
The Proteome of Circulating Large Extracellular Vesicles in Diabetes and Hypertension
 and
and
Abstract
:1. Introduction
2. Results
2.1. Physiological and Biochemical Measures
2.2. Characterization of EV Isolates
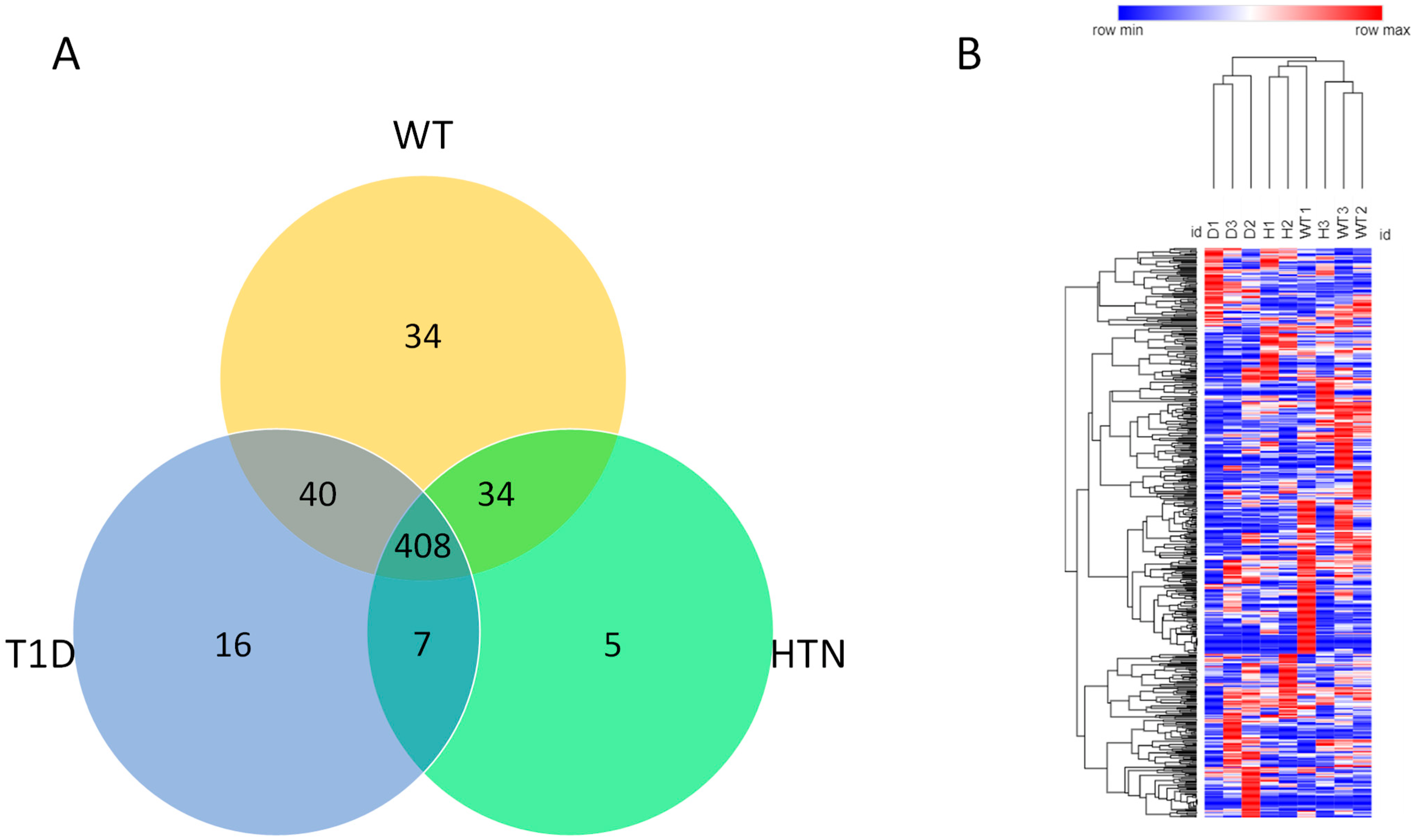
2.3. Proteomics Analysis and Associated Signaling Pathways
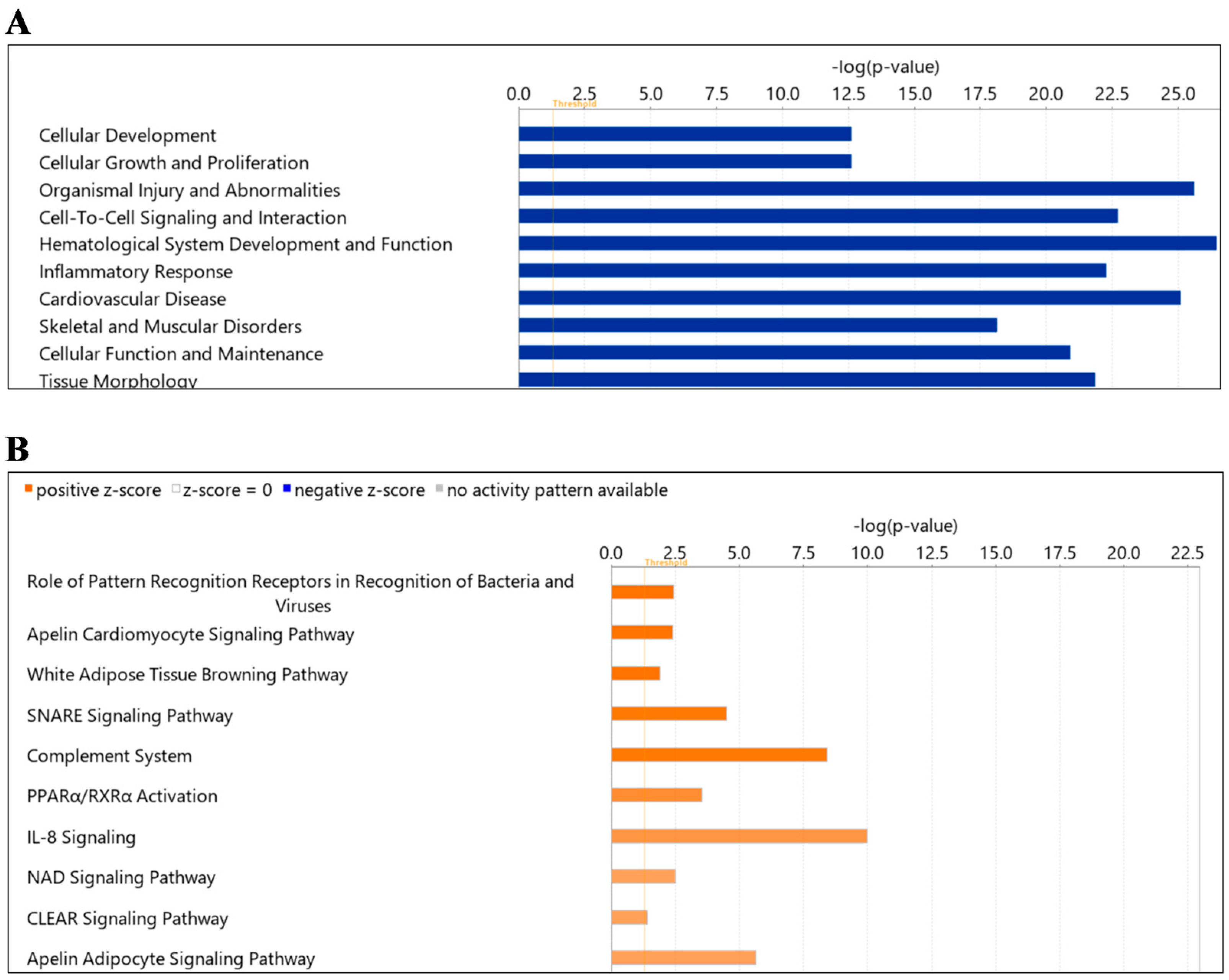
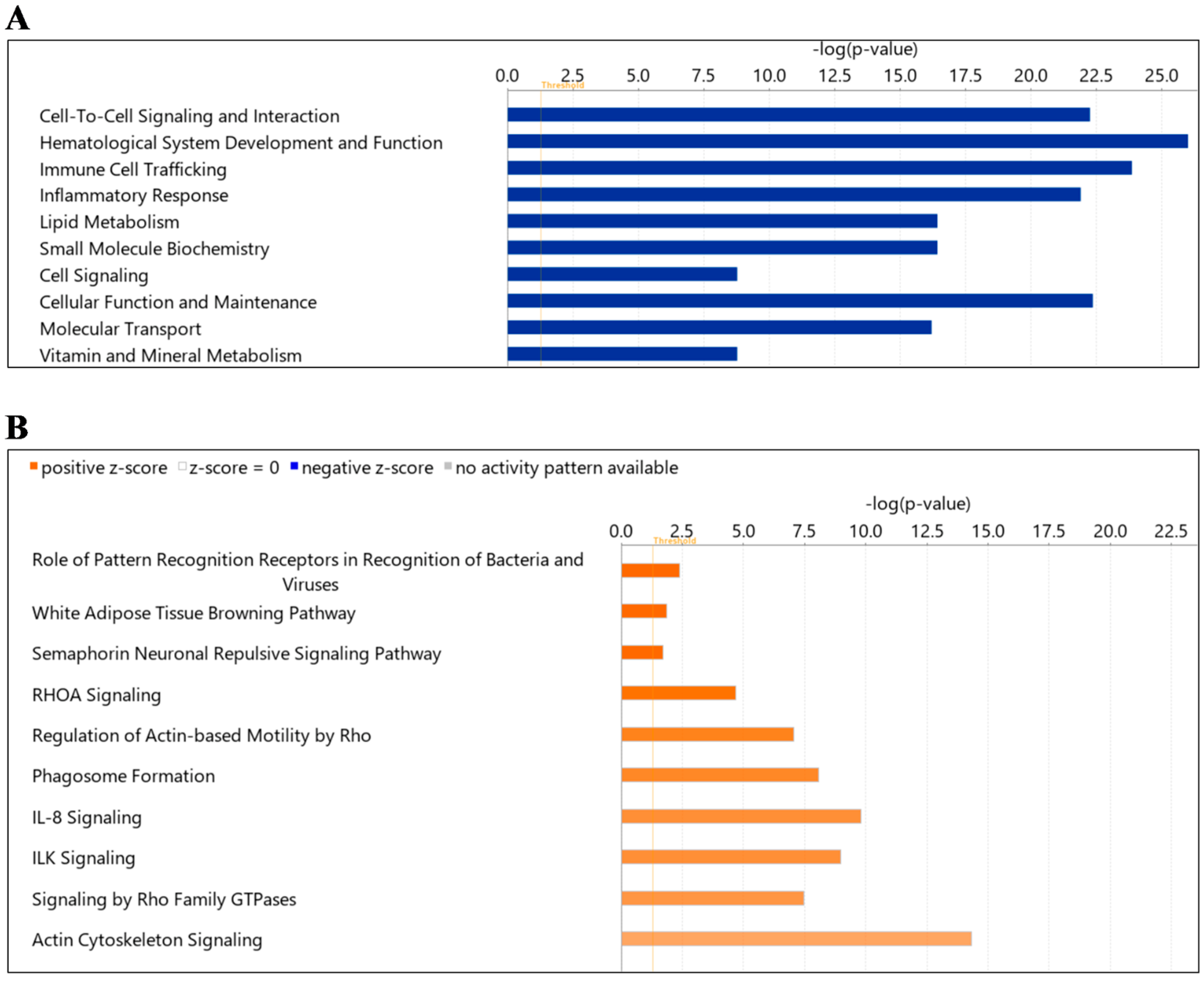
2.4. Protein Ingenuity Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Blood Pressure Measurement
4.3. Physiological Parameters
4.4. EV Isolation
4.5. Nanoparticle Tracking Analysis
4.6. Electron Microscopy
4.7. Western Blot Analysis
4.8. Proteomic Assessment of EVs
4.9. Bioinformatics Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strain, W.D.; Paldánius, P.M. Diabetes, Cardiovascular Disease and the Microcirculation. Cardiovasc. Diabetol. 2018, 17, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic Kidney Disease: Challenges, Progress, and Possibilities. Clin. J. Am. Soc. Nephrol. 2017, 12, 2032–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, P.; Ekmekcioglu, C. Impact of Salt Intake on the Pathogenesis and Treatment of Hypertension. Adv. Exp. Med. Biol. 2017, 956, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Pugh, D.; Gallacher, P.J.; Dhaun, N. Management of Hypertension in Chronic Kidney Disease. Drugs 2019, 79, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Cheung, B.M.Y.; Li, C. Diabetes and Hypertension: Is There a Common Metabolic Pathway? Curr. Atheroscler. Rep. 2012, 14, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.Z.; Jose, P.A.; Yang, J.; Zeng, C. Importance of Extracellular Vesicles in Hypertension. Exp. Biol. Med. 2021, 246, 342–353. [Google Scholar] [CrossRef]
- Xiao, Y.; Zheng, L.; Zou, X.; Wang, J.; Zhong, J.; Zhong, T. Extracellular Vesicles in Type 2 Diabetes Mellitus: Key Roles in Pathogenesis, Complications, and Therapy. J. Extracell. Vesicles 2019, 8, 1625677. [Google Scholar] [CrossRef] [Green Version]
- Burger, D.; Schock, S.; Thompson, C.S.; Montezano, A.C.; Hakim, A.M.; Touyz, R.M. Microparticles: Biomarkers and Beyond. Clin. Sci. 2013, 124, 423–441. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Buzas, E.I.; Bemis, L.T.; Bora, A.; Lasser, C.; Lotvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of Sample Collection, Isolation and Analysis Methods in Extracellular Vesicle Research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef]
