
Knockout Mutants of OsPUB7 Generated Using CRISPR/Cas9 Revealed Abiotic Stress Tolerance in Rice
 , ,
, ,
Abstract
:1. Introduction
2. Results
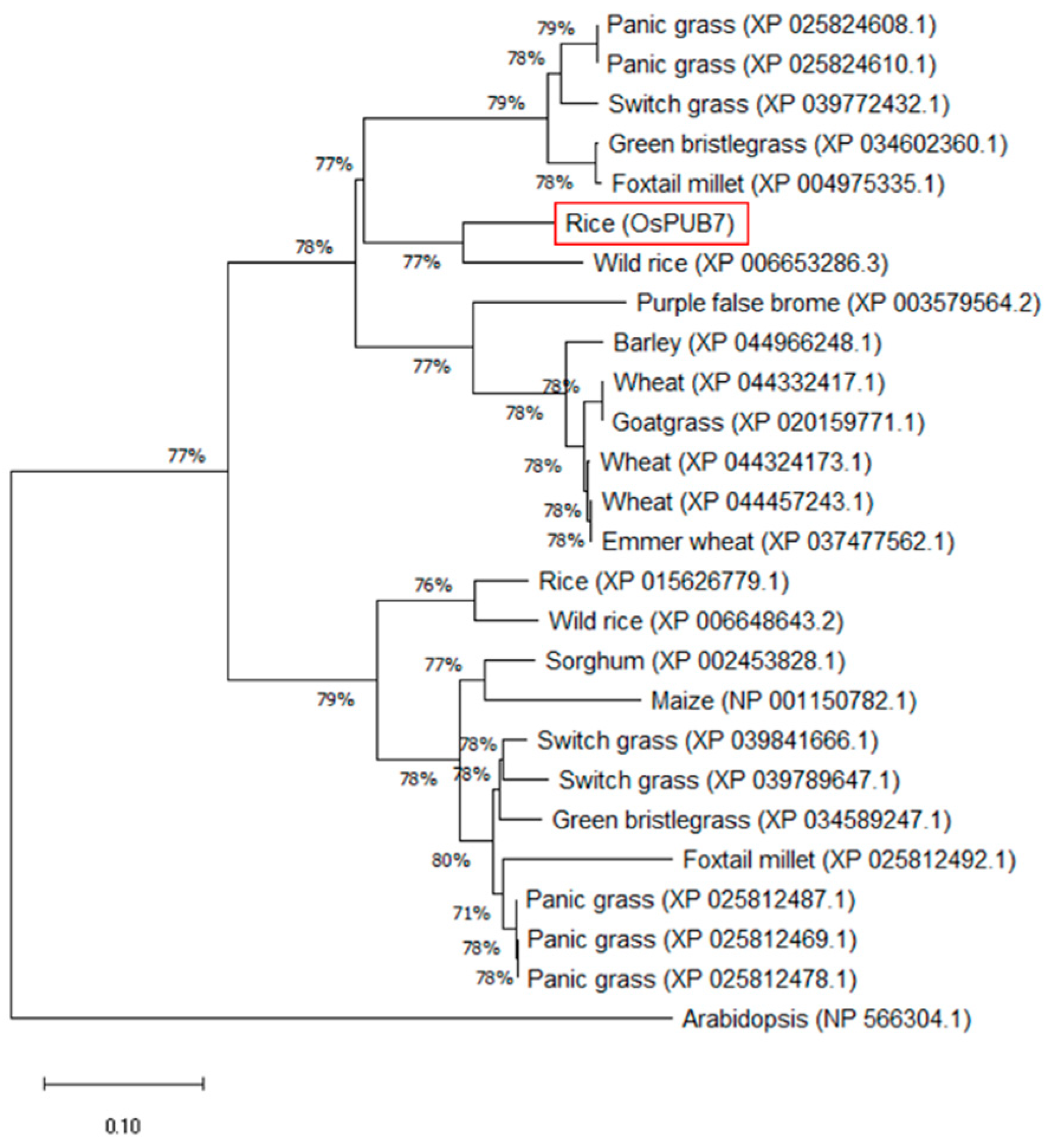
2.1. Analysis of the Genetic Relationship of OsPUB7 among Plant Species
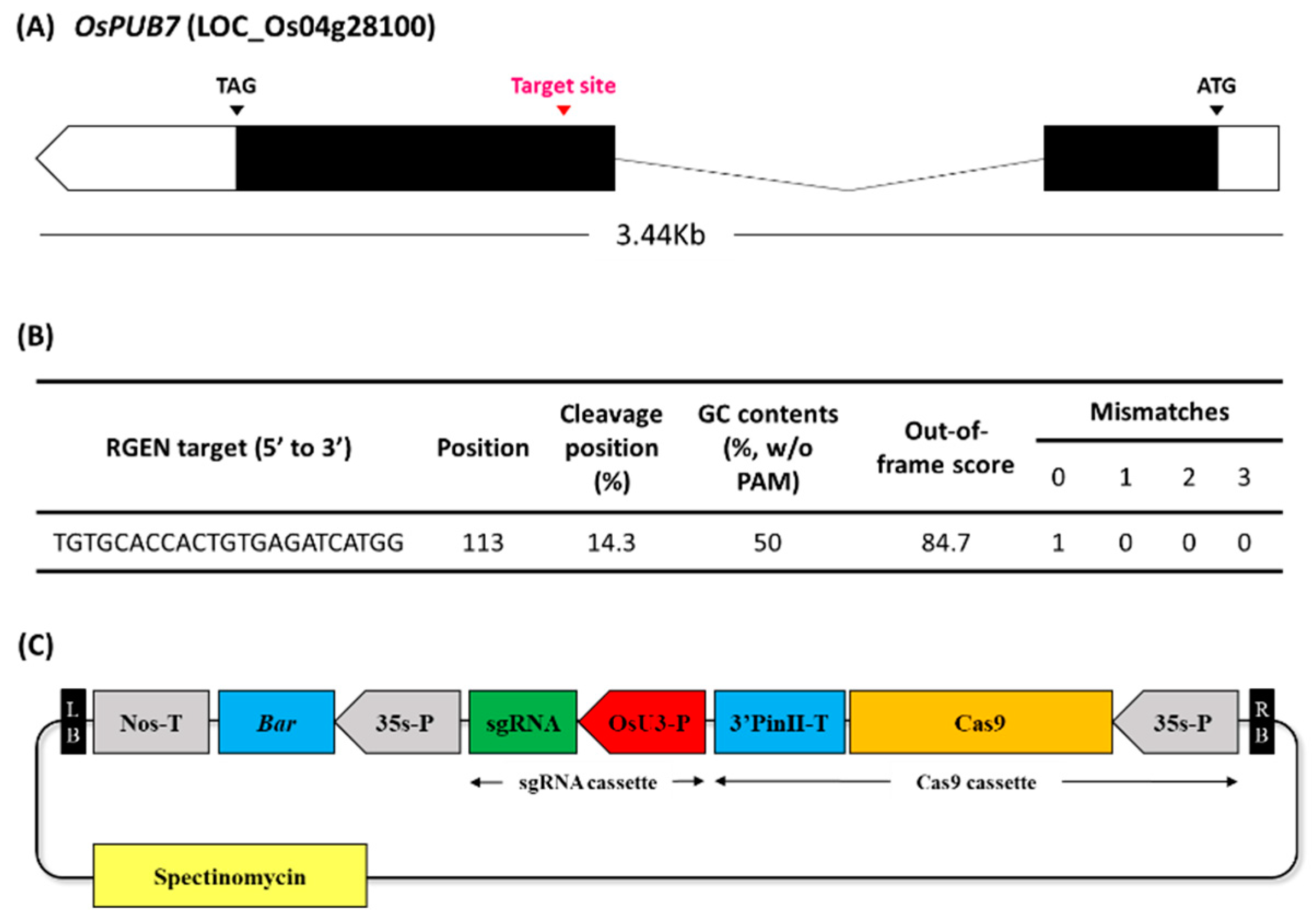
2.2. Production of OsPUB7-Edited Lines and Analysis of Mutation Types
2.2.1. CRISPR/Cas9-Mediated Editing of OsPUB7
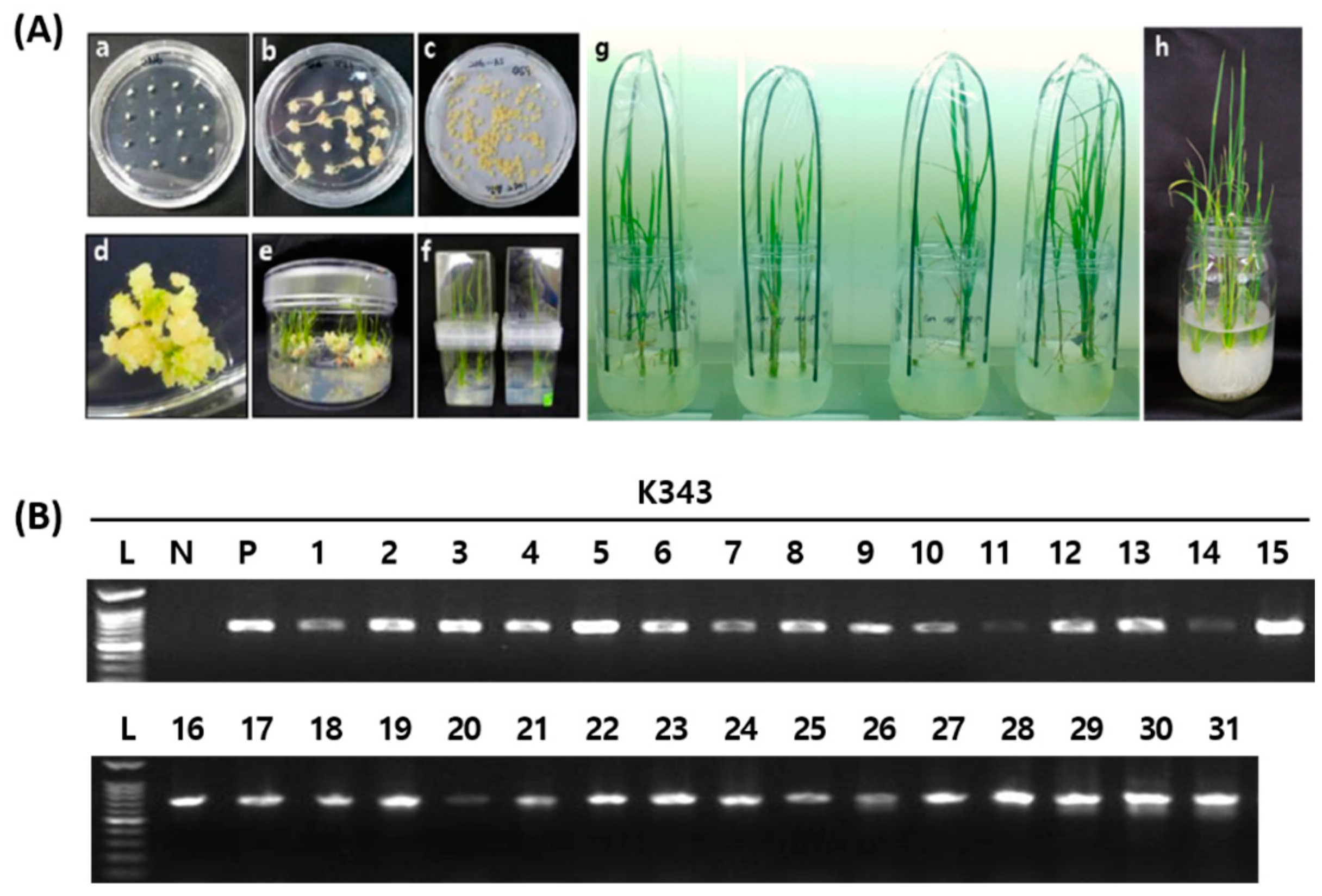
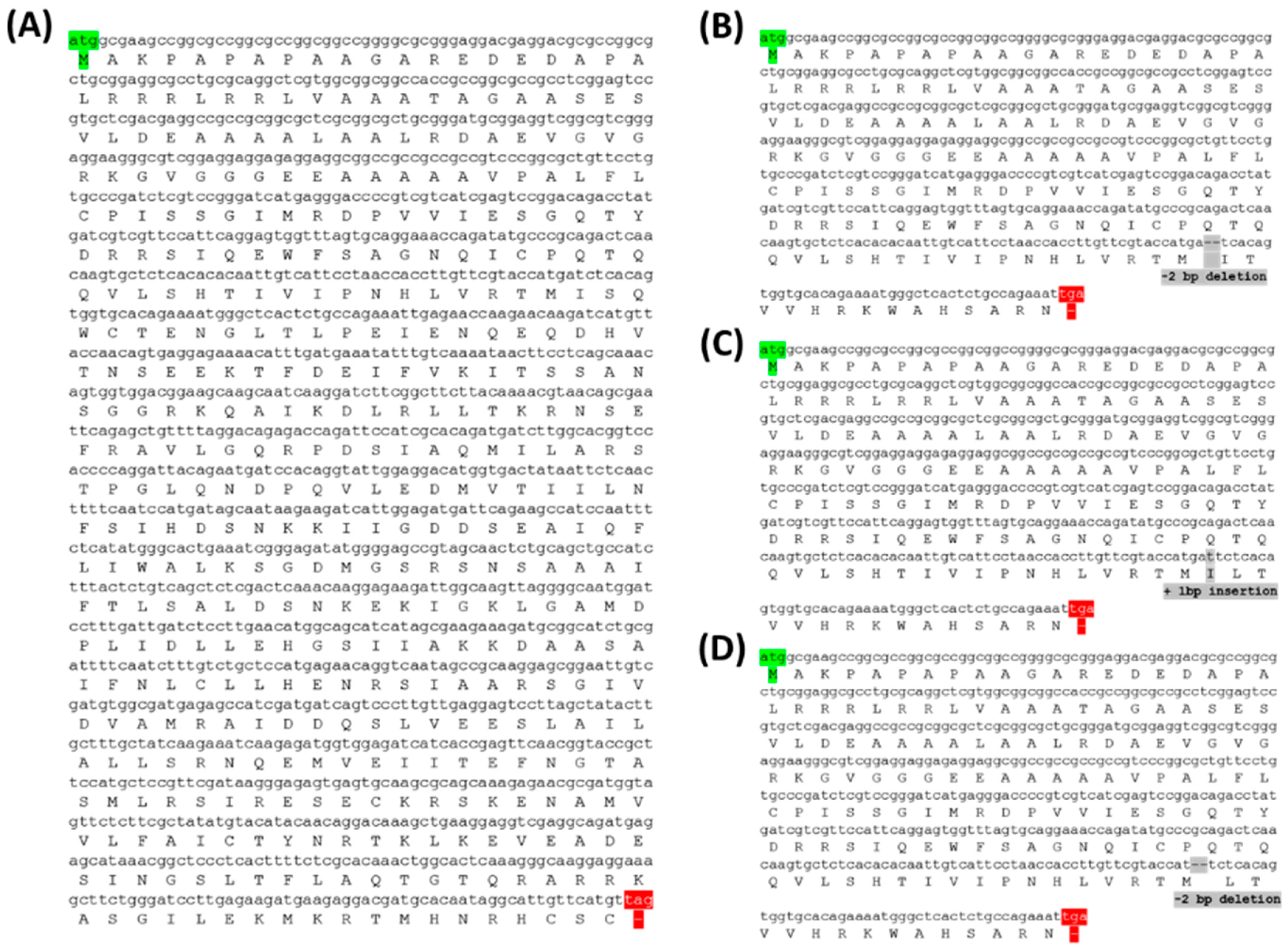
2.2.2. Analysis of Mutation Types Using Next-Generation Sequencing (NGS)
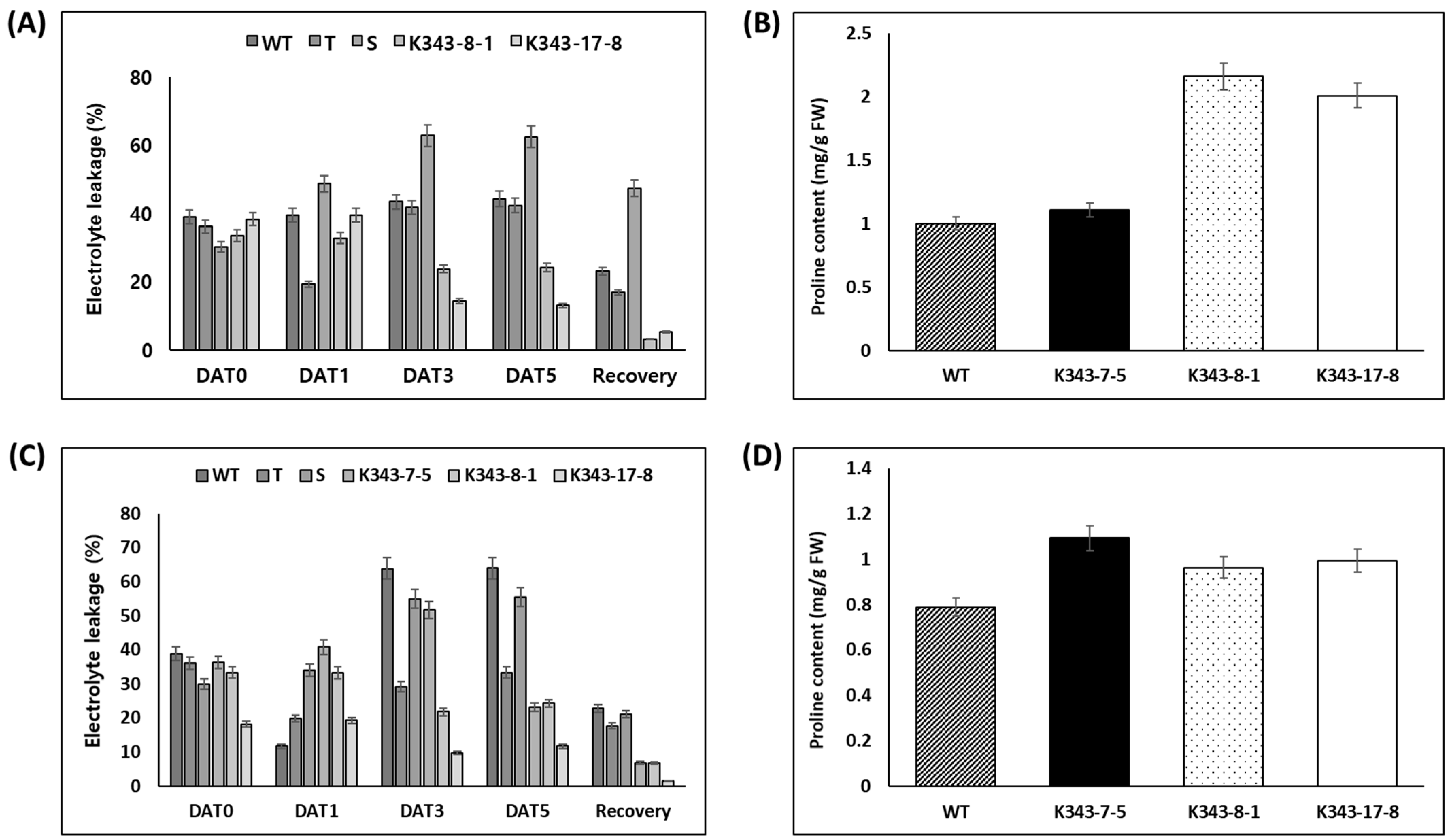
2.3. Selection of Tolerant Gene-Edited Lines and Examination of Physiological Characteristics According to Abiotic Stress Treatment
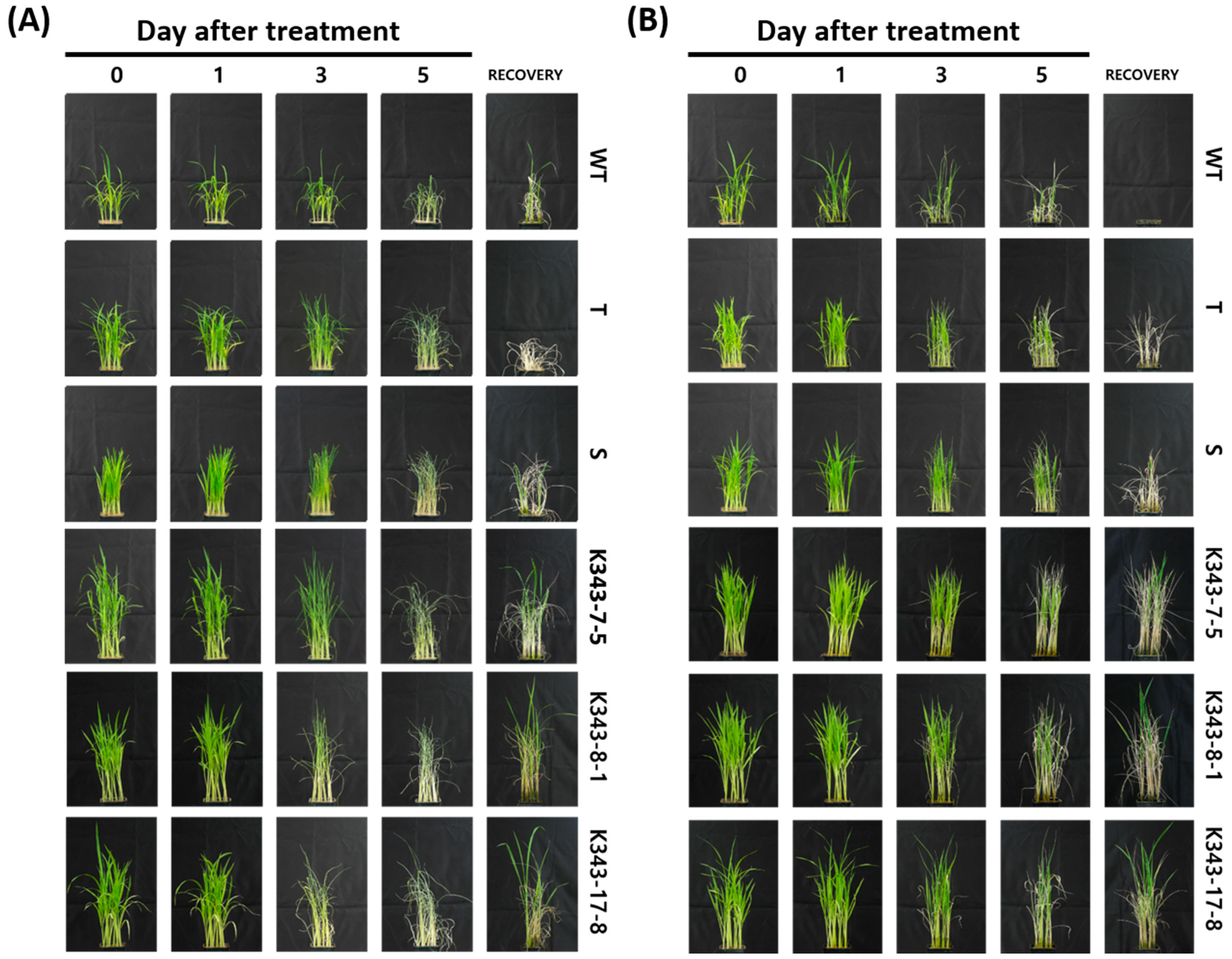
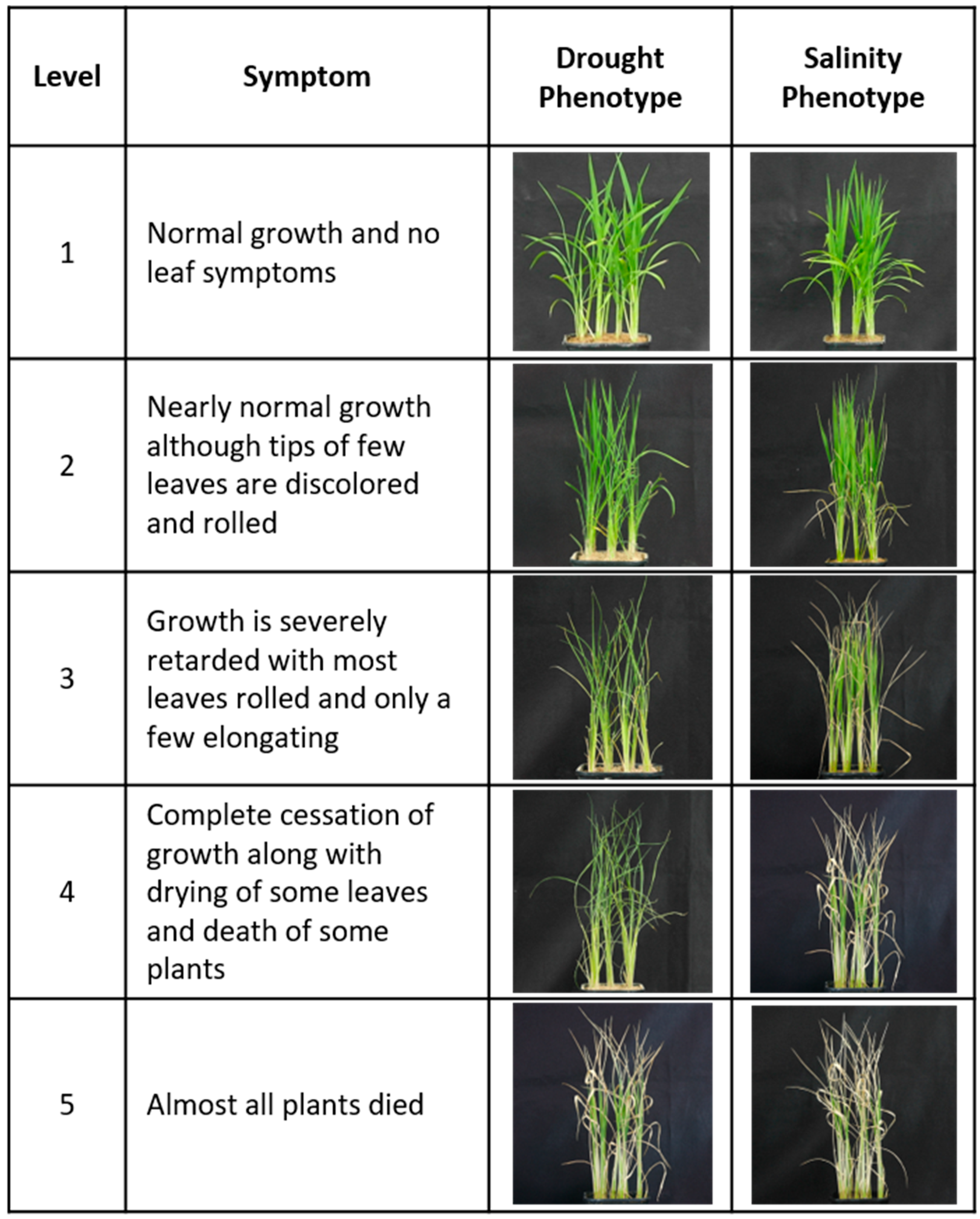
2.3.1. Screening of Drought and Salinity Stress
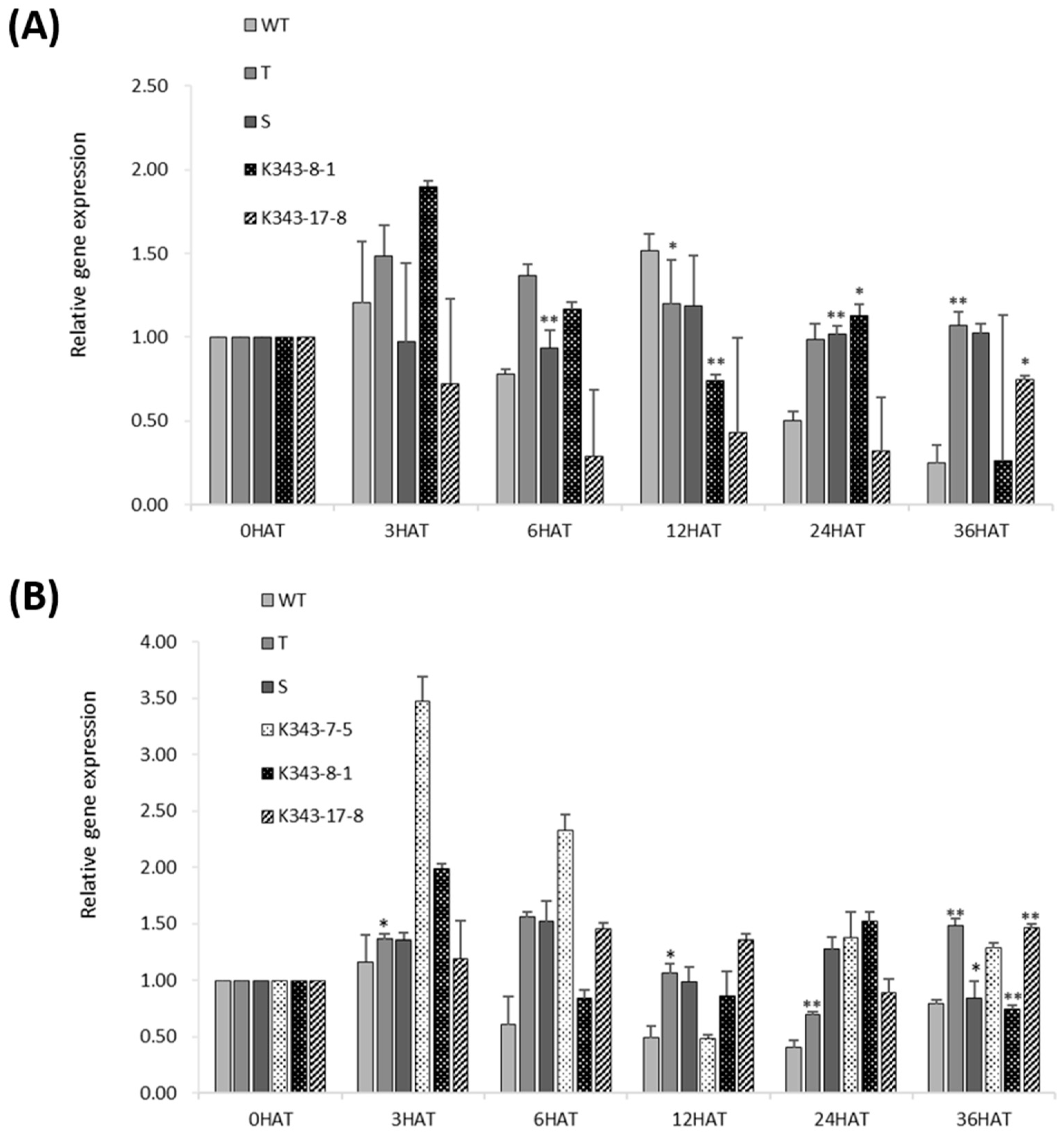
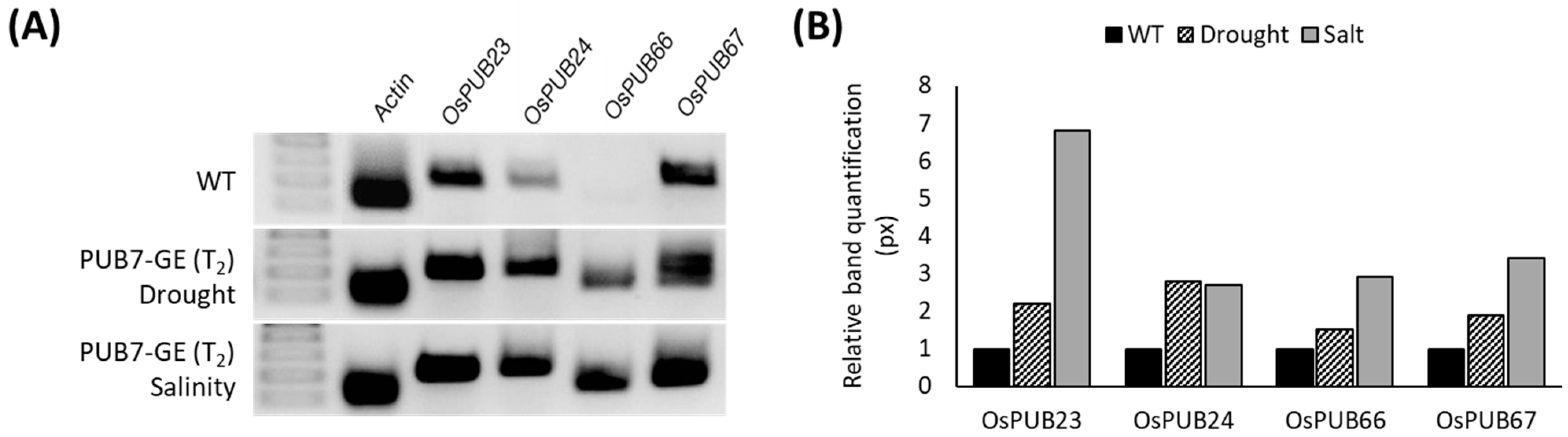
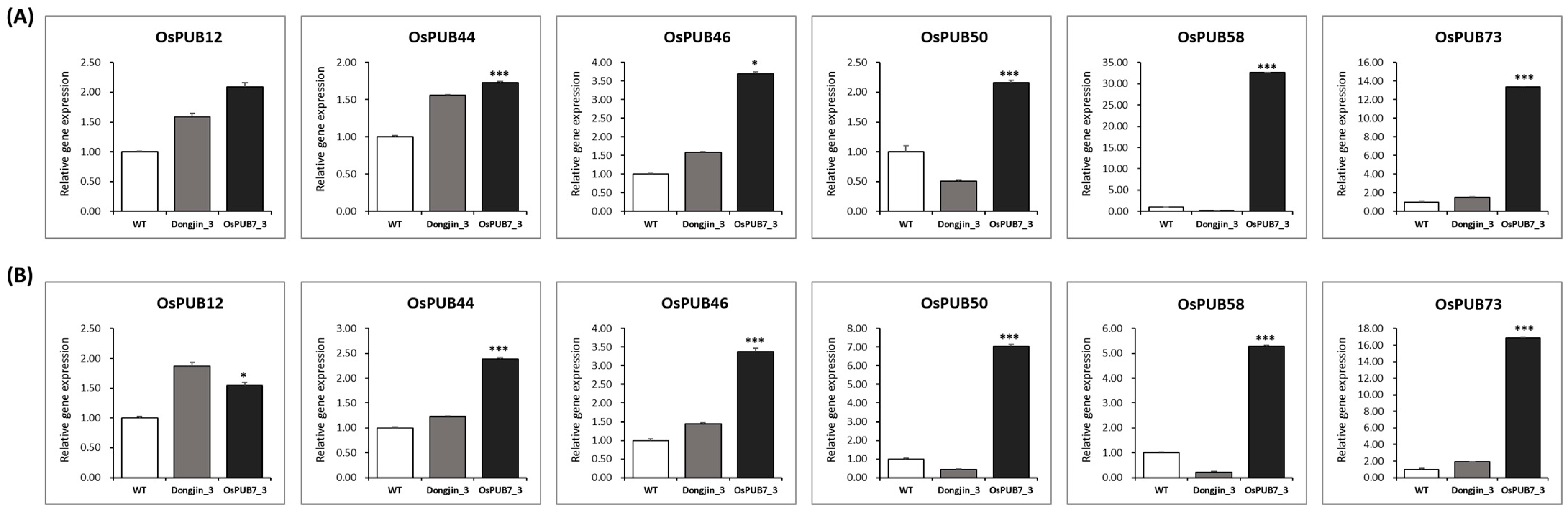
2.3.2. Determination of Physiological Characteristics and Analysis of mRNA Expression
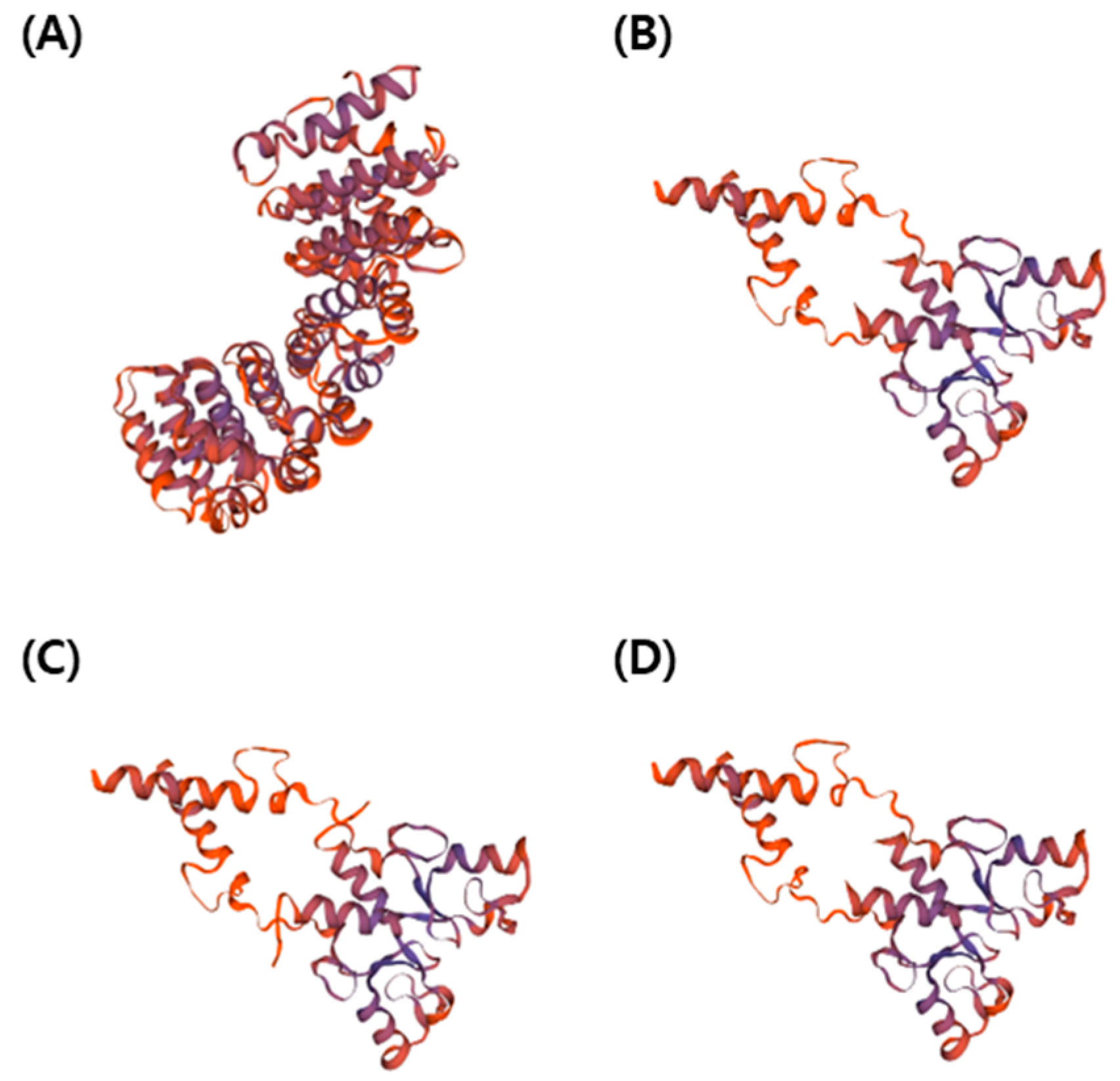
2.4. Analysis of Protein Structure in PUB7-GE
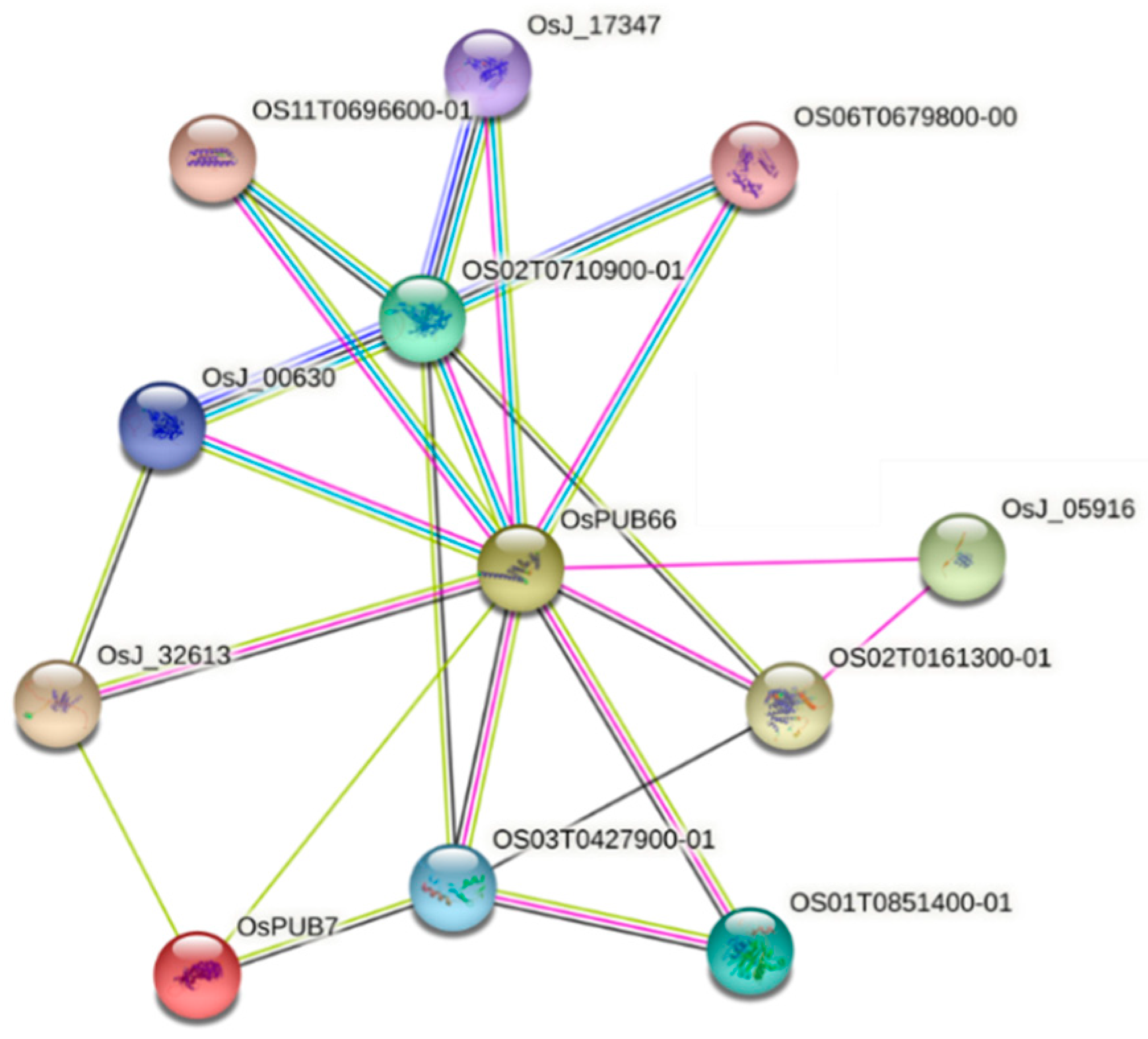
2.5. Analysis of Protein–Protein Interactions
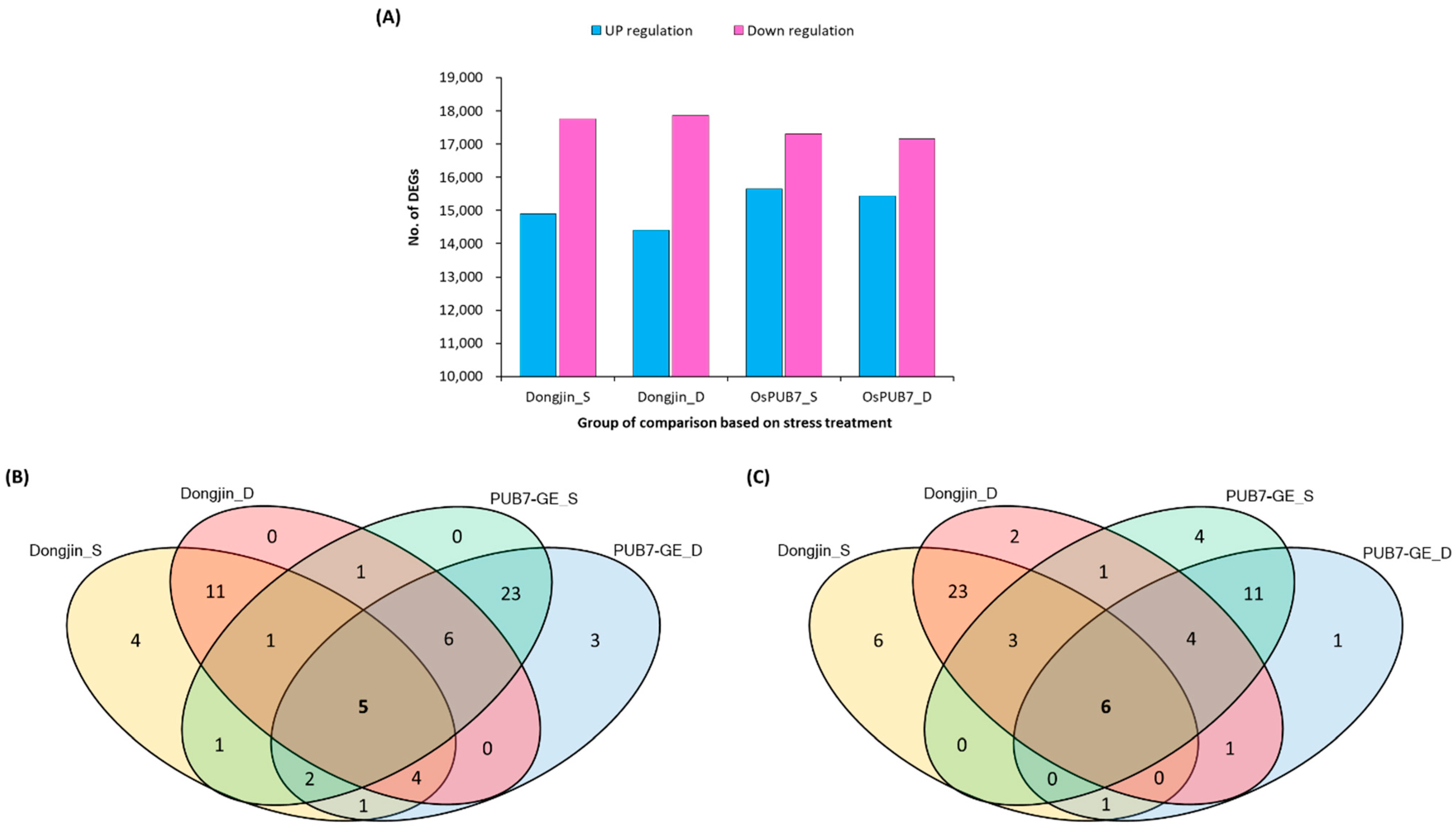
2.6. Analysis of RNA-Sequencing Data of PUB7-GE
2.7. Investigation of Agronomic Traits
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Analysis of Phylogenetic Tree
4.3. Selection of Target Sequences and Vector Construction
4.4. Agrobacterium-Mediated Transformation and Analysis of Mutation Types Using NGS
4.5. Screening for Drought and Salt Stress Resistance
4.6. Measurement of the Physiological Parameters of Transgenic Rice
4.7. mRNA Expression Analysis
4.8. Analysis of OsPUB7-GE
