J. Clin. Endocrinol. Metab. 2022, 107, 297–308. [Google Scholar] [CrossRef]
- Mishra, S.; Yadav, T.; Rani, V. Exploring miRNA based approaches in cancer diagnostics and therapeutics. Crit. Rev. Oncol. Hematol. 2016, 98, 12–23. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Denes, J.; Kasuki, L.; Trivellin, G.; Colli, L.M.; Takiya, C.M.; Stiles, C.E.; Barry, S.; de Castro, M.; Gadelha, M.R.; Korbonits, M. Regulation of aryl hydrocarbon receptor interacting protein (AIP) protein expression by MiR-34a in sporadic somatotropinomas. PLoS ONE 2015, 10, e0117107. [Google Scholar] [CrossRef]
- Bogner, E.M.; Daly, A.F.; Gulde, S.; Karhu, A.; Irmler, M.; Beckers, J.; Mohr, H.; Beckers, A.; Pellegata, N.S. miR-34a is upregulated in AIP-mutated somatotropinomas and promotes octreotide resistance. Int. J. Cancer 2020, 147, 3523–3538. [Google Scholar] [CrossRef]
- Palumbo, T.; Faucz, F.R.; Azevedo, M.; Xekouki, P.; Iliopoulos, D.; Stratakis, C.A. Functional screen analysis reveals miR-26b and miR-128 as central regulators of pituitary somatomammotrophic tumor growth through activation of the PTEN-AKT pathway. Oncogene 2013, 32, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.G.; He, D.S.; Zhou, J.; Yao, B.; Xiao, W.W.; Chen, C.H.; Zhu, Y.H.; Wang, H.J. Differential expression of microRNAs in GH-secreting pituitary adenomas. Diagn. Pathol. 2010, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Micko, A.S.; Wöhrer, A.; Wolfsberger, S.; Knosp, E. Invasion of the cavernous sinus space in pituitary adenomas: Endoscopic verification and its correlation with an MRI-based classification. J. Neurosurg. 2015, 122, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.C.A.; Vasquez, M.L.; Wildemberg, L.E.; Vázquez-Borrego, M.C.; Bitana, L.; Camacho, A.H.D.S.; Silva, D.; Ogino, L.L.; Ventura, N.; Chimelli, L.; et al. Molecular evidence and clinical importance of β-arrestins expression in patients with acromegaly. J. Cell. Mol. Med. 2018, 22, 2110–2116. [Google Scholar] [CrossRef] [PubMed]
- Rindi, G.; Mete, O.; Uccella, S.; Basturk, O.; La Rosa, S.; Brosens, L.A.A.; Ezzat, S.; de Herder, W.W.; Klimstra, D.S.; Papotti, M.; et al. Overview of the 2022 WHO Classification of Neuroendocrine Neoplasms. Endocr. Pathol. 2022, 33, 115–154. [Google Scholar] [CrossRef]
- Obari, A.; Sano, T.; Ohyama, K.; Kudo, E.; Qian, Z.R.; Yoneda, A.; Rayhan, N.; Mustafizur Rahman, M.; Yamada, S. Clinicopathological features of growth hormone-producing pituitary adenomas: Difference among various types defined by cytokeratin distribution pattern including a transitional form. Endocr. Pathol. 2008, 19, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Gatto, F.; Feelders, R.A.; van der Pas, R.; Kros, J.M.; Waaijers, M.; Sprij-Mooij, D.; Neggers, S.J.; van der Lelij, A.J.; Minuto, F.; Lamberts, S.W.; et al. Immunoreactivity score using an anti-sst2A receptor monoclonal antibody strongly predicts the biochemical response to adjuvant treatment with somatostatin analogs in acromegaly. J. Clin. Endocrinol. Metab. 2013, 98, E66–E71. [Google Scholar] [CrossRef]
