

Effect of 3D Spheroid Culturing on NF-κB Signaling Pathway and Neurogenic Potential in Human Amniotic Fluid Stem Cells


Abstract
:1. Introduction
2. Results
2.1. Evaluation of Morphological Characteristics and Expression of Cell Surface Markers in hAFSCs Grown in 2D and 3D Cultures
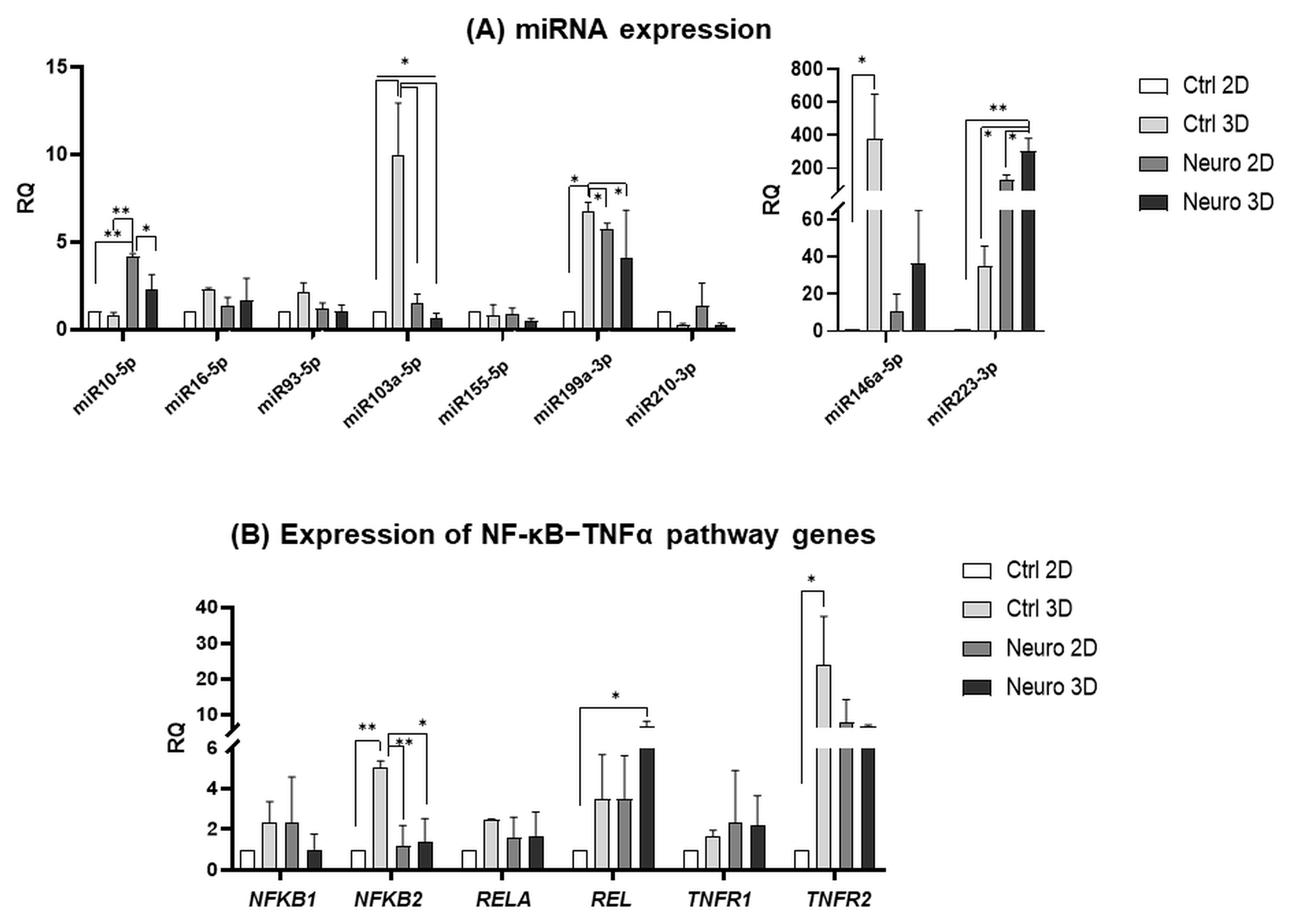
2.2. Gene and miRNA Expression Patterns in Control and Neuro-Differentiated hAFSCs Grown in 2D and 3D Cultures
2.3. Modulation of Protein Expression in Control and Neuro-Differentiated hAFSCs Grown in 2D and 3D Cultures
2.4. Examination of hAFSCs Secretion Profile in 2D and 3D Cultures
3. Discussion
4. Materials and Methods
4.1. Isolation, Cultivation and Spheroid Formation of Human Amniotic Fluid Stem Cells
4.2. Differentiation Assay
4.3. Flow Cytometry Analysis
4.4. RNA Isolation and RT-qPCR
4.5. Evaluation of Micro RNAs
4.6. Immunofluorescence Analysis
4.7. Protein Isolation and Western Blot Analysis
4.8. Secretome Preparation for Mass Spectrometry
4.9. LC-MS-Based Protein Identification
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joerger-Messerli, M.S.; Marx, C.; Oppliger, B.; Mueller, M.; Surbek, D.V.; Schoeberlein, A. Mesenchymal Stem Cells from Wharton’s Jelly and Amniotic Fluid. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 30–44. [Google Scholar] [CrossRef]
- Yan, Z.-J.; Hu, Y.-Q.; Zhang, H.-T.; Zhang, P.; Xiao, Z.-Y.; Sun, X.-L.; Cai, Y.-Q.; Hu, C.-C.; Xu, R.-X. Comparison of the Neural Differentiation Potential of Human Mesenchymal Stem Cells from Amniotic Fluid and Adult Bone Marrow. Cell. Mol. Neurobiol. 2013, 33, 465–475. [Google Scholar] [CrossRef]
- Alessio, N.; Pipino, C.; Mandatori, D.; Di Tomo, P.; Ferone, A.; Marchiso, M.; Melone, M.A.; Peluso, G.; Pandolfi, A.; Galderisi, U. Mesenchymal stromal cells from amniotic fluid are less prone to senescence compared to those obtained from bone marrow: An in vitro study. J. Cell. Physiol. 2018, 233, 8996–9006. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Chun, S.Y.; Kil Lee, J.; Lim, H.J.; Bae, J.-S.; Chung, H.-Y.; Atala, A.; Soker, S.; Yoo, J.J.; Kwon, T.G. Human amniotic fluid stem cell injection therapy for urethral sphincter regeneration in an animal model. BMC Med. 2012, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Loukogeorgakis, S.P.; De Coppi, P. Concise Review: Amniotic Fluid Stem Cells: The Known, the Unknown, and Potential Regenerative Medicine Applications. Stem Cells 2017, 35, 1663–1673. [Google Scholar] [CrossRef]
- Abe, Y.; Ochiai, D.; Sato, Y.; Otani, T.; Fukutake, M.; Ikenoue, S.; Kasuga, Y.; Tanaka, M. Amniotic fluid stem cells as a novel strategy for the treatment of fetal and neonatal neurological diseases. Placenta 2021, 104, 247–252. [Google Scholar] [CrossRef]
- Laschke, M.W.; Menger, M.D. Life is 3D: Boosting Spheroid Function for Tissue Engineering. Trends Biotechnol. 2016, 35, 133–144. [Google Scholar] [CrossRef]