4.9. Protein–Protein Interaction Network Analysis
4.10. RNA-Sequencing Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiqui-tin-proteasome system. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 182–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, S.M.; Genest, O.; Wickner, S. Protein rescue from aggregates by powerful molecular chaperone ma-chines. Nat. Rev. Mol. Cell Biol. 2013, 14, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.; Millar, A. Protein turnover in plant biology. Nat. Plants 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santner, A.; Estelle, M. The ubiquitin-proteasome system regulates plant hormone signaling. Plant J. 2010, 61, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. The ubiquitin-proteasome proteolytic pathway. Cell 1994, 79, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Schwartz, D.; Elias, J.E.; Thoreen, C.C.; Cheng, D.; Marsischky, G.; Roelofs, J.; Finley, D.; Gygi, S.P. A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol. 2003, 21, 921–926. [Google Scholar] [CrossRef]
- Sadowski, M.; Suryadinata, R.; Tan, A.R.; Roesley, S.N.A.; Sarcevic, B. Protein monoubiquitination and polyubiquitination generate structural diversity to control distinct biological processes. IUBMB Life 2012, 64, 136–142. [Google Scholar] [CrossRef]
- Kirisako, T.; Kamei, K.; Murata, S.; Kato, M.; Fukumoto, H.; Kanie, M.; Sano, S.; Tokunaga, F.; Tanaka, K.; Iwai, K. A ubiquitin ligase complex assembles linear polyubiquitin chains. EMBO J. 2016, 25, 4877–4887. [Google Scholar] [CrossRef]
- Berndsen, C.; Wolberger, C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014, 21, 301–307. [Google Scholar] [CrossRef]
