
Temporins: Multifunctional Peptides from Frog Skin

1
Istituto di Scienze e Tecnologie Chimiche “G. Natta”, CNR, Via M. Bianco 9, 20131 Milano, Italy
2
Dipartimento di Scienze Farmaceutiche, Università degli Studi di Milano, Via Venezian 21, 20133 Milano, Italy
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(6), 5426; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065426
Submission received: 9 February 2023
/
Revised: 3 March 2023
/
Accepted: 8 March 2023
/
Published: 12 March 2023
(This article belongs to the Special Issue Antimicrobial Peptides: Structure and Mechanism of Biological Activity 2.0)
Abstract
:Temporins are short peptides secreted by frogs from all over the world. They exert antimicrobial activity, mainly against Gram-positive bacteria, including resistant pathogens; recent studies highlight other possible applications of these peptides as anticancer or antiviral agents. This review is meant to describe the main features of temporins produced by different ranid genera. Due to the abundance of published papers, we focus on the most widely investigated peptides. We report studies on their mechanism of action and three-dimensional structure in model systems mimicking bacterial membranes or in the presence of cells. The design and the antimicrobial activity of peptide analogues is also described, with the aim of highlighting elements that are crucial to improve the bioactivity of peptides while reducing their toxicity. Finally, a short section is dedicated to the studies aimed at applying these peptides as drugs, to produce new antimicrobial materials or in other technological uses.
1. Introduction
Frogs’ skin is a rich source of bioactive compounds, such as alkaloids, biogenic amines and peptides such as neuropeptides and antimicrobial peptides (AMPs) secreted by the cutaneous exocrine apparatus [1,2]. The production of AMPs is not common to all frog families; among the Neobatrachia, AMPs have been identified in the Cicroglossidae, Hylidae, Hyperoliidae, Leptodactylidae, Myobatrachidae and Ranidae families, while they have not been found in Bufonidae, Ceratophyridae, Eleutherodactylidae, Microhylidae, Pyxicephalidae or Rhacophoridae [3,4]. Each frog produces its own set of peptides that may act in synergy [5]. Typically, 10 to 20 peptides belonging to different families can be found in frogs’ secretions; the composition and structure of the peptides are different within each family, including linear and cyclic peptides, disulphide bridged peptides or sequences without cysteines [6]. The diversity of secreted peptides was hypothesized to be a strategy to gain maximum protection from different microorganisms and reduce the chance of inducing resistance to individual peptides [7]. The pattern of secreted peptides can be employed as a biomarker of a particular frog population [8]. In some species, AMP production is regulated by the thyroid hormones, and may be seasonal and affected by environmental factors [9,10]. The synthesis of peptides occurs in the multinucleated cells of the dermatous skin glands. Peptides are stored in the secretory granules and released by a holocrine mechanism [7]. The physiological role of secreted AMPs is still debated, although the majority of published studies are focused on their protective function against pathogens. Studies on the activity of these peptides against pathogens present in the animal environment are scarce compared to those on pathogens involved in human diseases [4,5]. The activity of frogs’ secreted peptides was demonstrated against A. hydrophilia, a bacterial strain present in frog skin, for combinations of temporin A and temporin L, temporin B and temporin L, bombinin-like peptide and brevinin-2-like peptide. In addition, temporin A, esculentin-2P, dermaseptin-B1 and ranauterin-2P were found to be active against frog viruses.
The observation that AMPs are co-secreted with neuropeptides and exert a cytolytic action suggests that the so-called AMPs support the neuropeptides’ action against predators by enhancing their delivery to the endocrine and nervous systems of the predators [1].
The sequences of these peptides are different in different amphibians, and the abundance of amino acids seems to be related to the geographical location of the amphibians. While alanine is abundant in peptides from South American frogs, leucine is more abundant in peptides secreted from European frogs [11].
A phylogenetic analysis of genes encoding skin AMP peptides from Hylidae and Ranidae shows that the genes evolved from a common locus. Peptides such as caerins from Australian hylids, brevinins, ranalexin, temporins from ranin and dermaseptins from South American hylids are derived from a common precursor sequence belonging to the preprodermaseptins family. Molecular cloning from cDNA libraries obtained from skin secretions reveals that AMPs are derived from tripartite precursors containing a signal peptide, a spacer, also named the acidic propiece, and the active peptide. The signal sequence is highly conserved between related species, whereas the spacer bioactive peptide differs between anaurans species. The diversity of active peptide sequences, likely being the result of adaptation to challenges by pathogens, is such that even phylogenetically related species produce non-identical peptide sequences [12,13].
Based on the sequence, peptides are divided into 14 families: Brevinin-1, Brevinin-2, Esculentin-1, Esculentin-2, Japonicin-1, Japonicin-2, Nigrocin-2, Palustrin-1, Palustrin-2, Ranacyclin, Ranalexin, Ranateurin-1, Ranateurin-2 and Temporin.
In this review, we will focus on the temporin family. Temporins are short peptides (8–17 amino acids long), showing broad spectrum activity against several microorganisms harmful for human health. The ease with which they are obtained and modified by chemical synthesis renders these peptides ideal starting materials to produce new antimicrobial agents.
Temporins were isolated from skin secretions of frogs such as Rana erythraea and Rana esculenta for the first time [14]. In 1996, after screening a cDNA library of the skin of Rana temporaria, precursors of three temporin peptides (temporin B, G and H) were identified. The precursors start with a signal peptide, and the mature peptides are preceded by the Lys-Arg sequence, a processing site for convertase, and terminate with the Gly-Lys sequence, which is processed by carboxypeptidases to produce amidated C-terminal peptides. The predicted mature sequences corresponded to two 13-amino-acid peptides and one decapeptide. The three peptides, together with other similar peptides (temporin A, temporin C, temporin D, temporin E, temporin F, L and K) were isolated from skin secretions of stimulated R. temporaria frogs. Isolated peptides were found to be all amidated at the C-terminal end; biological assays performed on synthetic peptides revealed that all peptides, apart from temporin D and H, show antibacterial activity. Since then, more than 150 peptides belonging to the temporin family have been isolated from ranid frogs of different genera such as Amolops, Hylarana, Lithobathes, Odorrana, Pelophylax, Rana and Hylarana [15]. The names of peptides follow the frog species names, as reported in the literature [16]. The sampling of peptides typically occurred upon mild transdermal electrical stimulation of live frogs, which causes contraction of the adrenergic myocytes with consequent release of their content. Samples were fractionated through HPLC, tested and identified through mass spectrometry (MS) techniques. Table 1 reports temporin sequences grouped by frog genera. Temporins share the following common features: the length, most of them are 13 amino acids long; the presence of at least one basic residue (lysine or arginine); the lack of cysteine. Some peptides contain non-proteinogenic amino acids, for example, temporin M from the Russian brown frog Rana temporaria has an oxyproline residue in the place of the proline found in Temporin M isolated from European Rana temporaria. The length of peptides is variable, ranging from 8 amino acids in temporin SHf from Pelophylax saharicus to 17 amino acids in temporin LTc from Hylarana latouchi [13].
Due to the large amount of data present in the literature on temporins, we cite examples of temporins produced from the different frog genera, and we discuss in more detail peptides for which more data have been reported. Particular attention was paid to the most recent studies describing structural details along with mechanism of action studies.
2. Temporins from Rana
Peptides isolated from frogs belonging to the Rana genus are the most widely investigated. Here, we will discuss peptides isolated from the Chinese brown frog Rana chensinensis and from the European frog Rana temporaria. We will describe the main features of temporins 1CE, A, B, G and L and their synthetic analogues. These peptides are all active against Gram-positive bacteria, do not have a defined secondary or 3D structure in buffer and fold in the presence of bacterial membranes or bacterial membrane mimics.
2.1. Temporin 1CE Family
Eight temporins were isolated from a skin cDNA library of the Chinese brown frog Rana chensinensis [18,19] (see Table 1). Studies on the bioactivity are reported on only five peptides (Table 2).
2.1.1. Temporin 1 CEa
Temporin 1CEa is a 15-amino-acid peptide, the most active among the temporin 1CEs, showing higher antimicrobial activity against Gram-positive than Gram-negative bacteria (Table 2) and significant haemolytic activity (lethal dose 50 (LD50) = 99 μM) [18,20,21]. Temporin 1CEa also shows anti-inflammatory activity; it binds to lipopolysaccharide (LPS), as revealed through isothermal titration calorimetry (ITC) experiments, mainly through electrostatic attraction inhibiting the inflammatory effects of LPS [20]. The dissociation constant value of temporin 1CEa for LPS is around 0.1 µM. Temporin 1CEa downregulates the MyD88-dependent signalling pathway, reducing the release of proinflammatory molecules as tumour necrosis factor-α (TNFα) and interleukin-6. The peptide has an interesting anti-cancer (antiproliferative) activity, with IC50 ranging from 30 to 60 μM against several cell lines [21]. It is particularly active against human breast cancer cells. Transmission electron microscopy (TEM) analysis revealed that in one hour, the peptide causes membrane breakage with leakage of intracellular components. In contrast, no damage to erythrocytes was observed at 50 μM and no cytotoxicity was observed against normal human endothelial cells. Studies in human A375 melanoma cells showed that Temporin 1CEa targets phosphatidylserine (PtdSer) overexpressed on the cancer cell surface. Temporin 1CEa binds PtdSer, as assessed through ITC, and this interaction elicits peptide helical structure favouring the membrane binding and inhibitory activity [22].
The molecular mechanism at the base of the anticancer activity of temporin 1CEa was analysed in different human breast cancer cell lines [23]. The peptide binds to cell membranes, disrupting membrane integrity, inducing permeability and causing the release of intracellular Ca2+. Scanning electron microscopy (SEM) and TEM studies showed that peptide at high concentrations disrupts cell membranes, inducing depolarization. In this condition, the peptide is internalized and targets the mitochondrial membrane (which is negatively charged), triggering mitochondrial membrane depolarization and reactive oxygen species (ROS) overproduction. Temporin 1CEa is unordered in water and assumes a helix structure in 50% trifluoroethanol (TFE)/water mixture [21].
2.1.2. Temporins 1-CEb, 1-CEc, 1-CEh
Temporin 1CEb is a 12-mer peptide with an overall 1+ charge (also known as amurin-3 from Rana amurensis). This peptide is active against Gram-positive bacteria and less active against Gram-negative bacteria, and it presents significant haemolytic activity (LD50: 112 μM) [18]. It is unordered in water, but it assumes a helical structure in the presence of 50% TFE or 30 mM sodium dodecyl sulphate (SDS), suggesting that this conformation could be assumed when the peptide interacts with bacterial membranes. Temporin 1CEb was the starting point to develop more potent analogues, as described below.
Temporin 1CEc has low antibacterial activity and also low haemolytic activity (HC50 ≥ 500 μM). For this reason, it inspired the design of new analogues (see infra).
Very recently, a new Temporin 1CE family member was identified and characterized: Temporin 1CEh [24]. The peptide shows inhibitory activity, mainly against Gram-positive bacteria, but it is endowed with an elevated haemolytic activity (HC50 = 152.6 μM).

{kind=link}
{kind=link}
{kind=link}
Table 2.
Antimicrobial activity against selected microorganisms of peptides from Rana chensinensis.
| Peptide | Antimicrobial Activity as MIC (µM) | |||
|---|---|---|---|---|
| S. aureus 22401 | Ref | E. coli 44102 | Ref | |
| Temporin 1CEa | 14.4 | [18] | >100 | [19] |
| Temporin 1CEb | 41 | [18] | >100 | [18] |
| Temporin 1CEc | >100 | [18] | >100 | [18] |
| Temporin 1CEe | 6.250 | [19] | 50.0 | [19] |
| Temporin 1CEh | 8 | [24] | 128 | [24] |
MIC = minimum inhibitory concentration.
2.1.3. Temporin 1 CEa Analogues
Temporin 1CEa analogues were designed with the aim of improving anti-cancer activity against human breast cancer cells and specificity while reducing haemolysis [25]. Hydrophobicity and cationicity were modulated by introducing Lys and Leu residues on the polar and apolar surface of the peptide, respectively. All designed peptides are unordered in water but fold in a helical conformation in the presence of 50% TFE or SDS. Structure–activity studies revealed that increasing the peptide positive charge enhances cancer cell specificity, whereas moderate hydrophobicity is required for reducing the haemolytic activity. Six analogues were tested, and the peptide LK3 (FKKLKKIANIINSIFKK-NH2), with lysine residues in position 2, 3 and 16 (in place of valine, aspartic acid and glycine), emerged as the most interesting, being more active than the parent peptide with an inhibitory concentration (IC)50 of 27.72 μM (MCF-7 cell line) and 25.04 μM (Bcap-37 cell line) and lower haemolytic activity, with an LC50 of 4.468 µM vs. the 95.74 µM of temporin 1CEa.
2.1.4. Temporin 1CEb Analogues
Temporin 1CEb analogues were designed to obtain a novel compound with improved bioactivity against pathogens and reduced haemolytic activity [26]. The primary amino acid sequence was modified in order to increase the overall positive charge (introducing Lys) and decrease hydrophobicity to modulate haemolysis. In particular, the analogue L-K6 (KKILSKIKKLLK-NH2) presenting a 7+ charge was very promising. It was more active than the parent peptide against several Gram-negative and Gram-positive bacteria (minimum inhibitory concentration (MIC) = 3.25 μM) and almost not haemolytic (LD50 > 1000 μM). The L-K6 peptide assumes a helical conformation in membrane-like environments such as in the presence of SDS (30 mM) or TFE as a cosolvent (50%). The peptide enhanced the permeability and induced the depolarization of the bacterial membrane. This peptide showed promising application in the treatment of dental caries, being able to kill oral pathogens such as Streptococcus mutans (MIC: 3.13 μM) as well as Candida albicans (MIC: 6.25 μM) [27]. More interestingly, L-K6 was also able to inhibit biofilm formation and reduce the biomass of an existing biofilm of S. mutans; furthermore, it could decrease the inflammatory response mediated by LPS, modulating the MyD88-dependent pathway and inhibiting the expression of pro-inflammatory molecules [28].
The role played by single or multiple tryptophan residues introduced in place of the apolar amino acids of the L-K6 peptide was also evaluated [29]. Analogues with a single-point mutation (I1W and I4W) were endowed with improved antibacterial activity, especially against Gram-negative bacteria, such as Klebsiella pneumoniae, Enterobacter cloacae and Proteus mirabilis; in contrast, the peptide L-K6 is inactive. The presence of tryptophan does not affect the haemolytic activity of this class of peptides. The introduction of a tryptophan residue in the N-terminal region increases the affinity for negatively charged lipids, with the side chain of the tryptophan inserting into the bilayer as demonstrated by fluorescence and quenching studies. The penetration of the tryptophan side chain into the membrane improves the ability of the peptide analogues to break the negatively charged liposomes; this could explain the ability of these tryptophan analogues to kill bacteria. In addition, the replacement of leucine in position 5 with tryptophan (L5W) mainly improves the antibacterial activity against Gram-positive bacteria. Finally, the introduction of two tryptophan residues in the analogue L-K6 afforded peptides I1WL5W and I4WL5W, featuring strong antimicrobial activity against both Gram-positive and Gram-negative bacteria and low cytotoxicity [30]. The mechanism of action of these peptides consists of binding to the bacterial membrane, as suggested by fluorescence studies, mainly due to electrostatic interactions, followed by the insertion into the lipid phase of the aromatic side chains, which triggers membrane destruction. These results were further confirmed through SEM analysis on Escherichia coli and Staphylococcus aureus bacteria.