- Ezquerra, S.; Zuleta, A.; Arancibia, R.; Estay, J.; Aulestia, F.; Carrion, F. Functional Properties of Human-Derived Mesenchymal Stem Cell Spheroids: A Meta-Analysis and Systematic Review. Stem Cells Int. 2021, 2021, 8825332. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Sancilio, S.; Di Mauro, M.; Iannetti, G.; Dolci, S.; Amabile, G.; Di Baldassarre, A.; Ghinassi, B. Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells. Int. J. Mol. Sci. 2020, 21, 6317. [Google Scholar] [CrossRef]
- Mohammed, E.E.A.; Beherei, H.; El-Zawahry, M.; Farrag, A.R.; Kholoussi, N.; Helwa, I.; Gaber, K.; Allam, M.A.; Mabrouk, M.; Aleem, A.K.A. Combination of Human Amniotic Fluid Derived-Mesenchymal Stem Cells and Nano-hydroxyapatite Scaffold Enhances Bone Regeneration. Open Access Maced. J. Med. Sci. 2019, 7, 2739–2750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shieh, H.F.; Graham, C.D.; Brazzo, J.A.; Zurakowski, D.; Fauza, D.O. Comparisons of human amniotic mesenchymal stem cell viability in FDA-approved collagen-based scaffolds: Implications for engineered diaphragmatic replacement. J. Pediatr. Surg. 2017, 52, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Zinin, N.; Adameyko, I.; Wilhelm, M.; Fritz, N.; Uhlén, P.; Ernfors, P.; Henriksson, M.A. MYC proteins promote neuronal differentiation by controlling the mode of progenitor cell division. EMBO Rep. 2014, 15, 383–391. [Google Scholar] [CrossRef]
- Gao, L.; Yang, X.; Zhang, H.; Yu, M.; Long, J.; Yang, T. Inhibition of miR-10a-5p suppresses cholangiocarcinoma cell growth through downregulation of Akt pathway. OncoTargets Ther. 2018, 11, 6981–6994. [Google Scholar] [CrossRef] [PubMed]
- Schubbert, S.; Jiao, J.; Ruscetti, M.; Nakashima, J.; Wu, S.; Lei, H.; Xu, Q.; Yi, W.; Zhu, H.; Wu, H. Methods for PTEN in Stem Cells and Cancer Stem Cells BT. In PTEN: Methods and Protocols; Salmena, L., Stambolic, V., Eds.; Springer New York: New York, NY, USA, 2016; pp. 233–285. [Google Scholar]
- Harraz, M.M.; Xu, J.-C.; Guiberson, N.; Dawson, T.M.; Dawson, V.L. MiR-223 regulates the differentiation of immature neurons. Mol. Cell. Ther. 2014, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Liang, C.; Hua, J.; Zhang, B.; Liu, J.; Zhang, Y.; Wei, M.; Yu, X.; Xu, J.; Shi, S. A miR-146a-5p/TRAF6/NF-kB p65 axis regulates pancreatic cancer chemoresistance: Functional validation and clinical significance. Theranostics 2020, 10, 3967–3979. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Tang, Q. Reduced expression of microRNA-199a-3p is associated with vascular endothelial cell injury induced by type 2 diabetes mellitus. Exp. Ther. Med. 2018, 16, 3639–3645. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Niu, P.; Zhao, Y.; Cheng, Y.; Chen, W.; Lin, L.; Lu, J.; Cheng, X.; Xu, Z. Impact of MiR-223-3p and MiR-2909 on Inflammatory Factors IL-6, IL-1ß, and TNF-α, and the TLR4/TLR2/NF-ΚB/STAT3 Signaling Pathway Induced by Lipopolysaccharide in Human Adipose Stem Cells. PLoS ONE 2019, 14, e0212063. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, W.; Kong, X.; Li, J.; Chen, X.; Ge, Y.; Zhong, W.; Fang, S. Protective effects of α 2 macroglobulin on human bone marrow mesenchymal stem cells in radiation injury. Mol. Med. Rep. 2018, 18, 4219–4228. [Google Scholar] [CrossRef]