- Ciechanover, A. The ubiquitin–proteasome pathway: On protein death and cell life. EMBO J. 1998, 17, 7151–7160. [Google Scholar] [CrossRef] [Green Version]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta-Mol. Cell Res. 2004, 1695, 55–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecker, S.H.; Goldberg, A.L.; Mitch, W.E. Protein degradation by the ubiquitin-proteasome pathway in normal and disease states. J. Am. Soc. Nephrol. 2006, 177, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Kim, W.T. Classification and interaction modes of 40 rice E2 ubiquitin-conjugating enzymes with 17 rice ARM-U-box E3 ubiquitin ligases. Biochem. Biophys. Res. Commun. 2014, 444, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Patterson, C. A new gun in town: The U box is a ubiquitin ligase domain. Sci. STKE 2002, 2002, PE4. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Koonin, E.V. The U box is a modified RING finger: A common domain in ubiquitination. Curr. Biol. 2000, 10, R132–R134. [Google Scholar] [CrossRef] [Green Version]
- Smalle, J.; Vierstra, R.D. The ubiquitin 26S proteasome proteolytic pathway. Annu. Rev. Plant Biol. 2004, 55, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Callis, J. Ubiquitin ligases mediate growth and development by promoting protein death. Curr. Opin. Plant Biol. 2007, 10, 624–632. [Google Scholar] [CrossRef]
- Koegl, M.; Hoppe, T.; Schlenker, S.; Ulrich, H.D.; Mayer, T.U.; Jentsch, S. A novel ubiquitination factor, E4, is involved in multiubiquitin chain assembly. Cell 1999, 96, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, S.; Yada, M.; Matsumoto, M.; Ishida, N.; Nakayama, K.I. U Box Proteins as a New Family of Ubiquitin-Protein Ligases. J. Biol. Chem. 2001, 276, 33111–33120. [Google Scholar] [CrossRef] [Green Version]