The peptide L-K6 exhibits anticancer activity, especially against MCF-7 cancer cells [31]. The mechanism of action consists of a selective binding to the cancer cell surface, determined by the presence of PtdSer, and internalization mainly via a clathrin-independent macropinocytosis, causing nuclear damage without affecting the cytoskeleton, as revealed using super-resolution microscopy. Inside the cancer cell, L-K6 likely targets the DNA, inducing chromatin decondensation and DNA double strand breaks. In a subsequent in vitro study, it was shown that L-K6 induces sensitivity to doxorubicin in resistant breast cancer cells MCF-7/ADR, suggesting a possible combined use of L-K6 and chemotherapeutic agents to overcome drug resistance in cancer chemotherapy [32].
Another interesting analogue of temporin 1CEb is the peptide D-K5 described by Shang and co-workers, derived from L-K6, with a D-Lys in position 7 [26]. It was selected to prepare a bifunctional molecule presenting the antibacterial peptide D-K5 linked through a PEG spacer (8-amino-3,6-dioxaoctanoic acid) to a Leu-enkephalin analogue (dalargin, DAL), a peptide with cytoprotective and pro-proliferative activity for application in infected wound healing [33]. Conjugation affects the antimicrobial properties of the original peptide to a small extent; however, the bifunctional peptide shows significant cytotoxicity towards keratinocytes and fibroblasts when used at higher concentrations. Successively, the immunomodulatory properties of the conjugate DAL-PEG-DK5 were studied [34]. In vitro analyses demonstrated that the DAL-PEG-DK5 peptide shows anti-inflammatory activity against LPS-induced inflammation. The peptide interacts with LPS, and this interaction drives peptide aggregation, which hampers LPS recognition by the host cell, thereby inhibiting intracellular signalling and decreasing the expression of pro-inflammatory cytokines. In vivo experiments showed that the DAL-PEG-DK5 peptide protects mice from septic shock mediated by LPS. Finally, it was shown that this peptide exerts antimicrobial activity against methicillin-resistant Staphylococcus aureus (MRSA) [35]. In particular, DAL-PEG-DK5 showed good bacteriostatic activity against several MRSA strains isolated from infected patients, with an MIC of 40 μg/mL against the S. aureus USA300 strain. Furthermore, the peptide was also able to disrupt the biofilm of the USA300 strain. The mechanism of action consists of bacterial membrane permeabilization and disruption. The peptide is able to accumulate in the cytoplasm of infected human keratinocytes, leading to a significant reduction (71%) in intracellular MRSA, suggesting a potential application for the treatment of staphylococcal skin infections.
2.1.5. Temporin 1CEc Analogues
Temporin 1CEc analogues were designed to improve natural peptide antimicrobial activity, modulating the total positive charge and hydrophobicity of the peptides [36]. Structure–activity relationship studies showed that for this peptide series, a balance between hydrophobicity and cationicity is required to obtain improved biological activity. Peptide analogues 2K2L (IIPLPLKKFLKKL-NH2) and 2K4L (IILLLLKKFLKKL-NH2) have broad spectrum antibacterial activity with low or moderate haemolytic effect. Interestingly, they are also active against multi-drug resistant bacteria such as Staphylococcus epidermidis (MRSE1208). In this case, they inhibit biofilm formation. The peptides achieve antibacterial activity, disrupting the bacterial cell membrane as revealed through SEM and confocal laser scanning microscopy experiments on MRSE1208. The analysis of membrane permeability in the presence of the two peptides suggested the formation of pores. Finally, an analogue of temporin 1CEh named T1CEh-KKPWW (KKWVPWKKIANIL-NH2) was reported. It presents improved antibacterial activity against Gram-negative bacteria and low haemolysis [24].
2.2. Temporin A
2.2.1. Temporin A Biological Activity
The temporin A peptide was isolated from Rana temporaria in 1996 [37]. The biological activity of this peptide is quite broad. In fact, temporin A is active against bacteria, fungi and protozoa and, in addition, it has been shown to present anticancer and insulinotropic activity (Table 3). Temporin A is mainly active against Gram-positive bacteria, such as S. aureus and Bacillus megateriurn, specific E. coli strains and clinically important strains such as methicillin-resistant S. aureus and vancomycin-resistant Enterococcus faecium and Enterococcus faecalis. The measured MIC values against these bacteria range from 2.5 to 20 µM [38]. More importantly, temporin A presents low lytic activity against human red blood cells (LC > 120 μM).
Several studies have been performed on MRSA strains, specifically to treat superficial wounds [39]. In this regard, it has been shown that temporin A, in addition to its antibacterial properties, can stimulate the healing of tissues. In fact, it reduces the number of S. aureus cells inside HaCaT keratinocytes, but at the same time induces the migration of HaCaT keratinocytes mediated by EGFR [40]. In vivo experiments show that temporin A is effective in the treatment of MRSA-infected surgical wounds in mice; when topically administered, it promotes healing [41]. Temporin A was investigated as a prophylactic agent in infections from Staphylococci [42]. Finally, temporin A inhibits biofilm formation on polystyrene surface and central venous catheters, also in synergy with a cecropin A-melittin hybrid peptide (CA(1–7)M(2–9)NH2) [43].
In addition, temporin A activity has been studied in combination with other antibiotics compounds, very often showing remarkable synergism. For example, the peptide showed synergy with imipenem (against Pseudomonas aeruginosa), ceftazidime (against P. aeruginosa, S. aureus), polymyxin E (against P. aeruginosa) and with linezolid against S. epidermidis ATCC 12228 in a subcutaneous rat pouch model [44,45]. Moreover, it is active against E. faecalis, also in combination with co-amoxiclav and imipenem [44]. It improves survival in a mouse model of staphylococcal sepsis when administered in combination with imipenem [46]. Temporin A enhances the antibacterial activity of gentamicin against S. aureus and, to a lower extent, against P. aeruginosa biofilm, suggesting application as adjuvant agent in the treatment of biofilm-associated infections [47]. Temporin A can also act in combination with other temporin peptides, such as a temporin B analogue or temporin L, against different Gram-positive and Gram-negative bacteria [48,49]. Notably, synergism with a modified temporin B (TB-YK) has been reported. In particular, the peptide mixture showed antimicrobial and anti-inflammatory activity in vivo against Gram-positive (S. aureus, Listeria monocytogenes) and Gram-negative (Salmonella enterica, E. coli) bacteria [49]. Temporin A is active against fungi such as C. albicans and channel catfish virus and the non-enveloped frog virus-3 [50]. A computational study revealed that the peptide could interact with Middle East respiratory syndrome coronavirus (MERS-CoV) protein and, hence, it could have a role in the treatment of this pathology [51]. Temporin A has strong antiparasitic action against protozoa of the Leishmania genus, triggering membrane destruction. For this reason, it should prevent the pathogen from developing resistance [52,53]. Furthermore, it is a chemoattractant for phagocytic leukocytes through its interaction with the Formyl Peptide Receptor-Like 1 receptor, suggesting a role in host immune response to infection [54]. Finally, temporin A showed interesting activity against human cells. It was demonstrated that it presents anticancer activity towards the non-small cancer lung cell (NSCLC) lines, and was able to induce insulin release from rat- and human-derived beta cells with potential use for type 2 diabetes treatment [55,56]. The interaction between temporin A and A549 cells, lung-derived cells, was studied through scanning ion conductive microscopy, revealing inhomogeneous binding of temporin A at the cell surface [57].
2.2.2. Temporin A Structure and Mechanism of Action
The mechanism of action of temporin A is based on the molecular interaction with the bacterial cell membrane. The peptide in water is unordered, and it assumes a helical conformation when dissolved with a cosolvent (such as TFE) or a detergent (SDS), suggesting that this will be the active conformation in the presence of bacteria. The positively charged peptide is attracted by the negative charges exposed on the bacterial membrane, and the change from water to a more hydrophobic environment induces helix stabilization in the amphipathic peptide. Structural studies in SDS micelles showed that the peptide is located at the micelle–water interface; in contrast, in the presence of dodecylphosphocholine (DPC) micelles, temporin A does not fold and is not able to penetrate the hydrophobic core, suggesting a molecular explanation for its low haemolytic activity. Increasing the concentration of the peptide on the bacterial membrane, the peptide starts to assemble and destroys the membrane through a carpet-like mechanism [53,58]. In the presence of LPS micelles, temporin A assumes an oligomeric helical conformation; this could explain the limited ability of the peptide to permeate the membranes of Gram-negative bacteria, explaining its low antibacterial activity [59]. Temporin L prevents the LPS-induced self-association of temporin A; this observation suggests an explanation of the synergism and activity of the peptide mixture against Gram-negative bacteria [60].
2.2.3. Temporin A Analogues
Several analogues have been designed to understand structure–function relationships and to develop a more potent antimicrobial agent. Single or double mutants, dimeric, peptides with inverted chirality (all-D) and retro-inverse analogues have been studied, but no significant advantages with respect to the natural peptide have been found [15].
2.3. Temporin B
2.3.1. Temporin B Biological Activity
Temporin B is a 13-amino-acid peptide isolated from the skin of the European frog Rana temporaria. It is mainly active against pathogens such as Gram-positive bacteria, including S. aureus, B. megaterium and Streptococcus pyogenes, and also against fungi such as C. albicans (Table 4). Experiments carried out with bacteria isolated from the natural flora of the frogs reveal that temporin B is strongly active against Acinetobacter junii Bo-2 [61]. Temporin B effectively kills ATCC-derived and multidrug-resistant clinical isolates of S. aureus in HaCaT keratinocytes without injuring host cells. Interestingly, the peptide exerted a wound-healing effect when tested on keratinocyte monolayers [40]. At very high concentrations, it shows activity against E. coli; the activity increases when tested against E. coli strain mutants with defective cell walls. Antibacterial assays against an MDR clinical isolate of P. aeruginosa show a lack of activity in vitro, while in experiments on Caenohabditis elegans infected with the same bacterium, the peptide was found to be able to reduce the lethality of infected nematodes [62].
The peptide acts in synergy with temporin L against Gram-negative bacteria such as E. coli, P. aeruginosa and Aeromonas hydrophyla. The synergic action assumes a physiological significance and explains why the same organism produces multiple forms of AMPs.
The biological activity of temporin B was also investigated against viruses, cancer cells and parasites. The peptide displays anti-Leishmania activity at a micromolar concentration; it was hypothesized that it permeates the plasma membrane of the parasite [46]. Recent experiments show that temporin B inhibits herpes simplex virus 1 (HSV-1) infection in human epithelial cells. The peptide disrupts the viral envelope, as suggested in TEM experiments. In addition, it inhibits the attachment and the entry of the virus into host cells [64]. The lack of cytotoxicity and haemolytic activity render this peptide interesting for future therapeutic applications.
2.3.2. Temporin B Structure and Mechanism of Action
The secondary structure of the peptide was investigated through CD spectroscopy in different conditions. In buffer at neutral pH, the peptide does not show conformational preferences; in the presence of 30% TFE or negatively charged micelles composed of SDS and 1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine (SOPC)/1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoglycerol (POPG) large unilamellar vesicles (LUV), the peptide assumes a helical conformation [54,61,65]. Interestingly, when the peptide is incubated with either E. coli LPS or E. coli cells, there is no indication of a preferred conformation [60,66]. NMR studies of temporin B in LPS micelles suggest oligomerization of the peptide, involving residues at the N- and C-terminal ends [67]. When temporin L and temporin B are combined, temporin B appears as a monomer. Based on these results and on fluorescence experiments, it was hypothesized that temporin L prevents the aggregation of temporin B within LPS, therefore allowing translocation of the peptide across the outer membrane to reach the cytoplasmic membrane [60,67]. The structure of temporin B in LPS micelles was determined employing data collected on the mixture of temporin B plus temporin L. The peptide assumes a helical conformation, exhibiting a large hydrophobic face comprising residues Leu1, Ile4, Val5, Leu8 and Leu12, and on the opposite side, the two polar residues (Asn7 and Ser11), along with Lys 10. Saturation transfer difference (STD) NMR experiments indicate that only N-terminal hydrophobic residues and polar residues establish interactions with LPS, supporting the hypothesis that the self-association of the peptide by non-polar residues might be responsible for the limited interactions of the peptide with LPS micelles [67].
Initial attempts to understand the mechanism of action of temporin B were performed by studying the interactions of the peptide with lipids as model biomembranes. The peptide causes lysis of zwitterionic or acidic phospholipids, as revealed by leakage experiments with phosphatidylcholine (PtdCho), phosphatidylserine (PtdSer) and phosphatidylglycerol (PtdGro) [61]. Studies on LUV composed of SOPC and SOPC/POPG by circular dichroism reveal that the peptide folds into an α-helix upon interaction with the vesicles. Using temporin B conjugated to the fluorescent probe Texas red through an extra cysteine residue at the N-terminus, the formation of amyloid-like fibres was observed in the presence of SOPC/POPG micelles but not phosphatidylcholine (PC) [65]. The insertion of the peptide into lipid monolayers located at air/water interfaces was investigated by measuring the change in surface pressure upon injection of the peptide into SOPC/POPG micelles [68]. Temporin B penetrates monolayers, and penetration increases with increasing POPG content. The interaction of temporin B with lipids causes lipid lateral segregation and the formation of lipid-enriched microdomains. Electrostatic interactions between the positively charged peptide and the negatively charged membranes reduce repulsion between peptides, resulting in their aggregation in the membrane. Experiments with giant unilamellar vesicles (GUV) containing SOPC and POPG reveal that the peptide rapidly aggregates on the surface of the membrane and does not cause GUV dissolution [69]. This observation supports the hypothesis that temporin B destabilizes the membrane and unlike a detergent, it does not destroy it, nor forms pores. Studies on glass-supported phospholipid bilayers revealed that the interactions of temporin B with these model membranes results in the segregation of membrane-bound peptides and formation of lipid fibrillar protrusions [70], suggesting that the interaction of peptides with lipids drives membrane shape transformations. Recent studies performed using all-atom and coarse-grained molecular dynamics simulations aimed at investigating the interaction of temporin B with mixed zwitterionic and anionic phospholipids membranes support previous experimental results, confirming the ability of temporin B to produce clusters on the membrane surface and promote the extrusion of lipids [71].
The interaction of temporin B with E. coli LPS was investigated through ITC; in physiological conditions, i.e., at 37 °C, when the LPS is in the liquid crystalline phase, the interaction of the peptide with LPS O26:B6 is an endothermic entropically driven process, likely involving the disruption of hydrogen bonds in ordered water molecules around lipid A [72]. The titration of LPS O111:B4, which has longer carbohydrate chains as compared to O26:B6, is instead an exothermic process in the same conditions, driven by electrostatic interactions. Depending on the prevalence of hydrophobic vs. electrostatic forces, the interaction of peptides with LPS will be enthalpically or entropically driven. The ability of temporin B to interact with LPS was also proven in fluorescence experiments. Studies performed on a rhodamine-labelled and also on a dansyl-labelled temporin B in the presence of E. coli LPS show that the peptide self-associates in these conditions [60]. Binding studies of temporin B to LPS are reported in different papers. Binding stoichiometry studies revealed that an excess of peptide binds to LPS micelles; the dissociation constants of nitrobenzodiazole (NBD) and dansyl-labelled temporin B were comparable and found to be in the micromolar range [67,73]. Binding studies of the peptide to E. coli cells revealed a lack of interactions, consistent with the lack of activity against E. coli. TEM studies carried out on E. coli cells incubated with temporin B revealed no changes in the bacterial cell morphology. Overall, these data indicate that the ability of temporin B to establish interactions with E. coli LPS is not necessarily related to its ability to kill bacterial cells [73].