- Bitzer, M.; Gersdorff GVon Liang, D.; Dominguez-Rosales, A.; Beg, A.A.; Rojkind, M.; Böttinger, E.P. A mechanism of suppression of TGF-β/SMAD signaling by NF-κB/RelA. Genes Dev. 2000, 14, 187–197. [Google Scholar] [CrossRef]
- Gao, S.; Sun, Y.; Zhang, X.; Hu, L.; Liu, Y.; Chua, C.Y.; Phillips, L.M.; Ren, H.; Fleming, J.B.; Wang, H.; et al. IGFBP2 activates the NF-κB pathway to drive epithelial-mesenchymal transition and invasive character in pancreatic ductal adenocarcinoma. Cancer Res. 2016, 76, 6543–6554. [Google Scholar] [CrossRef] [Green Version]
- Han, H.-W.; Asano, S.; Hsu, S.-H. Cellular Spheroids of Mesenchymal Stem Cells and Their Perspectives in Future Healthcare. Appl. Sci. 2019, 9, 627. [Google Scholar] [CrossRef]
- Domnina, A.; Novikova, P.; Obidina, J.; Fridlyanskaya, I.; Alekseenko, L.; Kozhukharova, I.; Lyublinskaya, O.; Zenin, V.; Nikolsky, N. Human mesenchymal stem cells in spheroids improve fertility in model animals with damaged endometrium. Stem Cell Res. Ther. 2018, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Niibe, K.; Ohori-Morita, Y.; Zhang, M.; Mabuchi, Y.; Matsuzaki, Y.; Egusa, H. A Shaking-Culture Method for Generating Bone Marrow Derived Mesenchymal Stromal/Stem Cell-Spheroids With Enhanced Multipotency in vitro. Front. Bioeng. Biotechnol. 2020, 8, 590332. [Google Scholar] [CrossRef]
- Amador-Arjona, A.; Cimadamore, F.; Huang, C.-T.; Wright, R.; Lewis, S.; Gage, F.H.; Terskikh, A.V. SOX2 primes the epigenetic landscape in neural precursors enabling proper gene activation during hippocampal neurogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, E1936–E1945. [Google Scholar] [CrossRef] [PubMed]
- Bley, N.; Hmedat, A.; Müller, S.; Rolnik, R.; Rausch, A.; Lederer, M.; Hüttelmaier, S. Musashi–1—A Stemness RBP for Cancer Therapy? Biology 2021, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, M.; Lachinani, L.; Dormiani, K.; Nasr-Esfahani, M.H.; Gure, A.O.; Ghaedi, K. Intracellular functions of RNA-binding protein, Musashi1, in stem and cancer cells. Stem Cell Res. Ther. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Sakakibara, S.-I.; Imai, T.; Suzuki, A.; Nakamura, Y.; Sawamoto, K.; Ogawa, Y.; Toyama, Y.; Miyata, T.; Okano, H. Musashi1: An Evolutionally Conserved Marker for CNS Progenitor Cells Including Neural Stem Cells. Dev. Neurosci. 2000, 22, 139–153. [Google Scholar] [CrossRef]
- Kawase, S.; Kuwako, K.; Imai, T.; Renault-Mihara, F.; Yaguchi, K.; Itohara, S.; Okano, H. Regulatory Factor X Transcription Factors Control Musashi1 Transcription in Mouse Neural Stem/Progenitor Cells. Stem Cells Dev. 2014, 23, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Zaytseva, O.; Kim, N.H.; Quinn, L.M. Myc in brain development and cancer. Int. J. Mol. Sci. 2020, 21, 7742. [Google Scholar] [CrossRef] [PubMed]