- Fehlmann, T.; Reinheimer, S.; Geng, C.; Su, X.; Drmanac, S.; Alexeev, A.; Zhang, C.; Backes, C.; Ludwig, N.; Hart, M.; et al. cPAS-based sequencing on the BGISEQ-500 to explore small non-coding RNAs. Clin. Epigenetics 2016, 8, 123. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Puerta, E.; Lebrón, R.; Rueda, A.; Gómez-Martín, C.; Giannoukakos, S.; Jaspez, D.; Medina, J.M.; Zubkovic, A.; Jurak, I.; Fromm, B.; et al. sRNAbench and sRNAtoolbox 2019: Intuitive fast small RNA profiling and differential expression. Nucleic Acids Res. 2019, 47, W530–W535. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Fougner, S.L.; Casar-Borota, O.; Heck, A.; Berg, J.P.; Bollerslev, J. Adenoma granulation pattern correlates with clinical variables and effect of somatostatin analogue treatment in a large series of patients with acromegaly. Clin. Endocrinol. 2012, 76, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Paragliola, R.M.; Corsello, S.M.; Salvatori, R. Somatostatin receptor ligands in acromegaly: Clinical response and factors predicting resistance. Pituitary 2017, 20, 109–115. [Google Scholar] [CrossRef]
- Fan, X.; Mao, Z.; He, D.; Liao, C.; Jiang, X.; Lei, N.; Hu, B.; Wang, X.; Li, Z.; Lin, Y.; et al. Expression of somatostatin receptor subtype 2 in growth hormone-secreting pituitary adenoma and the regulation of miR-185. J. Endocrinol. Investig. 2015, 38, 1117–1128. [Google Scholar] [CrossRef]
- Braicu, C.; Gulei, D.; Raduly, L.; Harangus, A.; Rusu, A.; Berindan-Neagoe, I. Altered expression of miR-181 affects cell fate and targets drug resistance-related mechanisms. Mol. Asp. Med. 2019, 70, 90–105. [Google Scholar] [CrossRef]
- Rezaei, T.; Amini, M.; Hashemi, Z.S.; Mansoori, B.; Rezaei, S.; Karami, H.; Mosafer, J.; Mokhtarzadeh, A.; Baradaran, B. microRNA-181 serves as a dual-role regulator in the development of human cancers. Free. Radic. Biol. Med. 2020, 152, 432–454. [Google Scholar] [CrossRef]
- Eigler, T.; Ben-Shlomo, A. Somatostatin system: Molecular mechanisms regulating anterior pituitary hormones. J. Mol. Endocrinol. 2014, 53, R1–R19. [Google Scholar] [CrossRef]
- Huang, X.; Schwind, S.; Santhanam, R.; Eisfeld, A.K.; Chiang, C.L.; Lankenau, M.; Yu, B.; Hoellerbauer, P.; Jin, Y.; Tarighat, S.S.; et al. Targeting the RAS/MAPK pathway with miR-181a in acute myeloid leukemia. Oncotarget 2016, 7, 59273–59286. [Google Scholar] [CrossRef]
- Fu, Y.; Xin, Z.; Ling, Z.; Xie, H.; Xiao, T.; Shen, X.; Lin, J.; Xu, L.; Jiang, H. A CREB1-miR-181a-5p loop regulates the pathophysiologic features of bone marrow stromal cells in fibrous dysplasia of bone. Mol. Med. 2021, 27, 81. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Tan, F.; Yu, N.; Pei, H. STAT1 Inhibits MiR-181a Expression to Suppress Colorectal Cancer Cell Proliferation Through PTEN/Akt. J. Cell. Biochem. 2017, 118, 3435–3443. [Google Scholar] [CrossRef]
- Hollander, M.C.; Blumenthal, G.M.; Dennis, P.A. PTEN loss in the continuum of common cancers, rare syndromes and mouse models. Nat. Rev. Cancer 2011, 11, 289–301. [Google Scholar] [CrossRef]
- Li, X.; Han, J.; Zhu, H.; Peng, L.; Chen, Z. miR-181b-5p mediates TGF-β1-induced epithelial-to-mesenchymal transition in non-small cell lung cancer stem-like cells derived from lung adenocarcinoma A549 cells. Int. J. Oncol. 2017, 51, 158–168. [Google Scholar] [CrossRef]
