
Cytokinin Response Factor 9 Represses Cytokinin Responses in Flower Development
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CRFs Are Found in Early Divergent Land Plants and Streptophyte Algae
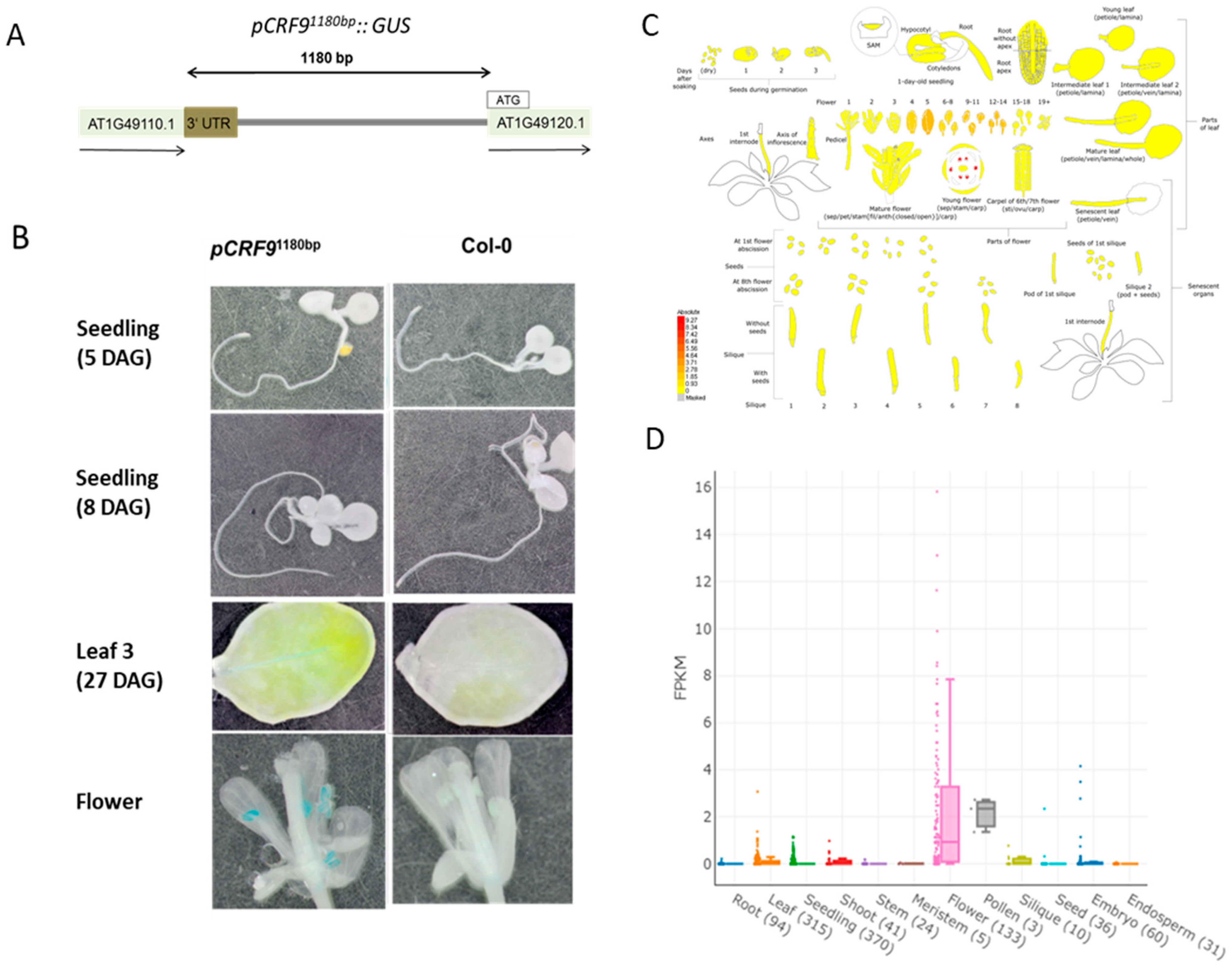
2.2. CRF9 Is Mainly Expressed in the Flower
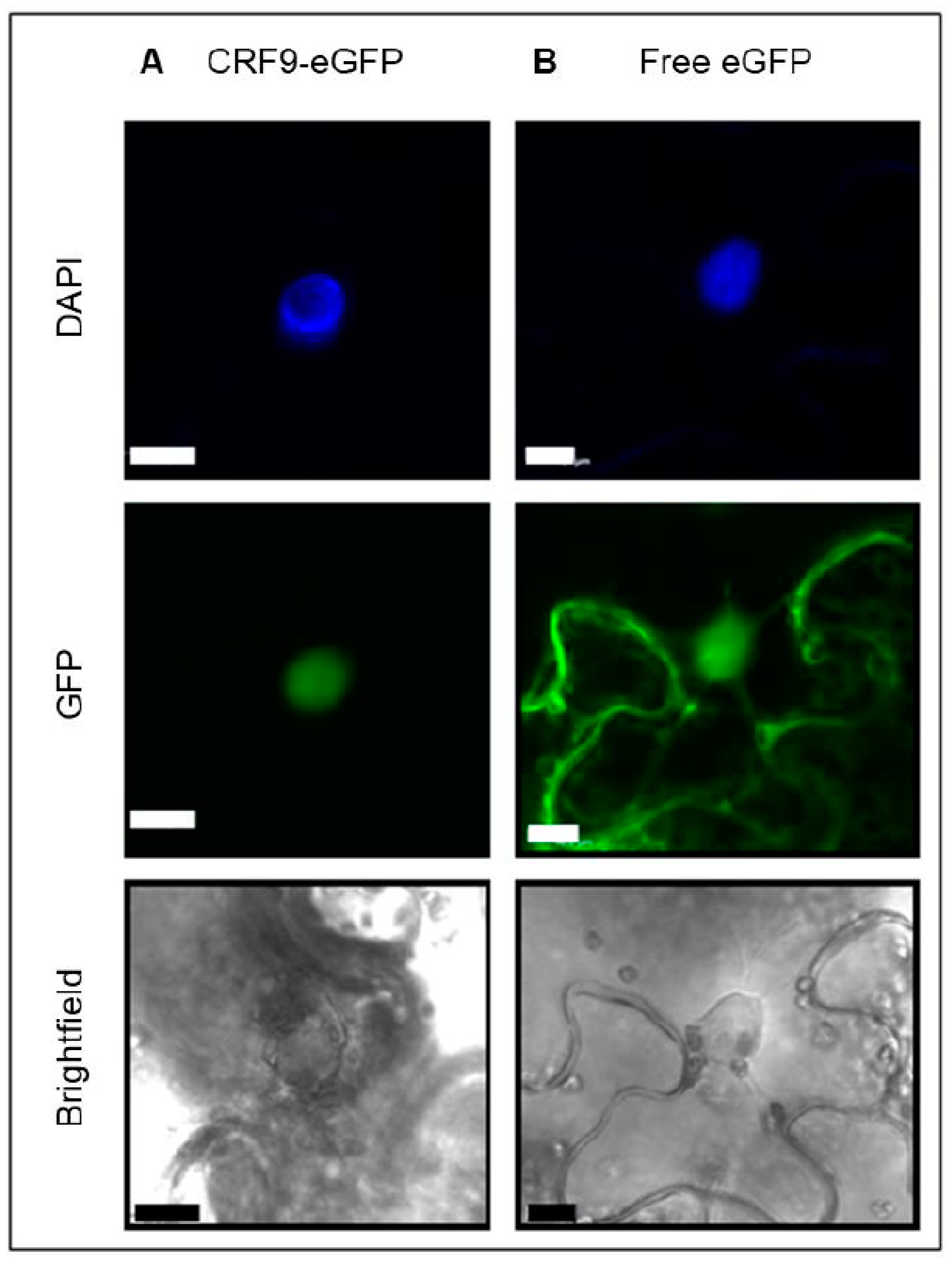
2.3. The Putative Transcription Factor CRF9 Is Located in the Nucleus
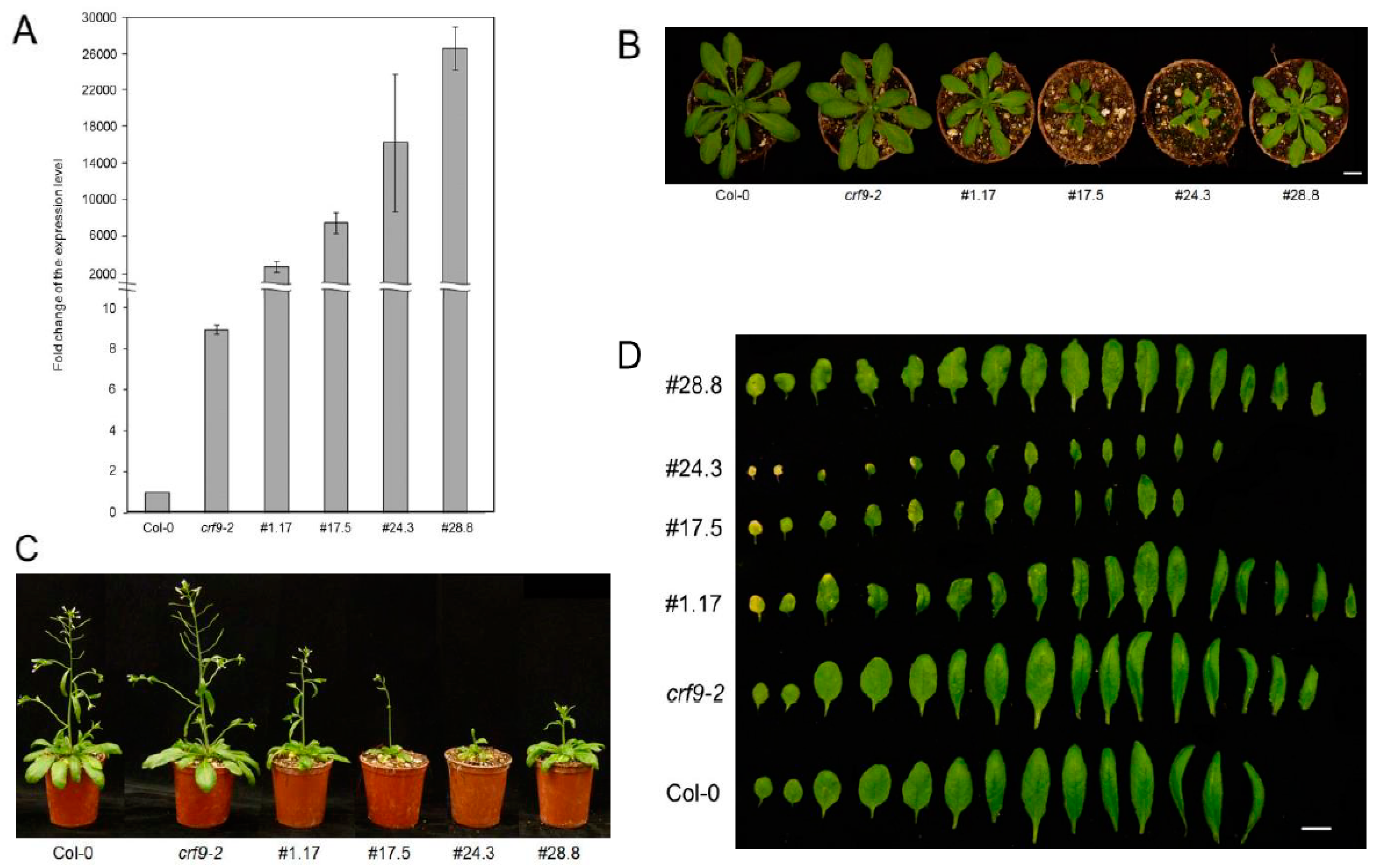
2.4. CRF9 Plays a Role in Plant Development
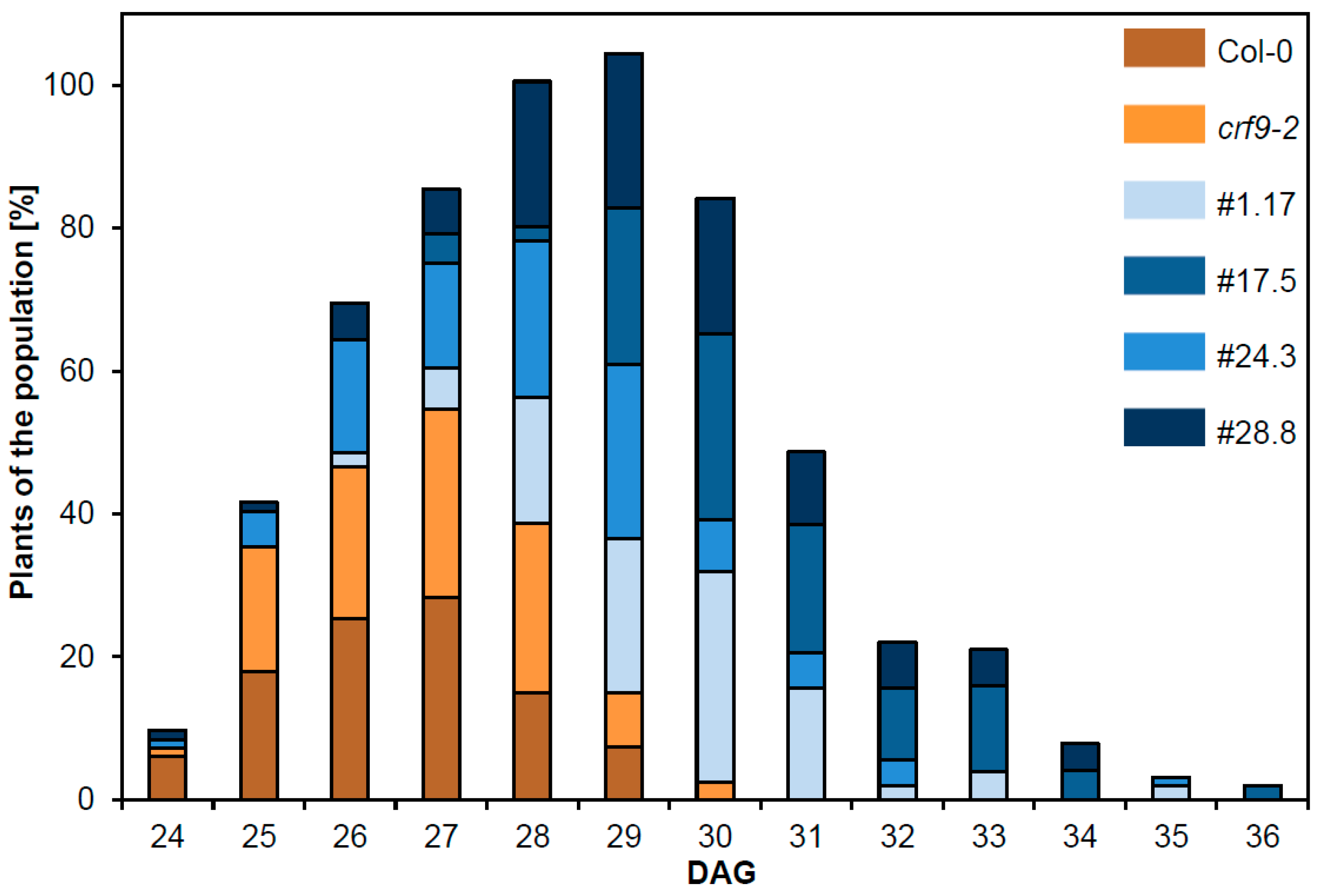
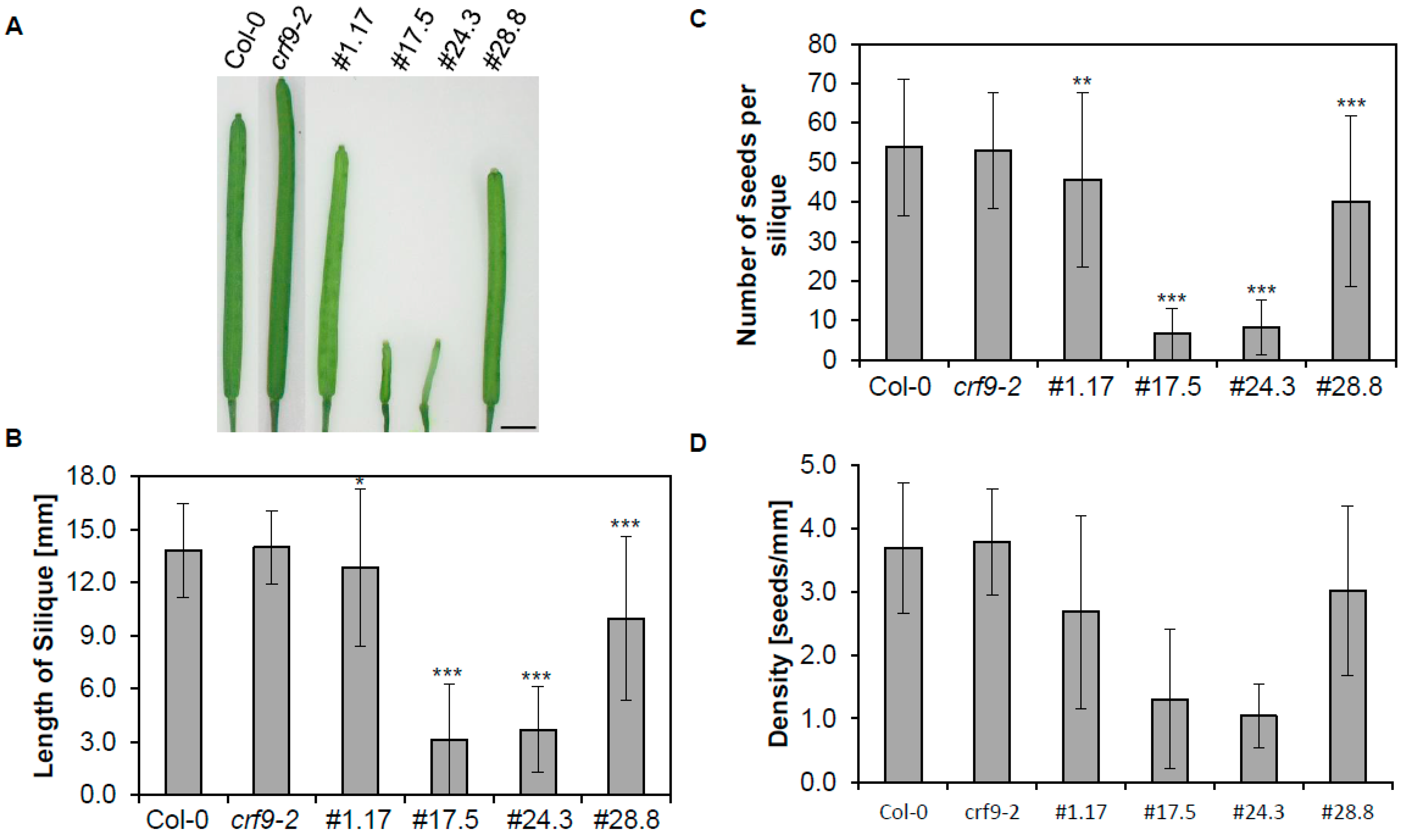
2.5. Reproductive Traits Are Affected by CRF9
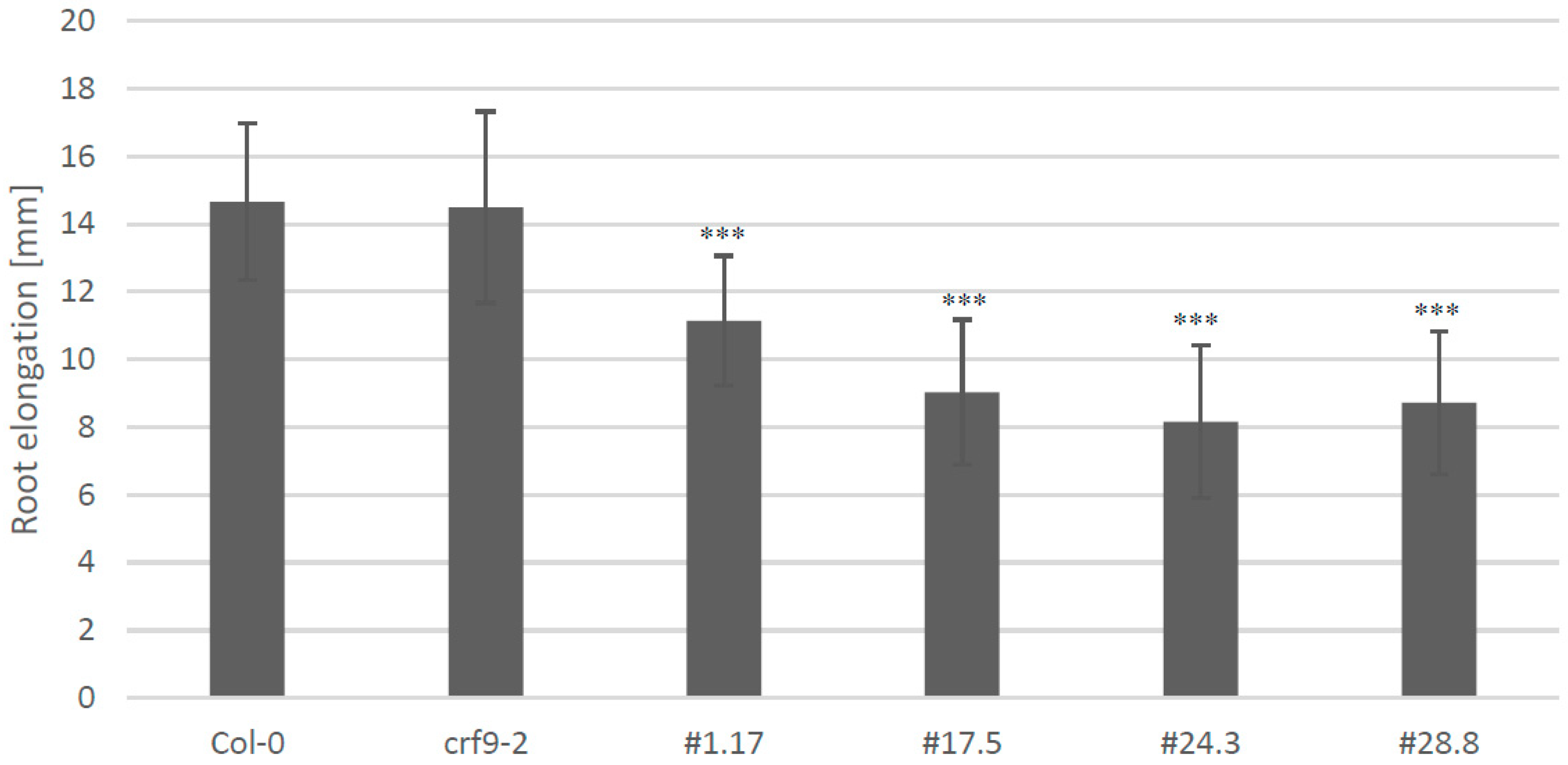
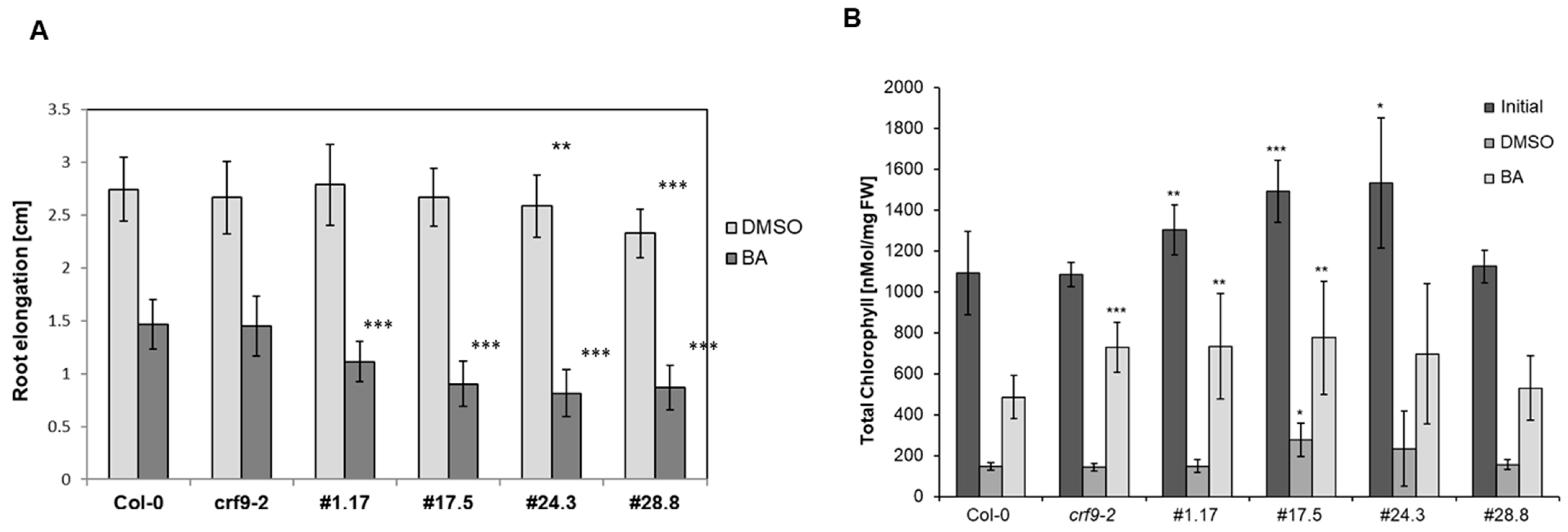
2.6. CRF9 Affects the Cytokinin Response in Roots
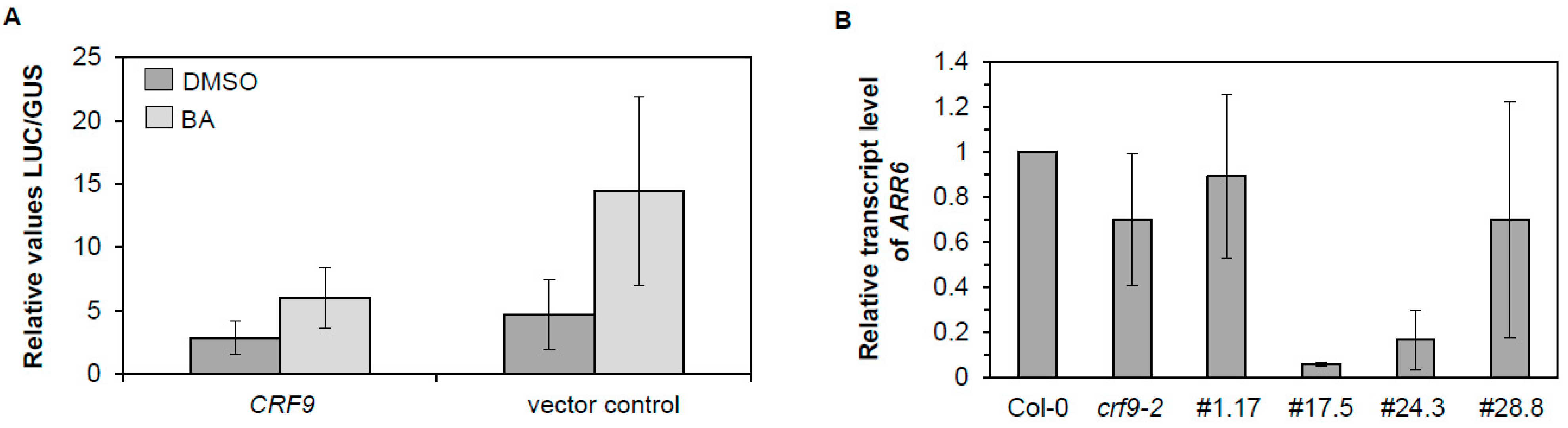
2.7. CRF9 Is a Transcriptional Repressor of ARR6
3. Discussion
3.1. CRF9 Plays a Role in Reproductive Growth
3.2. CRF9 Is a Transcriptional Regulator of the Cytokinin Response
3.3. CRF9 Belongs to a Branch of CRFs Specific for Modern Land Plants
4. Materials and Methods
4.1. Phylogenetic Analysis
4.2. Plant Materials and Growth Conditions
4.3. Transgenic Plants
4.4. Subcellular Localization in Nicotiana Benthamiana
4.5. Root Elongation Assay
4.6. Histochemical Analysis
4.7. Dark-Induced Senescence Analysis