- Conacci-Sorrell, M.; Ngouenet, C.; Eisenman, R.N. Myc-Nick: A Cytoplasmic Cleavage Product of Myc that Promotes α-Tubulin Acetylation and Cell Differentiation. Cell 2010, 142, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Jovičić, A.; Roshan, R.; Moisoi, N.; Pradervand, S.; Moser, R.; Pillai, B.; Luthi-Carter, R. Comprehensive expression analyses of neural cell-type-specific miRNAs identify new determinants of the specification and maintenance of neuronal phenotypes. Ann. Intern. Med. 2013, 158, 5127–5137. [Google Scholar]
- Yu, X.; Nagai, J.; Khakh, B.S. Improved tools to study astrocytes. Nat. Rev. Neurosci. 2020, 21, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, P.; Zhou, Z.; Natarelli, L.; Wei, Y.; Nazari-Jahantigh, M.; Zhu, M.; Grommes, J.; Steffens, S.; Weber, C.; Schober, A. Endothelial Dicer promotes atherosclerosis and vascular inflammation by miRNA-103-mediated suppression of KLF4. Nat. Commun. 2016, 7, 10521. [Google Scholar] [CrossRef]
- Noorolyai, S.; Baghbani, E.; Maleki, L.A.; Kojabad, A.B.; Shanehbansdi, D.; Shahgoli, V.K.; Mokhtarzadeh, A.; Baradaran, B. Restoration of miR-193a-5p and miR-146 a-5p Expression Induces G1 Arrest in Colorectal Cancer through Targeting of MDM2/p53. Adv. Pharm. Bull. 2020, 10, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Li, P.; Li, B.; Miao, D.; Deng, Q. RelA promotes proliferation but inhibits osteogenic and chondrogenic differentiation of mesenchymal stem cells. FEBS Lett. 2020, 594, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Ma, Z.; Ding, Y.; Bedarida, T.; Chen, L.; Xie, Z.; Song, P.; Zou, M.H. Circulating miR-103a-3p contributes to angiotensin II-induced renal inflammation and fibrosis via a SNRK/NF-κB/p65 regulatory axis. Nat. Commun. 2019, 10, 2145. [Google Scholar] [CrossRef]
- De Luca, F. Regulatory role of NF-κB in growth plate chondrogenesis and its functional interaction with Growth Hormone. Mol. Cell. Endocrinol. 2020, 514, 110916. [Google Scholar] [CrossRef]
- Crisostomo, P.R.; Wang, Y.; Markel, T.A.; Wang, M.; Lahm, T.; Meldrum, D.R. Human mesenchymal stem cells stimulated by TNF-α, LPS, or hypoxia produce growth factors by an NFκB- but not JNK-dependent mechanism. Am. J. Physiol. Cell Physiol. 2008, 294, 675–682. [Google Scholar] [CrossRef]
- Becker-Weimann, S.; Xiong, G.; Furuta, S.; Han, J.; Kuhn, I.; Akavia, U.-D.; Pe’Er, D.; Bissell, M.J.; Xu, R. NFkB disrupts tissue polarity in 3D by preventing integration of microenvironmental signals. Oncotarget 2013, 4, 2010–2020. [Google Scholar] [CrossRef]
- Chan, Y.-H.; Lee, Y.-C.; Hung, C.-Y.; Yang, P.-J.; Lai, P.-C.; Feng, S.-W. Three-dimensional Spheroid Culture Enhances Multipotent Differentiation and Stemness Capacities of Human Dental Pulp-derived Mesenchymal Stem Cells by Modulating MAPK and NF-kB Signaling Pathways. Stem Cell Rev. Rep. 2021, 17, 1810–1826. [Google Scholar] [CrossRef]
- Domnina, A.; Ivanova, J.; Alekseenko, L.; Kozhukharova, I.; Borodkina, A.; Pugovkina, N.; Smirnova, I.; Lyublinskaya, O.; Fridlyanskaya, I.; Nikolsky, N. Three-Dimensional Compaction Switches Stress Response Programs and Enhances Therapeutic Efficacy of Endometrial Mesenchymal Stem/Stromal Cells. Front. Cell Dev. Biol. 2020, 8, 473. [Google Scholar] [CrossRef]