2.3.3. Temporin B Analogues
Extensive work has been performed on the development of temporin B analogues. The introduction of positively charged amino acids results in a broader spectrum of action of the peptide. In 2009, Capparelli and co-workers reported an analogue modified by a tyrosine and two lysines at the N-terminus (TB-YK) [54]. This analogue was active against Gram-positive and Gram-negative bacteria. Experiments in vitro and in vivo demonstrated that this peptide acts in synergy with temporin A, in conditions that closely mimic bacterial infections in humans. Alanine scanning studies, aimed at understanding the amino acids crucial for the activity of temporin B, revealed that Lys in position 10 is fundamental for the activity of the peptide, and that substitution of the glycine residue in position 6 by an alanine resulted in a peptide with slightly improved activity against Gram-positive bacteria [63]. The introduction of two lysines at the N-terminus of the peptide in combination with the replacement of glycine by alanine resulted in a peptide (TB_KKG6A) also showing activity against the Gram-negative bacteria E. coli and P. aeruginosa [74].
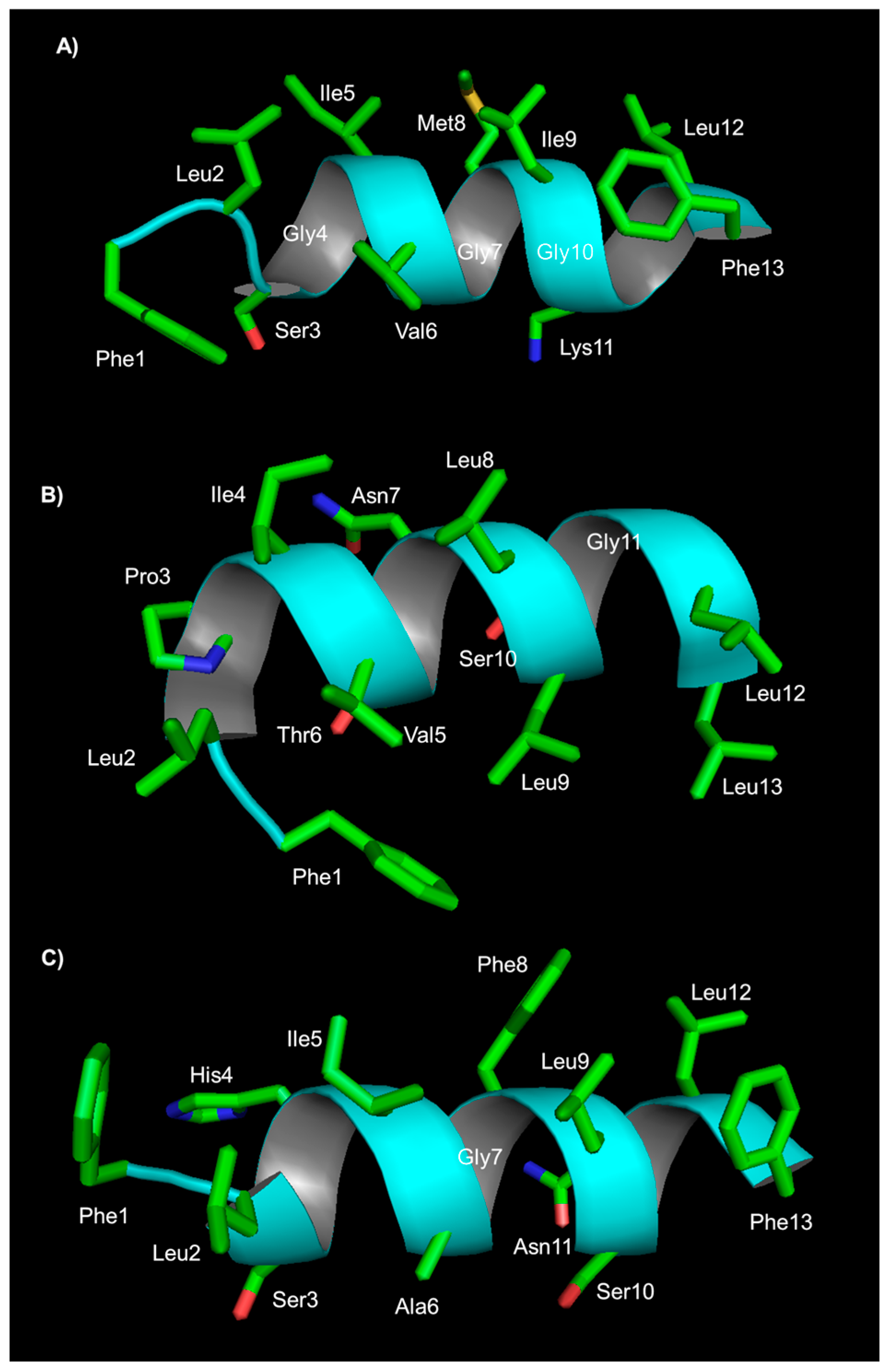
Structural studies on the peptide TB_KKG6A were carried out through NMR in the presence of E. coli LPS and E. coli cells; these studies provide high-resolution molecular details of the interaction between the peptide and bacteria. The peptide aggregates in solution and becomes monomeric when it establishes interactions with the bacterial outer membrane. A transition from a random coil to a helical structure is observed either with LPS or with cells of E. coli, but the three-dimensional structure in the two conditions is different: TB_KKG6A is a kinked helix when it interacts with isolated LPS, while it is an uninterrupted helix when it interacts with E. coli cells (Figure 1). In the latter case, the helix shows a hydrophobic face containing Val5, Leu8, Leu 9 and Leu 12, and a hydrophilic face comprising residues Asn7, Lys10 and Ser11. The N-terminal end with two lysines is mostly disordered. STD experiments revealed that amino acids on the hydrophilic face are in contact with the LPS. Altogether, structural studies suggest that positively charged residues at the N-terminus are responsible for anchoring the peptide to the negatively charged phospholipids, and Pro3, Ala6, Asn7 and Lys10 specifically bind to the membrane. Binding studies of the peptide in the presence of E. coli LPS and E. coli cells revealed that the peptide binds to LPS and to cells with a very high affinity; the stoichiometry of binding to cells is about 1 × 106 peptides per bacterial cell, suggesting that peptides cover the cell membrane. The analysis of the morphology of E. coli cells incubated with TB_KKG6A through TEM shows the formation of cellular filaments. One hypothesis is that membrane disgregulation causes the inhibition of cell division; the process of cell division is, in fact, mediated by the formation of a “Z ring”, composed of polymerized FtsZ protein, at the site of division. If the protein FtsZ is not associated with the membrane, Z rings are not functional for division, and long filaments of cells are observed [75].
Further studies on temporin B analogues in which the amino acids in position 3, 6 and 7 were modified allowed the identification of the analogue TB_KKG6K with improved activity toward Gram-positive and Gram-negative bacteria. TB_KKG6K inhibits the growth of human pathogenic yeast and kills planktonic and sessile cells of C. albicans [76,77]. The introduction of D-lysines in place of L-lysine in TB_KKG6K to increase the stability to proteolytic degradation did not result in a substantial change in the peptide antimicrobial activity. Interestingly, this peptide was better tolerated than its L-lysine counterpart in vitro, in primary human keratinocytes and in 3D human epidermal models. Studies aimed at understanding the fungal cell membrane activity of TB_KKG6K were carried out using model membrane systems. The peptide preferentially interacts with anionic membranes; fluorescence microscopy experiments carried out on a fluorescein-labelled peptide and with a lipophilic dye FM4-64 suggest that the peptide compromises the integrity of the membrane and is rapidly taken up by cells through an energy-independent mechanism. The peptide does not cause cell lysis but triggers the disintegration of subcellular structures. These results suggest the existence of an intracellular target of the peptide.
A different temporin B analogue named TB_L1FK was designed through a statistics-based computational strategy, aimed at obtaining peptides showing physico-chemical features such as hydrophobicity and net charge similar to the natural peptide temporin B, and low cytotoxicity. TB_L1FK was obtained by replacing the N-terminal leucine with a phenylalanine and the C-terminal leucine with a lysine, and deleting Asn7 [78]. The antimicrobial activity of this peptide was investigated together with that of TB_KKG6A. The two peptides reduced the ability of S. aureus to form biofilm, while only TB_KKG6A reduced the biomass of P. aeruginosa biofilms. Both peptides in combination with EDTA caused a reduction in colony-forming unit (CFU) number in preformed biofilms of S. aureus and P. aeruginosa. Interestingly, no haemolytic activity was detected on human red blood cells, and TB_L1FK showed a lower cytotoxic activity against human epithelial cells.
2.4. Temporin G
Temporin G is a 13-amino-acid peptide isolated from the skin of Rana Temporaria [37]. Its antimicrobial activity was initially investigated against nosocomial multidrug-resistant strains; it exerts bactericidal activity against Gram-positive bacteria such as E. faecium and S. aureus and Gram-negative bacteria as Stenotrophomonas maltophilia and Acinetobacter baumannii at low concentrations (Table 5) [79]. The activity of the peptides is strongly reduced or lost when tested in human serum.
Temporin G can eradicate S. aureus biofilms; fluorescence experiments performed using the membrane-impermeable dye Sytox green revealed that the peptide induces the perturbation of S. aureus biofilm membranes [81]. When used in combination with tobramycin, a commercially available antibiotic, the peptide at a sub-inhibitory concentration enhances the activity of the antibiotic. Tests on human immortalized keratinocytes show that the peptide is slightly cytotoxic only at high concentrations, causing a reduction in metabolically active cells. The activity of the peptide was also tested on fungal strains: MIC50 between 4 and 16 µM were found against C. albicans, while a reduced activity was detected against non-Candida species [80]. The peptide acts by destabilizing the cell membrane structure, likely causing the formation of small local breaks. It is also effective in inhibiting the yeast-to-hyphal transition, and hampers the formation of biofilms in Candida. Finally, the peptide is active against filamentous pathogenic fungi such as Microsporum gypseum or Trichophyton mentagrophytes. Recently, the antiviral activity of temporin G was explored [82]. The peptide inhibits the early stages of the PR8A influenza virus replication in A459 cells; it was hypothesized that the peptide interacts either with the subunit HA1 of the protein haemagglutinin and this leads to the inhibition of the attachment of the virus to the host cell, or with the HA2 subunit and terminal regions of HA1 to prevent the conformational change of HA essential for the endocytosis of the virus into the host cells. Finally, temporin G inhibits the replication of Herpes simplex virus type 1, affecting the early stages of the virus life cycle and virion as well. The peptide triggers a reduction in infection by John Cunningham polyomavirus (JCPyV) [83]. The peptide stimulates insulin release from 1.1B4 human-derived pancreatic β cells, without cytotoxicity [56]. Experiments in mice show that temporin G injection improves glucose tolerance and causes an increase in insulin secretion, suggesting its potential application in the treatment of type 2 diabetes.
No analogue has been reported so far to the best of our knowledge.
2.5. Temporin L
2.5.1. Temporin L Biological Activity
Temporin L is a 13-amino-acid peptide isolated from Rana Temporaria [37]. It is active against Gram-positive bacteria and the fungus Candida (Table 6). In contrast to most of the natural temporins, it also shows activity against Gram-negative bacteria. Temporin L at a subMIC concentration inhibits biofilm formation in Pseudomonas fluorescens in static and dynamic conditions and is able to perturb preformed biofilm architecture [84,85]. In order to investigate the antiendotoxic effect of temporin L, its interaction with LPS was investigated in vitro and in vivo. Fluorescence displacement assays revealed that the peptide binds to purified E. coli LPS and lipid A and penetrates into LPS monolayers. The antimicrobial activity of the peptide against E. coli is reduced in the presence of LPS, confirming the affinity of temporin L toward LPS. In vivo experiments on rat models indicate that treatment with a combination of temporin L and imipenem and piperacillin reduces the plasma levels of the endotoxin and TNF-alpha, resulting in high survival rates of the animals [86].
Temporin L causes lysis of human erythrocytes, with an efficiency higher than temporin A and B. It is also cytotoxic to cancer cell lines such as Hut-78, U-937 and K562 [87]. Cell sensitivity to the peptide is dependent on the cell line, but the kinetics of killing are very rapid in all cases. K-562 and U-937 cells were more resistant as compared to Hut-78.
The antiviral activity of temporin L was investigated against different viruses, including enveloped, naked, DNA and RNA viruses. The peptide is active against herpes viruses, paramyxoviruses, influenza virus and coronaviruses, including SARS-CoV-2, showing a reduced or null activity against nonenveloped viruses [85]. Based on these results, it was hypothesized that the activity of the peptide is related to its ability to interact with the viral membrane.
2.5.2. Temporin L Structure and Mechanisms of Action
To understand its mechanism of action against bacteria, membrane permeabilization studies were performed by detecting the release of fluorescent dyes from lipid vesicles of different compositions, charges and sizes, such as the electrically neutral PC or PC/cholesterol and the negatively charged PC/POPG. The peptide interacts preferentially with neutral zwitterionic vesicles and perturbs membrane integrity; leakage of large dyes occurs to a lesser extent compared to that of small molecules, suggesting the formation of local pores or breaks in the membrane rather than its disruption by a detergent-like mechanism. It was hypothesized that temporin L induces a profound reorganization of the bilayer, establishing contacts with the lipid chains. In vesicles containing acidic phospholipids, the formation of clusters with lipid-bound peptide molecules is observed [68,87]. Using fluorescence spectroscopy, exploiting the tryptophan present in temporin L, Zhao and co-workers investigated the interaction of the peptide with SOPC/cholesterol, SOPC and SOPC/POPG liposomes [88]. The interaction of temporin with liposomes determines a blue shift in the emission of the tryptophan and fluorescence quenching, suggesting that the peptide is partially inserted into the liposomes. The peptide binds to SOPC liposomes and interactions strengthen in the presence of POPG. The results of these experiments support the hypothesis that the peptide interacts with the lipid bilayer in a defined fashion and does not induce significant lipid segregation, suggesting a barrel–stave mode of interaction with the bacterial membrane. Experiments carried out in parallel on temporin B and L on supported phospholipid bilayers composed of PC and PG indicate the formation of lipid tubular protrusions, suggesting that peptide–lipid interactions play a key role in membrane shape transformation [70].
Using CD and NMR techniques, the interactions of temporin L with micelles of SDS and DPC, employed as mimic of bacterial and mammalian membranes, respectively, were investigated [58]. The peptide has a random coil structure in water and is an α-helix in SDS and dodecylphosphorylcholine (DPC) micelles. NMR in SDS reveals the existence of an α-helical structure encompassing residues 3 to 11, while the N and C-terminal ends appear less defined. In DPC, all residues are in an α-helix conformation. The topological orientation of the peptide in the micelles was detected using paramagnetic probes and NMR: in SDS, it is located at the liquid–water interface, while in DPC, it is perpendicular to the micelle surface with its N-terminus buried in the hydrophobic core of the micelle and the C-terminus in the water layer. These results are consistent with the experimentally observed tendency of temporin L to interact with mammalian membranes, such as those of erythrocytes. Fluorescence experiments confirmed the ability of temporin L to interact with E. coli LPS with a calculated dissociation constant of 4.4 µM, and suggest the formation of an antiparallel dimer [67]. NMR studies of temporin L in LPS micelles indicate that the peptide assumes an amphipathic α-helical structure, showing the hydrophobic and aromatic residues Trp4, Phe 5, Leu9, Phe8 and Ile12 on one face, and the hydrophilic residues Gln3, Ser6, Lys7 and Arg11 on the other face. Interactions between the residues of the non-polar face stabilize the dimer (Figure 2). CD studies carried out for the peptide in the presence of E. coli cells reveal that this peptide, as some analogues of temporin B, folds into a helix upon interaction with cells. CD spectra recorded on a mixture of temporin L with S. epidermidis cells suggest the folding of the peptide in a helical conformation, although signals are not very intense [89].