- Mudgil, Y.; Shiu, S.H.; Stone, S.L.; Salt, J.N.; Goring, D.R. A Large Complement of the Predicted Arabidopsis ARM Repeat Proteins Are Members of the U-Box E3 Ubiquitin Ligase Family. Plant Physiol. 2004, 134, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Yee, D.; Goring, D.R. The diversity of plant U-box E3 ubiquitin ligases: From upstream activators to down-stream target substrates. J. Exp. Bot. 2009, 60, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.Y.; Cheng, J.K.; Zhu, Y.J.; Ding, Y.L.; Meng, J.J.; Chen, Z.Z.; Xie, Q.; Guo, Y.; Li, J.; Yang, S.; et al. Degradation of the ABA co-receptor ABI1 by PUB12/13 U-box E3ligases. Nat. Commun. 2015, 6, 8630–8642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, D.H.; Ahn, M.Y.; Park, K.Y.; Kim, E.Y.; Kim, W.T. TheN-terminal UND motif of the Arabidopsis U-Box E3 Ligase PUB18 Is critical forthe negative regulation of ABA-mediated stomatal movement anddetermines its ubiquitination specificity for Exocyst Subunit Exo70B1. Plant Cell 2016, 28, 2952–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.F.; Li, Y.B.; Fei, F.; Wang, Z.K.; Wang, W.; Cao, A.Z.; Liu, Y.; Han, S.; Xing, L.P.; Wang, H.Y.; et al. E3 ubiquitin ligase gene CMPG1-V from Haynaldia villosa L. contributes to powdery mildew resistance in common wheat (Triticum aestivum L.). Plant J. 2015, 84, 154–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, D.; Cao, Y.; Sun, X.; Espinoza, C.; Nguyen, C.T.; Liang, Y.; Stacey, G. Arabidopsis E3 ubiquitin ligase PLANT U-BOX13 (PUB13) regulates chitin receptor Lysin motif receptor kinase5 (LYK5) protein abundance. New Phytol. 2017, 214, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.R.; Park, C.H.; Venu, R.C.; Gough, J.; Wang, G.L. Classification, expression pattern, and E3 ligase activity assay of rice U-box-containing proteins. Mol. Plant 2008, 1, 800–815. [Google Scholar] [CrossRef] [Green Version]