- Joshi, P.; Bodnya, C.; Rasmussen, M.L.; Romero-Morales, A.I.; Bright, A.; Gama, V. Modeling the function of BAX and BAK in early human brain development using iPSC-derived systems. Cell Death Dis. 2020, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Savickiene, J.; Treigyte, G.; Baronaite, S.; Valiuliene, G.; Kaupinis, A.; Valius, M.; Arlauskiene, A.; Navakauskienė, R. Human Amniotic Fluid Mesenchymal Stem Cells from Second- and Third-Trimester Amniocentesis: Differentiation Potential, Molecular Signature, and Proteome Analysis. Stem Cells Int. 2015, 2015, 319238. [Google Scholar] [CrossRef]
- Gasiūnienė, M.; Zentelytė, A.; Treigytė, G.; Baronaitė, S.; Savickienė, J.; Utkus, A.; Navakauskienė, R. Epigenetic alterations in amniotic fluid mesenchymal stem cells derived from normal and fetus-affected gestations: A focus on myogenic and neural differentiations. Cell Biol. Int. 2019, 43, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated in 3D Secretome | Downregulated in 3D Secretome | |||
|---|---|---|---|---|
| No. | Protein Name | Fold Change | Protein Name | Fold Change |
| 1 | Alpha-2-macroglobulin (A2M) | 11–18 | Plasminogen activator inhibitor 1 (SERPINE1) | 25–484 |
| 2 | Complement C3 (C3) | 5–15 | Transforming growth factor-beta-induced protein (TGFBI) | 82–409 |
| 3 | Ryanodine receptor 3 (RYR3) | 4–14 | Secreted protein acidic and rich in cysteine (SPARC) | 49–195 |
| 4 | Leucine-rich repeat serine/threonine-protein kinase 2 (LRRK2) | 4–10 | Fibronectin (FN1) | 72–83 |
| 5 | Dynein axonemal heavy chain 2 (DNAH2) | 2–5 | Metalloproteinase inhibitor 1 (TIMP1) | 36–83 |
| 6 | Immunoglobulin kappa constant (IGKC) | 3 | Collagen alpha-1(V) chain (COL5A1) | 35–36 |
| 7 | E3 ubiquitin-protein ligase RNF213 (RNF213) | 1.5–3.2 | Collagen alpha-1(I) chain (COL1A1) | 20–28 |
| 8 | Centlein (CNTLN) | 1.9–2.2 | Galectin-1 (LGALS1) | 8–17 |
| 9 | Putative uncharacterized protein LINC02901 (C6orf99) | 1.8–2.2 | Nucleobindin-1 (NUCB1) | 4–8 |
| 10 | Dynein axonemal heavy chain 12 (DNAH12) | 1.6–2.2 | Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | 7–9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiulienė, G.; Zentelytė, A.; Beržanskytė, E.; Navakauskienė, R. Effect of 3D Spheroid Culturing on NF-κB Signaling Pathway and Neurogenic Potential in Human Amniotic Fluid Stem Cells. Int. J. Mol. Sci. 2023, 24, 3584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043584
Valiulienė G, Zentelytė A, Beržanskytė E, Navakauskienė R. Effect of 3D Spheroid Culturing on NF-κB Signaling Pathway and Neurogenic Potential in Human Amniotic Fluid Stem Cells. International Journal of Molecular Sciences. 2023; 24(4):3584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043584
Chicago/Turabian StyleValiulienė, Giedrė, Aistė Zentelytė, Elizabet Beržanskytė, and Rūta Navakauskienė. 2023. "Effect of 3D Spheroid Culturing on NF-κB Signaling Pathway and Neurogenic Potential in Human Amniotic Fluid Stem Cells" International Journal of Molecular Sciences 24, no. 4: 3584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043584