- Rontogianni, S.; Synadaki, E.; Li, B.; Liefaard, M.C.; Lips, E.H.; Wesseling, J.; Wu, W.; Altelaar, M. Proteomic Profiling of Extracellular Vesicles Allows for Human Breast Cancer Subtyping. Commun. Biol. 2019, 2, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhadad, M.A.; Jonasson, C.; Huth, C.; Wilson, R.; Gieger, C.; Matias, P.; Grallert, H.; Graumann, J.; Gailus-Durner, V.; Rathmann, W.; et al. Deciphering the Plasma Proteome of Type 2 Diabetes. Diabetes 2020, 69, 2766–2778. [Google Scholar] [CrossRef] [PubMed]
- Zanini, J.C.; Pietzner, M.; Langenberg, C. Integrating Genetics and the Plasma Proteome to Predict the Risk of Type 2 Diabetes. Curr. Diabetes Rep. 2020, 20, 60. [Google Scholar] [CrossRef]
- Gummesson, A.; Björnson, E.; Fagerberg, L.; Zhong, W.; Tebani, A.; Edfors, F.; Schmidt, C.; Lundqvist, A.; Adiels, M.; Bäckhed, F.; et al. Longitudinal Plasma Protein Profiling of Newly Diagnosed Type 2 Diabetes. EBioMedicine 2021, 63, 103147. [Google Scholar] [CrossRef]
- Overgaard, A.J.; Hansen, H.G.; Lajer, M.; Pedersen, L.; Tarnow, L.; Rossing, P.; McGuire, J.N.; Pociot, F. Plasma Proteome Analysis of Patients with Type 1 Diabetes with Diabetic Nephropathy. Proteome Sci. 2010, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento de Oliveira, V.; Lima-Neto, A.B.M.; van Tilburg, M.F.; de Oliveira Monteiro-Moreira, A.C.; Duarte Pinto Lobo, M.; Rondina, D.; Fernandes, V.O.; Montenegro, A.P.D.R.; Montenegro, R.M.; Guedes, M.I.F. Proteomic Analysis to Identify Candidate Biomarkers Associated with Type 1 Diabetes. Diabetes Metab. Syndr. Obes. 2018, 11, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Gajjala, P.R.; Jankowski, V.; Heinze, G.; Bilo, G.; Zanchetti, A.; Noels, H.; Liehn, E.; Perco, P.; Schulz, A.; Delles, C.; et al. Proteomic-Biostatistic Integrated Approach for Finding the Underlying Molecular Determinants of Hypertension in Human Plasma. Hypertension 2017, 70, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Zhai, D.; Cabezas, E.; Welsh, K.; Nouraini, S.; Satterthwait, A.C.; Reed, J.C. Humanin Peptide Suppresses Apoptosis by Interfering with Bax Activation. Nature 2003, 423, 456–461. [Google Scholar] [CrossRef]
- Yen, K.; Lee, C.; Mehta, H.; Cohen, P. The Emerging Role of the Mitochondrial-Derived Peptide Humanin in Stress Resistance. J. Mol. Endocrinol. 2013, 50, R11–R19. [Google Scholar] [CrossRef] [Green Version]
- Pena, M.J.; Jankowski, J.; Heinze, G.; Kohl, M.; Heinzel, A.; Bakker, S.J.L.; Gansevoort, R.T.; Rossing, P.; de Zeeuw, D.; Heerspink, H.J.L.; et al. Plasma Proteomics Classifiers Improve Risk Prediction for Renal Disease in Patients with Hypertension or Type 2 Diabetes. J. Hypertens. 2015, 33, 2123–2132. [Google Scholar] [CrossRef]