The ability of the peptide to permeate the cytoplasmic membrane of E. coli cells was investigated by monitoring the leakage of cytoplasmic β-galactosidase: the peptide increases the permeability of the inner membrane at concentrations in which it shows significant bactericidal activity [90]. Using a triple-staining assay to detect contemporarily total, viable and membrane-perturbed bacteria, Mangoni and co-workers found that a sublethal concentration of temporin L alters the integrity of the bacterial membrane to a significant extent without triggering cell death, and induces modifications to the cell shape, as shown through SEM experiments.
Using a combination of molecular modelling and electrophysiology experiments such as patch–clamp, the interaction of temporin L with model membranes was investigated [91]. The structure of the peptide in model membranes is helical. The peptide conformational flexibility was investigated through molecular dynamic simulations in 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine (POPE)/POPG and POPG; the N-terminus of the peptide appears to be endowed with high flexibility and is involved in the penetration of the membrane. The association of the peptide through aromatic hydrophobic residues, namely Phe1, Phe5 and Phe 8, occurs. Patch–clamp experiments in 1,2-diphytanoyl-sn-glycero-3-phospho-(1′-rac-glycerol)(DPhPG) membranes, used as a mimic of Gram-negative bacteria cytoplasmic membrane, suggest that temporin L has channel-like activity, forming pores with a size twice the diameter of a chloride ion.
A deeper investigation of the mechanism of action of temporin L on Gram-negative bacteria was carried out using a combination of functional proteomic, fluorescence, docking, TEM, DLS and small-angle neutron scattering (SANS) to visualize the effect of the peptide on E. coli cells [92]. Initial studies allowed the identification of proteins belonging to the divisome as interactors of temporin L. The analysis of the morphology of bacteria incubated with the peptide revealed the formation of elongated structures, typically formed when cell division is impaired. Molecular docking, fluorescence experiments and biochemical assays supported the hypothesis that the protein FtsZ, responsible for the first step of bacterial cell division, is the target of temporin L. SANS analyses, aimed at investigating the effect of the peptide on E. coli cells focusing on structures formed in the range 2 to 300 nm, indicate that there is no difference in the lamellar structure of E. coli upon treatment with the peptide, which means no breaks in the membrane, and suggest a change in the spatial arrangement of the proteins targeted by the peptide. Overall, the mechanism of action appears to be the following: the peptide crosses the outer membrane and binds to FtsZ, triggering the inhibition of cell division and cell death [85,93,94].
2.5.3. Temporin L Analogues
Many temporin L analogues were synthesized with the aim of reducing its cytotoxicity and, of course, improve its antimicrobial activity. Selected analogues will be described. Several studies started from the analysis of the structure of the peptide and investigated the relationship between the structure and the biological activity of the peptide. In 2008, in a study carried out in parallel on temporin A and temporin L, Carotenuto and co-workers hypothesized that the ability of temporin L to interact with mammalian membranes, therefore being cytotoxic, was related to its regular helical structure. With the aim of inducing a turn in the structure of the peptide, Gln3 was replaced by a proline; the resulting peptide Pro3TL showed increased activity against Gram-positive bacteria and yeast cells, and also reduced haemolytic activity with respect to the parent peptide [58]. The analysis of the structure of this peptide in SDS and DPC revealed a stable helical conformation between residues 6 and 13 and a flexible N-terminus, supporting the initial working hypothesis. The substitution of L-Leu9 by the D enantiomer yields a peptide devoid of haemolytic activity, showing improved activity against C. albicans and reduced activity against bacteria as compared to Pro3TL [95]. The structure of this peptide in SDS and DPC is a kinked helix. The structure is stabilized by an unusual i to i + 5 hydrogen bond between Phe5 and Gly10. This peptide shows anti-inflammatory activity in zymosan-induced peritonitis in in vivo models of mice [96]. In addition, the peptide is active against enveloped viruses [85]. The introduction of ornithine in position 11 of Pro3TL in the place of Leu11, or the replacement of Pro3 by 4-amino-proline resulted in a peptide with further reduced haemolytic activity and antimicrobial activity similar to that of Pro3TL [97,98].
The analogue of temporin L with norleucine (Nle) in position 1 and two more substitutions (NLe1, DLeu9, DLys10) differs from all the others in terms of structure; it is not a helical peptide. CD data suggest the formation of beta sheet structures in the presence of 1,2-dioleoyl-sn-glycero-3-phospho-(1′-rac-glycerol) (DOPG)/cardiolipin (CL) and DOPG/1,2-Dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE) LUV. The increase in the hydrophobicity of the peptide due to the presence of NLe was related to an increased anti-inflammatory activity of the peptide in murine macrophage cell lines [99].
Srivastava and co-workers identified a phenylalanine zipper-like motif in temporin L. To determine the role of this structural motif, analogues in which phenylalanines in positions “a” and “d” of the zipper motif were replaced by alanine or leucine residues were obtained. It was found that the phenylalanine zipper motif affects the ability of the peptide to interact with LPS [100]. The introduction of leucines in place of phenylalanines in the analogue F5,8L TempL results in the introduction of a leucine zipper motif, which leads to increased LPS binding activity, increased ability of these peptides to dissociate LPS aggregates and, finally, enhanced endotoxic activity of the peptide. This peptide exhibits an alpha helical structure in the presence of LPS, with a percentage of helicity higher than temporin L. The haemolytic activity of the double-substituted F5,8L TempL is higher than temporin L, while the antibacterial activity is not improved. Peptides F5L TempL and F8L Temp L show moderately reduced haemolytic activity as compared to temporin L. In contrast, the replacement of phenylalanine by alanine impairs the ability of the peptide to bind to and fold in the presence of LPS, and reduces the production of LPS-induced pro-inflammatory cytokines. Interestingly, the peptide F5,8L TempL shows drastically reduced haemolytic and antibacterial activities.
Another interesting analogue of temporin L was designed by Kumari et al., starting from the analogue Q3K, which shows potent antiendotoxic activity and cytotoxicity comparable to the natural temporin L, changing the phenylalanine residues located at the hydrophobic face of the peptide (F8) and in the “d” position of the phenylalanine zipper by the cationic lysine residue and swapping the tryptophan residue W4 with serine S6. The analogue, named SW, Q3K, F8K, shows improved antimicrobial activity, comparable antiendotoxic activity and reduced haemolytic activity as compared to temporin L [101]. The low cytotoxicity and antimicrobial activity were also demonstrated in mice challenged with lethal doses of P. aeruginosa. This peptide is amphipathic and has clusters of hydrophobic and hydrophilic amino acids, resulting in a disordered secondary structure in bacterial membrane mimetic lipid vesicles. Overall data reported by Kumari and co-workers suggest new directions for the design of noncytotoxic antimicrobial peptides, highlighting the importance of a high hydrophobic moment together with a low secondary structure content.
3. Temporins from Hylarana
Peptides produced by four frogs belonging to the Hylarana family have been isolated. Limited data are available (Table 7); for example, in the case of temporin PTa from Hylarana picturata, we know that it is a 14-amino-acid peptide and that it is active against both Gram-positive bacteria, such as S. aureus, and Gram-negative bacteria, such as E. coli [102]. More information is available on peptides produced by Hylarana guentheri, named temporin GHa, GHb, GHc and GHd.
Temporins GHa to GHd were identified by analysing the sequence of cDNA isolated from Hylarana guentheri, a frog living in South China [103]. These are 13-amino-acid peptides, lacking positively charged residues such as arginine or lysine. According to secondary structure predictions, all peptides but temporin GHb show a certain degree of helicity. They have a broad spectrum of antimicrobial activity, being able to kill both Gram-positive and Gram-negative bacteria, including the methicillin-resistant S. aureus, and fungi. The activity of these peptides is independent of salt concentrations and remains almost unaltered when they are incubated with human serum and S. aureus V8 protease. The analysis of the morphology of different strains of E. coli and S. aureus treated with peptides suggests that peptides likely act by perturbing the cell membrane.
Temporin GHa Analogues
Analogues of temporin GHa were obtained by replacing histidines in position 4 or 11 or both with lysines [102]. The peptide temporin GHaK with both histidines replaced shows increased activity against the tested strains of Gram-positive and Gram-negative bacteria. In addition, these peptides prevent the adhesion and formation of biofilms by S. aureus. Bacterial membrane permeability was increased upon incubation with the peptide. In a recent work, arginines were introduced in sequences to replace histidines in position 4 and 11 to give GHaR, as well as amino acids in positions 6, 7 and 8 to give temporins GHaR6R, GHaR7R, GHaR8R and GHaR9R [102]. The secondary structure of these peptides is predicted to be alpha helical. An analogue of temporin GHaR named GHaR9W, in which Leu9 was replaced by tryptophan, was also obtained. In general, the introduction of positive charges causes an increase in the antimicrobial activity of the peptides; all derivatives are more active than temporin GHa. Haemolytic activity appears to be high for GHaR and reduced in GHaR6R. SEM analyses revealed that these peptides severely perturb the membrane of S. mutans.
Temporin GHaR analogues show potent activity against MRSA, permeabilizing its cytoplasmic membrane, and are able to inhibit MRSA biofilm formation and eradicate mature biofilms. Interestingly, these peptides trigger the downregulation of the expression of the genes icaADBC involved in biofilm formation [105].
4. Temporins from Lithobates
Several temporins from the Lithobates genera have been investigated so far; data on the biological activities of these compounds are reported (Table 8), but to the best of our knowledge, their mechanism of action is not known. For example, temporins 1Ca to 1Ce isolated from skin extracts of Rana clamitans are 13-amino-acid peptides [106]. The analysis of their antimicrobial activity reveals that these peptides are active against S. aureus, and not against E. coli and C. albicans.
The temporin 1ARa isolated from skin secretions of Lithobates areolata is active against S. aureus. At high concentrations, it is active against E. coli and is inactive against C. albicans [107]. It shows haemolytic activity (HC50) at 210 µM concentration on human erythrocytes. No structure or mechanism has been reported so far.
Temporins CPa and CPb are secreted by the gopher frog L. capito [109]. These are 13-amino-acid peptides. The temporin CPa has an atypical primary structure (IPPFIKKVLTTVF-NH2) and activity, being more active against Gram-negative than Gram-positive bacteria. Haemolytic activity measured against human erythrocytes as HL50 is >120 µM. Studies on the anticancer activity of these peptides carried out using mouse breast cancer cells (4T1) indicate that LC50 were >80 µM.
5. Temporins from Odorrana
Skin secretion of Odorrana hainanensis contains peptides belonging to the temporin family, namely temporin HN1 (AILTTLANWARKFL-NH2) and HN2 (NILNTIINLAKKIL-NH2) [110]. Both contain 14 amino acids and two positively charged amino acids. These peptides are mainly active against Gram-positive bacteria and fungi such as C. albicans, with temporin HN2 being the most active. Unlike other temporins, these are poor in leucines and contain alanines (Table 9). The haemolytic activity of these peptides is weak. According to predictions, these temporins should have an α-helical structure. The Chinese bamboo leaf odorous frog Odorrana versabilis secretes many antimicrobial peptides, including temporin 1VE [111]. Other than the sequence, there is no information on its activity or structure.
6. Temporins from Pelophylax
Studies on temporin secreted by frogs of the Pelophylax family are mainly focused on peptides secreted by the Pelophylax saharica.
6.1. Temporin SH Family
Peptides belonging to the family of temporin SH were identified in the skin of the North African frog Pelophylax saharica [112]. Currently, six members have been identified and characterized (see Table 1). They present a length ranging from 8 to 17 residues with different net charges. These peptides are active against Gram-positive and Gram-negative bacteria, yeasts and fungi with MIC values below 30 µM (see Table 10). They also show activity against Leishmania, L. pneumophila and Helicobacter pylori. In some cases, anticancer activity was detected. Peptides Temporin SHa and SHf are the most interesting members of this family.
6.1.1. Temporin SHa Biological Activity
Temporin SHa presents a net charge of 2+, a broad-spectrum of activity and high potency against Gram-negative and Gram-positive bacteria (Table 10). It is particularly active against E. coli and P. aeruginosa (MIC = 31 µM). The haemolytic activity, measured as LC50, is 25 µM. Temporin SHa shows high activity against clinical strains of H. pylori and no toxicity on human gastric cells in vitro and on human gastric explants. Its activity is not reduced in the presence of mucins or human gastric mucus. Instead, the presence of pepsin prevents temporin SHa activity [117]. The peptide is active against several species of Legionella, with MIC values ranging from 3 to 6 μM. Experiments on L. pneumophila cells showed that the SHa peptide induces cell permeabilization, which would mainly account for the bactericidal activity of this peptide. Temporin SHa is able to reduce the amount of L. pneumophila inside eukaryotic cells, inducing a host response, as it is able to penetrate into the cytoplasm but is not able to enter vacuoles; for these reasons, it protects the bacteria from the action of the peptide [118].
Furthermore, it showed antiparasitic activity (IC50 = 20 µM) against the promastigote and amastigote stages of Leishmania infantum and Leishmania Mexicana (4 and 42 μM, respectively) [119]. The difference in activity was attributed to the different composition of the membrane of the two parasite forms. Specifically, the peptide temporin SHa interacts with the proteophosphoglycans present on the surface of the cell in the promastigote form but absent in the cell surface of the amastigote stage [119]. Temporin SHa shows anti-viral activity against herpes simplex virus type 1, directly acting on the virus (virucidal activity) instead of eliciting an immunomodulatory response [120]. Temporin SHa shows anticancer activity against specific cancer cell lines such as MCF-7 (human breast cancer; IC50 = 14.5 μM), HeLa (IC50 = 18.4 μM) and NCI-H460 (NSLC; IC50 = 34.5 μM).
6.1.2. Temporin SHa Structure and Mechanism of Action
The NMR structure of temporin SHa was determined in 80 mM SDS. In water, a small amount of secondary structure is present. In the presence of SDS, the peptide adopts an amphipathic helical secondary structure involving residues 3 to 12 (Figure 3). Analysis with paramagnetic probes to detect the position of the peptide within the micelle suggests that temporin SHa adopts a nearly parallel orientation with respect to the micelle surface. Binding studies with multilamellar vesicles suggest a binding mode based on in-plane insertion into the lipid bilayer [113]. The decrease in hydrophobicity, replacing large aliphatic residues (leucine and valine) with alanine in the [A2,6,9]SHa analogue peptide, reduces the ability of the peptide to fold in an α-helical structure in the presence of the membrane model system, and prevents antimicrobial activity, suggesting a primary role of the interaction between he hydrophobic side chains and lipid acyl chains.
6.1.3. Temporin SHf Biological Activity
Another interesting and peculiar member of the temporin SH family is the peptide temporin SHf. It is a short 8-mer peptide rich in the aromatic residue phenylalanine [116]. It shows broad antimicrobial activity against Gram-positive and Gram-negative bacteria and yeasts. Exceptions are E. coli D21, E. coli ATCC 35218, P. aeruginosa and the fungus Aspergillus flavus. No activity was detected against the promastigote and axenic amastigote forms of Leishmania infantum. Temporin SHf is not cytotoxic against human erythrocytes (LC50 = 200 µM) and has no haemolytic activity at a 50 μM concentration.
6.1.4. Temporin SHa Structure and Mechanism of Action
Structural analysis of temporin SHa in DPC micelles reveals that the peptide adopts an α-helix conformation involving residues 3 to 8, with the peptide interacting with the surface of the micelle and inserting Phe 3, Leu 4 and Phe 8 residues inside the micelle [116]. The mechanism of action consists of membrane permeabilization with release of the intracellular content. Calorimetric studies carried out in the presence of 1,2-dimyristoyl-sn-glycero-3-phosphocholine (DMPC), a model for mammalian cell membranes, or dimyristoyl phosphatidylglycerol (DMPG), a model for negatively charged bacterial membrane, suggest that the peptide temporin SHf interacts strongly and exclusively with DMPG, indicating a selectivity towards the bacterial cell membrane.