4.8. Protoplast Transactivation Assay
4.9. Floral Transition Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mok, D.W.S.; Mok, M.C. Cytokinin Metabolism and Action. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef]
- Werner, T.; Schmülling, T. Cytokinin action in plant development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef]
- Wybouw, B.; de Rybel, B. Cytokinin—A Developing Story. Trends Plant Sci. 2019, 24, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Du, K.; Kang, X.; Wei, H. The diverse roles of cytokinins in regulating leaf development. Hortic. Res. 2021, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Svolacchia, N.; Sabatini, S. Cytokinins. Curr. Biol. 2023, 33, R10–R13. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. Book 2014, 12, e0168. [Google Scholar] [CrossRef] [Green Version]
- Keshishian, E.A.; Rashotte, A.M. Plant cytokinin signaling. Essays Biochem. 2015, 58, 13–27. [Google Scholar]
- To, J.P.C.; Kieber, J.J. Cytokinin signaling: Two components and more. Trends Plant Sci. 2008, 13, 85–92. [Google Scholar] [CrossRef]
- Hellmann, E.; Gruhn, N.; Heyl, A. The more, the merrier: Cytokinin signaling beyond Arabidopsis. Plant Signal. Behav. 2010, 5, 1384–1390. [Google Scholar] [CrossRef] [Green Version]