- Yi, Q.; Xie, W.; Sun, W.; Liao, Y. A Concise Review of MicroRNA-383: Exploring the Insights of Its Function in Tumorigenesis. J. Cancer 2022, 13, 313–324. [Google Scholar] [CrossRef]
- Wu, S.; Gu, Y.; Huang, Y.; Wong, T.C.; Ding, H.; Liu, T.; Zhang, Y.; Zhang, X. Novel Biomarkers for Non-functioning Invasive Pituitary Adenomas were Identified by Using Analysis of microRNAs Expression Profile. Biochem. Genet. 2017, 55, 253–267. [Google Scholar] [CrossRef]
- Liu, J.; Dou, Y.; Sheng, M. Inhibition of microRNA-383 has tumor suppressive effect in human epithelial ovarian cancer through the action on caspase-2 gene. Biomed. Pharmacother. 2016, 83, 1286–1294. [Google Scholar] [CrossRef]
- Kopeina, G.S.; Zhivotovsky, B. Caspase-2 as a master regulator of genomic stability. Trends Cell Biol. 2021, 31, 712–720. [Google Scholar] [CrossRef]
- Wang, X.; Ding, G.; Lai, W.; Liu, S.; Shuai, J. MicroRNA-383 upregulation protects against propofol-induced hippocampal neuron apoptosis and cognitive impairment. Exp. Ther. Med. 2018, 15, 3181–3188. [Google Scholar] [CrossRef]
- Roberts, O.; Paraoan, L. PERP-ing into diverse mechanisms of cancer pathogenesis: Regulation and role of the p53/p63 effector PERP. Biochim. Et Biophys. Acta Rev. Cancer 2020, 1874, 188393. [Google Scholar] [CrossRef]
- Leonart, L.P.; Riveros, B.S.; Krahn, M.D.; Pontarolo, R. Pharmacological Acromegaly Treatment: Cost-Utility and Value of Information Analysis. Neuroendocrinology 2021, 111, 388–402. [Google Scholar] [CrossRef]
- Ferone, D.; de Herder, W.W.; Pivonello, R.; Kros, J.M.; van Koetsveld, P.M.; de Jong, T.; Minuto, F.; Colao, A.; Lamberts, S.W.; Hofland, L.J. Correlation of in vitro and in vivo somatotropic adenoma responsiveness to somatostatin analogs and dopamine agonists with immunohistochemical evaluation of somatostatin and dopamine receptors and electron microscopy. J. Clin. Endocrinol. Metab. 2008, 93, 1412–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controlled (n = 13) | Uncontrolled (n = 19) | p-Value | |
|---|---|---|---|
| Median age (years) | 41 (23–59) | 33 (21–56) | 0.05 |
| Gender (male %) | 38.5 (5/13) | 63.1 (12/19) | 0.15 |
| Median pretreatment GH (ng/mL) | 2.8 (0.7–15.8) | 5.1 (0.5–35) | 0.2 |
| Median GH decrease (%) | 88.6 (32.4–94.9) | 34.0 (−523.2–86.0) | 0.003 |
| Median pretreatment IGF-I (xULN) | 2.4 (1.3–3.0) | 2.8 (1.3–4.0) | 0.08 |
| Median IGF-I decrease (%) | 71.0 (51.5–81.1) | 32.4 (−67.6–75.9) | 0.001 |
| Median largest tumor diameter (cm) | 2.0 (1.0–6.1) | 2.6 (1.0–4.7) | 0.6 |
| Invasive tumor (%) | 25 (2/8) | 61.5 (8/13) | 0.18 |
| Sparsely granulated (%) | 25 (3/9) | 62.5 (10/14) | 0.1 |
| Median SST2 (IRS) | 12 (6–12) | 7 (0–12) | 0.06 |
| Median SST5 (IRS) | 4 (2–12) | 9 (0–12) | 0.6 |
| Controlled (n = 5) | Uncontrolled (n = 5) | |
|---|---|---|
| Median age (years) | 37.0 (35.0–59.0) | 25.0 (21.0–33.0) |
| Gender (male %) | 40.0 (2/5) | 40.0 (2/5) |
| Median pretreatment GH (ng/mL) | 1.8 (0.7–4.0) | 8.0 (2.8–21.9) |
| Median GH decrease (%) | 83.0 (32.4–94.1) | −18.7 (−523.2–17.8) |
| Median pretreatment IGF-I (xULN) | 2.4 (1.7–2.7) | 2.8 (1.8–3.7) |
| Median IGF-I decrease (%) | 74.8 (65.8–81.1) | −41.9 (−67.6–42.8) |