- Park, J.J.; Yi, J.; Yoon, J.; Cho, L.H.; Ping, J.; Jeong, H.J.; Cho, S.K.; Kim, W.T.; An, G. OsPUB15, an E3 ubiquitin ligase, functions to reduce cellular oxidative stress during seedling establishment. Plant J. Cell Mol. Biol. 2011, 65, 194–205. [Google Scholar] [CrossRef]
- Lv, Q.; Li, X.; Jin, X.; Sun, Y.; Wu, Y.; Wang, W.; Huang, J. Rice OsPUB16 modulates the ‘SAPK9-OsMADS23-OsAOC’ pathway to reduce plant water-deficit tolerance by repressing ABA and JA biosynthesis. PLoS Genet. 2022, 18, e1010520. [Google Scholar] [CrossRef]
- Cong, J.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Physiol. Behav. 2013, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedeek, K.E.M.; Mahas, A.; Mahfouz, M. Plant genome engineering for targeted improvement of crop traits. Front. Plant Sci. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, M.; Mikami, M.; Toki, S. Biallelic Gene Targeting in Rice. Plant Physiol. 2016, 170, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kang, K.K.; Cho, Y.G. Molecular and Functional Analysis of U-box E3 Ubiquitin Ligase Gene Family in Rice (Oryza sativa). Int. J. Mol. Sci. 2021, 22, 12088. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Barbara, J.W.; Lior, P. Transcript as-sembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef] [Green Version]
- Waltz, E. Gene-edited CRISPR mushroom escapes US regulation. Nature 2016, 532, 293. [Google Scholar] [CrossRef] [Green Version]
- Kapahnke, M.; Banning, A.; Tikkanen, R. Random splicing of several exons caused by a single base change in the target exon of CRISPR/cas9 mediated gene knockout. Cells 2016, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Maquat, L.E. Nonsense-mediated mRNA decay: Splicing, translation and mRNP dynamics. Nat. Rev. Mol. Cell Biol. 2004, 5, 89–99. [Google Scholar] [CrossRef]