6.1.5. Temporin SH Analogues
Several analogues of temporin SHa were obtained by introducing D-alanine in the place of amino acids in position 4, 7 and 10 to give [G4a]-SHa, [G7a]-SHa and [G10a]-SHa peptides. These analogues were tested, and their mechanism of action evaluated against methicillin-resistant S. aureus [121]. [G10a]-SHa showed an MIC comparable with the natural peptide (3.58 vs. 3.62 μM), but performed better in physiological conditions such as 30% foetal bovine serum, 30% human serum and 10% human blood (in these conditions, the natural peptide was unstable). In addition, [G10a]-SHa is very active against clinical strains of H. pylori and maintains its activity after treatment with pepsin [117]. However, it has a more pronounced cytotoxicity and haemolytic profile with respect to the parent and other analogue peptides. Overall, the [G10a]-SHa peptide seems to be more suitable for skin applications. Atomic force microscopy (AFM) experiments showed that the mechanism of action of these peptides consists of membrane rupturing, triggering leakage of cytoplasmic content. [G10a]-SHa is cytotoxic to NIH-3T3 murine fibroblast cell line and more active than temporin SHa against the indicated cancer cell lines. The conjugation of [G10a]-SHa with a breast cancer-targeting peptide furnishes a hybrid peptide with no cytotoxicity towards non-cancer cells but still active against MCF-7 cell lines [122].
The peptide [K3]SHa contains a lysine in place of a serine in position 3; it is non-haemolytic and it is more active than the parent peptide against several Gram-positive and Gram-negative bacterial strains, including multidrug-resistant strains of the clinically relevant pathogenic species S. aureus. This peptide kills bacteria and induces a potent membrane permeabilization/depolarization, likely through a detergent-like mechanism. The improved antimicrobial activity with respect to the parent peptide could be, in part, ascribed to a stronger interaction with the anionic membrane. Temporin [K3]SHa retains the ability to fold in an α-helical structure in the presence of membrane mimetic due to enhanced electrostatic interaction [123]. Interestingly, temporin [K3]SHa shows antifungal activity, with an MIC in the range 5.5–45 μM. In particular, it acts rapidly against C. albicans, inducing permeabilization of the membrane [124]. It also shows leishmanicidal activity, being more potent than temporin SHa. A growth inhibitory effect was observed with promastigotes of the Leishmania infantum, Leishmania major, Leishmania amazonensis and Leishmania braziliensis species, with amastigote forms of L. infantum and other trypanosomatids, such as Trypanosoma brucei gambiense and Trypanosomacruzi [123]. The anti-Leishmania activity was also analysed; it is mainly due to a membranolytic mechanism at peptide concentrations above IC50, whereas below IC50, it can induce apoptosis-like cell death. Finally, temporin [K3]SHa shows anti-viral activity [120].
Several temporin SHf analogues were designed in order to improve the antimicrobial activity without increasing the haemolytic activity [125]. Structure analysis studies revealed that the analogue [p-tBuF2, R5]SHf, presenting the unnatural amino acid p-terbutyl-phenylalanine in position 2 and arginine in position 5, retains the helical conformation in the presence of SDS and shows improved antimicrobial activity with respect to the natural peptide, being also active against Gram-negative bacteria such as E. coli ATCC 25922 at physiological salt concentration (150 mM NaCl) and in 30% serum.
7. Biomedical/Biotechnological Applications of Temporins
Applications of temporins as drugs are still limited, likely due to their quick in vivo degradation. The formulation of temporins into liposomal vectors or otherwise has been reported [126]. Some examples will be reported here. The encapsulation of temporin B into chitosan nanoparticles was described; encapsulation of the peptide reduces its cytotoxicity against mammalian cells. Sustained antibacterial activity was demonstrated against various strains of S. epidermidis; the gradual release of the peptide results in a reduction in viable bacterial count and inhibition of the regrowth of residual cells [126]. The use of temporins for topical applications to combat Candida infections was recently hypothesized [77]. Studies aimed at formulating the peptide temporin L have been reported. In 2020, Brancaccio and co-workers described the use of cyclodextrins as tools to improve the pharmacokinetic profile and reduce the toxicity of temporin L. Cyclodextrins complexed to temporin L were demonstrated to effectively hamper the attachment of biofilms produced by clinical isolates of P. aeruginosa and MRSA and eradicate bacteria. The enzyme-triggered release of a caged temporin L was proposed and tested in HEK293T cells [94].
The analogue of temporin L named [Pro3]Temporin L was encapsulated in poly(2-hydroxyethyl methacrylate) microgel particles to improve delivery and control the peptide release. The analysis of antibacterial activity against S. aureus showed that encapsulation was effective; the peptide maintains its biological activity and a sustained release of the peptide was observed [127].
More recently, L-K6, an analogue of temporin 1CEa, was embedded in a liposome presenting on its surface folic acid as a cancer-targeting agent and evaluated in vitro and in vivo on human breast cancer MCF-7 cells [128]. The peptide was selectively targeted to cancer cells, and tumour growth was reduced by about three times in ten days of observation. Even though L-K6 was able to selectively target MCF-7 cancer cells, the use of a decorated liposome was needed to reduce enzymatic degradation [31].
Peptides can be immobilized on solid surfaces to produce antimicrobial materials, which is useful for the production of implantable materials [129]. Temporins SHa, [K3]SHa and D-[K3]SHa were covalently linked on a gold surface; immobilized peptides retain antimicrobial activity against Listeria ivanovii. Immobilization was performed by exploiting amide bond formation, reacting either the amin of the amino terminus or lysine side chains, or the carboxy C-terminus of a modified peptide. The surface decorated with temporin D-[K3]SHa showed effective antiadhesive properties reducing the bacterial adhesion to the modified gold surface by up to 50%. Immobilization through the C-terminal end was the most efficient [130,131]. A strategy to immobilize temporin SHa on a titanium surface was reported [132]. Chemoselectively immobilized temporin SHa on titanium surfaces showed the ability to inhibit the growth of E. coli and S. epidermidis. Better results are achieved when the peptide is grafted through a central lysine residue instead of via the N- and C-termini. In a different report, the temporin B analogue TB_KKG6A was immobilized onto microcrystalline cellulose; the functionalization of the cellulose confers antimicrobial activity against E. coli cells to the polymer [133].
Temporin A associated through electrostatic interactions with the enzyme organophosphorus hydrolase was found to overcome bacterial resistance acting on the quorum sensing mechanism mediated by N-acyl homoserine lactones. This nanocomplex showed improved antibacterial activity with respect the peptide alone when tested against Bacillus subtilis and Pseudomonas sp. [134]. Successively, this nanocomplex was entrapped in poly(vinyl alcohol) cryogel (PVA-CG)/bacterial cellulose material, forming a composite antibacterial material or bacterial cellulose [135,136].
Rana chensinensis skin peptides with antimicrobial properties including peptides belonging to the temporin 1C family were used to prepare different biomaterials for wound healing application. Composite nanofibres, loaded with peptides, were prepared through coaxial electrospinning of polyvinylpyrrolidone and sodium alginate, and through uniaxial electrospinning of polyvinylpyrrolidone and polycaprolactone also loaded with silver nanoparticles [137,138]. In vitro and in vivo experiments showed that the nanofibres release the bioactive compounds, are biocompatible and promote wound healing. Hybrid microspheres were also prepared using a polyvinyl alcohol gelatine hydrogel matrix and sodium alginate (SA) frog egg-like microspheres. This system can deliver Rana chensinensis skin peptides and induces wound repair in vivo [139].
Temporins have also been exploited to produce biosensors. Temporin PTa was implemented in an electrochemical biosensor for the detection and differentiation of bacterial strains [140]. The sensor is based on a gold electrode. The transducing layer contains a monolayer of mercaptobenzoic acid and gold-capped magnetic nanoparticles (Fe3O4Au), decorated with 3-(aminopropyl)triethoxysilane and finally derivatized with the peptide. Analysis is performed through electrochemical spectroscopy impedance and cyclic voltammetry. The sensor detects Gram-negative bacteria such as K. pneumoniae and A. baumannii with a limit of 10 CFU/mL and discriminates other organisms such as B. subtilis and C. albicans, exploiting structural differences in the bacterial membranes.
Temporin L was used as a membrane-interacting domain to develop a PET probe for in vivo imaging of granzyme B activity. Enzymatic cleavage allows radiolabelled temporin L to assume a helical conformation and insert into adjacent cellular membranes [141].
8. Conclusions
Antimicrobial peptides belonging to the temporin family have been isolated from the skin of frogs from all over the world or have been identified from cDNA libraries of skin secretions. Irrespective of the geographical location of the frog or the frog genus, temporin sequences are rich in leucines and isoleucines, while lacking cysteine. Most of the temporins contain a single positively charged amino acid (typically one lysine). These features determine their 3D structure and their antimicrobial activity. While most temporins effectively kill Gram-positive bacteria, some, such as temporin L from Rana temporaria or temporin CPa from L. capito, show strong activity against Gram-negative bacteria. Most temporins have no preferential structure in buffer and assume a helical structure in membrane-mimicking environments (e.g., in POPG vesicles or DOPG/PC mixtures) or in the presence of components of the bacterial outer membrane (such as LPS). The formation of beta-sheet structures has been hypothesized for temporin Rb in DOPG/DOPE/CL LUV based on CD analysis [101]. The interactions of peptides with the bacterial outer membrane have been demonstrated in many cases using fluorescence microscopy techniques, while intracellular targets have been identified in only a few cases. Due to their short sequences, these peptides are ideal candidates for structure–activity studies and, therefore, an ideal starting point to produce new and more active compounds. Changes in the peptide primary structure have been related to increases in their antimicrobial or haemolytic activity; in particular, many studies demonstrate that an increase in the positive charges of peptides translates into an increase in activity against Gram-negative bacteria.
In consideration of the growing interest of the scientific community in these molecules, based on the exponential increase in published papers on temporins and their analogues, in the near future, we expect to see a deeper exploitation of temporins in technological and medical applications.
Author Contributions
Conceptualization, writing, review and editing, L.D.D. and A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Konig, E.; Bininda-Emonds, O.R.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef] [PubMed]
- Erspamer, V.; Ottolenghi, A. Preliminary researches on the mechanism of the antidiuretic action of enteramine. Experientia 1951, 7, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Iwamuro, S.; King, J.D. Dermal cytolytic peptides and the system of innate immunity in anurans. Ann. N. Y. Acad. Sci. 2009, 1163, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Structural diversity and species distribution of host-defense peptides in frog skin secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial Peptides: Amphibian Host Defense Peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. (Landmark Ed.) 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [Green Version]
- Vanhoye, D.; Bruston, F.; Nicolas, P.; Amiche, M. Antimicrobial peptides from hylid and ranin frogs originated from a 150-million-year-old ancestral precursor with a conserved signal peptide but a hypermutable antimicrobial domain. Eur. J. BioChem. 2003, 270, 2068–2081. [Google Scholar] [CrossRef]
- Samgina, T.Y.; Vasileva, I.D.; Trebse, P.; Torkar, G.; Surin, A.K.; Meng, Z.; Zubarev, R.A.; Lebedev, A.T. Mass Spectrometry Differentiation between Rana arvalis Populations Based on Their Skin Peptidome Composition. J. Am. Soc. Mass Spectrom. 2022, 33, 1480–1491. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, A.; Conlon, J.M.; Kawasaki, H.; Iwamuro, S. Developmental and triiodothyronine-induced expression of genes encoding preprotemporins in the skin of Tago’s brown frog Rana tagoi. Gen. Comp. Endocrinol. 2006, 146, 242–250. [Google Scholar] [CrossRef]
- Matutte, B.; Storey, K.B.; Knoop, F.C.; Conlon, J.M. Induction of synthesis of an antimicrobial peptide in the skin of the freeze-tolerant frog, Rana sylvatica, in response to environmental stimuli. FEBS Lett. 2000, 483, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G. Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction. Antibiotics 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Cortazar-Chinarro, M.; Meyer-Lucht, Y.; Van der Valk, T.; Richter-Boix, A.; Laurila, A.; Hoglund, J. Antimicrobial peptide and sequence variation along a latitudinal gradient in two anurans. BMC Genet. 2020, 21, 38. [Google Scholar] [CrossRef]
- Samgina, T.Y.; Vasileva, I.D.; Kovalev, S.V.; Trebse, P.; Torkar, G.; Surin, A.K.; Zubarev, R.A.; Lebedev, A.T. Differentiation of Central Slovenian and Moscow populations of Rana temporaria frogs using peptide biomarkers of temporins family. Anal. Bioanal. Chem. 2021, 413, 5333–5347. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; Mignogna, G.; Barra, D.; Bossa, F. Antimicrobial peptides from skin secretions of Rana esculenta. Molecular cloning of cDNAs encoding esculentin and brevinins and isolation of new active peptides. J. Biol. Chem. 1994, 269, 11956–11961. [Google Scholar] [CrossRef]
- Romero, S.M.; Cardillo, A.B.; Martinez Ceron, M.C.; Camperi, S.A.; Giudicessi, S.L. Temporins: An Approach of Potential Pharmaceutic Candidates. Surg. Infect. (Larchmt) 2020, 21, 309–322. [Google Scholar] [CrossRef]
- Conlon, J.M. Reflections on a systematic nomenclature for antimicrobial peptides from the skins of frogs of the family Ranidae. Peptides 2008, 29, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef]
- Shang, D.; Yu, F.; Li, J.; Zheng, J.; Zhang, L.; Li, Y. Molecular cloning of cDNAs encoding antimicrobial peptide precursors from the skin of the Chinese brown frog, Rana chensinensis. Zool. Sci. 2009, 26, 220–226. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, Y.; Li, Z.; Su, Q. Molecular cloning of novel antimicrobial peptide genes from the skin of the Chinese brown frog, Rana chensinensis. Zool. Sci. 2011, 28, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, Y.; Kang, Y.; Shang, D. Antimicrobial peptide temporin-1CEa isolated from frog skin secretions inhibits the proinflammatory response in lipopolysaccharide-stimulated RAW264.7 murine macrophages through the MyD88-dependent signaling pathway. Mol. Immunol. 2021, 132, 227–235. [Google Scholar] [CrossRef]
