

Stress Management in Plants: Examining Provisional and Unique Dose-Dependent Responses



Department of Molecular Biology and Genetics, Laboratory of Regulation of Gene Expression, Institute of Plant Physiology and Genetics, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(6), 5105; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065105
Submission received: 11 February 2023
/
Revised: 5 March 2023
/
Accepted: 6 March 2023
/
Published: 7 March 2023
(This article belongs to the Special Issue Response to Environmental Stress in Plants)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The purpose of this review is to critically evaluate the effects of different stress factors on higher plants, with particular attention given to the typical and unique dose-dependent responses that are essential for plant growth and development. Specifically, this review highlights the impact of stress on genome instability, including DNA damage and the molecular, physiological, and biochemical mechanisms that generate these effects. We provide an overview of the current understanding of predictable and unique dose-dependent trends in plant survival when exposed to low or high doses of stress. Understanding both the negative and positive impacts of stress responses, including genome instability, can provide insights into how plants react to different levels of stress, yielding more accurate predictions of their behavior in the natural environment. Applying the acquired knowledge can lead to improved crop productivity and potential development of more resilient plant varieties, ensuring a sustainable food source for the rapidly growing global population.
1. Introduction
Climate change is a multifaceted phenomenon that affects plant and animal species, as well as their habitats and ecosystems due to altered weather patterns and an increased frequency of extreme weather events. It also contributes to the spread of pests and diseases [1]. Being rooted in one place, plants are highly sensitive to fluctuations in temperature, rainfall, radiation, and other environmental factors, which induce a range of short-term or medium-term reactions, such as the acclimation processes, or long-term phenomena, such as transgenerational adaptation [2]. These reactions can impact the physiological state of plants, affecting their growth and development and leading to reduced seed production and germination, decreased nutrient uptake, and water use efficiency, increased vulnerability to pests and diseases and, in extreme cases, death [3]. The kinetics of the normal biological response is contingent on the intensity and duration of the stressor (acute or chronic) [4]. Plants have developed intricate defense mechanisms to withstand natural adversities and secure their survival against various challenges [5]. Altering the anatomy and morphology of plants through modification of their structural features is a potential strategy for combating the consequences of climate change [6,7,8]. At the genetic level, plants have developed mechanisms to repair or tolerate DNA damage, allowing them to maintain genome integrity [3,9]. A recent review by Hartmann et al. [2] traces the impact of different stressors on the stability of plant genome, focusing on the crucial underlying mechanisms. They indicate that environmental stress can cause changes in plant DNA, including mutations, epigenetic modifications, and alterations in gene expression. These changes can have a profound impact on plant growth, development, and ability to adapt to changing conditions. The crucial role of the potential interplay between genetic and epigenetic elements in understanding plant stress responses has been discussed. The effects of environmental stimuli on plants can sometimes be atypical and multifaceted, requiring more comprehensive analysis.
This review endeavors to give an insight into current knowledge of plant genome stability, with particular emphasis on the provisional and unique dose-dependent stress responses that are essential in determining plant growth and development when exposed to low or high stress levels.
2. Principal Stressors Impacting Plants
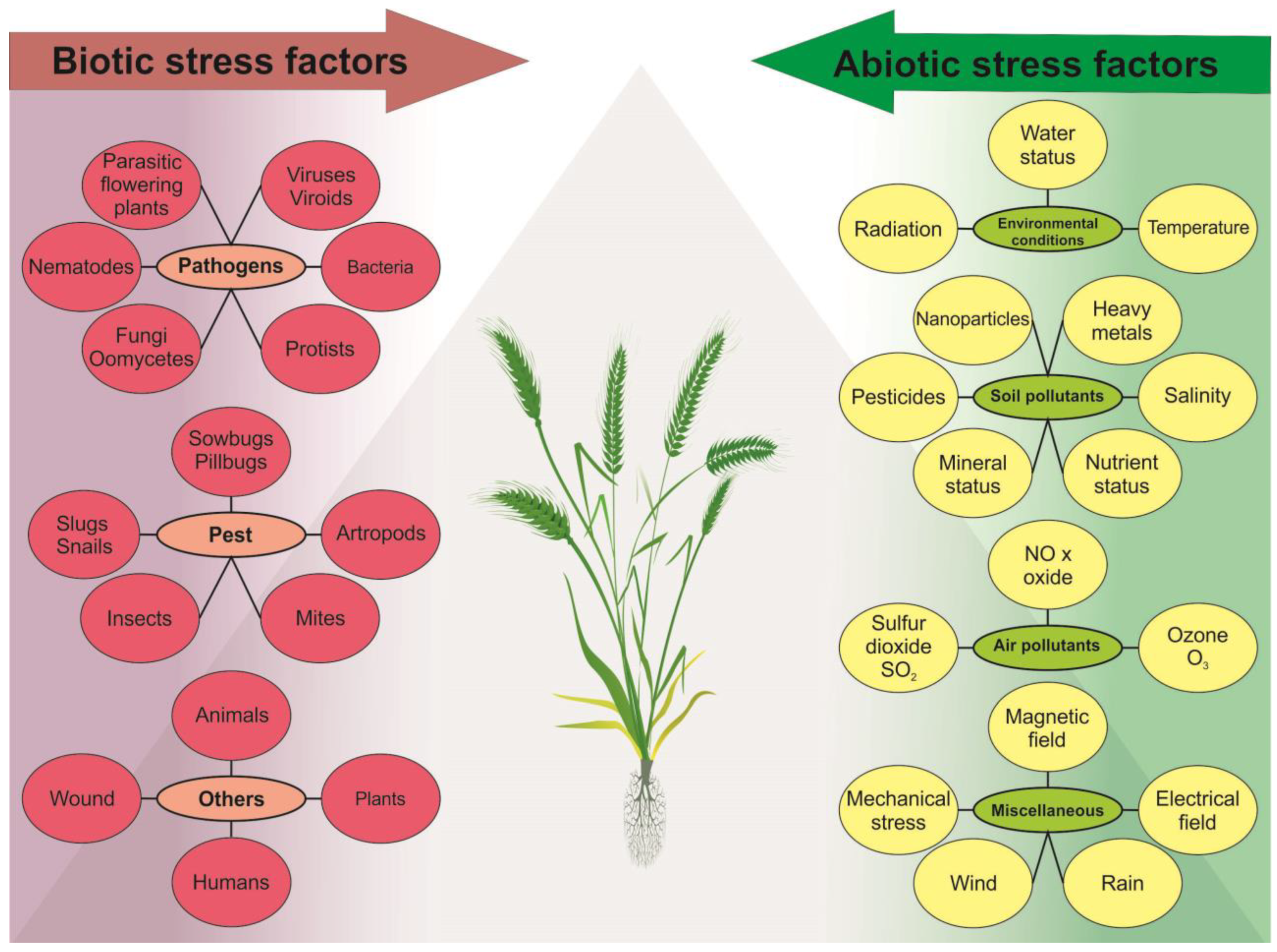
Any extreme event or climate variation can disrupt the optimal conditions for plants, leading to reduced plant growth, leaf and root development, and reduced crop yield [4,5]. Plants are affected by a variety of stress factors, which can be grouped into two main categories: biotic and abiotic.
Biotic stress factors include pathogens, pests, and inter-species rivalry, which cause harm to the plant through infections, herbivory, and resource competition [10,11]. The growth and health of plants can be greatly impacted by various types of pathogens, such as bacteria, viruses, and fungi, leading to a wide range of symptoms, including wilting, discoloration, and even plant death. Pests such as insects, mites, and nematodes can also inflict significant damage to plants by feeding on plant tissue, causing harm to leaves, stems, and even roots, as well as transmitting diseases, thereby exacerbating overall harm to the plant [12].
Biotic stress can cause detrimental effects on plant growth and development, including but not limited to decreasing plant biomass, disrupting photosynthesis, and altering plant morphology [13,14]. This generally leads to the production of defense compounds, such as phenolics and alkaloids, which can negatively affect the nutritional quality of crops. The effects of biotic stress on plants depend on the type of stress, the severity of the stress, and the ability of plants to elicit appropriate defense mechanisms [15].
During biotic stress, plants activate various hormonal pathways to cope with the stressor [13,16]. The Jasmonic acid (JA) pathway plays a crucial role in plant defense against insect herbivores, necrotrophic fungi, and some bacterial pathogens. Upon perceiving the presence of these stressors, plants release JA, which triggers a cascade of signaling events, ultimately leading to the expression of defense-related genes and the production of specialized metabolites, such as phytoalexins, and protease inhibitors [15,17]. Salicylic acid (SA) is another major phytohormone that plays a vital role in plant defense against biotic stress. It triggers the expression of genes responsible for the production of pathogenesis-related proteins that directly attack the pathogen or restrict its spread [18]. Additionally, the host defense system can be reinforced by administering exogenous SA treatment [19]. The ethylene pathway is also activated in response to a range of biotic stressors, such as insect herbivory, pathogen infection, and mechanical damage. This results in increased ethylene levels and the production of defense-related proteins, secondary metabolites, and the activation of signaling pathways involved in plant defense [20]. The mentioned regulatory pathways involved in plant defense exhibit notable differences, yet they also intersect to offer protection against pathogens. Furthermore, phytohormones such as abscisic acid (ABA), auxin, brassinosteroids, cytokinins, gibberellins, and peptide hormones are also involved in modulating plant immune responses [15,21,22,23]. Among these, JA is particularly important in triggering the plant defense system and cross-talks with other phytohormonal pathways [24].
Abiotic stressors encompass a wide range of environmental factors, including water stress, temperature fluctuations, imbalanced nutrient and mineral levels, and exposure to various forms of radiation, such as non-ionizing and ionizing radiation (IR) (high and low linear energy transfer, LET), as well as ultraviolet radiation (UV). Water stress (drought and flooding) affects many aspects of plant growth and development induced by genetic and molecular changes, which alter various biochemical and physiological processes [25] and have both individual and community-wide effects on plants [26]. Temperature stress can result from exposure to extreme cold, freezing temperatures, or high heat, which damage plant tissues and disrupt growth [27,28]. All macronutrient deficiencies inhibit plant growth and development [29]. Exposure to non-ionizing and IR and ultraviolet radiation can harm plants through photoinhibition and photo-oxidation processes [30]. All abiotic stress factors usually lead to genotoxic and oxidative stress, which damages plant DNA and other cellular components [28,31]. Some abiotic stressors affecting plants have anthropogenic origin, namely ozone, carbon dioxide (CO2), emissions of sulfur and nitrogen oxides (SOX and NOX), formaldehydes, ammonia and various complex mixtures, soil pollutants such as soil salinity [4,32,33,34], and the use of nanoparticles in agriculture, such as nano-fertilizer and nano-pesticides [35]. In addition, plants have specialized mechanisms to counteract the effects of sulfur dioxide (SO2) from acid rain [36]. At low concentrations, SO2 can act as a signaling molecule in plants and play a role in seed germination, stomatal opening and closing, and plant response to various environmental stimuli [37]. Similarly, low concentrations of nitric oxide (NO) have a signaling role in plants. NO is non-toxic and has an impact on plant growth and development, senescence, ethylene production, resistance to diseases, and stress tolerance [38]. The combination of natural and anthropogenic factors leads to the spread of heavy metals (Cd, Cr, Pb, Al, Hg, etc.) in soil through volcanic eruptions, industrial activities, and modern agricultural practices. These elements play no role in fundamental metabolic processes and do not have major physiological functions in plants. All of these factors can trigger changes spontaneously, intentionally or as a result of metabolic activity. The diversity of biotic and abiotic factors in plants is illustrated in Figure 1.
3. Impact of Abiotic Stress Factors on the Plant Genome: Direct Effects
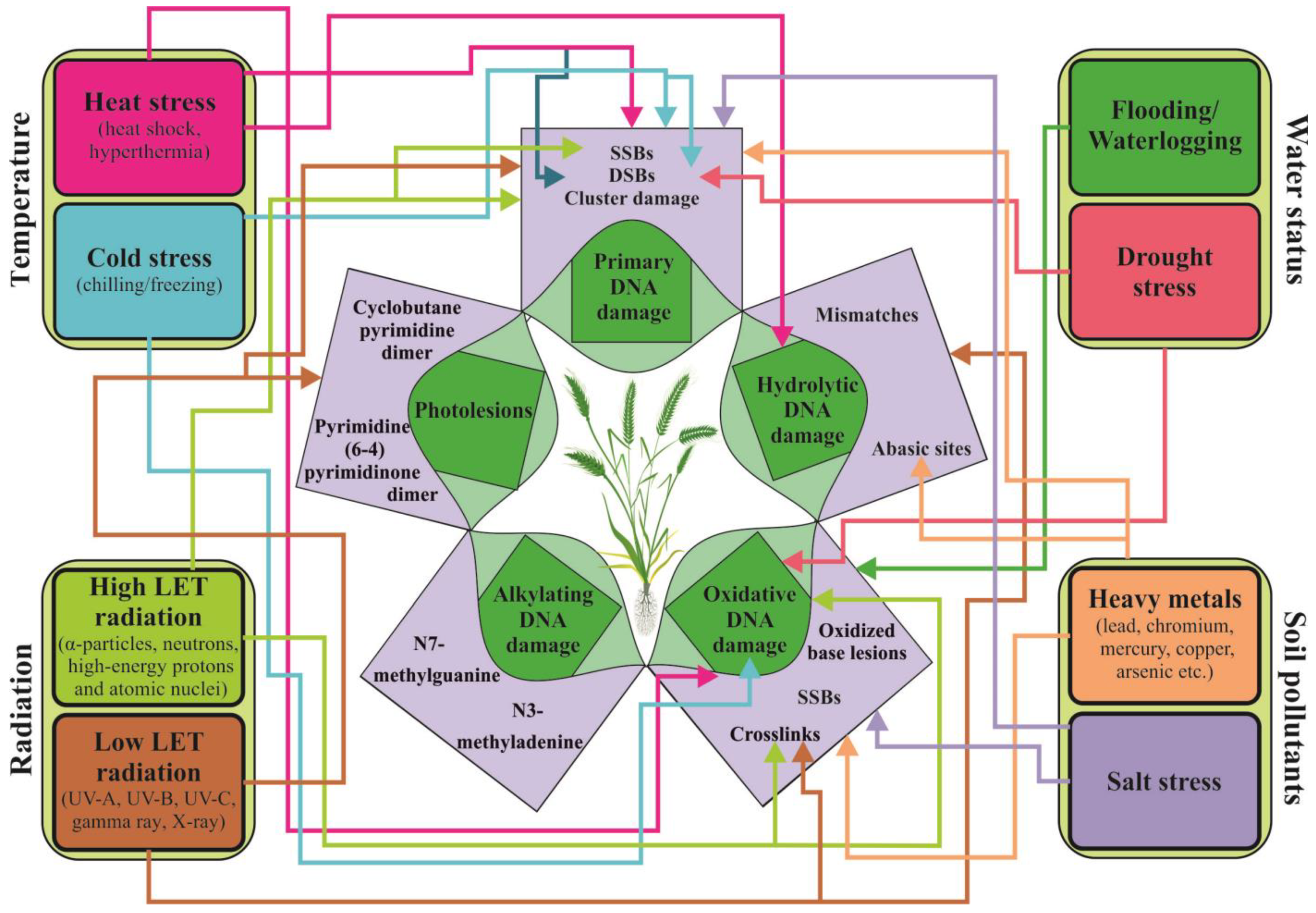
Environmental factors or treatment with agents can affect plant biological macromolecules (DNA, proteins, and lipids) in a direct or indirect manner, leading to oxidative, genotoxic, and cytotoxic effects on plant cells. These effects can cause genomic instability, mutation, carcinogenesis, or even cell death, which can negatively impact plant health and result in reduced crop yield [33,39]. The inability of cells to preserve the integrity of their own genome is linked to primary damage in their biomolecules. The nature of primary DNA damage is similar, regardless of it occurs in microbial, animal, or plant genomes. Whether it originates from external or internal sources, damage can be hydrolytic (cleavage of glycosidic bonds, deamination of cytosine analogs, and depurination) [39]; alkylating [40]; oxidative damage [41]; damage caused by low or high LET radiation (single and double strand breaks, SSBs and DSBs, cluster damage); UV-induced damage (photolesions); or damage caused by base analogs and intercalating agents, crosslinking agents, and protein inhibitors, as illustrated in Figure 2 [42,43,44]. Plant ability to respond to DNA damage caused by stress is dependent on various factors, including their anatomy and morphology, behavioral abilities, physiological resilience, phenotypic flexibility, and the effectiveness of DNA repair mechanisms [44,45,46]. Most environmental factors have a complex mode of action and can directly affect the sugar-phosphate backbone of DNA and lead to the formation of SSBs and DSBs or necessitate metabolic activation.