- Xu, M.-D.; Wu, X.-Z.; Zhou, Y.; Xue, Y.; Zhang, K.-Q. Proteomic Characteristics of Circulating Microparticles in Patients with Newly-Diagnosed Type 2 Diabetes. Am. J. Transl. Res. 2016, 8, 209–220. [Google Scholar]
- Jayabalan, N.; Lai, A.; Nair, S.; Guanzon, D.; Scholz-Romero, K.; Palma, C.; McIntyre, H.D.; Lappas, M.; Salomon, C. Quantitative Proteomics by SWATH-MS Suggest an Association between Circulating Exosomes and Maternal Metabolic Changes in Gestational Diabetes Mellitus. Proteomics 2019, 19, e1800164. [Google Scholar] [CrossRef] [Green Version]
- Zierfuss, B.; Höbaus, C.; Herz, C.T.; Pesau, G.; Koppensteiner, R.; Schernthaner, G.-H. Thrombospondin-4 Increases with the Severity of Peripheral Arterial Disease and is Associated with Diabetes. Heart Vessels 2020, 35, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Razzaque, M.S.; Koji, T.; Horita, Y.; Nishihara, M.; Harada, T.; Nakane, P.K.; Taguchi, T. Synthesis of Type III Collagen and Type IV Collagen by Tubular Epithelial Cells in Diabetic Nephropathy. Pathol. Res. Pract. 1995, 191, 1099–1104. [Google Scholar] [CrossRef]
- Hämäläinen, P.; Saltevo, J.; Kautiainen, H.; Mäntyselkä, P.; Vanhala, M. Erythropoietin, Ferritin, Haptoglobin, Hemoglobin and Transferrin Receptor in Metabolic Syndrome: A Case Control Study. Cardiovasc. Diabetol. 2012, 11, 116. [Google Scholar] [CrossRef] [Green Version]
- Yassine, H.N.; Trenchevska, O.; He, H.; Borges, C.R.; Nedelkov, D.; Mack, W.; Kono, N.; Koska, J.; Reaven, P.D.; Nelson, R.W. Serum Amyloid a Truncations in Type 2 Diabetes Mellitus. PLoS ONE 2015, 10, e0115320. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.-C.; Zhang, Z.-Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the Apelin Pathway as a Novel Therapeutic Approach for Cardiovascular Diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin-APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef]
- Gaisano, H.Y. Recent New Insights into the Role of SNARE and Associated Proteins in Insulin Granule Exocytosis. Diabetes Obes. Metab. 2017, 19 (Suppl. S1), 115–123. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Khan, A.; Ostenson, C.-G.; Berggren, P.-O.; Efendic, S.; Meister, B. Down-Regulated Expression of Exocytotic Proteins in Pancreatic Islets of Diabetic GK Rats. Biochem. Biophys. Res. Commun. 