- Peccarelli, M.; Kebaara, B.W. Regulation of natural mRNAs by the nonsense-mediated mRNA decay pathway. Eukaryot. Cell 2014, 13, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.H.; Ryu, M.Y.; Jammes, F.; Hwang, J.H.; Turek, M.; Kang, B.G.; Kwak, J.M.; Kim, W.T. Roles of four Arabidopsis U-box E3 ubiquitin ligases in negative regulation of abscisic acid-mediated drought stress responses. Plant Physiol. 2012, 160, 556–568. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhao, J.; Li, L.; Gao, Y.; Zhao, L.; Patil, S.B.; Fang, J.; Zhang, W.; Yang, Y.; Li, M.; et al. The Arabidopsis U-box E3 ubiquitin ligase PUB30 negatively regulates salt tolerance by facilitating BRI1 kinase inhibitor 1 (BKI1) degradation. Plant Cell Environ. 2017, 40, 2831–2843. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Pandey, G.K. Expansion and function of repeat domain proteins during stress and development in plants. Front. Plant Sci. 2016, 6, 1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Singh, A.; Shankar, A.; Pandey, A.; Baranwal, V.; Kapoor, S.; Tyagi, A.K.; Pandey, G.K. Comprehensive expression analysis of rice Armadillo gene family during abiotic stress and development. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2014, 21, 267–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehti-Shiu, M.; Zou, C.; Hanada, K.; Shiu, S.H. Evolutionary history and stress regulation of plant receptor-like kinase/pelle genes. Plant Physiol. 2009, 150, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Ramegowda, V.; Krishnan, A.; Pereira, A. Rice GROWTH UNDER DROUGHT KINASE is required for drought tolerance and grain yield under normal and drought stress conditions. Plant Physiol. 