Hydrolytic DNA damage can occur spontaneously or as a result of stress caused by heating, alkylation of bases, or the action of N-glycosylases [44]. Some of these changes lead to the formation of abasic (apurinic or apyrimidinic, AP) sites. In maize root tip cells, spontaneous hydrolytic DNA damage occurs at a frequency of 3.75 × 105 per genome/per cell during the first 20 h of seed imbibition [47]. According to Britt [39], hydrolytic damage is a common event in the plant genome due to constant exposure to oxygen and UV light. Under stress conditions, the incidence of this type of DNA damage considerably increases.
Exposure to high temperatures can result in an accumulation of hydrolytic damage (such as deaminated cytosine and AP sites) and oxidative damage (such as 8-oxoguanine) in the plant genome [48]. However, plants have the ability to effectively cope with a significant amount of DNA damage through mechanisms that include tolerance or an enhanced repair capacity [49].
Oxidative damage in plant cells is one of the best studied phenomena. Sunlight, specifically its most energetic components (low and high LET; UV radiation), along with air pollutants such as ozone, possess enough energy to excite electrons and ionize water molecules. This results in the formation of water radiolysis products (reactive oxygen species, ROS), hydrated electrons (eaq−), ionized water (H2O+), hydroperoxyl radicals (HO2•), hydroxyl radicals (•OH), hydrogen radicals (H•), reactive nitrogen species (RNS) [49,50], and hydrogen peroxide (H2O2). These events occur within a very short period of time (~10−8 s) [51]. During stress, there is typically an increase in the imbalance between the production and scavenging of ROS [52,53,54]. Recently, two pathways of ROS generation in plant cells have been identified: through plasma membrane-bound nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and peroxidase; or during “electron leak” in chloroplasts, mitochondria, and peroxisomes [55]. Furthermore, different types of ROS have varying levels of reactivity towards DNA. Møller et al. [56] found that among all ROS, only •OH is able to quickly react with DNA molecules. Furthermore, singlet oxygen can also react with DNA, but it primarily does so with guanine. ROS-induced damage to the genome can take many forms, including more than 100 different types of base damage (e.g., single pyrimidine and purine base lesions, inter- and intrastrand crosslinks, purine 5′, 8-cyclonucleosides, DNA–protein adducts, etc.) [57]. Extreme temperatures (frost, cold and heat stress), salinity, drought, water and nutrient imbalance, and high light intensity can trigger oxidative damage in the plant genome [58]. Accumulation of ROS can disrupt the photosynthetic system, carbohydrate metabolism, and some plant cell structures and mechanisms, regardless of whether oxidative damage is caused by endogenous processes or external agents [49,59].
To combat the detrimental effects of oxidative damage, plants have developed a variety of antioxidant regulatory systems, both enzymatic and non-enzymatic [58]. Furthermore, plants possess mechanisms for tolerance, such as ignoring or neglecting induced DNA damage without repair, as discussed by Roldán-Arjona and Ariza [49]. Oxidative stress is often accompanied by nitrosative stress [60]. Activation of RNS, such as NO and nitric dioxide (NO2), as well as nonradicals, such as nitrous acid (HNO2) and dinitrogen tetroxide (N2O4), when combined with superoxide radical (O2•) can lead to disruptions in lipids, thiols, proteins, and DNA bases [56,61]. These signaling molecules are typically associated with the response to abiotic and biotic stresses, but they also play a crucial role in regulating various processes in plants, such as metabolism, growth and development, solute transport, autophagy, and programmed cell death (PCD) [62]. Despite their importance in plant stress response, their potential role in IR stress has not been fully explored [61]. Primary alkylated DNA lesions are a common occurrence in plant genomes, regardless of their endogenous or exogenous origin. They result from the addition of alkyl groups (such as methyl or ethyl group) to oxygen and nitrogen atoms within DNA bases and phosphodiester bonds [43]. These lesions have genotoxic, cytotoxic, or mutagenic properties because they can obstruct gene transcription and replication process within the plant genome.
Secondary damage to the genome, such as AP sites, DNA strand breaks, and interstrand crosslinks, can also occur [63]. In human genomes, the most common alkylated DNA lesions are N7-methylguanine and N3-methyladenine [43,64]. These types of DNA damage can block replication and transcription, leading to the formation of AP sites [64,65]. AP sites can be produced spontaneously or enzymatically during base excision repair (BER). In Arabidopsis, it has been suggested that different groups of endonucleases are active for the removal of AP sites [65].
The use of alkylation damage in plants is often utilized in breeding and genetic modification programs. There are different types of alkylating agents, which are classified by the number of reactive sites as monofunctional, bifunctional, and polyfunctional [66]. These agents can also be classified by their specific nucleophilic substitution reactions, either as monomolecular nucleophilic substitution (SN1) or bimolecular nucleophilic substitution 2 (SN2) [67]. SN1 agents can affect both nitrogen and oxygen atoms on the bases, while SN2 agents mainly affect ring nitrogen atoms (N1, N3 and N7) on the bases. These mutations have been utilized to improve cereal crops and medicinal plants, as well as to breed Capsicum annuum in order to obtain economically valuable traits. Examples of SN1 alkylating agents include N-methyl-N-nitrosourea (MNU), ethyl methanesulfonate (EMS), and N-methyl-N′-nitro-N-nitrosoguanidine (MNNG), while an example of SN2 agents is methyl methanesulfonate (MMS) [68,69].
The main components of solar radiation, namely UV-B and short-wavelength UV-C, induce direct photolesions, such as cyclobutane pyrimidine dimers (CPDs) or pyrimidine (6-4) pyrimidone photoproducts (6-4 PPs), as well as indirect DNA damage through the accumulation of ROS. UV-A radiation primarily causes SSBs, alkali-labile lesions, and oxidative DNA damage due to the low level of absorption by DNA [70].
Exposure to IR, both high and low LET, primarily causes base damage and direct DNA strand breaks, including SSBs and a small number of DSBs (<5%). These breaks occur randomly throughout the plant genome. Depending on the intensity of exposure, AP sites and cluster damage may also occur [71]. It is known that low doses of low LET radiation do not usually cause breaks in the DNA chain. When such breaks do occur, they are more likely to be SSBs than DSBs [72]. SSBs are less severe for the genome as they can be repaired quickly, usually within a minute, as the cell has a copy of intact DNA and can restore its structure during repair. However, if there are more than 100,000 SSBs, cell-cycle arrest may occur. In contrast, DSBs are primarily caused by the breakage of phosphodiester bonds between the sugar residues of two complementary DNA, which occur 10–20 bp apart in both DNA strands [73]. It is known that one to ten DSBs can lead to cell-cycle arrest and ultimately cell death [74]. DNA strand breaks can also be caused by other complex types of abiotic stressors. For instance, heat stress can lead to SSBs and DBSs [48], while cold stress can cause DNA damage, including DSBs in root stem cells [75].
Clustered DNA damage refers to complex DNA injury (including DSBs and non-DSBs) that occurs when there are at least two or more lesions of the DNA helix resulting from single exposure to IR or treatment with radiomimetic agents [76,77]. This type of damage includes harm to DNA bases, SSBs, and AP, as well as modifications to sugar residues [76]. It is believed that cluster damage mainly occurs as an indirect effect of radiation exposure and is highly dependent on the neutralization of free radicals and the structure of chromatin [78,79]. Research on the impacts of IR on plants offers an opportunity to study changes in regions affected by radiation pollution, such as Chernobyl, Fukushima, and the Marshall Islands, as well as in naturally radioactive areas [80,81], to track the adaptation process. Additionally, using radiation in a controlled environment allows radiation mutation breeding to create new plant varieties with desirable traits [82]. In addition to traditional forms of radiation, such as X-rays and gamma rays, there is growing interest in using high-energy particle radiation to enhance the quality of ornamental plants [83], and economically important crops [83,84,85], as well as for algae biofuel production [86].
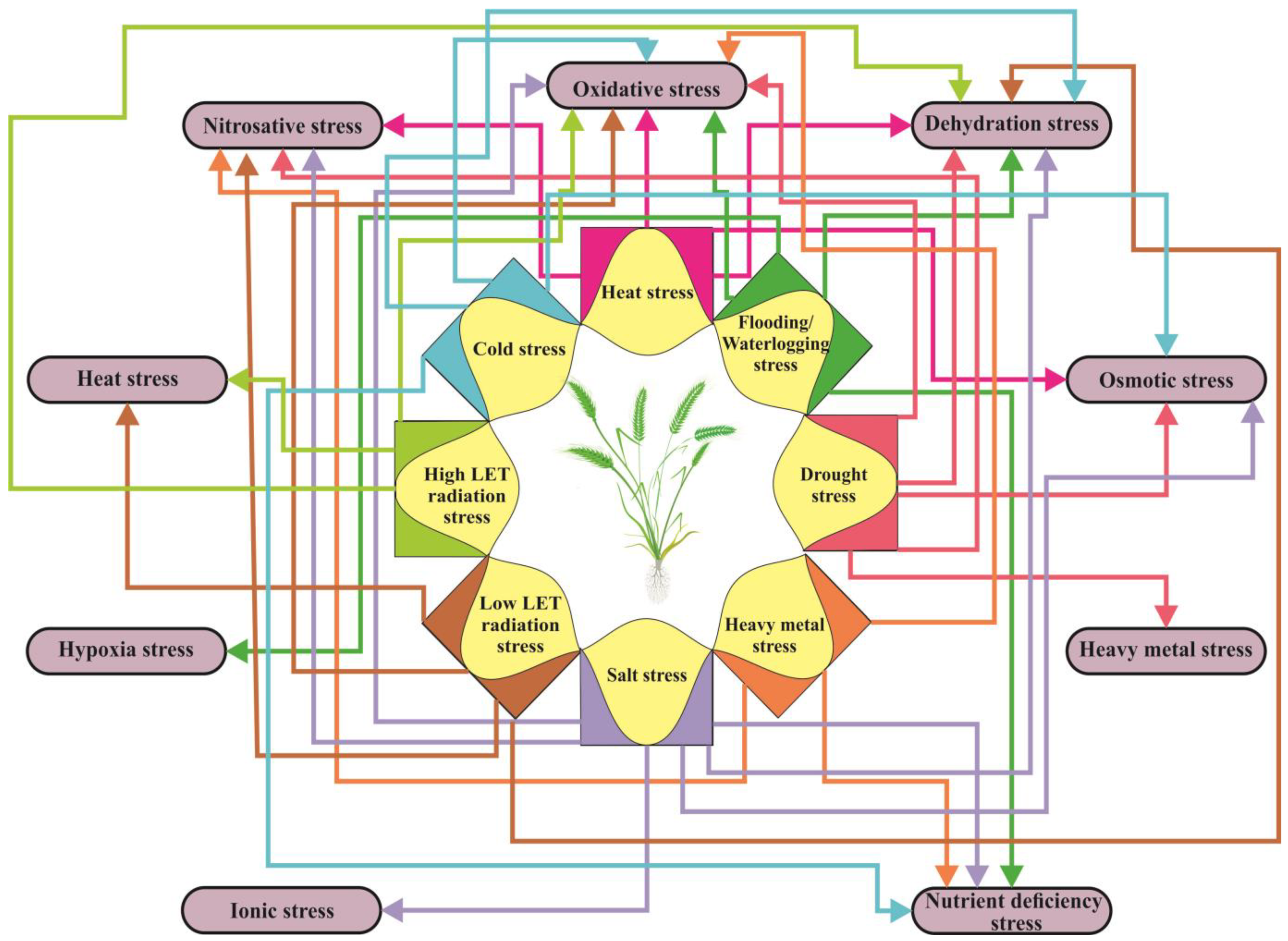
Plants often need to respond to multiple abiotic factors simultaneously, and the consequences of these impacts can lead to secondary and tertiary stress. The most common secondary stress in plants is oxidative stress (Figure 3), which is caused by an overproduction and accumulation of ROS resulting from an imbalance in internal homeostasis. Heat stress causes a variety of changes in plants, including denaturation of proteins and membrane instability, which in turn cause redox stress and an accumulation of ROS [87]. Exposure to radiation can also contribute to the occurrence of heat stress. Salt stress, caused by high soil salinity, leads to both osmotic and ionic stress. The resulting osmotic stress provokes a decrease in plant water content, which is also known as dehydration stress [88]. Dehydration stress primarily occurs during droughts, but it also occurs as a secondary stress during cold, frost, and heat, as well as due to salt. It also occurs as a tertiary stress following radiation stress [89]. Disturbances in K+/Na+ homeostasis, a result of ionic stress, can alter plant metabolism, disrupt membrane structure, and change enzyme activity. These changes can lead to an increase in ROS levels, resulting in the development of oxidative stress as a secondary stress [90], as well as nutritional imbalances [88] (Figure 3). Drought and salt stress elicit similar metabolic responses in plants [91].