2002, 291, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, S.; Nakamichi, Y.; Yamamura, C.; Matsushima, S.; Watanabe, T.; Ozawa, S.; Furukawa, H.; Ishida, H. Decreased Expression of T-SNARE, Syntaxin 1, and SNAP-25 in Pancreatic Beta-Cells is Involved in Impaired Insulin Secretion from Diabetic GK Rat Islets: Restoration of Decreased t-SNARE Proteins Improves Impaired Insulin Secretion. Diabetes 1999, 48, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Ostenson, C.-G.; Gaisano, H.; Sheu, L.; Tibell, A.; Bartfai, T. Impaired Gene and Protein Expression of Exocytotic Soluble N-Ethylmaleimide Attachment Protein Receptor Complex Proteins in Pancreatic Islets of Type 2 Diabetic Patients. Diabetes 2006, 55, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.B.; MacPhail, R.M.; Sheu, L.; Wheeler, M.B.; Gaisano, H.Y. Beta-Cell Hypertrophy in Fa/Fa Rats is Associated with Basal Glucose Hypersensitivity and Reduced SNARE Protein Expression. Diabetes 1999, 48, 997–1005. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Tian, Y.; Huang, W.; Tong, N.; Fu, X. Integrative Biology of Extracellular Vesicles in Diabetes Mellitus and Diabetic Complications. Theranostics 2022, 12, 1342–1372. [Google Scholar] [CrossRef]
- Chung, S.; Park, C.W. Role of Peroxisome Proliferator-Activated Receptor α in Diabetic Nephropathy. Diabetes Metab. J. 2011, 35, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Gu, Z. The Role of Peroxisome Proliferator-Activated Receptors in Kidney Diseases. Front. Pharmacol. 2022, 13, 832732. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, H.W.; Ko, S.H.; Chung, H.W.; Lim, S.W.; Yang, C.W.; Chang, Y.S.; Sugawara, A.; Guan, Y.; Breyer, M.D. Accelerated Diabetic Nephropathy in Mice Lacking the Peroxisome Proliferator-Activated Receptor Alpha. Diabetes 2006, 55, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Cacicedo, J.M.; Ido, Y. Impaired Nicotinamide Adenine Dinucleotide (NAD+) Metabolism in Diabetes and Diabetic Tissues: Implications for Nicotinamide-Related Compound Treatment. J. Diabetes Investig. 2020, 11, 1403–1419. [Google Scholar] [CrossRef]
- Graw, J.A.; Yu, B.; Rezoagli, E.; Warren, H.S.; Buys, E.S.; Bloch, D.B.; Zapol, W.M. Endothelial Dysfunction Inhibits the Ability of Haptoglobin to Prevent Hemoglobin-Induced Hypertension. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1120–H1127. [Google Scholar] [CrossRef] [Green Version]
- Wirth, A. Rho Kinase and Hypertension. Biochim. Biophys. Acta 2010, 1802, 1276–1284. [Google Scholar] [CrossRef]
- Lee, D.L.; Webb, R.C.; Jin, L. Hypertension and RhoA/Rho-Kinase Signaling in the Vasculature: Highlights from the Recent Literature. Hypertension 2004, 44, 796–799. [Google Scholar] [CrossRef]