- Wang, C.; Li, H.B.; Li, S.; Tian, L.L.; Shang, D.J. Antitumor effects and cell selectivity of temporin-1CEa, an antimicrobial peptide from the skin secretions of the Chinese brown frog (Rana chensinensis). Biochimie 2012, 94, 434–441. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Y.W.; Zhang, L.; Gong, X.G.; Zhou, Y.; Shang, D.J. Melanoma cell surface-expressed phosphatidylserine as a therapeutic target for cationic anticancer peptide, temporin-1CEa. J. Drug Target. 2016, 24, 548–556. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, Y.; Li, S.; Li, H.; Tian, L.; Wang, H.; Shang, D. Anticancer mechanisms of temporin-1CEa, an amphipathic alpha-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sci. 2013, 92, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhou, X.; Xi, X.; Zai, Y.; Zhou, M.; Chen, X.; Ma, C.; Chen, T.; Wang, L.; Kwok, H.F. In Vitro & In Vivo Studies on Identifying and Designing Temporin-1CEh from the Skin Secretion of Rana chensinensis as the Optimised Antibacterial Prototype Drug. Pharmaceutics 2022, 14, 604. [Google Scholar] [PubMed]
- Yang, Q.Z.; Wang, C.; Lang, L.; Zhou, Y.; Wang, H.; Shang, D.J. Design of potent, non-toxic anticancer peptides based on the structure of the antimicrobial peptide, temporin-1CEa. Arch. Pharm. Res. 2013, 36, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Shang, D.; Li, X.; Sun, Y.; Wang, C.; Sun, L.; Wei, S.; Gou, M. Design of potent, non-toxic antimicrobial agents based upon the structure of the frog skin peptide, temporin-1CEb from Chinese brown frog, Rana chensinensis. Chem. Biol. Drug Des. 2012, 79, 653–662. [Google Scholar] [CrossRef]
- Shang, D.; Liang, H.; Wei, S.; Yan, X.; Yang, Q.; Sun, Y. Effects of antimicrobial peptide L-K6, a temporin-1CEb analog on oral pathogen growth, Streptococcus mutans biofilm formation, and anti-inflammatory activity. Appl. Microbiol. Biotechnol. 2014, 98, 8685–8695. [Google Scholar] [CrossRef]
- Dong, W.; Zhu, X.; Zhou, X.; Yang, Y.; Yan, X.; Sun, L.; Shang, D. Potential role of a series of lysine-/leucine-rich antimicrobial peptide in inhibiting lipopolysaccharide-induced inflammation. Biochem. J. 2018, 475, 3687–3706. [Google Scholar] [CrossRef]
- Bi, X.; Wang, C.; Ma, L.; Sun, Y.; Shang, D. Investigation of the role of tryptophan residues in cationic antimicrobial peptides to determine the mechanism of antimicrobial action. J. Appl. Microbiol. 2013, 115, 663–672. [Google Scholar] [CrossRef]
- Bi, X.; Wang, C.; Dong, W.; Zhu, W.; Shang, D. Antimicrobial properties and interaction of two Trp-substituted cationic antimicrobial peptides with a lipid bilayer. J. Antibiot. (Tokyo) 2014, 67, 361–368. [Google Scholar] [CrossRef]
- Wang, C.; Dong, S.; Zhang, L.; Zhao, Y.; Huang, L.; Gong, X.; Wang, H.; Shang, D. Cell surface binding, uptaking and anticancer activity of L-K6, a lysine/leucine-rich peptide, on human breast cancer MCF-7 cells. Sci. Rep. 2017, 7, 8293. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Huang, L.L.; Li, R.J.; Wang, Y.; Wu, X.X.; Shang, D.J. Synergistic Therapy of Doxorubicin with Cationic Anticancer Peptide L-K6 Reverses Multidrug Resistance in MCF-7/ADR Cancer Cells In Vitro via P-glycoprotein Inhibition. Int. J. Pept. Res. Ther. 2021, 27, 2291–2301. [Google Scholar] [CrossRef]
- Kosikowska, P.; Pikula, M.; Langa, P.; Trzonkowski, P.; Obuchowski, M.; Lesner, A. Synthesis and Evaluation of Biological Activity of Antimicrobial--Pro-Proliferative Peptide Conjugates. PLoS ONE 2015, 10, e0140377. [Google Scholar] [CrossRef]
- Golda, A.; Kosikowska-Adamus, P.; Babyak, O.; Lech, M.; Wysocka, M.; Lesner, A.; Potempa, J.; Koziel, J. Conjugate of Enkephalin and Temporin Peptides as a Novel Therapeutic Agent for Sepsis. Bioconjug. Chem. 2018, 29, 4127–4139. [Google Scholar] [CrossRef] [PubMed]
- Golda, A.; Kosikowska-Adamus, P.; Kret, A.; Babyak, O.; Wojcik, K.; Dobosz, E.; Potempa, J.; Lesner, A.; Koziel, J. The Bactericidal Activity of Temporin Analogues Against Methicillin Resistant. Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 4761. [Google Scholar] [CrossRef] [Green Version]
- Ji, F.; Zhao, Y.; Jiang, F.; Shang, D. Membrane mechanism of temporin-1CEc, an antimicrobial peptide isolated from the skin secretions of Rana chensinensis, and its systemic analogs. Bioorg. Chem. 2022, 119, 105544. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L. Temporins, anti-infective peptides with expanding properties. Cell. Mol. Life Sci. 2006, 63, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.; Silberring, J.; Soliymani, R.; Heikkinen, S.; Kilpelainen, I.; Lankinen, H.; Kuusela, P. Antibacterial activities of temporin A analogs. Febs. Lett. 2000, 479, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Luca, V.; Segev-Zarko, L.A.; Shai, Y.; Mangoni, M.L. Temporins A and B stimulate migration of HaCaT keratinocytes and kill intracellular Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 2520–2527. [Google Scholar] [CrossRef] [Green Version]
- Simonetti, O.; Cirioni, O.; Goteri, G.; Ghiselli, R.; Kamysz, W.; Kamysz, E.; Silvestri, C.; Orlando, F.; Barucca, C.; Scalise, A.; et al. Temporin A is effective in MRSA-infected wounds through bactericidal activity and acceleration of wound repair in a murine model. Peptides 2008, 29, 520–528. [Google Scholar] [CrossRef]
- Ghiselli, R.; Giacometti, A.; Cirioni, O.; Mocchegiani, F.; Orlando, F.; Kamysz, W.; Del Prete, M.S.; Lukasiak, J.; Scalise, G.; Saba, V. Temporin A as a prophylactic agent against methicillin sodium-susceptible and methicillin sodium-resistant Staphylococcus epidermidis vascular graft infection. J. Vasc. Surg. 2002, 36, 1027–1030. [Google Scholar] [CrossRef] [Green Version]
- Ciandrini, E.; Morroni, G.; Cirioni, O.; Kamysz, W.; Kamysz, E.; Brescini, L.; Baffone, W.; Campana, R. Synergistic combinations of antimicrobial peptides against biofilms of methicillin-resistant Staphylococcus aureus (MRSA) on polystyrene and medical devices. J. Glob. Antimicrob. Resist. 2020, 21, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; Silvestri, C.; Licci, A.; Riva, A.; Lukasiak, J.; Scalise, G. In vitro activity of amphibian peptides alone and in combination with antimicrobial agents against multidrug-resistant pathogens isolated from surgical wound infection. Peptides 2005, 26, 2111–2116. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Orlando, F.; D’Amato, G.; Kamysz, W.; Mocchegiani, F.; Sisti, V.; Silvestri, C.; Lukasiak, J.; et al. Temporin A soaking in combination with intraperitoneal linezolid prevents vascular graft infection in a subcutaneous rat pouch model of infection with Staphylococcus epidermidis with intermediate resistance to glycopeptides. Antimicrob. Agents Chemother. 2004, 48, 3162–3164. [Google Scholar] [CrossRef] [Green Version]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Kamysz, W.; Orlando, F.; Mocchegiani, F.; Silvestri, C.; Licci, A.; Lukasiak, J.; Saba, V.; et al. Temporin A alone and in combination with imipenem reduces lethality in a mouse model of staphylococcal sepsis. J. Infect. Dis. 2005, 192, 1613–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paduszynska, M.A.; Greber, K.E.; Paduszynski, W.; Sawicki, W.; Kamysz, W. Activity of Temporin A and Short Lipopeptides Combined with Gentamicin against Biofilm Formed by Staphylococcus aureus and Pseudomonas aeruginosa. Antibiotics 2020, 9, 566. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta 2009, 1788, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Capparelli, R.; Romanelli, A.; Iannaccone, M.; Nocerino, N.; Ripa, R.; Pensato, S.; Pedone, C.; Iannelli, D. Synergistic antibacterial and anti-inflammatory activity of temporin A and modified temporin B in vivo. PLoS ONE 2009, 4, e7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinchar, V.G.; Bryan, L.; Silphadaung, U.; Noga, E.; Wade, D.; Rollins-Smith, L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004, 323, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Marimuthu, S.K.; Nagarajan, K.; Perumal, S.K.; Palanisamy, S.; Subbiah, L. Insilico Alpha-Helical Structural Recognition of Temporin Antimicrobial Peptides and Its Interactions with Middle East Respiratory Syndrome-Coronavirus. Int. J. Pept. Res. Ther. 2020, 26, 1473–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangoni, M.L.; Saugar, J.M.; Dellisanti, M.; Barra, D.; Simmaco, M.; Rivas, L. Temporins, small antimicrobial peptides with leishmanicidal activity. J. Biol. Chem. 2005, 280, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Chadbourne, F.L.; Raleigh, C.; Ali, H.Z.; Denny, P.W.; Cobb, S.L. Studies on the antileishmanial properties of the antimicrobial peptides temporin A, B and 1Sa. J. Pept. Sci. 2011, 17, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wade, D.; Kurosaka, K.; Wang, Z.Y.; Oppenheim, J.J.; Yang, D. Temporin A and related frog antimicrobial peptides use formyl peptide receptor-like 1 as a receptor to chemoattract phagocytes. J. Immunol. 2004, 173, 2652–2659. [Google Scholar] [CrossRef] [Green Version]
- Swithenbank, L.; Cox, P.; Harris, L.G.; Dudley, E.; Sinclair, K.; Lewis, P.; Cappiello, F.; Morgan, C. Temporin A and Bombinin H2 Antimicrobial Peptides Exhibit Selective Cytotoxicity to Lung Cancer Cells. Scientificn (Cairo) 2020, 2020, 3526286. [Google Scholar] [CrossRef]
- Musale, V.; Casciaro, B.; Mangoni, M.L.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Conlon, J.M. Assessment of the potential of temporin peptides from the frog Rana temporaria (Ranidae) as anti-diabetic agents. J. Pept. Sci. 2018, 24, e3065. [Google Scholar] [CrossRef]
- Xu, L.F.; Li, Y.; Jin, R.; Jiang, D.P.; Jiang, D.C. High spatial resolution observation of Temporin A at cell membranes using scanning ion conductive microscopy. Electrochem. Commun. 2022, 134, 107181. [Google Scholar] [CrossRef]
- Carotenuto, A.; Malfi, S.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.; Mangoni, M.L.; Gaddi, L.M.; Novellino, E.; Grieco, P. A different molecular mechanism underlying antimicrobial and hemolytic actions of temporins A and L. J. Med. Chem. 2008, 51, 2354–2362. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, R.; Joshi, M.; Mohanram, H.; Bhunia, A.; Mangoni, M.L.; Bhattacharjya, S. NMR structure of temporin-1 ta in lipopolysaccharide micelles: Mechanistic insight into inactivation by outer membrane. PLoS ONE 2013, 8, e72718. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, Y.; Barra, D.; Simmaco, M.; Shai, Y.; Mangoni, M.L. A synergism between temporins toward Gram-negative bacteria overcomes resistance imposed by the lipopolysaccharide protective layer. J. Biol. Chem. 2006, 281, 28565–28574. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Rinaldi, A.C.; Di Giulio, A.; Mignogna, G.; Bozzi, A.; Barra, D.; Simmaco, M. Structure-function relationships of temporins, small antimicrobial peptides from amphibian skin. Eur. J. Biochem. 2000, 267, 1447–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uccelletti, D.; Zanni, E.; Marcellini, L.; Palleschi, C.; Barra, D.; Mangoni, M.L. Anti-Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: A plausible mode of action in vitro and in vivo. Antimicrob. Agents Chemother. 2010, 54, 3853–3860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avitabile, C.; Netti, F.; Orefice, G.; Palmieri, M.; Nocerino, N.; Malgieri, G.; D’Andrea, L.D.; Capparelli, R.; Fattorusso, R.; Romanelli, A. Design, structural and functional characterization of a Temporin-1b analog active against Gram-negative bacteria. Biochim. Biophys. Acta 2013, 1830, 3767–3775. [Google Scholar] [CrossRef] [Green Version]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, e02367-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, R.; Domanov, Y.; Kinnunen, P.K. Fluorescent temporin B derivative and its binding to liposomes. J. Fluoresc. 2007, 17, 223–234. [Google Scholar] [CrossRef]
- Malgieri, G.; Avitabile, C.; Palmieri, M.; D’Andrea, L.D.; Isernia, C.; Romanelli, A.; Fattorusso, R. Structural basis of a temporin 1b analogue antimicrobial activity against Gram negative bacteria determined by CD and NMR techniques in cellular environment. ACS Chem. Biol. 2015, 10, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.; Saravanan, R.; Mohanram, H.; Mangoni, M.L.; Bhattacharjya, S. NMR structures and interactions of temporin-1Tl and temporin-1Tb with lipopolysaccharide micelles: Mechanistic insights into outer membrane permeabilization and synergistic activity. J. Biol. Chem. 2011, 286, 24394–24406. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Rinaldi, A.C.; Di Giulio, A.; Simmaco, M.; Kinnunen, P.K. Interactions of the antimicrobial peptides temporins with model biomembranes. Comparison of temporins B and L. Biochemistry 2002, 41, 4425–4436. [Google Scholar] [CrossRef]
- Mahalka, A.K.; Kinnunen, P.K. Binding of amphipathic alpha-helical antimicrobial peptides to lipid membranes: Lessons from temporins B and L. Biochim. Biophys. Acta 2009, 1788, 1600–1609. [Google Scholar] [CrossRef] [Green Version]
- Domanov, Y.A.; Kinnunen, P.K. Antimicrobial peptides temporins B and L induce formation of tubular lipid protrusions from supported phospholipid bilayers. Biophys. J. 2006, 91, 4427–4439. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Ma, M.; Shao, Z.; Zhang, J.; Fu, L.; Li, X.; Fang, W.; Gao, L. Structure and Formation Mechanism of Antimicrobial Peptides Temporin B- and L-Induced Tubular Membrane Protrusion. Int. J. Mol. Sci. 2021, 22, 11015. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Epand, R.F.; Rosenfeld, Y.; Peleg, A.; Barra, D.; Epand, R.M.; Shai, Y. Lipopolysaccharide, a key molecule involved in the synergism between temporins in inhibiting bacterial growth and in endotoxin neutralization. J. Biol. Chem. 2008, 283, 22907–22917. [Google Scholar] [CrossRef] [Green Version]
- Avitabile, C.; D’Andrea, L.D.; Saviano, M.; Olivieri, M.; Cimmino, A.; Romanelli, A. Binding studies of antimicrobial peptides to Escherichia coli cells. Biochem. Biophys. Res. Commun. 2016, 478, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Bezzerri, V.; Avitabile, C.; Dechecchi, M.C.; Lampronti, I.; Borgatti, M.; Montagner, G.; Cabrini, G.; Gambari, R.; Romanelli, A. Antibacterial and anti-inflammatory activity of a temporin B peptide analogue on an in vitro model of cystic fibrosis. J. Pept. Sci. 2014, 20, 822–830. [Google Scholar] [CrossRef]
- Hurley, K.A.; Santos, T.M.A.; Nepomuceno, G.M.; Huynh, V.; Shaw, J.T.; Weibel, D.B. Targeting the Bacterial Division Protein FtsZ. J. Med. Chem. 2016, 59, 6975–6998. [Google Scholar] [CrossRef] [PubMed]
- Kakar, A.; Holzknecht, J.; Dubrac, S.; Gelmi, M.L.; Romanelli, A.; Marx, F. New Perspectives in the Antimicrobial Activity of the Amphibian Temporin B: Peptide Analogs Are Effective Inhibitors of Candida albicans Growth. J. Fungi. 2021, 7, 457. [Google Scholar] [CrossRef] [PubMed]
- Kakar, A.; Sastre-Velasquez, L.E.; Hess, M.; Galgoczy, L.; Papp, C.; Holzknecht, J.; Romanelli, A.; Varadi, G.; Malanovic, N.; Marx, F. The Membrane Activity of the Amphibian Temporin B Peptide Analog TB_KKG6K Sheds Light on the Mechanism That Kills Candida albicans. mSphere 2022, 7, e0029022. [Google Scholar] [CrossRef]
- Grassi, L.; Maisetta, G.; Maccari, G.; Esin, S.; Batoni, G. Analogs of the Frog-skin Antimicrobial Peptide Temporin 1Tb Exhibit a Wider Spectrum of Activity and a Stronger Antibiofilm Potential as Compared to the Parental Peptide. Front. Chem. 2017, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Maisetta, G.; Di Luca, M.; Gaddi, L.M.; Esin, S.; Florio, W.; Brancatisano, F.L.; Barra, D.; Campa, M.; Batoni, G. Comparative analysis of the bactericidal activities of amphibian peptide analogues against multidrug-resistant nosocomial bacterial strains. Antimicrob. Agents Chemother. 2008, 52, 85–91. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, F.D.; Casciaro, B.; De Angelis, M.; Marcocci, M.E.; Palamara, A.T.; Nencioni, L.; Mangoni, M.L. Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. Int. J. Mol. Sci. 2022, 23, 6345. [Google Scholar] [CrossRef]
- Casciaro, B.; Loffredo, M.R.; Cappiello, F.; Fabiano, G.; Torrini, L.; Mangoni, M.L. The Antimicrobial Peptide Temporin G: Anti-Biofilm, Anti-Persister Activities, and Potentiator Effect of Tobramycin Efficacy Against Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 9410. [Google Scholar] [CrossRef]