4. Dose–Response Models in Plant Ecotoxicology
Investigating the impacts of chemical agents on plants, known as plant ecotoxicology, can be challenging due to the complexity of interpreting the relationship between the dose of the agent and the observed biological effects. Basic models, such as dose–response models, can aid in understanding this relationship by showing how different doses of a chemical agent can lead to different levels of DNA damage in plants. The impact of chemicals on plants can vary depending on the type of agent, concentration, duration of exposure (acute or chronic)], and the mode of action in which the chemical acts on the plant [92,93]. Furthermore, the choice of plant species, testing method (in vivo or in vitro), and the way the chemical is absorbed and metabolized by the plant cells can also affect the results of ecotoxicological research and the effects of the chemical on plants.
Agents can be classified as genotoxic or non-genotoxic based on how they affect cells [93]. Genotoxic agents cause damage to DNA and proteins, and this damage can be classified as primary or secondary depending on whether or not an inflammatory response is present [94]. Primary genotoxins interact directly with cellular components and DNA and lead to the formation of harmful molecules, such as ROS/RNS. They can damage DNA directly or indirectly through the production of free radicals in the mitochondria and membrane-bound NADPH oxidases [94]. Toxicity tests, both in vivo (using live animals) and in vitro (using cell/tissue cultures of human or animal origin), aim to determine the effective dose of the agent that leads to a carcinogenic effect. Non-genotoxic or epigenetic agents primarily affect cell behavior rather than DNA. These agents include tumor promoters, endocrine modifiers, receptor mediators, immunosuppressants or elicitors, and can cause tissue-specific toxicity and inflammatory responses [95]. Most of the agents have little or no impact on plant health as plants possess mechanisms to prevent the spread of tumor cells [96]. On the other hand, some of these agents can be used to achieve specific effects in plant cells through transformation. A key characteristic of these agents is that they have a threshold dose, below which they do not produce a biological effect [93]. Similarly, plants have a threshold radiation dose of 10 mGy·d−1 (417 μGy/h) before adverse effects occur [97]. Genotoxic agents directly damage DNA and chromosomes and have mutagenic effects that do not require metabolic activation. Pro-carcinogenic agents, which do require metabolic activation to cause cancer, also belong to this group. Some inorganic substances, such as metals or metalloids, can also be genotoxic. Their level and distribution in soil and water are crucial for plant growth as some are essential micronutrients [98]. However, excessive amounts could be harmful to both plants and humans. When metabolized by plants, metals/metalloids can enter the food chain and accumulate in the human body, causing harm. High concentrations of these substances in soil also lead to contamination and decreased crop yield [98]. In plant model systems, various agents cause sublethal and lethal effects [92]. These effects often have a specific dose–response relationship, which can be linear or non-linear, with a threshold or non-threshold level. As previously mentioned, non-genotoxic agents typically have a threshold dose, but it has recently been observed that some genotoxic carcinogens also have threshold doses [93].
Haber’s law, one of the earliest models used to describe the relationship between dose and response, states that toxic effects are related to both the concentration and duration of exposure. This model suggests that even small doses of a substance can have an effect, which means that exposure to low levels of a toxic substance over a long period of time can be just as harmful as exposure to a higher level for a shorter period of time. However, in reality, many toxic effects are found to be more influenced by the concentration of the substance rather than the duration of exposure [99]. This means that the amount of a toxic substance in a given environment is more crucial in determining its potential for harm than the length of time when the substance is present. It is important to note that the dose–response relationship is not always linear, and the toxic effects of a substance can also depend on the route of exposure, the organism, and other factors.
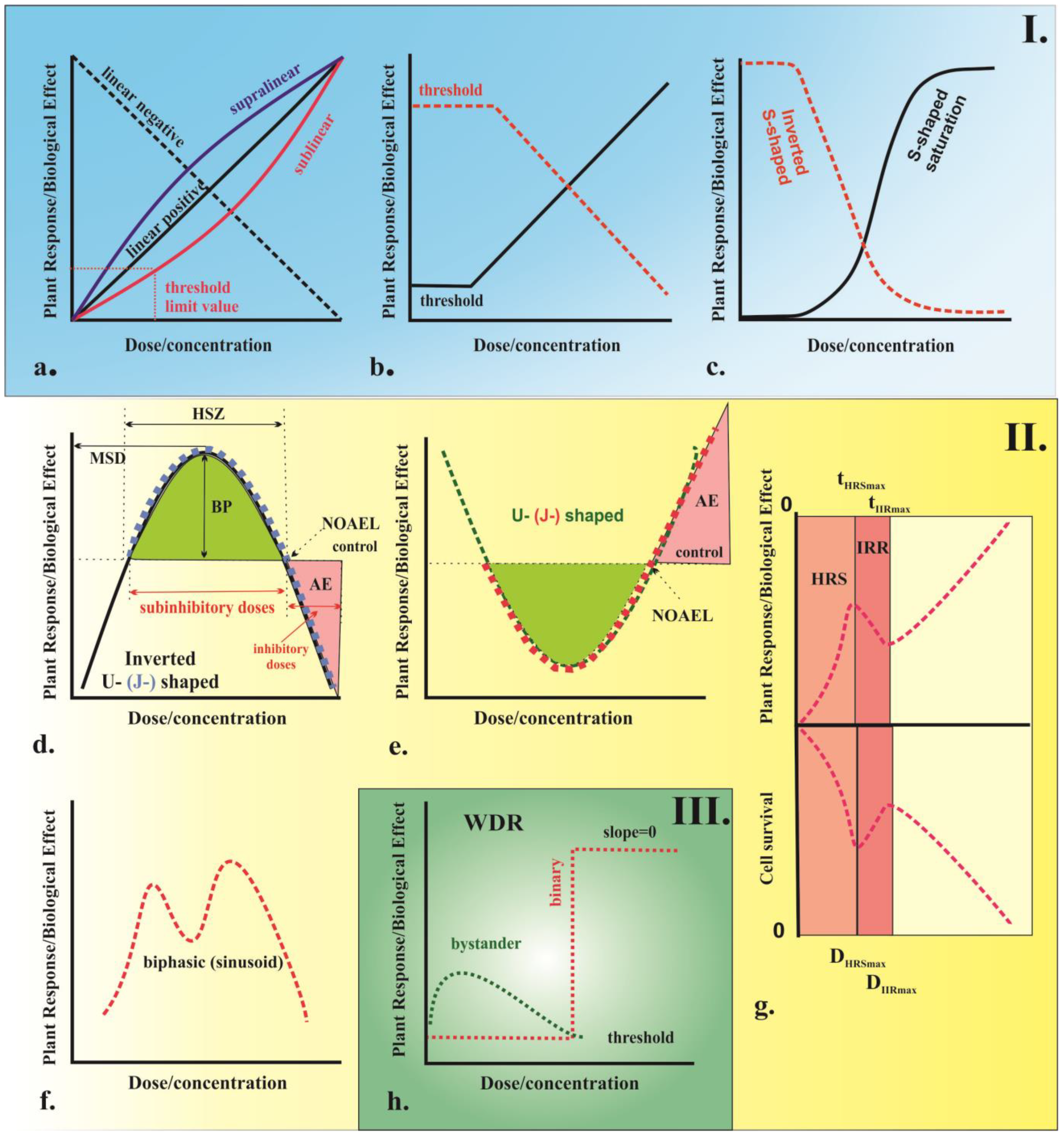
Plants, like all biological systems, are complex organisms and their response to external factors depends on various structural and behavioral factors. There are also notable differences in the way plants and animals respond to agents. According to Karban et al. [100], plants tend to have a higher threshold for agents and a lower sensitivity compared to animal models. The relationship between the amount of an agent (a drug or chemical) applied to an organism and the resulting biological response can be represented by a dose–response curve as illustrated in Figure 4. This type of graph illustrates the relationship between the increased dose or the concentration of the agent and the corresponding increase in biological response. There are two main types of dose–response curves, namely graded and quantal. Graded dose–response curves describe the continuous relationship between increased biological response and increased dose or concentration in the single biological unit. These curves are characterized by four parameters as follows: potency (also known as the half-maximal effective concentration or dose, denoted as EC50 or ED50), which is the dose that produces the maximum effect; slope, which describes how steep the curve is; maximum, which is the highest level of response that can be achieved; and threshold dose, which is the minimum dose required to produce any response [101]. On the other hand, quantal dose–response curves describe the relationship between the proportion of organisms experienced or a not particular effect, known as an “all-or-nothing” phenomenon. Depending on the variation in biological response per unit dose or concentration, different types of dose–response curves can be observed. These can include monotonic curves, where the slope does not change sign; non-monotonic, where the slope changes sign [102]; and curves with no dose–response relationship (WDR), characterized by a zero slope [103]. Examples of different types of curves include linear, non-linear, threshold, sigmoidal, saturation, and U-shaped curves.
In plants, different stressors can produce various dose–response relationships depending on the specific responses and the ability or inability to overcome a particular stressor. Linear response models (Figure 4a,b) with or without threshold dose or concentration (linear no-threshold, LNT; linear threshold, LT) have been used for over 90 years as the standard model for assessing the risk of chemicals, radiation, and environmental agents [107,108]. It should be noted, however, that this model may not always be appropriate for all types of stressors and its limitations should be taken into account. The LNT model, originally developed to explain evolutionary processes [108], has become the main model for assessing the risk of IR effects used by the World Health Organization (WHO) and the Environmental Protection Agency (EPA) as the standard for human health protection. In this model, the biological effect is assumed to be proportional and have a linear or linear-quadratic curve [109]. The linear no-threshold model is based on the principle that radiation is extremely hazardous and there is no safe level of exposure. Even low doses of radiation can result in heritable genetic mutations and tumorigenesis. The target effects of radiation exposure on DNA molecules also follow this pattern [110], which is directly related to the high-dose effects of radiation on human populations. However, this does not necessarily apply to all types of stressors. Furthermore, this model does not always explain the observed short- and long-term effects of low-dose radiation [111], which have been observed in humans during space exploration, after the catastrophic nuclear disasters in Hiroshima and Nagasaki in 1945, and those affected by nuclear disasters at Chernobyl (1986) and Fukushima (2011) [109]. It is worth mentioning that alternative models, such as the hormesis or threshold model, may better explain these effects.
When evaluating the risk for plants, it is important to consider both the potential harm to the plant itself and the potential impact on human health through consumption or other means. For instance, plants grown in areas with high radiation levels or heavy pesticide use may contain dangerous levels of radionuclides or pesticides that could harm humans if consumed [80,112]. Furthermore, doses or concentrations that would affect humans have a greater impact on plants because they have a higher tolerance for radiation and chemical exposure. It should also be noted that various factors, such as the type of agent, the exposure level, the extent of contact with the plant, the accumulation in the plant, the distribution in different plant parts, and the plant ability to neutralize the agent, are crucial in determining the impact. It is also important to consider the context in which the exposure takes place and the specific plant species and variety being studied.
5. Abiotic Stress Factors Elicit Off-Target Effects in Plants In Vivo
The wide variety of plant stressors leads to a number of biological effects. Ideally, stressors only affect specific targets, such as the DNA molecule, resulting in targeted effects that can be seen within one or two generations. However, in reality, organism response is more complex and off-target effects, which are not fully understood, are also observed [113]. Despite limited research in this area, radiation exposure often leads to such effects in animal and human model systems, and similar effects are seen in plants due to various environmental stressors. The known off-target effects of radiation exposure, such as signal-mediated effects, stress-induced genomic instability, transgenerational effects, sensitivity to low doses, biphasic response or hormesis, and others, also occur in the plant genome under environmental stress as outlined by the study of Joiner [114]. More research is needed to fully understand the extent and mechanisms of these off-target effects in plants.
6. Signal-Mediated Effects in Plants
The signal-mediated effects deviate from the traditional target theory and there is ongoing discussion regarding the dose–response relationship. There are two opposing hypotheses. One states that these effects increase with dose, while the other argues that they do not. Some researchers suggest that the effects reach a saturation point at low doses and no further effects are seen at higher doses [104,115,116]. Others consider that a binary response, with a clear threshold dose below which no effects are observed, is also possible [104]. The shape of the dose–response curve describing these effects is still under investigation and can vary depending on the specific stressor and plant species. Typical dose–response curves describing these effects are shown in Figure 4h. The term “signaling-mediated effects” encompasses two physiological phenomena called bystander and abscopal effects, which refer to the ability of untreated cells to pick up stress signals from treated cells, displaying similar changes [117]. These two effects can be differentiated based on the location of the stress response, either in neighboring cells or in distant organs from the site of stress application (“out-of-field”), respectively [118]. The mechanisms that lead to signaling-mediated effects in plants are still being studied, with some theories suggesting that they result from intracellular gap–junction communication [119] or by extracellular low-molecular-weight factors released in the growth medium, such as ROS, cytokines, calcium ions, and short RNAs [120,121,122,123]. The transmitted signals can be harmful or beneficial to the organism, which are referred to as the “kiss of death” and “kiss of life”, respectively [119]. It is known that even low doses of high and low LET radiation can induce radiation bystander effects. Research on signaling-mediated effects in plants began relatively recently with reports on the occurrence of bystander/abscopal effects in plants [120,121]. In these studies, the researchers showed that post-embryonic developmental defects occur in Arabidopsis thaliana after exposure to α-particles and low-energy heavy ions in the shoot apical meristem embryo or intact seeds. Furthermore, some researchers have proposed that the activation of auxin-dependent transcription processes by ROS [124], as well as an increase in homologous recombination events caused by the induction of long-distance DNA damage [125,126], may be responsible for this phenomenon. These findings have been supported by research of Medicago truncatula when studying bystander effects [127]. Further examination of the mechanisms underlying bystander effects has revealed changes in DNA methylation patterns and histone modification [128,129], as well as hormone levels [130]. Evidence of chromosomal damage, including rearrangements and sister chromatid exchanges (SCEs), micronuclei, mutations, cell death, altered gene expression [131], and differentiation and changes in microRNA (miRNA) profiles, have also been found [50].