- Savoia, C.; Tabet, F.; Yao, G.; Schiffrin, E.L.; Touyz, R.M. Negative Regulation of RhoA/Rho Kinase by Angiotensin II Type 2 Receptor in Vascular Smooth Muscle Cells: Role in Angiotensin II-Induced Vasodilation in Stroke-Prone Spontaneously Hypertensive Rats. J. Hypertens. 2005, 23, 1037–1045. [Google Scholar] [CrossRef]
- Sit, S.-T.; Manser, E. Rho GTPases and Their Role in Organizing the Actin Cytoskeleton. J. Cell Sci. 2011, 124, 679–683. [Google Scholar] [CrossRef] [Green Version]
- Olazabal, I.M.; Caron, E.; May, R.C.; Schilling, K.; Knecht, D.A.; Machesky, L.M. Rho-Kinase and Myosin-II Control Phagocytic Cup Formation during CR, but Not FcgammaR, Phagocytosis. Curr. Biol. 2002, 12, 1413–1418. [Google Scholar] [CrossRef] [Green Version]
- Alique, M.; Civantos, E.; Sanchez-Lopez, E.; Lavoz, C.; Rayego-Mateos, S.; Rodrigues-Díez, R.; García-Redondo, A.B.; Egido, J.; Ortiz, A.; Rodríguez-Puyol, D.; et al. Integrin-Linked Kinase Plays a Key Role in the Regulation of Angiotensin II-Induced Renal Inflammation. Clin. Sci. 2014, 127, 19–31. [Google Scholar] [CrossRef]
- Lu, H.; Fedak, P.W.M.; Dai, X.; Du, C.; Zhou, Y.-Q.; Henkelman, M.; Mongroo, P.S.; Lau, A.; Yamabi, H.; Hinek, A.; et al. Integrin-Linked Kinase Expression is Elevated in Human Cardiac Hypertrophy and Induces Hypertrophy in Transgenic Mice. Circulation 2006, 114, 2271–2279. [Google Scholar] [CrossRef] [Green Version]
- Clancy, J.W.; Schmidtmann, M.; D’Souza-Schorey, C. The Ins and Outs of Microvesicles. FASEB BioAdvances 2021, 3, 399–406. [Google Scholar] [CrossRef]
- Sun, Y.; Tao, Q.; Wu, X.; Zhang, L.; Liu, Q.; Wang, L. The Utility of Exosomes in Diagnosis and Therapy of Diabetes Mellitus and Associated Complications. Front. Endocrinol. 2021, 12, 756581. [Google Scholar] [CrossRef]
- Arishe, O.O.; Priviero, F.; Wilczynski, S.A.; Webb, R.C. Exosomes as Intercellular Messengers in Hypertension. Int. J. Mol. Sci. 2021, 22, 11685. [Google Scholar] [CrossRef]
- Thibodeau, J.F.; Holterman, C.E.; Burger, D.; Read, N.C.; Reudelhuber, T.L.; Kennedy, C.R. A Novel Mouse Model of Advanced Diabetic Kidney Disease. PLoS ONE 2014, 9, e113459. [Google Scholar] [CrossRef] [PubMed]
- Burger, D.; Reudelhuber, T.L.; Mahajan, A.; Chibale, K.; Sturrock, E.D.; Touyz, R.M. Effects of a Domain-Selective ACE Inhibitor in a Mouse Model of Chronic Angiotensin II-Dependent Hypertension. Clin. Sci. 2014, 127, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Prescott, G.; Silversides, D.W.; Chiu, S.M.; Reudelhuber, T.L. Contribution of Circulating Renin to Local Synthesis of Angiotensin Peptides in the Heart. Physiol. Genom. 