- Rashotte, A.M.; Mason, M.G.; Hutchison, C.E.; Ferreira, F.J.; Schaller, G.E.; Kieber, J.J. A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two-component pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 11081–11085. [Google Scholar] [CrossRef] [Green Version]
- Hallmark, H.T.; Rashotte, A.M. Review—Cytokinin Response Factors: Responding to more than cytokinin. Plant Sci. 2019, 289, 110251. [Google Scholar] [CrossRef]
- Hughes, A.M.; Hallmark, H.T.; Plačková, L.; Novák, O.; Rashotte, A.M. Clade III cytokinin response factors share common roles in response to oxidative stress responses linked to cytokinin synthesis. J. Exp. Bot. 2021, 72, 3294–3306. [Google Scholar] [CrossRef]
- Keshishian, E.A.; Cliver, B.R.; McLaughlin, W.F.; Hallmark, H.T.; Plačková, L.; Goertzen, L.R.; Novák, O.; Cobine, P.A.; Leisner, C.P.; Rashotte, A.M. CYTOKININ RESPONSE FACTOR 2 is involved in modulating the salt stress response. Plant J. 2022, 110, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Cutcliffe, J.W.J.W.; Hellmann, E.; Heyl, A.; Rashotte, A.M.A.M. CRFs form protein-protein interactions with each other and with members of the cytokinin signaling pathway in Arabidopsis via the CRF domain. J. Exp. Bot. 2011, 62, 4995–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuendorf, J.E.; Schmülling, T. Meeting at the DNA: Specifying cytokinin responses through transcription factor complex formation. Plants 2021, 10, 1458. [Google Scholar] [CrossRef] [PubMed]