It has been shown that when Arabidopsis plants grown in Petri dishes are irradiated with UV-C and X-ray, communication between plants leads to phenomenon bystander effects [132]. Similar reactions have also been observed in response to various abiotic stress factors, as well as to biotic stressors, such as pathogen infection and pest infestation [133,134]. When one tobacco plant was infected with tobacco mosaic virus, it resulted in an increased frequency of homologous recombination events that spread to uninfected tissue in both a sensitive and resistant cultivar [133]. Other bystander-like effects have been observed in plant–plant interactions when volatile organic compounds emitted by herbivore-damaged cabbage plants trigger the production of defense traits in nearby undamaged plants in field conditions [135].
7. Transgenerational Memory Effects in Plants
Plants need to adapt better to constantly changing environments in order to pass on their acquired memory to future generations. This can occur through short-term memory factors, such as stress-induced signaling chemicals, proteins, RNAs, and metabolites in the germline, or through changes in certain epigenetic features, referred to as long-term memory factors [136,137]. Changes in the progeny of plants that occur one or two generations after stress are referred to as intergenerational stress response, while changes that occur in several generation or result in permanent changes, called epimutations, are referred to as transgenerational stress response [137]. The chronic effects of low-dose IR on organisms can result in genetic changes that are inherited by the next generation. Studies on the long-term consequences of nuclear power plant accidents, such as those in Chernobyl and Fukushima, have shown that transgenerational heritable effects of IR on plant germline are particularly evident. These effects include an increase in somatic homologous recombination frequency and activation of the ROS scavenging system in plants [138,139]. There is also an increase in the activity of transposable elements [140]. Factors that contribute to transgenerational memory can include not only different types of IR, but also various biotic and abiotic environmental factors [128,141,142,143,144,145]. Transmission of environmental information through epigenetic marks is crucial for transgenerational memory effects in plants. In recent years, there has been growing interest in studying these phenomena, which can have both negative and positive impacts, such as increased genome instability, higher stress tolerance, and cross-tolerance [146,147].
The epigenetic landscape of a plant that includes DNA methylation [148,149,150,151], post-translational histone modifications [152], and small RNAs [153] can be dynamically altered in response to environmental cues [154]. During cell division, DNA methylation patterns are copied to newly synthesized DNA strands and are heritably transmitted from the parent plants to their offspring. Differences in gene expression or phenotype within and between populations can be attributed to stable “epialleles” where the variation in gene expression is caused by epigenetic differences [151]. A well-studied example of such a phenomenon is “paramutation”, wherein one gene allele can transfer epigenetic information to another allele, resulting in a persistent and inheritable modification in gene expression [148,155]. Epigenetic marks can be removed or maintained during reproduction depending on developmental stage and tissue context of the gametes. For instance, DNA methylation can be erased in male gametes but maintained in female gametes. This differential erasure is called genomic imprinting and can result in distinct epigenetic inheritance patterns across paternal and maternal lineages [156]. Following transmission to the next generation, epigenetic marks are interpreted and translated into changes in gene expression and chromatin structure. One example is DNA methylation, which recruits proteins to modify histones, leading to changes in chromatin and gene expression. These modifications are critical for various developmental processes, stress responses, and adaptation to changing environments. They can be transmitted across generations through mitotic and meiotic divisions, contributing to transgenerational epigenetic inheritance. Histone modifications are now recognized as important players in shaping the phenotype and evolution of plant populations [151,152].
Epigenetic marks have varying capacities for long-term inheritance, with DNA methylation being more stably transmitted than other regulators [156]. While histone modifications are more likely to be reset during meiosis, they can work together with other regulators to ensure transgenerational effects. Transgenerational memory can be induced by coordinated epigenetic regulation driven by various factors, such as histone demethylases, heat-shock transcription factors, and trans-acting siRNA biogenesis, to enhance growth and attenuate plant immunity under increased temperature stress [151].
8. Stress-Induced Genomic Instability
Stress-induced instability describes de novo changes at the genomic and chromosomal levels in the progeny of stressed cells, leading to increased carcinogenic susceptibility in these cells. This instability can manifest as an abnormal cell state, such as an accumulation of inherited genomic changes, such as gene mutations, high levels of microsatellite or expanded simple tandem repeat (ESTR) mutation levels, and delayed cell death, or epigenetic modifications, such as changes in DNA methylation, chromatin remodeling, and gene expression [121,157,158]. In addition to genomic instability, stress can also result in chromosomal instability, characterized by high levels of chromosomal rearrangements, formation of micronuclei, increased somatic hyperrecombination, and changes in ploidy levels. Several endpoints have been used to study radiation-induced genome instability in plants, such as analyzing chromosomal aberrations in meiotic pollen mother cells in rice [159], micronuclei formation and cell proliferation in tobacco cells [160], and rearrangements between parental chromosomes in certain root meristem cells in interspecific tobacco hybrids [161]. Significant insights into plant genome instability have been gained through the examination of reconstructed karyotypes of plants produced through IR in breeding practices and analyzed by comet assay [162,163]. It has been proposed that exposure to radiation may cause genome instability, potentially due to an increased accumulation of inherited genomic rearrangements. The presence of specific mutations, such as deletions and translocations, leads to different levels of genomic instability in the irradiated progenitor offspring, which translates into different levels of sensitivity in the lines to IR [162,164]. Initially, a basic understanding of genomic instability in plants came from IR studies; however, further research has revealed that this type of instability can also occur as a result of other methods used in plant breeding, for instance, physical, chemical, or radiation mutagenesis. Recent studies have shown that stress-inducing factors, such as low concentrations of EMS [165], herbicides [166], and salt and heat stress [142], can also lead to genomic instability.
9. Bet-Hedging Strategy and Stress-Induced Transgenerational Tolerance of Plants
Plants have evolved various survival strategies to cope with environmental stress, including bet-hedging and transgenerational stress tolerance. These strategies increase phenotypic diversity and endurance, allowing plants to adapt and survive in changing environments [147]. Bet-hedging can occur through an intergenerational nonheritable effect, where the phenotype of the progeny is influenced by the environmental conditions experienced by the parent [137]. Otherwise, bet-hedging can occur through a transgenerational heritable effect, where the phenotype of the offspring is influenced by the natural surroundings experienced by previous generations [167]. For instance, the Cape Verde islands accession (Cvi-0) of Arabidopsis shows increased phenotypic diversity and endurance in response to high temperatures across two successive generations. The resulting offspring are better adapted to survive in harsh environments. S1 and S2 generations exhibit desirable traits, such as increased plant height, length, and more rosette leaves [168], but these characteristics may diminish in subsequent generations [137]. Similarly, transgenerational stress tolerance in plants refers to their ability to endure various degrees of environmental stress [169], primarily determined by the parental exposure to stress, known as priming stress [136]. Different environmental stressors can induce a phenomenon called primed state and tolerance [169,170,171,172]. These stressors can be applied during various stages of plant development, from seed germination to maturity. Several plant systems have been identified as causes for transgenerational plant tolerance, including hormonal signaling pathways, antioxidant defense systems, and epigenetic modifications [173]. The key aspect of this tolerance is an enhancement in plant growth, yield, performance, and defense [169], which is epigenetically controlled and can be sustained over multiple generations free of stress [173]. This enables plants to not only endure challenging environments but also to maintain an advantage over other individuals, while also increasing their overall fitness and chances of long-term survival [174].
10. Cross-Tolerance
Many studies focus on explaining the effects on plants after being exposed to a single stressor under controlled environmental conditions. However, these effects in natural and field environments are a result of a combination of various biotic and abiotic stressors [175]. Cross-tolerance (cross-resistance or cross-protection) is a phenomenon in which preliminary plant exposure to a mild primary stressor induces tolerance to subsequent exposure to another, more severe stressor. There are three types of cross-tolerances in plants as follows: 1. Transcriptional overlap between different stress responses [2,175]; 2. Induced cross-tolerance, in which the application of priming stress or suppression stress activates plant stress memory and makes it tolerant to subsequent exposure to a different stress [175,176]; and 3. Inherent cross-tolerance, which is characterized by genetic overlap between different stressors. The combined effect of two or more stressors can have a positive or negative impact on plant development and metabolism [177]. Some stress factors may have similar modes of action, using the same signaling molecules and mechanisms [178].
Due to biological plasticity, plants have the ability to readjust and enhance their metabolism and quality characteristics. In this process, the disruption homeostasis of ROS, RNS, and RCS (reactive carbonyl species) in plants is crucial. Severe stress can cause an overproduction of ROS (such as H2O2), leading to oxidative stress. In mild stress or in severe but short-term stress, ROS function as signaling molecules and trigger tolerance mechanisms in plants [2,175,176,178]. Other signaling molecules from the RNS family (e.g., NO) and RCS (e.g., methylglyoxal) play similar roles in building plant tolerance [176,178]. They lead to synergistic co-activation of glyoxalase and antioxidant systems in plants. Tolerance mechanisms also include the accumulation of plant hormones, osmolytes, and heat-shock proteins (HSPs), which contribute to the protection of cellular components, membranes, and proteins by restoring osmotic balance in cells or serving as chemical messengers in signal transduction pathways [7,179]. The cellular signal transduction network involves important players, such as HSPs, heat-shock transcription factors (HSFs), and mitogen-activated protein kinases (MAPKs), which yield specific signaling and connections between different types of stressors [176,178]. To help plants tolerate stress, the expression of stress-responsive genes is controlled by adjusting the levels of specific miRNAs, a process known as post-transcriptional gene regulation [178].
Phytohormones, including salicylic acid (SA), ethylene (ET), jasmonate (JA), abscisic acid (ABA), auxin (AUX), brassinosteroid (BR), gibberellic acid (GA), cytokinin (CK), and strigolactones (SLs), work together to activate defense gene expression and orchestrate effective plant defense responses against abiotic and biotic stress [180,181]. Abscisic acid (ABA) is considered the master hormonal switch that determines whether to prioritize abiotic or biotic stress responses based on the specific nature of the stressors [24]. ABA primarily regulates plant responses to drought, low temperature, and salinity, but it also mediates defense against pathogens [179,182,183]. Jasmonic acid (JA) enhances resistance against hemibiotrophic pathogens, improves tolerance to abiotic stress, and plays a crucial role in plant response to a range of stressors, including drought stress, ozone stress, UV stress, salinity stress, and cold and temperature stress [184]. JA interacts with other hormone signaling pathways, such as auxin, ethylene (ET), ABA, salicylic acid (SA), brassinosteroids (BRs), and gibberellin (GA), suggesting that JA may also function as a central signal in the network of phytohormones. Salicylic acid is usually involved in the regulation of pathogen-associated protein expression, but it also plays an important role in the response to abiotic stresses, including drought, low temperature, and salinity stress [185,186]. When plants are exposed to multiple stressors, such as drought and heat, they often exhibit increased levels of ethylene, which promotes the expression of stress-responsive genes and enhances plant ability to withstand multiple stressors [187]. Thus, cross-talk between phytohormones is a vital component of plant defense responses under abiotic and biotic stress.
11. Low-Dose Hyper-Radiosensitivity and Radioresistance
The concept of low-dose hyper-radiosensitivity is well-established in mammalian systems, where cells display resistance to high single doses of radiation but show sensitivity to small single doses (Figure 4g). This is typified by a limited number of exposures, such as high and low LET IR and chemotherapy drugs [114]. However, experiments on low-dose hyper-radiosensitivity in plant models have not been adequately described [114]. Eriksson [188] is one of the few researchers to do so, reporting this phenomenon in irradiated maize plants after exposure to a dose of 50 cGy where the frequency of mutation induction and lethality in pollen grains was higher than the spontaneous mutation rate. The differences in radiation response at low and high doses of radiation are thought to be due to the different sensitivity of the cell-cycle phases [189]. Further research is needed to fully understand the mechanisms underlying low-dose hyper-radiosensitivity in plant models and to examine the implications of this phenomenon for plant breeding and crop production.
12. Biphasic Dose–Response Effects in Plants: Hormesis, Stress-Induced Priming, and Adaptive Response
Although the existence of biphasic dose–response effects in plants has been known since Darwin’s time, it has long been incorrectly associated with homeopathy, causing a stagnation in its study. Another reason for the lack of sufficient knowledge is that this phenomenon has been studied by different disciplines. For many years, it was rejected by governmental regulatory agencies because it contradicts the established dose–response approach to risk assessment [190]. Recently, Calabrese and Agathokleous [190] reported that there are over 30 different terms used to describe the biphasic dose–response model, including U-shaped, adaptive responses, hormesis, priming, preconditioning, and others, which all describe different aspects of the same phenomenon [191].
In plant studies, various terms have been employed based on the type of stressors. For instance, Ancel and Lallemand [192] used the term “preconditioning” to refer to this phenomenon in plants following X-ray irradiation. As noted in the review by Calabrese and Baldwin [193], the concept of “chemical hormesis” can be traced back to the studies from the late 19th century, which demonstrate the stimulatory effects of sodium hypochlorite on seed germination and the influence of different metals on root growth. In the mid-1970s, the term “adaptive response” was utilized to describe the same phenomenon that occurs after chemical mutagens, and later in the 1980s, it was also used for IR [194]. When defense mechanisms were activated as a result of pathogens, arthropod attacks, or adverse environmental conditions, the term “defense priming” was introduced [195,196]. In recent years, this phenomenon has gained increased attention, leading to its deeper understanding.
Hormesis is considered a quantitative estimate of biological plasticity [197]. The basis of hormetic response is prior exposure to a low dose or concentration of a stress trigger (“priming” stress), which can reduce the toxic effects of subsequent exposure of a higher dose or concentration (“challenge” stress) of the same or a different stress trigger [194,198,199]. The hormetic dose–response curve is often depicted as a U-shaped or J-shaped curve, with the main features being the hormetic stimulatory zone (HSZ) with subinhibitory doses; the maximal stimulatory dose (MSD), which is the percentage change from the control dose (usually <200% of control response) [200]; and the selection of the no observable adverse effects level (NOAEL), or the zero equivalent point (ZEP) or thresholds, followed by inhibitory doses where adverse effects are observed [201,202].