2014, 166, 1634–1645. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.T.; Guo, P.; Xia, X.L.; Yin, W.L. PdERECTA, a leucine-rich repeat receptor-like kinase of poplar, confers enhanced water use efficiency in Arabidopsis. Planta 2011, 234, 229–241. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Bae, S.; Kim, J. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Abdula, S.E.; Jee, M.G.; Jang, D.W.; Cho, Y.G. High-efciency and Rapid Agrobacterium-mediated genetic trans-formation method using germinating rice seeds. J. Plant Biotechnol. 2011, 38, 251–257. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Won, K. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lim, K.; Kim, J.; Bae, S. Cas-analyzer: An online tool for assessing genome editing results using NGS data. Bioinformatics 2017, 33, 286–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Rice Research Institute (IRRI). Standard Evaluation System for Rice; IRRI: Los Banos, Philippines, 2002. [Google Scholar]
- IRRI. Standard Evaluation System for Rice 4th Edition; IRRI, INGER Genetic Resources Center: Manila, Philippines, 1996. [Google Scholar]
- Abdula, S.E.; Lee, H.; Ryu, H.; Kang, K.; Nou, I.S.; Sorrells, M.E.; Cho, Y. Overexpression of BrCIPK1 Gene En-hances Abiotic Stress Tolerance by Increasing Proline Biosynthesis in Rice. Plant Mol. Biol. Report. 2015, 34, 501–511. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef] [Green Version]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | No. of Plants Examined | No. of Plants with Mutations | Mutation Rate (%) | Putative Homozygous Mutations | Putative Bi-Allelic Mutations | Putative Heterozygous Mutations | |||
|---|---|---|---|---|---|---|---|---|---|
| No. of Plants | % | No. of Plants | % | No. of Plants | % | ||||