2000, 4, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, P.N.; Overbeek, P.A.; Means, A.R. Calmodulin-Induced Early-Onset Diabetes in Transgenic Mice. Cell 1989, 58, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Burger, D.; Thibodeau, J.-F.; Holterman, C.E.; Burns, K.D.; Touyz, R.M.; Kennedy, C.R.J. Urinary Podocyte Microparticles Identify Prealbuminuric Diabetic Glomerular Injury. J. Am. Soc. Nephrol. 2014, 25, 1401–1407. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, M.; Xiao, F.; Abujrad, H.; Al-Rewashdy, Y.; Tang, V.A.; Langlois, M.-A.; Sorisky, A.; Ooi, T.C.; Burger, D. Effect of Hemodialysis on Extracellular Vesicles and Circulating Submicron Particles. BMC Nephrol. 2019, 20, 294. [Google Scholar] [CrossRef] [Green Version]
- Akbari, S.; Abou-Arkoub, R.; Sun, S.; Hiremath, S.; Reunov, A.; McCormick, B.B.; Ruzicka, M.; Burger, D. Microparticle Formation in Peritoneal Dialysis. Can. J. Kidney Health Dis. 2017, 4, 2054358117699829. [Google Scholar] [CrossRef]
- Mohammad, S.; Hutchinson, K.A.; da Silva, D.F.; Bhattacharjee, J.; McInnis, K.; Burger, D.; Adamo, K.B. Circulating Small Extracellular Vesicles Increase after an Acute Bout of Moderate-Intensity Exercise in Pregnant Compared to Non-Pregnant Women. Sci. Rep. 2021, 11, 12615. [Google Scholar] [CrossRef]
- Lytvyn, Y.; Xiao, F.; Kennedy, C.R.J.; Perkins, B.A.; Reich, H.N.; Scholey, J.W.; Cherney, D.Z.; Burger, D. Assessment of Urinary Microparticles in Normotensive Patients with Type 1 Diabetes. Diabetologia 2017, 60, 581–584. [Google Scholar] [CrossRef] [Green Version]
- Burger, D.; Montezano, A.C.; Nishigaki, N.; He, Y.; Carter, A.; Touyz, R.M. Endothelial Microparticle Formation by Angiotensin II Is Mediated via Ang II Receptor Type I/NADPH Oxidase/Rho Kinase Pathways Targeted to Lipid Rafts. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1898–1907. [Google Scholar] [CrossRef] [Green Version]
- Abolbaghaei, A.; Mohammad, S.; da Silva, D.F.; Hutchinson, K.A.; Myette, R.L.; Adamo, K.B.; Burger, D. Impact of Acute Moderate-Intensity Aerobic Exercise on Circulating Extracellular Vesicles in Pregnant and Non-Pregnant Women. Appl. Physiol. Nutr. Metab. 2023, 48, 198–208. [Google Scholar] [CrossRef]
- Burger, D.; Turner, M.; Xiao, F.; Munkonda, M.N.; Akbari, S.; Burns, K.D. High Glucose Increases the Formation and Pro-Oxidative Activity of Endothelial Microparticles. Diabetologia 2017, 60, 1791–1800. [Google Scholar] [CrossRef] [Green Version]