The effects of hormesis on organisms can be either harmful, known as distress, or beneficial, known as eustress [202,203]. In plants, there are two main types of hormetic models, namely inverted U-shaped and U-shaped (Figure 4d,e). The inverted U-shaped curve describes a response in which low dose increases and high dose decreases plant growth and photosynthesis parameters, genotoxicity, and mutagenesis [204]. The U-shaped curve, on the other hand, shows a reduction in adverse effects at low doses and an enhancement of adverse effects at high doses, as can be observed in defense mechanisms such as activities of the major scavenging enzymes, such as ascorbate peroxidase (APX), guaiacol peroxidase (GPX), superoxide radicals, endo-proteinase isoenzymes, carbonyl and malondialdehyde groups, etc. [204]. A third type of hormesis dose–response curve with two dents has also been proposed, but it is specific to plants and there is limited evidence of its existence. This model has been observed in different plants under heavy metal stress [205,206]. The scientific literature is abundant with research on the hormesis behavior of plants, which is triggered by various stress factors. These effects are observed at different levels of biological organization, including cells, organs, organisms, and communities [107]. Hormetic responses have been observed in plants following exposure to a wide range of agents that affect plant growth and development, such as macro- and micronutrients [207,208], biostimulants [209], herbicides and fungicides [210,211,212], heavy metals and metal ions, nanoparticles [204,213,214,215,216], temperature [200], phytohormones [217], heat stress [218], light [219], and pathogens [195].
A growing body of research has shown that both IR and non-IR exposure can have hormetic effects on plants. In general, hormesis is associated with pretreatment of plants with relatively weak exposure (called conditioning clastogenic dose), which increases their resistance to radiation, followed by exposure to higher doses (or challenge dose) of the agents some hours later [220]. Hormesis can be induced by both low and high doses of radiation [221,222,223]. Typically, radiation hormesis in plants has a positive effect, resulting in increased germination, growth rate, height, weight, pigment content, flowering, fertility, accelerated development, and increased radiation resistance. The degree of hormesis depends on the genetic characteristics of the seeds or plant, moisture of the seeds, type of low-dose radiation, and duration of irradiation [224,225]. The adaptive response triggered by hormesis includes both short-term mechanisms, such as the use of existing proteins, and long-term mechanisms, such as the expression of genes encoding specific enzyme systems. Activation of HSPs, proteasomes, and kinase cascades can also occur [225,226]. During hormesis, several mechanisms are activated in plants, including the detoxification of ROS through increased levels of ABA, followed by increased levels of H2O2, activation of DNA repair mechanisms, removal of damaged cells through apoptosis, alteration of nitrogen metabolism, and stimulation of immune response [208,222,226,227,228]. The hormetic part of the adaptive response is associated with permanent genetic or epigenetic changes [229]. Recent studies suggest that epigenetic mechanisms play a role in plant adaptation and generation of transgenerational memories to stress [230]. It is worth mentioning that the complexity of hormesis requires additional research to fully comprehend its underlying mechanisms.
13. Conclusions and Perspectives
In summary, research into the responses of plants to stress is rapidly growing and many questions still need to be explored. The study of how plants handle stressors is crucial, not only for agriculture and the environment, but also for the survival of all living organisms. We summarized current knowledge on the direct effects of high doses or concentrations of stressors on plants (target effects), as well as the effects seen at low doses or concentrations (non-target effects), with a specific focus on the low-level effects that deviate from traditional linear models and do not have a clear threshold in the dose–response relationship. Prolonged exposure to stressors enables plants to adapt and become more resilient, resulting in increased resistance. The response of plants to stress factors is influenced by lifestyle and, in general, plants are more tolerant and able to withstand a wider range of stressors than animals or humans. This is due to the various developmental, physiological, biochemical, genetic, and epigenetic strategies they employ to overcome stress conditions.
The information provided will assist plant scientists in identifying and investigating these phenomena to gain a deeper understanding of the mechanisms behind plant stress response. This response is a fundamental aspect of the natural world and understanding it can yield predictions about the effects of excessive chemical use in agriculture and the potential impacts of untested substances. By considering these factors, scientists and policymakers can develop effective strategies for mitigating the effects of climate change on plants and preserving their habitats and ecosystems.
Author Contributions
M.G. and V.V. planned, designed, wrote, and edited the review. M.G. drew the images. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education and Science (MES) of Bulgaria, grant number D01-271/09.12.2022.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This work has been carried out in the framework of the National Science Program “Environmental Protection and Reduction of Risks of Adverse Events and Natural Disasters”, approved by the Resolution of the Council of Ministers № 577/17.08.2018 and supported by the Ministry of Education and Science (MES) of Bulgaria (Agreement № Д01-271/09.12.2022).
Conflicts of Interest
The authors declare that there are no conflict of interest.
References
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Berkowitz, O.; Whelan, J.; Narsai, R. Cross-species transcriptomic analyses reveals common and opposite responses in Arabidopsis, rice and barley following oxidative stress and hormone treatment. BMC Plant Biol. 2022, 22, 62. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Drury, G.E.; Bray, C.M.; West, C.E. Repairing breaks in the plant genome: The importance of keeping it together. New Phytol. 2011, 192, 805–822. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassileva, V.; Simova-Stoilova, L.; Demirevska, K.; Feller, U. Variety-specific response of wheat (Triticum aestivum L.) leaf mitochondria to drought stress. J. Plant Res. 2009, 122, 445–454. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.; Vassileva, V.; Feller, U. Selection and breeding of suitable crop genotypes for drought and heat periods in a changing climate: Which morphological and physiological properties should be considered? Agriculture 2016, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Bano, C.; Amist, N.; Singh, N.B. Morphological and anatomical modifications of plants for environmental stresses. In Molecular Plant Abiotic Stress: Biology and Biotechnology, 1st ed.; Roychoudhury, A., Tripathi, D., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 29–44. [Google Scholar]
- Manova, V.; Gruszka, D. DNA damage and repair in plants—From models to crops. Front. Plant Sci. 2015, 6, 885. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Mishev, K.; Dobrev, P.I.; Lacek, J.; Filepová, R.; Yuperlieva-Mateeva, B.; Kostadinova, A.; Hristeva, T. Hormonomic changes driving the negative impact of broomrape on plant host interactions with arbuscular mycorrhizal fungi. Int. J. Mol. Sci. 2021, 22, 13677. [Google Scholar] [CrossRef]
- Jones, R.A.; Barbetti, M.J. Influence of climate change on plant disease infections and epidemics caused by viruses and bacteria. CABI Rev. 2012, 7, 1–33. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Plant immune system: Crosstalk between responses to biotic and abiotic stresses the missing link in understanding plant defence. Curr. Issues Mol. Biol. 2017, 23, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Loo, E.P.I. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Saddique, M.; Kamran, M.; Shahbaz, M. Differential responses of plants to biotic stress and the role of metabolites. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 69–87. [Google Scholar]
- Suzuki, N. Hormone signaling pathways under stress combinations. Plant Signal. Behav. 2016, 11, e1247139. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Action of jasmonates in plant stress responses and development—Applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef]
- Hao, Q.; Wang, W.; Han, X.; Wu, J.; Lyu, B.; Chen, F.; Caplan, A.; Li, C.; Wu, J.; Wanget, W.; et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol. Plant Pathol. 2018, 19, 1995–2010. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance-Salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Watkins, J.M.; Hechler, P.J.; Muday, G.K. Ethylene-induced flavonol accumulation in guard cells suppresses reactive oxygen species and moderates stomatal aperture. Plant Physiol. 2014, 164, 1707–1717. [Google Scholar] [CrossRef] [Green Version]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Islam, W.; Naveed, H.; Zaynab, M.; Huang, Z.; Chen, H.Y. Plant defense against virus diseases; growth hormones in highlights. Plant Signal. Behav. 2019, 14, 1596719. [Google Scholar] [CrossRef]
- Chen, Y.L.; Fan, K.T.; Hung, S.C.; Chen, Y.R. The role of peptides cleaved from protein precursors in eliciting plant stress reactions. New Phytol. 2020, 225, 2267–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Intern. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassileva, V.; Demirevska, K.; Simova-Stoilova, L.; Petrova, T.; Tsenov, N.; Feller, U. Long-term field drought affects leaf protein pattern and chloroplast ultrastructure of winter wheat in a cultivar-specific manner. J. Agron. Crop Sci. 2012, 198, 104–117. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigorova, B.; Vassileva, V.; Klimchuk, D.; Vaseva, I.; Demirevska, K.; Feller, U. Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (Triticum aestivum L.) leaves. J. Plant Interact. 2012, 7, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, S.; Mohapatra, T. Interaction between macro-and micro-nutrients in plants. Front. Plant Sci. 2021, 12, 665583. [Google Scholar] [CrossRef]
- Singh, S.; Fatima, A.; Tiwari, S.; Prasad, S.M. Plant responses to radiation stress and its adaptive mechanisms. In Plant Life under Changing Environment: Responses and Management; Tripathi, D.K., Singh, V.P., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 105–122. [Google Scholar]
- Debnath, S.; Chandel, R.K.; Devi, K.; Khan, Z. Mechanism and molecular response of induced genotoxicity and oxidative stress in plants. In Induced Genotoxicity and Oxidative Stress in Plants; Khan, Z., Ansari, M.Y.K., Shahwar, D., Eds.; Springer: Singapore, 2021; pp. 213–227. [Google Scholar]
- Lichtenthaler, H.K. The stress concept in plants: An introduction. Ann. N. Y. Acad. Sci. 1998, 851, 187–198. [Google Scholar] [CrossRef]
- Tuteja, N.; Ahmad, P.; Panda, B.B.; Tuteja, R. Genotoxic stress in plants: Shedding light on DNA damage, repair and DNA repair helicases. Mutat. Res. 2009, 681, 134–149. [Google Scholar] [CrossRef]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci. Total Environ. 2020, 698, 134235. [Google Scholar] [CrossRef]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R.; Janeeshma, E.; Puthur, J.T.; Aliniaeifard, S.; Chauhan, D.K.; et al. Metal/metalloid-based nanomaterials for plant abiotic stress tolerance: An overview of the mechanisms. Plants 2022, 11, 316. [Google Scholar] [CrossRef]
- Lang, C.; Popko, J.; Wirtz, M.; Hell, R.; Herschbach, C.; Kreuzwieser, J.; Rennenberg, H.; Mendel, R.R.; Hänsch, R. Sulphite oxidase as key enzyme for protecting plants against sulphur dioxide. Plant Cell Environ. 2007, 30, 447–455. [Google Scholar] [CrossRef]
- Li, Z.G.; Li, X.E.; Chen, H.Y. Sulfur dioxide: An emerging signaling molecule in plants. Front. Plant Sci. 2022, 13, 891626. [Google Scholar] [CrossRef]
- Garcês, H.; Durzan, D.; Pedroso, M.C. Mechanical stress elicits nitric oxide formation and DNA fragmentation in Arabidopsis thaliana. Ann. Bot. (Lond.) 2001, 87, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Britt, A.B. DNA damage and repair in plants. Annu. Rev. Plant Biol. 1996, 47, 75–100. [Google Scholar] [CrossRef]
- Gichner, T.; Ptácek, O.; Stavreva, D.A.; Plewa, M.J. Comparison of DNA damage in plants as measured by single cell gel electrophoresis and somatic leaf mutations induced by monofunctional alkylating agents. Environ. Mol. Mutagen. 1999, 33, 279–286. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Leitão, J.M. Chemical mutagenesis. In Plant Mutation Breeding and Biotechnology; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; CABI: Oxford, UK, 2012; pp. 135–158. [Google Scholar]
- Schröpfer, S.; Knoll, A.; Trapp, O.; Puchta, H. DNA repair and recombination in plants. In The Plant Sciences; Howell, S., Ed.; Springer: New York, NY, USA, 2014; Volume 2, pp. 51–93. [Google Scholar]
- Verma, P.; Tandon, R.; Yadav, G.; Gaur, V. Structural aspects of DNA repair and recombination in crop improvement. Front. Genet. 2020, 11, 574549. [Google Scholar] [CrossRef]
- Huey, R.B.; Carlson, M.; Crozier, L.; Frazier, M.; Hamilton, H.; Harley, C.; Hoang, A.; Kingsolver, J.G. Plants versus animals: Do they deal with stress in different ways? Integr. Comp. Biol. 2002, 42, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Nisa, M.U.; Huang, Y.; Benhamed, M.; Raynaud, C. The plant DNA damage response: Signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 2019, 10, 653. [Google Scholar] [CrossRef] [Green Version]