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Jackowska, B.G.; Prezioso, C.; Protto, V.; De Angelis, M.; Di Leva, F.S.; Casciaro, B.; Carotenuto, A.; Mangoni, M.L.; Palamara, A.T.; et al. The Inhibition of DNA Viruses by the Amphibian Antimicrobial Peptide Temporin G: A Virological Study Addressing HSV-1 and JPCyV. Int. J. Mol. Sci. 2022, 23, 7194. [Google Scholar] [CrossRef]
- Di Somma, A.; Recupido, F.; Cirillo, A.; Romano, A.; Romanelli, A.; Caserta, S.; Guido, S.; Duilio, A. Antibiofilm Properties of Temporin-L on Pseudomonas fluorescens in Static and In-Flow Conditions. Int. J. Mol. Sci. 2020, 21, 8526. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Mocchegiani, F.; Orlando, F.; Silvestri, C.; Bozzi, A.; Di Giulio, A.; Luzi, C.; Mangoni, M.L.; et al. Interaction of antimicrobial peptide temporin L with lipopolysaccharide in vitro and in experimental rat models of septic shock caused by gram-negative bacteria. Antimicrob. Agents Chemother. 2006, 50, 2478–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368 Pt 1, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Kinnunen, P.K. Binding of the antimicrobial peptide temporin L to liposomes assessed by Trp fluorescence. J. Biol. Chem. 2002, 277, 25170–25177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avitabile, C.; D’Andrea, L.D.; Saviano, M.; Romanelli, A. Determination of the secondary structure of peptides in the presence of Gram positive bacterium S. epidermidis cells. RSC Adv. 2016, 6, 51407–51410. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Barra, D.; Simmaco, M.; Bozzi, A.; Di Giulio, A.; Rinaldi, A.C. Effects of the antimicrobial peptide temporin L on cell morphology, membrane permeability and viability of Escherichia coli. Biochem. J. 2004, 380 Pt 3, 859–865. [Google Scholar] [CrossRef] [Green Version]
- Manzo, G.; Ferguson, P.M.; Hind, C.K.; Clifford, M.; Gustilo, V.B.; Ali, H.; Bansal, S.S.; Bui, T.T.; Drake, A.F.; Atkinson, R.A.; et al. Temporin L and aurein 2.5 have identical conformations but subtly distinct membrane and antibacterial activities. Sci. Rep. 2019, 9, 10934. [Google Scholar] [CrossRef] [Green Version]
- Di Somma, A.; Avitabile, C.; Cirillo, A.; Moretta, A.; Merlino, A.; Paduano, L.; Duilio, A.; Romanelli, A. The antimicrobial peptide Temporin L impairs E. coli cell division by interacting with FtsZ and the divisome complex. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129606. [Google Scholar] [CrossRef]
- Brancaccio, D.; Pizzo, E.; Cafaro, V.; Notomista, E.; De Lise, F.; Bosso, A.; Gaglione, R.; Merlino, F.; Novellino, E.; Ungaro, F.; et al. Antimicrobial peptide Temporin-L complexed with anionic cyclodextrins results in a potent and safe agent against sessile bacteria. Int. J. Pharm. 2020, 584, 119437. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, S.; Kashibe, M.; Matsumoto, K.; Hori, Y.; Kikuchi, K. Enzyme-triggered compound release using functionalized antimicrobial peptide derivatives. Chem. Sci. 2017, 8, 3047–3053. [Google Scholar] [CrossRef] [Green Version]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.M.; Marcellini, L.; Luca, V.; Barra, D.; Novellino, E.; et al. The effect of D-amino acid substitution on the selectivity of temporin L towards target cells: Identification of a potent anti-Candida peptide. BBA-Biomembr. 2013, 1828, 652–660. [Google Scholar] [CrossRef]
- Bellavita, R.; Raucci, F.; Merlino, F.; Piccolo, M.; Ferraro, M.G.; Irace, C.; Santamaria, R.; Iqbal, A.J.; Novellino, E.; Grieco, P.; et al. Temporin L-derived peptide as a regulator of the acute inflammatory response in zymosan-induced peritonitis. Biomed. Pharm. 2020, 123, 109788. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Carotenuto, A.; Antignano, I.; Bellavita, R.; Casciaro, B.; Loffredo, M.R.; Merlino, F.; Novellino, E.; Mangoni, M.L.; Nocera, F.P.; et al. The Outcomes of Decorated Prolines in the Discovery of Antimicrobial Peptides from Temporin-L. ChemMedChem 2019, 14, 1283–1290. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.; Malfi, S.; Marcellini, L.; Barra, D.; Novellino, E.; et al. Structure-activity relationship, conformational and biological studies of temporin L analogues. J. Med. Chem. 2011, 54, 1298–1307. [Google Scholar] [CrossRef]
- Bellavita, R.; Buommino, E.; Casciaro, B.; Merlino, F.; Cappiello, F.; Marigliano, N.; Saviano, A.; Maione, F.; Santangelo, R.; Mangoni, M.L.; et al. Synthetic Amphipathic beta-Sheet Temporin-Derived Peptide with Dual Antibacterial and Anti-Inflammatory Activities. Antibiotics 2022, 11, 1285. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Kumar, A.; Tripathi, A.K.; Tandon, A.; Ghosh, J.K. Modulation of anti-endotoxin property of Temporin L by minor amino acid substitution in identified phenylalanine zipper sequence. Biochem. J. 2016, 473, 4045–4062. [Google Scholar] [CrossRef]
- Kumari, T.; Verma, D.P.; Afshan, T.; Verma, N.K.; Pant, G.; Ali, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. A Noncytotoxic Temporin L Analogue with In Vivo Antibacterial and Antiendotoxin Activities and a Nonmembrane-Lytic Mode of Action. ACS Infect. Dis. 2020, 6, 2369–2385. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Luo, W.; Zhong, H.; Wang, M.; Song, Y.; Deng, S.; Zhang, Y. Molecular cloning and characterization of antimicrobial peptides from skin of Hylarana guentheri. Acta Biochim. Biophys. Sin. (Shanghai) 2017, 49, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.; Wang, G. Ab initio design of potent anti-MRSA peptides based on database filtering technology. J. Am. Chem. Soc. 2012, 134, 12426–12429. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Xie, Z.; Tan, X.; Guo, R.; Song, Y.; Xie, X.; Wang, R.; Li, L.; Wang, M.; Zhang, Y. Temporin-Like Peptides Show Antimicrobial and Anti-Biofilm Activities against Streptococcus mutans with Reduced Hemolysis. Molecules 2020, 25, 5724. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Xie, S.; Jin, X.; Zhu, R.; Wei, S.; Song, Y.; Xie, X.; Wang, R.; Li, L.; Wang, M.; et al. Four temporin-derived peptides exhibit antimicrobial and antibiofilm activities against methicillin-resistant. Acta Biochim. Biophys. Sin. (Shanghai) 2022, 54, 350–360. [Google Scholar]
- Halverson, T.; Basir, Y.J.; Knoop, F.C.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin of the North American green frog Rana clamitans. Peptides 2000, 21, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.F.; Lips, K.R.; Knoop, F.C.; Fritzsch, B.; Miller, C.; Conlon, J.M. Antimicrobial peptides and protease inhibitors in the skin secretions of the crawfish frog, Rana areolata. Biochim. Biophys. Acta 2002, 1601, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Wang, X.; Lushnikova, T.; Zhang, Y.; Golla, R.M.; Narayana, J.L.; Wang, C.; McGuire, T.R.; Wang, G. Antibacterial, antifungal, anticancer activities and structural bioinformatics analysis of six naturally occurring temporins. Peptides 2018, 106, 9–20. [Google Scholar] [CrossRef]
- Conlon, J.M.; Meetani, M.A.; Coquet, L.; Jouenne, T.; Leprince, J.; Vaudry, H.; Kolodziejek, J.; Nowotny, N.; King, J.D. Antimicrobial peptides from the skin secretions of the New World frogs Lithobates capito and Lithobates warszewitschii (Ranidae). Peptides 2009, 30, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, Z.; Hu, Y.; Li, F.; Liu, L.; Zheng, H.; Meng, H.; Yang, S.; Yang, X.; Liu, J. Novel antimicrobial peptides isolated from the skin secretions of Hainan odorous frog, Odorrana hainanensis. Peptides 2012, 35, 285–290. [Google Scholar] [CrossRef]
- Chen, T.; Zhou, M.; Rao, P.; Walker, B.; Shaw, C. The Chinese bamboo leaf odorous frog (Rana (Odorrana) versabilis) and North American Rana frogs share the same families of skin antimicrobial peptides. Peptides 2006, 27, 1738–1744. [Google Scholar] [CrossRef]
- Abbassi, F.; Oury, B.; Blasco, T.; Sereno, D.; Bolbach, G.; Nicolas, P.; Hani, K.; Amiche, M.; Ladram, A. Isolation, characterization and molecular cloning of new temporins from the skin of the North African ranid Pelophylax saharica. Peptides 2008, 29, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Galanth, C.; Amiche, M.; Saito, K.; Piesse, C.; Zargarian, L.; Hani, K.; Nicolas, P.; Lequin, O.; Ladram, A. Solution structure and model membrane interactions of temporins-SH, antimicrobial peptides from amphibian skin. A NMR spectroscopy and differential scanning calorimetry study. Biochemistry 2008, 47, 10513–10525. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Raja, Z.; Oury, B.; Gazanion, E.; Piesse, C.; Sereno, D.; Nicolas, P.; Foulon, T.; Ladram, A. Antibacterial and leishmanicidal activities of temporin-SHd, a 17-residue long membrane-damaging peptide. Biochimie 2013, 95, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Andre, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional Characterization of Temporin-SHe, a New Broad-Spectrum Antibacterial and Leishmanicidal Temporin-SH Paralog from the Sahara Frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar] [CrossRef]
- Abbassi, F.; Lequin, O.; Piesse, C.; Goasdoue, N.; Foulon, T.; Nicolas, P.; Ladram, A. Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide. J. Biol. Chem. 2010, 285, 16880–16892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olleik, H.; Baydoun, E.; Perrier, J.; Hijazi, A.; Raymond, J.; Manzoni, M.; Dupuis, L.; Pauleau, G.; Goudard, Y.; Villeon, B.; et al. Temporin-SHa and Its Analogs as Potential Candidates for the Treatment of Helicobacter pylori. Biomolecules 2019, 9, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crepin, A.; Jegou, J.F.; Andre, S.; Ecale, F.; Croitoru, A.; Cantereau, A.; Berjeaud, J.M.; Ladram, A.; Verdon, J. In vitro and intracellular activities of frog skin temporins against Legionella pneumophila and its eukaryotic hosts. Sci. Rep. 2020, 10, 3978. [Google Scholar] [CrossRef] [Green Version]
- Eggimann, G.A.; Sweeney, K.; Bolt, H.L.; Rozatian, N.; Cobb, S.L.; Denny, P.W. The role of phosphoglycans in the susceptibility of Leishmania mexicana to the temporin family of anti-microbial peptides. Molecules 2015, 20, 2775–2785. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Leveque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K3]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, R.; Khan, A.I.; Nadeem-Ul-Haque, M.; Maresca, M.; Choudhary, M.I.; Shaheen, F.; Simjee, S.U. Serum Stable and Low Hemolytic Temporin-SHa Peptide Analogs Disrupt Cell Membrane of Methicillin-Resistant Staphylococcus aureus (MRSA). Probiotics Antimicrob. Proteins 2022, 14, 391–405. [Google Scholar] [CrossRef]
- Shaheen, F.; Nadeem-Ul-Haque, M.; Ahmed, A.; Simjee, S.U.; Ganesan, A.; Jabeen, A.; Shah, Z.A.; Choudhary, M.I. Synthesis of breast cancer targeting conjugate of temporin-SHa analog and its effect on pro- and anti-apoptotic protein expression in MCF-7 cells. Peptides 2018, 106, 68–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, Z.; Andre, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, K.; Verdon, J.; Ladram, A.; Arnault, S.; Rodier, M.H.; Cateau, E. Antifungal activity of [K(3)]temporin-SHa against medically relevant yeasts and moulds. Can. J. Microbiol. 2022, 68, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Andre, S.; Washington, S.K.; Darby, E.; Vega, M.M.; Filip, A.D.; Ash, N.S.; Muzikar, K.A.; Piesse, C.; Foulon, T.; O’Leary, D.J.; et al. Structure-Activity Relationship-based Optimization of Small Temporin-SHf Analogs with Potent Antibacterial Activity. ACS Chem. Biol. 2015, 10, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef] [Green Version]
- Parilti, R.; Caprasse, J.; Riva, R.; Alexandre, M.; Vandegaart, H.; Bebrone, C.; Dupont-Gillain, C.; Howdle, S.M.; Jerome, C. Antimicrobial peptide encapsulation and sustained release from polymer network particles prepared in supercritical carbon dioxide. J. Colloid. Interf. Sci. 2018, 532, 112–117. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.; Wang, B.; Wang, R.X.; Wang, C.; Shang, D.J. Characterization and Anticancer Activity of a Folic Acid Conjugated and Cationic Peptide L-K6 Encapsulated Cancer-Targeting Liposomal Drug Delivery System. Int. J. Pept. Res. Ther. 2022, 28, 82. [Google Scholar] [CrossRef]
- Rai, A.; Ferrao, R.; Palma, P.; Patricio, T.; Parreira, P.; Anes, E.; Tonda-Turo, C.; Martins, M.C.L.; Alves, N.; Ferreira, L. Antimicrobial peptide-based materials: Opportunities and challenges. J. Mater. Chem. B 2022, 10, 2384–2429. [Google Scholar] [CrossRef]
- Lombana, A.; Raja, Z.; Casale, S.; Pradier, C.M.; Foulon, T.; Ladram, A.; Humblot, V. Temporin-SHa peptides grafted on gold surfaces display antibacterial activity. J. Pept. Sci. 2014, 20, 563–569. [Google Scholar] [CrossRef]
- Oger, P.C.; Piesse, C.; Ladram, A.; Humblot, V. Engineering of Antimicrobial Surfaces by Using Temporin Analogs to Tune the Biocidal/antiadhesive Effect. Molecules 2019, 24, 814. [Google Scholar] [CrossRef] [Green Version]
- Masurier, N.; Tissot, J.B.; Boukhriss, D.; Jebors, S.; Pinese, C.; Verdie, P.; Amblard, M.; Mehdi, A.; Martinez, J.; Humblot, V.; et al. Site-specific grafting on titanium surfaces with hybrid temporin antibacterial peptides. J. Mater. Chem. B 2018, 6, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, P.; Bosco, F.; Clerici, F.; Polissi, A.; Gelmi, M.L.; Romanelli, A. Covalent Grafting of Antimicrobial Peptides onto Microcrystalline Cellulose. ACS Appl. Bio Mater. 2020, 3, 4895–4901. [Google Scholar] [CrossRef] [PubMed]
- Aslanli, A.; Lyagin, I.; Efremenko, E. Charges’ interaction in polyelectrolyte (nano)complexing of His(6)-OPH with peptides: Unpredictable results due to imperfect or useless concept? Int. J. Biol. Macromol. 2019, 140, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Aslanli, A.; Stepanov, N.; Razheva, T.; Podorozhko, E.A.; Lyagin, I.; Lozinsky, V.I.; Efremenko, E. Enzymatically Functionalized Composite Materials Based on Nanocellulose and Poly(Vinyl Alcohol) Cryogel and Possessing Antimicrobial Activity. Materials 2019, 12, 3619. [Google Scholar] [CrossRef] [Green Version]
- Aslanli, A.; Lyagin, I.; Stepanov, N.; Presnov, D.; Efremenko, E. Bacterial Cellulose Containing Combinations of Antimicrobial Peptides with Various QQ Enzymes as a Prototype of an “Enhanced Antibacterial” Dressing: In Silico and In Vitro Data. Pharmaceutics 2020, 12, 1155. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Cheng, Z.; Wen, R.; Zhao, X.; Yu, X.; Sun, L.; Zhang, Y.; Han, Z.; Yuan, Y.; Kang, L. Novel SA@Ca2+/RCSPs core-shell structure nanofibers by electrospinning for wound dressings. RSC Adv. 2018, 8, 15558–15566. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Li, R.; Liu, G.M.; Jia, W.Y.; Sun, M.L.; Liu, Y.; Luo, Y.G.; Cheng, Z.Q. Phase separation-based electrospun Janus nanofibers loaded with Rana chensinensis skin peptides/silver nanoparticles for wound healing. Mater. Des. 2021, 207, 109864. [Google Scholar] [CrossRef]
- Ji, X.; Liu, G.M.; Cui, Y.T.; Jia, W.Y.; Luo, Y.G.; Cheng, Z.Q. A hybrid system of hydrogel/frog egg-like microspheres accelerates wound healing via sustained delivery ofRCSPs. J. Appl. Polym. Sci. 2020, 137, 718377. [Google Scholar] [CrossRef]
- da Silva-Junio, A.G.; Frias, I.A.M.; Lima-Neto, R.G.; Migliolo, L.; PS, E.S.; Oliveira, M.D.L.; Andrade, C.A.S. Electrochemical biosensor based on Temporin-PTA peptide for detection of microorganisms. J. Pharm. Biomed. Anal. 2022, 216, 114788. [Google Scholar] [CrossRef]
- Zhao, N.; Bardine, C.; Lourenco, A.L.; Wang, Y.H.; Huang, Y.; Cleary, S.J.; Wilson, D.M.; Oh, D.Y.; Fong, L.; Looney, M.R.; et al. In Vivo Measurement of Granzyme Proteolysis from Activated Immune Cells with PET. ACS Cent. Sci. 2021, 7, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Superposition of the two structures of peptide TB_KKG6A, analogue of temporin B, obtained in the two different systems. In light grey, the structure in LPS; in gold, the TB_KKG6A structure obtained in the cellular context. The interacting residues that assume very similar orientations in the two environments are indicated in red. Reprinted with permission from Ref. [66]. Copyright 2015 American Chemical Society.