| Median largest tumor diameter (cm) | 2.0 (1.4–3.7) | 2.4 (1.0–4.1) |
| Invasive tumor (%) | 25.0 (1/4) | 66.7 (2/3) |
| Sparsely granulated (%) | 25.0 (1/4) | 75.0 (3/4) |
| Median SST2 (IRS) | 12 (6–12) | 5 (0–12) |
| Median SST5 (IRS) | 4 (4–12) | 9 (0–12) |
| Controlled (n = 8) | Uncontrolled (n = 14) | p-Value | |
|---|---|---|---|
| Median age (years) | 44.0 (23.0–58.0) | 38.0 (22.0–56.0) | 0.34 |
| Gender (male %) | 37.5 (3/8) | 78.6 (11/14) | 0.08 |
| Median pretreatment GH (ng/mL) | 2.8 (0.9–15.8) | 3.7 (0.53–65.2) | 0.64 |
| Median GH decrease (%) | 89.1 (44.6–94.9) | 47.1 (−95.8–85.9) | 0.002 |
| Median pretreatment IGF-I (xULN) | 2.4 (1.3–3.0) | 2.8 (1.4–4.0) | 0.19 |
| Median IGF-I decrease (%) | 68.6 (51.5–75.2) | 34.9 (−8.8–75.9) | 0.003 |
| Median largest tumor diameter (cm) | 2.6 (1.0–6.1) | 2.6 (1.2–4.7) | 0.66 |
| Invasive tumor (%) | 25.0 (1/4) | 60.0 (6/10) | 0.56 |
| Sparsely granulated (%) | 40.0 (2/5) | 58.3 (7/12) | 0.62 |
| Median SST2 (IRS) | 12 (6–12) | 9 (4–12) | 0.30 |
| Median SST5 (IRS) | 6 (2–12) | 9 (0–12) | 0.90 |
| miRNA | Controlled (n = 8) | Uncontrolled (n = 14) | p-Value |
|---|---|---|---|
| miR-10b-5p | 2.67 × 10−5 (0–5.30 × 10−3) | 1.76 × 10−5 (0–1.11 × 10−3) | 0.595 |
| miR-27b-3p | 4.37 × 10−2 (7.19 × 10−3–1.19 × 10−1) | 3.35 × 10−2 (4.19 × 10−3–1.14 × 10−1) | 0.482 |
| miR-181a-5p | 1.13 × 10−1 (7.31 × 10−3–1.12) | 2.75 × 10−2 (1.10 × 10−3–4.84) | 0.075 |
| miR-181b-5p | 3.55 × 10−2 (3.42 × 10−3–2.94 × 10−1) | 1.18 × 10−2 (2.33 × 10−3–6.20 × 10−1) | 0.071 |
| miR-199b-5p | 2.00 × 10−4 (0–1.35 × 10−2) | 3.60 × 10−5 (0–1.28 × 10−2) | 0.185 |
| miR-383-5p | 1.73 × 10−5 (0–5.78 × 10−3) | 2.97 × 10−3 (2.37 × 10−5–6.82 × 10−2) | 0.041 |
| miR-505-5p | 8.20 × 10−6 (0–6.35 × 10−4) | 9.08 × 10−6 (0–1.45 × 10−3) | 0.773 |
| miR-629-3p | 1.59 × 10−5 (0–1.25 × 10−4) | 2.62 × 10−6 (0–1.87 × 10−3) | 0.862 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriques, D.G.; Miranda, R.L.; Dezonne, R.S.; Wildemberg, L.E.; Camacho, A.H.d.S.; Chimelli, L.; Kasuki, L.; Lamback, E.B.; Guterres, A.; Gadelha, M.R. miR-383-5p, miR-181a-5p, and miR-181b-5p as Predictors of Response to First-Generation Somatostatin Receptor Ligands in Acromegaly. Int. J. Mol. Sci. 2023, 24, 2875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032875
Henriques DG, Miranda RL, Dezonne RS, Wildemberg LE, Camacho AHdS, Chimelli L, Kasuki L, Lamback EB, Guterres A, Gadelha MR. miR-383-5p, miR-181a-5p, and miR-181b-5p as Predictors of Response to First-Generation Somatostatin Receptor Ligands in Acromegaly. International Journal of Molecular Sciences. 2023; 24(3):2875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032875
Chicago/Turabian StyleHenriques, Daniel G., Renan Lyra Miranda, Rômulo Sperduto Dezonne, Luiz Eduardo Wildemberg, Aline Helen da Silva Camacho, Leila Chimelli, Leandro Kasuki, Elisa B. Lamback, Alexandro Guterres, and Monica R. Gadelha. 2023. "miR-383-5p, miR-181a-5p, and miR-181b-5p as Predictors of Response to First-Generation Somatostatin Receptor Ligands in Acromegaly" International Journal of Molecular Sciences 24, no. 3: 2875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032875