- Bray, C.M.; West, C.E. DNA repair mechanisms in plants: Crucial sensors and effectors for the maintenance of genome integrity. New Phytol. 2005, 168, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Kantidze, O.L.; Velichko, A.K.; Luzhin, A.V.; Razin, S.V. Heat stress-induced DNA damage. Acta Nat. 2016, 8, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Roldán-Arjona, T.; Ariza, R.R. Repair and tolerance of oxidative DNA damage in plants. Mutat. Res. 2009, 681, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Yamamori, T.; Yasui, H.; Yamazumi, M.; Wada, Y.; Nakamura, Y.; Nakamura, H.; Inanami, O. Ionizing radiation induces mitochondrial reactive oxygen species production accompanied by upregulation of mitochondrial electron transport chain function and mitochondrial content under control of the cell cycle checkpoint. Free Radic. Biol. Med. 2012, 53, 260–270. [Google Scholar] [CrossRef] [Green Version]
- O’Kane, D.; Gill, V.; Boyd, P.; Burdon, R. Chilling, oxidative stress and antioxidant responses in Arabidopsis thaliana callus. Planta 1996, 198, 371–377. [Google Scholar] [CrossRef]
- Skyba, M.; Petijová, L.; Košuth, J.; Koleva, D.P.; Ganeva, T.G.; Kapchina-Toteva, V.M.; Cellárová, E. Oxidative stress and antioxidant response in Hypericum perforatum L. plants subjected to low temperature treatment. J. Plant Physiol. 2012, 169, 955–964. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhu, L.; He, L.; Chen, D.; Zeng, D.; Chen, G.; Hu, J.; Zhang, G.; Ren, D.; Dong, G.; et al. DNA damage and reactive oxygen species cause cell death in the rice local lesions 1 mutant under high light and high temperature. New Phytol. 2019, 222, 349–365. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Lu, T.; Meng, Z.; Zhang, G.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Sub-high temperature and high light intensity induced irreversible inhibition on photosynthesis system of tomato plant (Solanum lycopersicum L.). Front. Plant Sci. 2017, 8, 365. [Google Scholar] [CrossRef] [Green Version]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [Green Version]
- Turkan, I. ROS and RNS: Key signalling molecules in plants. J. Exp. Bot. 2018, 69, 3313–3315. [Google Scholar] [CrossRef] [Green Version]
- Drabløs, F.; Feyzi, E.; Aas, P.A.; Vaagbø, C.B.; Kavli, B.; Bratlie, M.S.; Peña-Diaz, J.; Otterlei, M.; Slupphaug, G.; Krokan, H.E. Alkylation damage in DNA and RNA-repair mechanisms and medical significance. DNA Repair 2004, 3, 1389–1407. [Google Scholar] [CrossRef]
- Mishina, Y.; Duguid, E.M.; He, C. Direct reversal of DNA alkylation damage. Chem. Rev. 2006, 106, 215–332. [Google Scholar] [CrossRef]
- Barbado, C.; Córdoba-Cañero, D.; Ariza, R.R.; Roldán-Arjona, T. Nonenzymatic release of N7-methylguanine channels repair of abasic sites into an AP endonuclease-independent pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E916–E924. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, A.T. Chemical mutagenesis: From plants to human. Curr. Sci. 2005, 89, 312–317. [Google Scholar]
- Peng, Y.; Pei, H. DNA alkylation lesion repair: Outcomes and implications in cancer chemotherapy. J. Zhejiang Univ. Sci. B 2021, 22, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Kakar, K.; Xuan, T.D.; Quan, N.V.; Wafa, I.K.; Tran, H.-D.; Khanh, T.D.; Dat, T.D. Efficacy of N-methyl-N-nitrosourea (MNU) mutation on enhancing the yield and quality of rice. Agriculture 2019, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Siddique, M.I.; Back, S.; Lee, J.H.; Jo, J.; Jang, S.; Han, K.; Venkatesh, J.; Kwon, J.K.; Jo, Y.D.; Kang, B.C. Development and characterization of an ethyl methane sulfonate (EMS) induced mutant population in Capsicum annuum L. Plants 2020, 9, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Jha, M.; Tuteja, N. DNA damage and repair in plants under ultraviolet and ionizing radiations. Sci. World J. 2015, 2015, 250158. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.J.; Nickson, C.M.; Thompson, J.M.; Kacperek, A.; Hill, M.A.; Parsons, J.L. Complex DNA damage induced by high linear energy transfer alpha-particles and protons triggers a specific cellular DNA damage response. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Hada, M.; Georgakilas, A.G. Formation of clustered DNA damage after high-LET irradiation: A review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef]
- Rydberg, B.; Cooper, B.; Cooper, P.K.; Holley, W.R.; Chatterjee, A. Dose-dependent misrejoining of radiation-induced DNA double-strand breaks in human fibroblasts: Experimental and theoretical study for high- and low-LET radiation. Radiat. Res. 2005, 163, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Clarkin, K.C.; Wahl, G.M. Sensitivity and selectivity of the DNA damage sensor responsible for activating p53-dependent G1 arrest. Proc. Natl. Acad. Sci. USA 1996, 93, 4827–4832. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Savina, M.; Du, J.; Devendran, A.; Kannivadi Ramakanth, K.; Tian, X.; Sim, W.S.; Mironova, V.V.; Xu, J. A sacrifice-for-survival mechanism protects root stem cell niche from chilling stress. Cell 2017, 170, 102–113.e14. [Google Scholar] [CrossRef] [Green Version]
- Asaithamby, A.; Chen, D.J. Mechanism of cluster DNA damage repair in response to high-atomic number and energy particles radiation. Mutat. Res. 2011, 711, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Nickoloff, J.A.; Sharma, N.; Taylor, L. Clustered DNA double-strand breaks: Biological effects and relevance to cancer radiotherapy. Genes 2020, 11, 99. [Google Scholar] [CrossRef] [Green Version]
- Hada, M.; Sutherland, B.M. Spectrum of complex DNA damages depends on the incident radiation. Radiat. Res. 2006, 165, 223–230. [Google Scholar] [CrossRef]
- Magnander, K.; Hultborn, R.; Claesson, K.; Elmroth, K. Clustered DNA damage in irradiated human diploid fibroblasts: Influence of chromatin organization. Radiat. Res. 2010, 173, 272–282. [Google Scholar] [CrossRef]
- Georgieva, M.; Rashydov, N.M.; Hajduch, M. DNA damage, repair monitoring and epigenetic DNA methylation changes in seedlings of Chernobyl soybeans. DNA Repair 2017, 50, 14–21. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Møller, A.P. Plants in the light of ionizing radiation: What have we learned from Chernobyl, Fukushima, and other “hot” places? Front. Plant Sci. 2020, 11, 552. [Google Scholar] [CrossRef]
- Ma, L.; Kong, F.; Sun, K.; Wang, T.; Guo, T. From classical radiation to modern radiation: Past, present, and future of radiation mutation breeding. Front. Public Health 2021, 9, 768071. [Google Scholar] [CrossRef]
- Abe, T.; Ryuto, H.; Fukunishi, N. Ion beam radiation mutagenesis. In Plant Mutation Breeding and Biotechnology; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; Joint FAO/IAEA: Vienna, Austria, 2012; pp. 99–106. [Google Scholar]
- Kato, M.; Masamura, N.; Shono, J.; Okamoto, D.; Abe, T.; Imai, S. Production and characterization of tearless and non-pungent onion. Sci. Rep. 2016, 6, 23779. [Google Scholar] [CrossRef] [Green Version]
- Yamatani, H.; Kohzuma, K.; Nakano, M.; Takami, T.; Kato, Y.; Hayashi, Y.; Monden, Y.; Okumoto, Y.; Abe, T.; Kumamaru, T.; et al. Impairment of Lhca4, a subunit of LHCI, causes high accumulation of chlorophyll and the stay-green phenotype in rice. J. Exp. Bot. 2018, 69, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, T.; Ivanov, I.N.; Oshima, K.; Ishii, K.; Kawamoto, H.; Ota, S.; Yamazaki, T.; Hirata, A.; Kazama, Y.; Abe, T.; et al. Comparison of lipid productivity of Parachlorella kessleri heavy-ion beam irradiation mutant PK4 in laboratory and 150-L mass bioreactor, identification and characterization of its genetic variation. Algal Res. 2018, 35, 416–426. [Google Scholar] [CrossRef]
- Masouleh, S.S.S.; Sassine, Y.N. Molecular and biochemical responses of horticultural plants and crops to heat stress. Ornam. Hortic. 2020, 26, 148–158. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamińska, I.; Lukasiewicz, A.; Klimek-Chodacka, M.; Długosz-Grochowska, O.; Rutkowska, J.; Szymonik, K.; Baranski, R. Antioxidative and osmoprotecting mechanisms in carrot plants tolerant to soil salinity. Sci. Rep. 2022, 12, 7266. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Lyubenova, M.; Boteva, S. Biotests in ecotoxicology: Current practice and problems. In Toxicology—New Aspects to This Scientific Conundrum; Soloneski, S., Larramendy, M.L., Eds.; InTechOpen Limited: London, UK, 2016; pp. 147–177. [Google Scholar]
- Bevan, R.J.; Harrison, P.T.C. Threshold and non-threshold chemical carcinogens: A survey of the present regulatory landscape. Regul. Toxicol. Pharmacol. 2017, 88, 291–302. [Google Scholar] [CrossRef]
- Shukla, R.K.; Badiye, A.; Vajpayee, K.; Kapoor, N. Genotoxic potential of nanoparticles: Structural and functional modifications in DNA. Front. Genet. 2021, 12, 728250. [Google Scholar] [CrossRef]
- Hernández, L.G.; van Steeg, H.; Luijten, M.; van Benthem, J. Mechanisms of non-genotoxic carcinogens and importance of a weight of evidence approach. Mutat. Res. 2009, 682, 94–109. [Google Scholar] [CrossRef]
- Doonan, J.H.; Sablowski, R. Walls around tumours—Why plants do not develop cancer. Nat. Rev. Cancer 2010, 10, 794–802. [Google Scholar] [CrossRef]
- Caplin, N.; Willey, N. Ionizing radiation, higher plants, and radioprotection: From acute high doses to chronic low doses. Front. Plant Sci. 2018, 9, 847. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef]
- Raies, A.B.; Bajic, V.B. In silico toxicology: Computational methods for the prediction of chemical toxicity. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2016, 6, 147–172. [Google Scholar] [CrossRef] [Green Version]
- Karban, R.; Orrock, J.L.; Preisser, E.L.; Sih, A. A comparison of plants and animals in their responses to risk of consumption. Curr. Opin. Plant Biol. 2016, 32, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mathis, D.A.; Emmett-Oglesby, M.W. Quantal vs. graded generalization in drug discrimination: Measuring a graded response. J. Neurosci. Methods 1990, 31, 23–33. [Google Scholar] [CrossRef]
- Thomas, C.; Martin, J.; Devic, C.; Bräuer-Krisch, E.; Diserbo, M.; Thariat, J.; Foray, N. Impact of dose-rate on the low-dose hyper-radiosensitivity and induced radioresistance (HRS/IRR) response. Int. J. Radiat. Biol. 2013, 89, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Devic, C.; Ferlazzo, M.L.; Berthel, E.; Foray, N. Influence of individual radiosensitivity on the hormesis phenomenon: Toward a mechanistic explanation based on the nucleoshuttling of ATM protein. Dose-Response 2020, 18, 1559325820913784. [Google Scholar] [CrossRef]
- Prise, K.M.; O’Sullivan, J.M. Radiation-induced bystander signalling in cancer therapy. Nat. Rev. Cancer 2009, 9, 351–360. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Chevillotte, G.; Bernard, A.; Varret, C.; Ballet, P.; Bodin, L.; Roudot, A.C. Probabilistic assessment method of the non-monotonic dose-responses-Part I: Methodological approach. Food Chem. Toxicol. 2017, 106, 376–385. [Google Scholar] [CrossRef]
- Agathokleous, E.; Araminiene, V.; Belz, R.G.; Calatayud, V.; De Marco, A.; Domingos, M.; Feng, Z.; Hoshika, Y.; Kitao, M.; Koike, T.; et al. A quantitative assessment of hormetic responses of plants to ozone. Environ. Res. 2019, 176, 108527. [Google Scholar] [CrossRef]
- Calabrese, E.J. The linear No-Threshold (LNT) dose response model: A comprehensive assessment of its historical and scientific foundations. Chem. Biol. Interact. 2019, 301, 6–25. [Google Scholar] [CrossRef]
- Tran, L.; Seeram, E. Current perspectives on the use of the linear non-threshold (LNT) model in radiation protection. Int. J. Radiol. Med. Imag. 2017, 3, 123. [Google Scholar]
- Shuryak, I.; Brenner, D.J.; Blattnig, S.R.; Shukitt-Hale, B.; Rabin, B.M. Modeling space radiation induced cognitive dysfunction using targeted and non-targeted effects. Sci. Rep. 2021, 11, 8845. [Google Scholar] [CrossRef] [PubMed]
- Averbeck, D. Non-targeted effects as a paradigm breaking evidence. Mutat. Res. 2010, 687, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.; Bonchev, G.; Zehirov, G.; Vasileva, V.; Vassileva, V. Neonicotinoid insecticides exert diverse cytotoxic and genotoxic effects on cultivated sunflower. Environ. Sci. Pollut. Res. Int. 2021, 28, 53193–53207. [Google Scholar] [CrossRef]
- Ward, J.F. DNA damage produced by ionizing radiation in mammalian cells: Identities, mechanisms of formation, and reparability. Prog. Nucleic Acid Res. Mol. Biol. 1988, 35, 95–125. [Google Scholar]
- Joiner, M.C.; Marples, B.; Lambin, P.; Short, S.C.; Turesson, I. Low-dose hypersensitivity: Current status and possible mechanisms. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 379–389. [Google Scholar] [CrossRef]
- Soleymanifard, S.; Bahreyni Toossi, M.T.; Sazgarnia, A.; Mohebbi, S. The role of target and bystander cells in dose-response relationship of radiation-induced bystander effects in two cell lines. Iran. J. Basic Med. Sci. 2013, 16, 177–183. [Google Scholar]
- Farias, V.A.; Tovar, I.; Del Moral, R.; O’Valle, F.; Expósito, J.; Oliver, F.J.; Ruiz de Almodóvar, J.M. Enhancing the bystander and abscopal effects to improve radiotherapy outcomes. Front. Oncol. 2020, 9, 1381. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhou, T.; Liu, W.; Zuo, L. Molecular mechanism of bystander effects and related abscopal/cohort effects in cancer therapy. Oncotarget 2018, 9, 18637–18647. [Google Scholar] [CrossRef] [Green Version]
- Daguenet, E.; Louati, S.; Wozny, A.S.; Vial, N.; Gras, M.; Guy, J.B.; Vallard, A.; Rodriguez-Lafrasse, C.; Magné, N. Radiation-induced bystander and abscopal effects: Important lessons from preclinical models. Br. J. Cancer 2020, 123, 339–348. [Google Scholar] [CrossRef]
- Spray, D.C.; Hanstein, R.; Lopez-Quintero, S.V.; Stout, R.F.; Suadicani, S.O.; Thi, M.M. Gap junctions and bystander effects: Good samaritans and executioners. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Wu, L.; Chen, L.; Pei, B.; Wang, Y.; Zhan, F.; Wu, Y.; Yu, Z. Targeted irradiation of shoot apical meristem of Arabidopsis embryos induces long-distance bystander/abscopal effects. Radiat. Res. 2007, 167, 298–305. [Google Scholar] [CrossRef]
- Merrifield, M.; Kovalchuk, O. Epigenetics in radiation biology: A new research frontier. Front. Genet. 2013, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium signaling-mediated plant response to cold stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, M.; Neumann, R. Changes in gene expression as one of the key mechanisms involved in radiation-induced bystander effect. Biomed. Rep. 2018, 9, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Mei, T.; Yuan, H.; Zhang, W.; Chen, L.; Xue, J.; Wu, L.; Wang, Y. Bystander/abscopal effects induced in intact Arabidopsis seeds by low-energy heavy-ion radiation. Radiat. Res. 2008, 170, 372–380. [Google Scholar] [CrossRef]