- Benito-Martin, A.; Peinado, H. FunRich Proteomics Software Analysis, Let the Fun Begin! Proteomics 2015, 15, 2555–2556. [Google Scholar] [CrossRef]
- Goedhart, J.; Luijsterburg, M.S. VolcaNoseR Is a Web App for Creating, Exploring, Labeling and Sharing Volcano Plots. Sci. Rep. 2020, 10, 20560. [Google Scholar] [CrossRef]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive Statistics for Studies in Sports Medicine and Exercise Science. Med. Sci. Sports Exerc. 2009, 41, 3–13. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Background | Blood Pressure (mmHg) | Blood Glucose (mg/dL) | Heart Weight/Tibia Length (mg/mm) | Urinary Albumin/Creatinine (ug/mg) | Body Weight (g) |
|---|---|---|---|---|---|
| Wild-type | 114 ± 6 | 11.3 ± 0.7 | 8.4 ± 0.1 | 245 ± 69 | 32.4 ± 1.2 |
| OVE26 | 124 ± 5 | 29.9 ± 0.8 * | 7.3 ± 0.5 | 1026 ± 204 * | 27.3 ± 0.9 * |
| TTRhRen | 144 ± 8 * | 12.3 ± 1.0 | 9.3 ± 0.6 | 504 ± 166 | 32.3 ± 1.1 |
| Protein | Change | Fold Change (log2) | p Value |
|---|---|---|---|
| TSP4 | ↑ | 1.72 | 0.0018 |
| HPT | ↑ | 1.59 | 0.00099 |
| CO3A1 | ↑ | 1.51 | 0.0042 |
| ANK1 | ↓ | −1.51 | 0.0062 |
| SAA4 | ↓ | −1.78 | 0.028 |
| Protein | Change | Fold Change (log2) | p Value |
|---|---|---|---|
| HPT | ↑ | 2.40 | 0.00099 |
| PPN | ↑ | 1.70 | 0.0022 |
| ANK1 | ↓ | −1.87 | 0.0041 |
| SPTB1 | ↓ | −2.18 | 0.028 |
| SPTA1 | ↓ | −1.78 | 0.026 |
| Protein | Change | Fold Change (log2) | p Value |
|---|---|---|---|
| IGHA | ↑ | 3.51 | 0.0003 |
| TSP4 | ↑ | 3.53 | 0.0003 |
| CLC1B | ↑ | 1.84 | 0.0146 |
| HVM17 | ↑ | 3.01 | 0.001 |
| CO3A1 | ↑ | 1.93 | 0.0117 |
| PSA3 | ↑ | 1.57 | 0.027 |
| PSA7 | ↑ | 2.45 | 0.00359 |
| PMGE | ↑ | 1.67 | 0.02148 |
| SAMP | ↓ | −2.06 | 0.11502 |
| SAA4 | ↓ | −1.65 | 0.02237 |
| ZPI | ↓ | −2.67 | 0.00215 |
| Canonical Pathway | Log (p Value) | Ratio | Z-Score | Proteins |
|---|---|---|---|---|
| Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses | 2.45 | 0.0321 | 2 | C1QA, C1QB, C1QC, MBL2, TGFB1 |
| Apelin Cardiomyocyte Signaling Pathway | 2.40 | 0.0404 | 2 | GNAI2, MYL6, MYL9, TGFB1 |
| White Adipose Tissue Browning Pathway | 1.90 | 0.029 | 2 | CAMP, LDHA, LDHB, THRB |
| SNARE Signaling Pathway | 4.51 | 0.0515 | 1.89 | MYH10, MYH14, MYH9, MYL6, MYL9, RAB6A, RAB7A |
| Complement System | 8.43 | 0.189 | 1.89 | C1QA, C1QB, C1QC, C4BPA, MASP1, MASP2, MBL2 |
| PPARα/RXRα Activation | 3.53 | 0.0359 | 1.633 | ACADL, APOA1, APOA2, GNAQ, RAP1A, RAP1B, TGFB1 |
| IL-8 Signaling | 9.97 | 0.0667 | 1.508 | CDC42, EGFR, GNA13, GNAI2, GNAQ, GNAZ, RAC1, RAC2, RAP1A, RAP1B, RHOA, VCAM1 |
| NAD Signaling Pathway | 2.51 | 0.0331 | 1.342 | ACADL, ACADM, LDHA, LDHB, TGFB1 |