- Zwack, P.J.; Robinson, B.R.; Risley, M.G.; Rashotte, A.M. Cytokinin Response Factor 6 Negatively Regulates Leaf Senescence and is Induced in Response to Cytokinin and Numerous Abiotic Stresses. Plant Cell Physiol. 2013, 54, 971–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwack, P.J.; De Clercq, I.; Howton, T.C.; Hallmark, H.T.; Hurny, A.; Keshishian, E.A.; Parish, A.M.; Benkova, E.; Mukhtar, M.S.; Van Breusegem, F.; et al. Cytokinin Response Factor 6 Represses Cytokinin-Associated Genes during Oxidative Stress. Plant Physiol. 2016, 172, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Powell, R.V.; Willett, C.R.; Goertzen, L.R.; Rashotte, A.M. Lineage-specific conservation of cis-regulatory elements in Cytokinin Response Factors. Sci. Rep. 2019, 9, 13387. [Google Scholar] [CrossRef] [Green Version]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Li, S.-M.; Zheng, H.-X.; Zhang, X.-S.; Sui, N. Cytokinins as central regulators during plant growth and stress response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Clabaugh, I.; To, J.P.; Maxwell, B.B.; Chiang, Y.-H.; Schaller, G.E.; Loraine, A.; Kieber, J.J. Identification of Cytokinin-Responsive Genes Using Microarray Meta-Analysis and RNA-Seq in Arabidopsis. Plant Physiol. 2013, 162, 272–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, W.G.; Schmülling, T. Transcript profiling of cytokinin action in Arabidopsis roots and shoots discovers largely similar but also organ-specific responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-Wide Analysis of the ERF Gene Family. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashotte, A.M.; Goertzen, L.R. The CRF domain defines cytokinin response factor proteins in plants. BMC Plant Biol. 2010, 10, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Xie, M.; Wu, Y.; Cui, X.; Tang, M.; Yang, L.; Xiang, Y.; Li, Y.; Bai, Z.; Huang, J.; et al. Genetic mapping and regional association analysis revealed a CYTOKININ RESPONSE FACTOR 10 gene controlling flowering time in Brassica napus L. Ind. Crops Prod. 2023, 193, 116239. [Google Scholar] [CrossRef]
- Hellmann, E.; Swinka, C.; Heyl, A. Novel in vivo screening design for the rapid and cost-effective identification of transcriptional regulators. Physiol. Plant. 2017, 160, 2–10. [Google Scholar] [CrossRef]
- Mason, M.G.; Mathews, D.E.; Argyros, D.A.; Maxwell, B.B.; Kieber, J.J.; Alonso, J.M.; Ecker, J.R.; Schaller, G.E. Multiple type-B response regulators mediate cytokinin signal transduction in Arabidopsis. Plant Cell 2005, 17, 3007–3018. [Google Scholar] [CrossRef] [Green Version]