- Li, F.; Liu, P.; Wang, T.; Bian, P.; Wu, Y.; Wu, L.; Yu, Z. The induction of bystander mutagenic effects in vivo by alpha-particle irradiation in whole Arabidopsis thaliana plants. Radiat. Res. 2010, 174, 228–237. [Google Scholar] [CrossRef]
- Li, F.; Wang, T.; Xu, S.; Yuan, H.; Bian, P.; Wu, Y.; Wu, L.; Yu, Z. Abscopal mutagenic effect of low-energy-ions in Arabidopsis thaliana seeds. Int. J. Radiat. Biol. 2011, 87, 984–992. [Google Scholar] [CrossRef]
- Chen, H.; Li, F.; Yuan, H.; Xiao, X.; Yang, G.; Wu, L. Abscopal signals mediated bio-effects in low-energy ion irradiated Medicago truncatula seeds. J. Radiat. Res. 2010, 51, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Boyko, A.; Kathiria, P.; Zemp, F.J.; Yao, Y.; Pogribny, I.; Kovalchuk, I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants: (virus-induced plant genome instability). Nucleic Acids Res. 2007, 35, 1714–1725. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Wang, T.; Xu, S.; Xu, S.; Wu, L.; Wu, Y.; Bian, P. Radiation-induced epigenetic bystander effects demonstrated in Arabidopsis thaliana. Radiat. Res. 2015, 183, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Schilmiller, A.L.; Howe, G.A. Systemic signaling in the wound response. Curr. Opin. Plant Biol. 2005, 8, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Xu, W.; Deng, C.; Wu, J.; Wang, T. Gene transcriptomic profile in Arabidopsis thaliana mediated by radiation-induced bystander effects. Int. J. Radiat. Res. 2019, 17, 369–377. [Google Scholar]
- Zemp, F.J.; Sidler, C.; Kovalchuk, I. Increase in recombination rate in Arabidopsis thaliana plants sharing gaseous environment with X-ray and UVC-irradiated plants depends on production of radicals. Plant Signal. Behav. 2012, 7, 782–787. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, I.; Kovalchuk, O.; Kalck, V.; Boyko, V.; Filkowski, J.; Heinlein, M.; Hohn, B. Pathogen-induced systemic plant signal triggers DNA rearrangements. Nature 2003, 423, 760–762. [Google Scholar] [CrossRef]
- Grant, M.; Lamb, C. Systemic immunity. Curr. Opin. Plant Biol. 2006, 9, 414–420. [Google Scholar] [CrossRef]
- Giron-Calva, P.S.; Li, T.; Blande, J.D. Volatile-mediated interactions between cabbage plants in the field and the impact of ozone pollution. J. Chem. Ecol. 2017, 43, 339–350. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Chen, J.; Wang, X.; Cai, J.; Zhou, Q.; Dai, T.; Cao, W.; Jiang, D. Parental drought-priming enhances tolerance to post-anthesis drought in offspring of wheat. Front. Plant Sci. 2018, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, I. Transgenerational genome instability in plants. In Genome Stability, From Virus to Human Application, 2nd ed.; Kovalchuk, I., Kovalchuk, O., Eds.; Academic Press: London, UK, 2021; Volume 26, pp. 659–678. [Google Scholar]
- Kovalchuk, O.; Arkhipov, A.; Barylyak, I.; Karachov, I.; Titov, V.; Hohn, B.; Kovalchuk, I. Plants experiencing chronic internal exposure to ionizing radiation exhibit higher frequency of homologous recombination than acutely irradiated plants. Mutat. Res. 2000, 449, 47–56. [Google Scholar] [CrossRef]
- Ćuk, K.; Gogala, M.; Tkalec, M.; Vidaković-Cifrek, Ž. Transgenerational stress memory in Arabidopsis thaliana (L.) Heynh.: Antioxidative enzymes and HSP70. Acta Bot. Croat. 2010, 69, 183–197. [Google Scholar]
- Migicovsky, Z.; Kovalchuk, I. Changes to DNA methylation and homologous recombination frequency in the progeny of stressed plants. Biochem. Cell Biol. 2013, 91, 1–5. [Google Scholar] [CrossRef]
- Rahavi, M.R.; Migicovsky, Z.; Titov, V.; Kovalchuk, I. Transgenerational adaptation to heavy metal salts in Arabidopsis. Front. Plant Sci. 2011, 2, 91. [Google Scholar] [CrossRef] [Green Version]
- Suter, L.; Widmer, A. Environmental heat and salt stress induce transgenerational phenotypic changes in Arabidopsis thaliana. PLoS ONE 2013, 8, e60364. [Google Scholar] [CrossRef]
- Groot, M.P.; Kubisch, A.; Ouborg, N.J.; Pagel, J.; Schmid, K.J.; Vergeer, P.; Lampei, C. Transgenerational effects of mild heat in Arabidopsis thaliana show strong genotype specificity that is explained by climate at origin. New Phytol. 2017, 215, 1221–1234. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Chen, L.; Xia, H.; Wei, H.; Lou, Q.; Li, M.; Li, T.; Luo, L. Transgenerational epimutations induced by multi-generation drought imposition mediate rice plant’s adaptation to drought condition. Sci. Rep. 2017, 7, 39843. [Google Scholar] [CrossRef] [Green Version]
- Colzi, I.; Gonnelli, C.; Vergata, C.; Golia, G.; Coppi, A.; Castellani, M.B.; Giovino, A.; Buti, M.; Sabato, T.; Capuana, M.; et al. Transgenerational effects of chromium stress at the phenotypic and molecular level in Arabidopsis thaliana. J. Hazard Mater. 2023, 442, 130092. [Google Scholar] [CrossRef]
- Herman, J.J.; Sultan, S.E. Adaptive transgenerational plasticity in plants: Case studies, mechanisms, and implications for natural populations. Front. Plant Sci. 2011, 2, 102. [Google Scholar] [CrossRef] [Green Version]
- Bilichak, A.; Kovalchuk, I. Transgenerational response to stress in plants and its application for breeding. J. Exp. Bot. 2016, 67, 2081–2092. [Google Scholar] [CrossRef]
- Sarkies, P. Molecular mechanisms of epigenetic inheritance: Possible evolutionary implications. Semin. Cell Dev. Biol. 2020, 97, 106–115. [Google Scholar] [CrossRef]
- Boyko, A.; Blevins, T.; Yao, Y.; Golubov, A.; Bilichak, A.; Ilnytskyy, Y.; Hollunder, J.; Meins, F., Jr.; Kovalchuk, I. Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of Dicer-like proteins. PLoS ONE 2010, 5, e9514. [Google Scholar] [CrossRef]
- Schmitz, R.J.; Schultz, M.D.; Lewsey, M.G.; O’Malley, R.C.; Urich, M.A.; Libiger, O.; Schork, N.J.; Ecker, J.R. Transgenerational epigenetic instability is a source of novel methylation variants. Science 2011, 334, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mladenov, V.; Fotopoulos, V.; Kaiserli, E.; Karalija, E.; Maury, S.; Baranek, M.; Segal, N.A.; Testillano, P.S.; Vassileva, V.; Pinto, G.; et al. Deciphering the epigenetic alphabet involved in transgenerational stress memory in crops. Int. J. Mol. Sci. 2021, 22, 7118. [Google Scholar] [CrossRef] [PubMed]
- Lang-Mladek, C.; Popova, O.; Kiok, K.; Berlinger, M.; Rakic, B.; Aufsatz, W.; Jonak, C.; Hauser, M.T.; Luschnig, C. Transgenerational inheritance and resetting of stress-induced loss of epigenetic gene silencing in Arabidopsis. Mol. Plant. 2010, 3, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, L.; Han, T.; Ye, W.; Liu, Y.; Sun, Y.; Moose, S.P.; Song, Q.; Chen, Z.J. Small RNAs mediate transgenerational inheritance of genome-wide trans-acting epialleles in maize. Genome Biol. 2022, 23, 53. [Google Scholar] [CrossRef]
- Sun, M.; Yang, Z.; Liu, L.; Duan, L. DNA methylation in plant responses and adaption to abiotic stresses. Int. J. Mol. Sci. 2022, 23, 6910. [Google Scholar] [CrossRef]
- Pilu, R. Paramutation phenomena in plants. Semin. Cell Dev. Biol. 2015, 44, 2–10. [Google Scholar] [CrossRef]
- Lucibelli, F.; Valoroso, M.C.; Aceto, S. Plant DNA methylation: An epigenetic mark in development, environmental interactions, and evolution. Int. J. Mol. Sci. 2022, 23, 8299. [Google Scholar] [CrossRef]
- Draviam, V.M.; Xie, S.; Sorger, P.K. Chromosome segregation and genomic stability. Curr. Opin. Genet. Dev. 2004, 14, 120–125. [Google Scholar] [CrossRef]
- Aypar, U.; Morgan, W.F.; Baulch, J.E. Radiation-induced genomic instability: Are epigenetic mechanisms the missing link? Int. J. Radiat. Biol. 2011, 87, 179–191. [Google Scholar] [CrossRef]
- Mei, M.; Deng, H.; Lu, Y.; Zhuang, C.; Liu, Z.; Qiu, Q.; Qiu, Y.; Yang, T.C. Mutagenic effects of heavy ion radiation in plants. Adv. Space Res. 1994, 14, 363–372. [Google Scholar] [CrossRef]
- Yokota, Y.; Funayama, T.; Hase, Y.; Hamada, N.; Kobayashi, Y.; Tanaka, A.; Narumi, I. Enhanced micronucleus formation in the descendants of gamma-ray-irradiated tobacco cells: Evidence for radiation-induced genomic instability in plant cells. Mutat. Res. 2010, 691, 41–46. [Google Scholar] [CrossRef]
- Kitamura, S.; Inoue, M.; Ohmido, N.; Fukui, K.; Tanaka, A. Chromosomal rearrangements in interspecific hybrids between Nicotiana gossei Domin and N. tabacum L., obtained by crossing with pollen exposed to helium ion beams or gamma-rays. Nucl. Instr. Meth. Phys. Res. B 2003, 206, 548–552. [Google Scholar] [CrossRef]
- Stoilov, L.; Georgieva, M.; Manova, V.; Liu, L.; Gecheff, K. Karyotype reconstruction modulates the sensitivity of barley genome to radiation-induced DNA and chromosomal damage. Mutagenesis 2013, 28, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Nikolova, I.; Georgieva, M.; Kruppa, K.; Molnar-Lang, M.; Liu, L.; Manova, V.; Stoilov, L. Cytogenetic effects in barley root apical meristem after exposure of dry seeds to lithium ion beams. Genet. Plant Physiol. 2015, 5, 3–9. [Google Scholar]
- Georgieva, M.; Stoilov, L. Assessment of DNA strand breaks induced by bleomycin in barley by the comet assay. Environ. Mol. Mutagen. 2008, 49, 381–387. [Google Scholar] [CrossRef]
- Amosova, A.V.; Zoshchuk, S.A.; Volovik, V.T.; Shirokova, A.V.; Horuzhiy, N.E.; Mozgova, G.V.; Yurkevich, O.Y.; Artyukhova, M.A.; Lemesh, V.A.; Samatadze, T.E.; et al. Phenotypic, biochemical and genomic variability in generations of the rapeseed (Brassica napus L.) mutant lines obtained via chemical mutagenesis. PLoS ONE 2019, 14, e0221699. [Google Scholar] [CrossRef]
- Song, X.; Yan, J.; Zhang, Y.; Li, H.; Zheng, A.; Zhang, Q.; Wang, J.; Bian, Q.; Shao, Z.; Wang, Y.; et al. Gene flow risks from transgenic herbicide-tolerant crops to their wild relatives can be mitigated by utilizing alien chromosomes. Front. Plant Sci. 2021, 12, 670209. [Google Scholar] [CrossRef]
- Gianella, M.; Bradford, K.J.; Guzzon, F. Ecological, (epi)genetic and physiological aspects of bet-hedging in angiosperms. Plant Reprod. 2021, 34, 21–36. [Google Scholar] [CrossRef]
- Suter, L.; Widmer, A. Phenotypic effects of salt and heat stress over three generations in Arabidopsis thaliana. PLoS ONE 2013, 8, e80819. [Google Scholar] [CrossRef]
- Castro, B.M.; Moriuchi, K.S.; Friesen, M.L.; Badri, M.; Nuzhdin, S.V.; Strauss, S.Y.; Cook, D.R.; Wettberg, E. Parental environments and interactions with conspecifics alter salinity tolerance of offspring in the annual Medicago truncatula. J. Ecol. 2013, 101, 1281–1287. [Google Scholar] [CrossRef]
- Ou, X.; Zhang, Y.; Xu, C.; Lin, X.; Zang, Q.; Zhuang, T.; Jiang, L.; von Wettstein, D.; Liu, B. Transgenerational inheritance of modified DNA methylation patterns and enhanced tolerance induced by heavy metal stress in rice (Oryza sativa L.). PLoS ONE 2012, 7, e41143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.; Puthur, J.T. Seed priming as a cost effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Nafees, M.; Chen, J.; Darras, A.; Ferrante, A.; Hancock, J.T.; Ashraf, M.; Zaid, A.; Latif, N.; Corpas, F.J.; et al. Chemical priming enhances plant tolerance to salt stress. Front. Plant Sci. 2022, 13, 946922. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Walias, F.J.; García, M.; Moreno, M.; Giannoukos, I.; González, N.; Sanz-García, E.; Necira, K.; Canto, T.; Tenllado, F. Transgenerational tolerance to salt and osmotic stresses induced by plant virus infection. Int. J. Mol. Sci. 2022, 23, 12497. [Google Scholar] [CrossRef]
- Childs, D.Z.; Metcalf, C.J.; Rees, M. Evolutionary bet-hedging in the real world: Empirical evidence and challenges revealed by plants. Proc. Biol. Sci. 2010, 277, 3055–3064. [Google Scholar] [CrossRef] [Green Version]
- Perez, I.B.; Brown, P.J. The role of ROS signaling in cross-tolerance: From model to crop. Front. Plant Sci. 2014, 5, 754. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Li, Z.G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and possible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Kumar, R.R.; Praveen, S. Plant transcriptional regulation in modulating cross-tolerance to stress. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D.J., Fujita, M., Huang, B., Eds.; Academic Press: London, UK, 2020; pp. 231–245. [Google Scholar]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Jiang, J.; Wang, C.Q.; Dehesh, K. ORA59 and EIN3 interaction couples jasmonate-ethylene synergistic action to antagonistic salicylic acid regulation of PDF expression. J. Integr. Plant Biol. 2017, 59, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [Green Version]
- Salvi, P.; Manna, M.; Kaur, H.; Thakur, T.; Gandass, N.; Bhatt, D.; Muthamilarasan, M. Phytohormone signaling and crosstalk in regulating drought stress response in plants. Plant Cell Rep. 2021, 40, 1305–1329. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Tada, Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P.; Indoliya, Y.; Chauhan, A.S.; Singh, P.; Singh, P.K.; Singh, P.C.; Srivastava, S.; Pande, V.; Chakrabarty, D. Auxin-salicylic acid cross-talk ameliorates OsMYB–R1 mediated defense towards heavy metal, drought and fungal stress. J. Hazard. Mater. 2020, 399, 122811. [Google Scholar] [CrossRef]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene signaling under stressful environments: Analyzing collaborative knowledge. Plants 2022, 11, 2211. [Google Scholar] [CrossRef]
- Eriksson, G. Induction of waxy mutants in maize by acute and chronic gamma irradiation. Hereditas 1963, 50, 161–178. [Google Scholar] [CrossRef]
- Joiner, M.C.; Lambin, P.; Marples, B. Adaptive response and induced resistance. C. R. Acad. Sci. III 1999, 322, 167–175. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Agathokleous, E. Hormesis: Transforming disciplines that rely on the dose response. IUBMB Life 2022, 74, 8–23. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P. How does hormesis impact biology, toxicology, and medicine? NPJ Aging Mech. Dis. 2017, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ancel, P.; Lallemand, S. Sur la protection contre l’action des rayons X par une irradiation prealable (radiophylaxie). C. R. Soc. Biol. 1928, 99, 1588–1590. [Google Scholar]