| CLEAR Signaling Pathway | 1.41 | 0.0175 | 1.342 | EGFR, RAB7A, RAP1A, RAP1B, TGFB1 |
| Apelin Adipocyte Signaling Pathway | 5.66 | 0.0769 | 1.342 | GNAI2, GPX1, GPX3, GSTM1, RAC1, RAC2, SOD1 |
| Canonical Pathway | Log (p Value) | Ratio | Z-Score | Proteins |
|---|---|---|---|---|
| Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses | 2.39 | 0.0321 | 2 | C1QA, C1QB, C1QC, MBL2, TGFB1 |
| White Adipose Tissue Browning Pathway | 1.85 | 0.029 | 2 | CAMP, LDHA, LDHB, THRB |
| Semaphorin Neuronal Repulsive Signaling Pathway | 1.72 | 0.0265 | 2 | MYL6, MYL9, RAC1, RHOA |
| RHOA Signaling | 4.68 | 0.0565 | 1.89 | ARPC2, ARPC4, ARPC5, GNA13, MYL6, MYL9, RHOA |
| Regulation of Actin-based Motility by Rho | 7.04 | 0.0776 | 1.667 | ARPC2, ARPC4, ARPC5, CDC42, MYL6, MYL9, RAC1, RAC2, RHOA |
| Phagosome Formation | 8.09 | 0.0304 | 1.606 | ARPC2, ARPC4, ARPC5, CDC42, IGHE, IGHG3, IGHM, IGKC, LCAT, MYH10, MYH11, MYH14, MYH9, MYL6, MYL9, RAC1, RAC2, RAP1A, RAP1B, RHOA, TLN1 |
| IL-8 Signaling | 9.79 | 0.0667 | 1.508 | CDC42, EGFR, GNA13, GNAI2, GNAQ, GNAZ, MMP2, MYL9, RAC1, RAC2, RAP1A, RAP1B, RHOA, VCAM1 |
| ILK Signaling | 8.99 | 0.065 | 1.508 | CDC42, FLNA, ILK, MYH10, MYH11, MYH14, MYH9, MYL6, MYL9, PARVB, RAC1, RAC2, RHOA |
| Signaling by Rho Family GTPases | 7.46 | 0.0485 | 1.414 | ARPC2, ARPC4, ARPC5, CDC42, GNA13, GNAI2, GNAQ, GNAZ, MYL6, MYL9, RAC1, RAC2, RHOA |
| Actin Cytoskeleton Signaling | 14.30 | 0.0776 | 1.213 | ARPC2, ARPC4, ARPC5, CDC42, FLNA, GNA13, KNG1, MYH10, MYH11, MYH14, MYH9, MYL6, MYL9, RAC1, RAC2, RAP1A, RAP1B, RHOA, TLN1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abolbaghaei, A.; Turner, M.; Thibodeau, J.-F.; Holterman, C.E.; Kennedy, C.R.J.; Burger, D. The Proteome of Circulating Large Extracellular Vesicles in Diabetes and Hypertension. Int. J. Mol. Sci. 2023, 24, 4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054930
Abolbaghaei A, Turner M, Thibodeau J-F, Holterman CE, Kennedy CRJ, Burger D. The Proteome of Circulating Large Extracellular Vesicles in Diabetes and Hypertension. International Journal of Molecular Sciences. 2023; 24(5):4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054930
Chicago/Turabian StyleAbolbaghaei, Akram, Maddison Turner, Jean-François Thibodeau, Chet E. Holterman, Christopher R. J. Kennedy, and Dylan Burger. 2023. "The Proteome of Circulating Large Extracellular Vesicles in Diabetes and Hypertension" International Journal of Molecular Sciences 24, no. 5: 4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054930