| OsPUB7 | 31 | 15 | 48 | 4 | 26.7 | 6 | 40.0 | 5 | 33.3 |
| Gene | Line | Genotype | No. of Resistant Plants | No. of Susceptible Plants | No. of All Edited Plants | χ2 | Significance (α) |
|---|---|---|---|---|---|---|---|
| OsPUB7 | K343-7 | Bi-allelic | 267 | 54 | 321 | 11.449 | 0.003 |
| K343-8 | Bi-allelic | 130 | 42 | 172 | 0.031 | 0.07 | |
| K343-17 | Bi-allelic | 23 | 16 | 39 | 5.342 | 0.001 |
| Gene | No. of Line | Plant Height (cm) | Culm Length (cm) | Panicle Length (cm) | No. of Tiller |
|---|---|---|---|---|---|
| Dongjin | WT | 113.3 ± 1.3 | 94.5 ± 1.6 | 16.6 ± 0.9 | 12.4 ± 1.7 |
| OsPUB7-GE | K343-7-5 ns | 115.1 ± 1.7 * | 89.7 ± 0.6 * | 21.6 ± 0.6 * | 9.7 ± 0.5 ns |
| K343-8-1 ns | 111.9 ± 2.3 ns | 92.6 ± 2.7 ns | 19.0 ± 1.7 ns | 15.7 ± 1.0 ns | |
| K343-17-8 * | 118.5 ± 1.6 ns | 94.3 ± 1.2 ns | 18.5 ± 0.9 ns | 15.0 ± 0.8 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-S.; Ko, S.-R.; Jung, Y.J.; Kang, K.-K.; Lee, Y.-J.; Cho, Y.-G. Knockout Mutants of OsPUB7 Generated Using CRISPR/Cas9 Revealed Abiotic Stress Tolerance in Rice. Int. J. Mol. Sci. 2023, 24, 5338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065338
Kim M-S, Ko S-R, Jung YJ, Kang K-K, Lee Y-J, Cho Y-G. Knockout Mutants of OsPUB7 Generated Using CRISPR/Cas9 Revealed Abiotic Stress Tolerance in Rice. International Journal of Molecular Sciences. 2023; 24(6):5338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065338
Chicago/Turabian StyleKim, Me-Sun, Seo-Rin Ko, Yu Jin Jung, Kwon-Kyoo Kang, Yung-Jo Lee, and Yong-Gu Cho. 2023. "Knockout Mutants of OsPUB7 Generated Using CRISPR/Cas9 Revealed Abiotic Stress Tolerance in Rice" International Journal of Molecular Sciences 24, no. 6: 5338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065338