- Calabrese, E.J.; Baldwin, L.A. Chemical hormesis: Its historical foundations as a biological hypothesis. Hum. Exp. Toxicol. 2000, 19, 2–31. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Preconditioning is hormesis part I: Documentation, dose-response features and mechanistic foundations. Pharmacol. Res. 2016, 110, 242–264. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense priming: An adaptive part of induced resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Westman, S.M.; Kloth, K.J.; Hanson, J.; Ohlsson, A.B.; Albrectsen, B.R. Defence priming in Arabidopsis—A meta-analysis. Sci. Rep. 2019, 9, 13309. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Mattson, M.P. Hormesis provides a generalized quantitative estimate of biological plasticity. J. Cell Commun. Signal. 2011, 5, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef]
- Guéguen, Y.; Bontemps, A.; Ebrahimian, T.G. Adaptive responses to low doses of radiation or chemicals: Their cellular and molecular mechanisms. Cell Mol. Life Sci. 2019, 76, 1255–1273. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Harayama, H.; Calabrese, E.J. Temperature-induced hormesis in plants. J. For. Res. 2019, 30, 13–20. [Google Scholar] [CrossRef]
- Garzon, C.D.; Flores, F.J. Hormesis: Biphasic dose-responses to fungicides in plant pathogens and their potential threat to agriculture, fungicides—Showcases of integrated plant disease management from around the world. In Fungicides: Showcases of Integrated Plant Disease Management from around the World; Nita, M., Ed.; InTechOpen Limited: London, UK, 2013; pp. 311–328. [Google Scholar]
- Duarte-Sierra, A.; Tiznado-Hernández, M.E.; Jha, D.K.; Janmeja, N.; Arul, J. Abiotic stress hormesis: An approach to maintain quality, extend storability, and enhance phytochemicals on fresh produce during postharvest. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3659–3682. [Google Scholar] [CrossRef]
- Cook, R.; Calabrese, E.J. The importance of hormesis to public health. Cien. Saude Colet. 2007, 12, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Niazi, N.K.; Rinklebe, J.; Bundschuh, J.; Dumat, C.; Pinelli, E. Trace elements-induced phytohormesis: A critical review and mechanistic interpretation. Crit. Rev. Environ. Sci. Technol. 2019, 50, 1984–2015. [Google Scholar] [CrossRef]
- Tang, Y.-T.; Qiu, R.-L.; Zeng, X.-W.; Ying, R.-R.; Yu, F.-M.; Zhou, X.-Y. Lead, zinc, cadmium hyperaccumulation and growth stimulation in Arabis paniculata Franch. Environ. Exp. Bot. 2009, 66, 126–134. [Google Scholar] [CrossRef]
- Jia, L.; Liu, Z.; Chen, W.; Ye, Y.; Yu, S.; He, X. Hormesis effects induced by cadmium on growth and photosynthetic performance in a Hyperaccumulator, Lonicera japonica Thunb. J. Plant Growth Regul. 2015, 34, 13–21. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Plant hormesis and Shelford’s tolerance law curve. J. For. Res. 2021, 32, 1789–1802. [Google Scholar] [CrossRef]
- Wei, C.; Jiao, Q.; Agathokleous, E.; Liu, H.; Li, G.; Zhang, J.; Fahad, S.; Jiang, Y. Hormetic effects of zinc on growth and antioxidant defense system of wheat plants. Sci. Total Environ. 2022, 807, 50992. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [Green Version]
- Belz, R.G.; Piepho, H.P. Variability of hormetic dose responses of the antiauxin PCIB on Lactuca sativa in a plant bioassay. Weed Res. 2013, 53, 418–428. [Google Scholar] [CrossRef]
- Belz, R.G.; Duke, S.O. Herbicides and plant hormesis. Pest Manag. Sci. 2014, 70, 698–707. [Google Scholar] [CrossRef]
- Jalal, A.; Oliveira Junior, J.C.; Ribeiro, J.S.; Fernandes, G.C.; Mariano, G.G.; Trindade, V.D.R.; Reis, A.R.D. Hormesis in plants: Physiological and biochemical responses. Ecotoxicol. Environ. Saf. 2021, 207, 111225. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Kuang, L.; He, X.; Bai, W.; Ding, Y.; Zhang, Z.; Zhao, Y.; Chai, Z. Effects of rare earth oxide nanoparticles on root elongation of plants. Chemosphere 2010, 78, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, C.; Cabot, C.; Martos, S.; Gallego, B.; Barceló, J. Do toxic ions induce hormesis in plants? Plant Sci. 2013, 212, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Feng, Z.; Iavicoli, I.; Calabrese, E.J. The two faces of nanomaterials: A quantification of hormesis in algae and plants. Environ. Int. 2019, 131, 105044. [Google Scholar] [CrossRef] [PubMed]
- Małkowski, E.; Sitko, K.; Szopiński, M.; Gieroń, Ż.; Pogrzeba, M.; Kalaji, H.M.; Zieleźnik-Rusinowska, P. Hormesis in plants: The role of oxidative stress, auxins and photosynthesis in corn treated with Cd or Pb. Int. J. Mol. Sci. 2020, 21, 2099. [Google Scholar] [CrossRef] [Green Version]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Biphasic effect of abscisic acid on plants: A hormetic viewpoint. Botany 2018, 96, 637–642. [Google Scholar] [CrossRef]
- Sanyal, R.P.; Misra, H.S.; Saini, A. Heat-stress priming and alternative splicing-linked memory. J. Exp. Bot. 2018, 69, 2431–2434. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Guo, B. Effects of light intensity on the growth, photosynthetic characteristics, and flavonoid content of Epimedium pseudowushanense B.L.Guo. Molecules 2016, 21, 1475. [Google Scholar] [CrossRef] [Green Version]
- Cortés, F.; Domínguez, I.; Mateos, S.; Piñero, J.; Mateos, J.C. Evidence for an adaptive response to radiation damage in plant cells conditioned with X-rays or incorporated tritium. Int. J. Radiat. Biol. 1990, 57, 537–541. [Google Scholar] [CrossRef]
- Asare, A.T.; Mensah, F.; Acheampong, S.; Asare-Bediako, E.; Armah, J. Effects of gamma irradiation S. on agromorphological characteristics of okra (Abelmoschus esculentus L. Moench.). Adv. Agric. 2017, 2017, e2385106. [Google Scholar] [CrossRef] [Green Version]
- Volkova, P.Y.; Duarte, G.T.; Soubigou-Taconnat, L.; Kazakova, E.A.; Pateyron, S.; Bondarenko, V.S.; Bitarishvili, S.V.; Makarenko, E.S.; Churyukin, R.S.; Lychenkova, M.A.; et al. Early response of barley embryos to low- and high-dose gamma irradiation of seeds triggers changes in the transcriptional profile and an increase in hydrogen peroxide content in seedlings. J. Agron. Crop Sci. 2020, 206, 277–295. [Google Scholar] [CrossRef]
- Antúnez-Ocampo, O.M.; Cruz-Izquierdo, S.; Mendoza-Onofre, L.E.; Sandoval-Villa, M.; Santacruz-Varela, A.; Cruz-Torres, E.D.L.; Peña-Lomelí, A. Growth dynamics of morphological and reproductive traits of Physalis peruviana L. M1 plants obtained from seeds irradiated with gamma rays. Not. Bot. Horti Agrobo. 2020, 48, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Geras’kin, S.; Churyukin, R.; Volkova, P. Radiation exposure of barley seeds can modify the early stages of plants’ development. J. Environ. Radioact. 2017, 177, 71–83. [Google Scholar] [CrossRef]
- Volkova, P.Y.; Bondarenko, E.V.; Kazakova, E.A. Radiation hormesis in plants. Curr. Opin. Toxicol. 2022, 30, 100334. [Google Scholar] [CrossRef]
- Lushchak, V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants and animals. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 175–190. [Google Scholar] [CrossRef]
- Feinendegen, L.E.; Pollycove, M.; Sondhaus, C.A. Responses to low doses of ionizing radiation in biological systems. Nonlinearity Biol. Toxicol. Med. 2004, 2, 143–171. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Zhang, L.; Feng, W.; Xu, H.; Wang, L.; Jiao, Z. ROS and ABA signaling are involved in the growth stimulation induced by low-dose gamma irradiation in Arabidopsis seedling. Appl. Biochem. Biotechnol. 2015, 175, 1490–1506. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Burke, P.; Arkhipov, A.; Kuchma, N.; James, S.J.; Kovalchuk, I.; Pogribny, I. Genome hypermethylation in Pinus silvestris of Chernobyl-a mechanism for radiation adaptation? Mutat. Res. 2003, 529, 13–20. [Google Scholar] [CrossRef]
- Kinoshita, T.; Seki, M. Epigenetic memory for stress response and adaptation in plants. Plant Cell Physiol. 2014, 55, 1859–1863. [Google Scholar] [CrossRef]
Figure 1.
Plants can be subjected to stress due to both biotic factors, such as pathogens, pests, and direct animal and human influences, and abiotic factors, such as diverse climatic changes, soil and air pollution, and magnetic fields. These stressors can act individually, simultaneously, or in various combinations, and disrupt plant homeostasis, impede growth, and impact development in wild populations and cultivated field crops.
Figure 1.
Plants can be subjected to stress due to both biotic factors, such as pathogens, pests, and direct animal and human influences, and abiotic factors, such as diverse climatic changes, soil and air pollution, and magnetic fields. These stressors can act individually, simultaneously, or in various combinations, and disrupt plant homeostasis, impede growth, and impact development in wild populations and cultivated field crops.

Figure 2.
Overview of the main types of DNA lesions observed in plant genomes in response to abiotic stress. Radiation as a stressor can lead to the accumulation of various types of DNA damage in plant cells, including direct DNA damage (SSBs, DSBs, and cluster damage), photolesions (6-4 photoproducts and cyclobutane-pyrimidine dimers), base damage, and interstrand crosslinks, depending on the radiation properties (high or low LET). Temperature, water, and salt stress can cause direct DNA lesions and oxidative damage due to an overproduction of ROS. Heat stress can also lead to hydrolytic lesions, similar to those caused by heavy metal stress. Primary alkylated damage is not connected to the stress factors shown in the figure as it results from the application of alkylating agents, which are not typical stress factors for plants. The relationships between specific DNA lesions and stress factors are indicated by arrows.
Figure 2.
Overview of the main types of DNA lesions observed in plant genomes in response to abiotic stress. Radiation as a stressor can lead to the accumulation of various types of DNA damage in plant cells, including direct DNA damage (SSBs, DSBs, and cluster damage), photolesions (6-4 photoproducts and cyclobutane-pyrimidine dimers), base damage, and interstrand crosslinks, depending on the radiation properties (high or low LET). Temperature, water, and salt stress can cause direct DNA lesions and oxidative damage due to an overproduction of ROS. Heat stress can also lead to hydrolytic lesions, similar to those caused by heavy metal stress. Primary alkylated damage is not connected to the stress factors shown in the figure as it results from the application of alkylating agents, which are not typical stress factors for plants. The relationships between specific DNA lesions and stress factors are indicated by arrows.

Figure 3.
A simplified illustration of the interaction between plant responses induced by primary abiotic stress factors and those caused by secondary and tertiary stressors. The primary stress factors are indicated in the inner section of the figure, while the secondary and tertiary stressors are shown in the outer section. The arrows depict the relationship between the various stressors.
Figure 3.
A simplified illustration of the interaction between plant responses induced by primary abiotic stress factors and those caused by secondary and tertiary stressors. The primary stress factors are indicated in the inner section of the figure, while the secondary and tertiary stressors are shown in the outer section. The arrows depict the relationship between the various stressors.

Figure 4.
Hypothetical curves of the dose–response relationship. I. Monotonic type (blue background). (a) Linear, sublinear and supralinear curves with no clear threshold. The lowest dose at which an effect can be observed is named the threshold limit value (TLV); (b) threshold curves; (c) S-shaped curves (saturation curve). The S-shaped curves are characterized by an increasing trend, reflecting the beneficial effects on the plant organism as the dose/concentration gradually increases. II. Non-monotonic (yellow background). (d) U-shaped or J-shaped curves; (e). inverted U- or β-shaped curves. This type of curve reflects the hormetic response at low doses; (f) biphasic curve; (g) N-shaped dose–response curves of HRS/IRR response in terms of DSBs and cell survival (adapted from Thomas et al. [104] and Devic et al. [105]). III. Curves without dose–response relationship (WDR) (green background). (h) Binary curve and bystander response (adapted from Prise and O’Sullivan [106]).
Figure 4.
Hypothetical curves of the dose–response relationship. I. Monotonic type (blue background). (a) Linear, sublinear and supralinear curves with no clear threshold. The lowest dose at which an effect can be observed is named the threshold limit value (TLV); (b) threshold curves; (c) S-shaped curves (saturation curve). The S-shaped curves are characterized by an increasing trend, reflecting the beneficial effects on the plant organism as the dose/concentration gradually increases. II. Non-monotonic (yellow background). (d) U-shaped or J-shaped curves; (e). inverted U- or β-shaped curves. This type of curve reflects the hormetic response at low doses; (f) biphasic curve; (g) N-shaped dose–response curves of HRS/IRR response in terms of DSBs and cell survival (adapted from Thomas et al. [104] and Devic et al. [105]). III. Curves without dose–response relationship (WDR) (green background). (h) Binary curve and bystander response (adapted from Prise and O’Sullivan [106]).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Georgieva, M.; Vassileva, V. Stress Management in Plants: Examining Provisional and Unique Dose-Dependent Responses. Int. J. Mol. Sci. 2023, 24, 5105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065105
AMA Style
Georgieva M, Vassileva V. Stress Management in Plants: Examining Provisional and Unique Dose-Dependent Responses. International Journal of Molecular Sciences. 2023; 24(6):5105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065105
Chicago/Turabian StyleGeorgieva, Mariyana, and Valya Vassileva. 2023. "Stress Management in Plants: Examining Provisional and Unique Dose-Dependent Responses" International Journal of Molecular Sciences 24, no. 6: 5105. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065105
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.