- Kroll, C.K.; Brenner, W.G. Cytokinin Signaling Downstream of the His-Asp Phosphorelay Network: Cytokinin-Regulated Genes and Their Functions. Front. Plant Sci. 2020, 11, 604489. [Google Scholar] [CrossRef]
- Kim, J. CYTOKININ RESPONSE FACTORs Gating Environmental Signals and Hormones. Trends Plant Sci. 2016, 21, 993–996. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, H.; Shi, L.; Xu, F.; Ding, G. Genome-wide dissection of the CRF gene family in brassica napus indicates that bnacrf8s specifically regulate root architecture and phosphate homeostasis against phosphate fluctuation in plants. Int. J. Mol. Sci. 2020, 21, 3660. [Google Scholar] [CrossRef]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kaltenegger, E.; Leng, S.; Heyl, A. The effects of repeated whole genome duplication events on the evolution of cytokinin signaling pathway. BMC Evol. Biol. 2018, 18, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyl, A.; Ramireddy, E.; Brenner, W.G.G.; Riefler, M.; Allemeersch, J.; Schmülling, T.; Schmulling, T. The Transcriptional Repressor ARR1-SRDX Suppresses Pleiotropic Cytokinin Activities in Arabidopsis. Plant Physiol. 2008, 147, 1380–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, S.; Bartrina, I.; Schmülling, T. Cytokinin regulates vegetative phase change in Arabidopsis thaliana through the miR172/TOE1-TOE2 module. Nat. Commun. 2021, 12, 5816. [Google Scholar] [CrossRef]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Šimášková, M.; O’Brien, J.A.; Khan, M.; Van Noorden, G.; Ötvös, K.; Vieten, A.; De Clercq, I.; Van Haperen, J.M.A.; Cuesta, C.; Hoyerová, K.; et al. Cytokinin response factors regulate PIN-FORMED auxin transporters. Nat. Commun. 2015, 6, 8717. [Google Scholar] [CrossRef] [Green Version]
- Cucinotta, M.; Manrique, S.; Guazzotti, A.; Quadrelli, N.E.; Mendes, M.A.; Benkova, E.; Colombo, L. Cytokinin response factors integrate auxin and cytokinin pathways for female reproductive organ development. Development 2016, 143, 4419–4424. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kleinboelting, N.; Huep, G.; Kloetgen, A.; Viehoever, P.; Weisshaar, B. GABI-Kat SimpleSearch: New features of the Arabidopsis thaliana T-DNA mutant database. Nucleic Acids Res. 2012, 40, D1211–D1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapuscinski, J. DAPI: A DNA-Specific Fluorescent Probe. Biotech. Histochem. 1995, 70, 220–233. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Wu, J.; Liu, H.; Duan, X.; Ding, Y.; Wu, H.; Bai, Y.; Sun, X. Prediction of DNA-binding residues in proteins from amino acid sequences using a random forest model with a hybrid feature. Bioinformatics 2009, 25, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Cho, Y.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Christensen, A.H.; Quail, P.H. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgenic Res. 1996, 5, 213–218. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swinka, C.; Hellmann, E.; Zwack, P.; Banda, R.; Rashotte, A.M.; Heyl, A. Cytokinin Response Factor 9 Represses Cytokinin Responses in Flower Development. Int. J. Mol. Sci. 2023, 24, 4380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054380
Swinka C, Hellmann E, Zwack P, Banda R, Rashotte AM, Heyl A. Cytokinin Response Factor 9 Represses Cytokinin Responses in Flower Development. International Journal of Molecular Sciences. 2023; 24(5):4380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054380
Chicago/Turabian StyleSwinka, Christine, Eva Hellmann, Paul Zwack, Ramya Banda, Aaron M. Rashotte, and Alexander Heyl. 2023. "Cytokinin Response Factor 9 Represses Cytokinin Responses in Flower Development" International Journal of Molecular Sciences 24, no. 5: 4380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054380