Figure 1.
Superposition of the two structures of peptide TB_KKG6A, analogue of temporin B, obtained in the two different systems. In light grey, the structure in LPS; in gold, the TB_KKG6A structure obtained in the cellular context. The interacting residues that assume very similar orientations in the two environments are indicated in red. Reprinted with permission from Ref. [66]. Copyright 2015 American Chemical Society.

Figure 2.
Structure of temporin L in LPS. Residues involved in dimer stabilization are indicated in magenta [67].
Figure 2.
Structure of temporin L in LPS. Residues involved in dimer stabilization are indicated in magenta [67].

Figure 3.
NMR structure in SDS of peptide (A) temporin SHa, (B) temporin SHb and (C) temporin SHc [113].
Figure 3.
NMR structure in SDS of peptide (A) temporin SHa, (B) temporin SHb and (C) temporin SHc [113].

Table 1.
Temporin sequences divided by frog genera as reported in the DBAASP database and recent reviews [15,17].
| Genus | Specie | Peptide Name | Sequence a |
|---|---|---|---|
| Amolops | |||
| A. chunganensis | Temporin CG1 | FLPFVGNLLKGLL | |
| Temporin CG2 | FFPIVGKLLSGLF | ||
| Temporin CG3 | FLPIVGKLLSGLF | ||
| Temporin CG4 | FLPILGNLLNGLL | ||
| Temporin CG5 | FLPFVGNLLNGLL | ||
| A. daiyunensis | Temporin DY1 | FFPMLADLVSKIF | |
| A. jingdongensis | Temporin AJ1 | FLPIVTGLLSSLL | |
| Temporin AJ2 | FFPIGGKLLFGLL | ||
| Temporin AJ3 | FFPIVGKLLSGLL | ||
| Temporin AJ4 | FFPIVGKLLFGLL | ||
| Temporin AJ5 | FPPIVGKLLFGLL | ||
| Temporin AJ6 | FLPIVGKLLSGLL | ||
| Temporin AJ7 | FLPIVGKLPSGLL | ||
| Temporin AJ8 | FFPIVGKRLYGLL | ||
| Temporin AJ9 | FLPIVGKLLSGLTGLL | ||
| Temporin AJ10 | FLPIVGKLLSGLSGLL | ||
| Temporin AJ11 | FFPIGGKLLSGLTGLL | ||
| A. lifanensis | Temporin LF1 | FLPFVGKLLSGLL | |
| A. loloensis | Temporin ALa | FLPIVGKLLSGLSGLL | |
| Temporin ALb | FLPIVGKLLSGLL | ||
| Temporin ALc | LLPIVGKGLSGKL | ||
| Temporin ALd | FLPIAGKLLSGLSGLL | ||
| Temporin ALe | FFPIVGKLLFGLSGLL | ||
| Temporin ALf | FFPIVGKLLSGLSGLL | ||
| Temporin ALg | FFPIVGKLLFGLFGLL | ||
| Temporin ALh | FLPIVGKLLSGLSGLS | ||
| Temporin ALi | FFPIVGKLLSGLL | ||
| Temporin ALj | FFPIVGKLLFGLL | ||
| Temporin ALk | FFPIVGKLLS | ||
| Euphlyctis | |||
| E. cyanophlyctis | Temporin ECa | FLPGLLAGLL | |
| Hylarana | |||
| H. guentheri | Temporin GH | FLPLLFGAISHLL | |
| Temporin GHa | FLQHIIGALGHLF | ||
| Temporin GHb | FIHHIIGALGHLF | ||
| Temporin GHc | FLQHIIGALTHIF | ||
| Temporin GHd | FLQHIIGALSHFF | ||
| Temporin GUa | FLQHIIGALSHIF | ||
| Temporin GUb | FLPLLFGAISHIL | ||
| Temporin GUc | FFPLIFGALSSILPKIL | ||
| H. latouchii | Temporin LT1 | FLPGLIAGIAKML | |
| Temporin LT2 | FLPIALKALGSIFPKIL | ||
| Temporin LTa | FFPLVLGALGSILPKIF | ||
| Temporin LTb | FIITGLVRGLTKLF | ||
| Temporin LTc | SLSRFLSFLKIVYPPAF | ||
| Temporin LTe | FLAGLIGGLAKML | ||
| Temporin HLa | FFPLIFGALSSILPKIL | ||
| Temporin HLb | FLQHIIGALSHIF | ||
| H. picturata | Temporin PTA | FFGSVLKLIPKIL | |
| H. taipehensis | Temporin TP1 | FLPVLGKVIKLVGGLL | |
| Temporin TP2 | FLPLLVGAISSILPKIF | ||
| Temporin TP3 | FLPLLFGALSTLLPKIF | ||
| H. spinulosa | Temporin SN1 | FFPFLLGALGSLLPKIF | |
| Temporin SN2 | FITGLIGGLMKAL | ||
| Temporin SN3 | FISGLIGGLMKAL | ||
| Temporin SN4 | FITGLISGLMKAL | ||
| Temporin SN5 | FFPLVLGALGSILPKIF | ||
| H. maosonensis | Temporin MS1 | FLTGLIGGLMKALGK | |
| Temporin MS4 | FLSGLIGGLAKMLGK | ||
| Limnonectes | |||
| L. kuhlii | Temporin LK1 | FFPLLFGALSSMMPKLF | |
| Lithobates | |||
| L. areolatus | Temporin 1AR | FLPIVGRLISGLL | |
| L. capito | Temporin CPa | IPPFIKKVLTTVF | |
| Temporin CPb | FLPIVGRLISGIL | ||
| L. catesbeianus | Temporin La | LLRHVVKILEKYL | |
| L. clamitans | Temporin 1Ca | FLPFLAKILTGVL | |
| Temporin 1Cb | FLPLFASLIGKLL | ||
| Temporin 1Cc | FLPFLASLLTKVL | ||
| Temporin 1Cd | FLPFLASLLSKVL | ||
| Temporin 1Ce | FLPFLATLLSKVL | ||
| L. grylio | Temporin 1Ga | SILPTIVSFLSKVF | |
| Temporin 1Gb | SILPTIVSFLSKFL | ||
| Temporin 1Gc | SILPTIVSFLTKFL | ||
| Temporin 1Gd | FILPLIASFLSKFL | ||
| L. heckscheri | Temporin 1HKa | SIFPAIVSFLSKFL | |
| L. okaloosae | Temporin 1OLa | FLPFLKSILGKIL | |
| Temporin 1OLb | FLPFFASLLGKLL | ||
| L. onca | Temporin ONa | FLPTFGKILSGLF | |
| L. palustris | Temporin 1PLa | FLPLVGKILSGLI | |
| L. pipiens | Temporin 1P | FLPIVGKLLSGLL | |
| L. septentrionalis | Temporin 1SPa | FLPLVGKILSGLI | |
| Temporin 1SPb | FLSAITSLLGKLL | ||
| Temporin 1SPc | FLSAITSILGKLF | ||
| L. sphenacephalus | Temporin 1S | LLFGKIISRLLGN | |
| L. virgatipes | Temporin 1Va | FLSSIGKILGNLL | |
| Temporin 1Vb | FLSIIAKVLGSLF | ||
| Temporin 1Vc | FLPLVTMLLGKLF | ||
| Odorrana | |||
| O. hainanensis | Temporin HN1 | AILTTLANWARKFL | |
| Temporia HN2 | NILNTIINLAKKIL | ||
| O. versabilis | Temporin 1VE | FLPLVGKILSGLI | |
| Pelophylax | |||
| P. esculentus | Temporin PE | FLPIVAKLLSGLL | |
| Temporin 1Re | FLPGLLAGLL | ||
| Temporin 1Ec | FLPVIAGLLSKLF | ||
| Temporin 1Ee | FLPVIAGVLSKLF | ||
| P. fukienensis | Temporin PF | FLPLIAGLFGKIF | |
| P. hubeiensis | Temporin HB1 | FLPLLAGLAAKWF | |
| Temporin HB2 | FLPFLAGLFGKIF | ||
| P. nigromaculatus | Temporin 1KM | FIPLVSGLFSRLL | |
| Temporin 1GY | VIPIVSGLLSSLL | ||
| P. ridibundus | Temporin Ra | FLKPLFNAALKLLP | |
| Temporin Rb | FLPVLAGVLSRA | ||
| P. saharicus | Temporin SHa | FLSGIVGMLGKLF | |
| Temporin SHb | FLPIVTNLLSGLL | ||
| Temporin SHc | FLSHIAGFLSNLF | ||
| Temporin SHd | FLPAALAGIGGILGKLF | ||
| Temporin SHe | FLPALAGIAGLLGKIF | ||
| Temporin SHf | FFFLSRIF | ||
| Rana | |||
| R. aurora | Temporin 1AUa | FLPIIGQLLSGLL | |
| R. boylii | Temporin 1BYa | FLPIIAKVLSGLL | |
| R. cascadae | Temporin 1CSa | FLPIVGKLLSGLL | |
| Temporin 1CSb | FLPIIGKLLSGLL | ||
| Temporin 1CSc | FLPLVTGLLSGLL | ||
| Temporin 1CSd | NFLGTLVNLAKKIL | ||
| R. chensinensis | Temporin 1CEa | FVDLKKIANIINSIF | |
| Temporin 1CEb | ILPILSLIGGLL | ||
| Temporin 1CEc | IIPLPLGYFAKKT | ||
| Temporin 1CEd | ILPLIASLIGGLL | ||
| Temporin 1CEe | ILPIIGKILSTIFGK | ||
| Temporin 1CEf | ILPILGKILSTIL | ||
| Temporin 1CEg | ILPIFSWIGHLF | ||
| Temporin 1CEh | FVDLKKIANILNSIF | ||
| R. draytonii | Temporin 1DRa | HFLGTLVNLAKKIL | |
| Temporin 1DRb | NFLGTLVNLAKKIL | ||
| Temporin 1DRc | FLPIIASVLSSLL | ||
| R. dybowskii | Temporin 1DYa | FIGPIISALASLFG | |
| Temporin CDYa | LPLVGNLLNDLL | ||
| Temporin CDYb | ILPILSLIGGLL | ||
| Temporin CDYc | VLPLVGNLLNDLL | ||
| Temporin CDYd | FIGPLISALASLFKG | ||
| Temporin CDYe | FIGPIISALASLFGG | ||
| R. italica | Temporin ITa | FLGAIAQALTSLLGKL | |
| R. japonica | Temporin 1Ja | ILPLVGNLLNDLL | |
| R. luteiventris | Temporin 1La | VLPLISMALGKLL | |
| Temporin 1Lb | NFLGTLINLAKKIM | ||
| Temporin 1Lc | FLPILINLIHKGLL | ||
| R. muscosa | Temporin 1M | FLPIVGKLLSGLL | |
| R. nigromaculata | Temporin 1RNa | ILPIRSLIKKLL | |
| Temporin 1RNb | FLPLKKLRFGLL | ||
| R. nigrovittata | Temporin RN1 | FLPLVLGALSGILPKIL | |
| Temporin RN2 | FFPLLFGALSSLLPKLF | ||
| Temporin RN3 | FFPLLFGALSSHLPKLF | ||
| R. ornativentris | Temporin 1Oa | FLPLLASLFSRLL | |
| Temporin 1Ob | FLPLIGKILGTIL | ||
| Temporin 1Oc | FLPLLASLFSRLF | ||
| Temporin 1Od | FLPLLASLFSGLF | ||
| Temporin 1Oe | ILPLLGNLLNGLL | ||
| Temporin 1Of | SLLLKGLASIAKLF | ||
| Temporin 1Og | FLSSLLSKVVSLFT | ||
| R. pirica | Temporin 1PRa | ILPILGNLLNGLL | |
| Temporin 1PRb | ILPILGNLLNSLL | ||
| R. pretiosa | Temporin PRa | FLPILGNLLSGLL | |
| Temporin PRb | FLPIITNLLGKLL | ||
| Temporin PRc | NFLDTLINLAKKFI | ||
| Temporin PRe | FLPLAMALGKLL | ||
| R. sakuraii | Temporin 1Ska | FLPVILPVIGKLLNGIL | |
| Temporin 1SKb | FLPVILPVIGKLLSGIL | ||
| R. tagoi | Temporin 1TGa | FLPILGKLLSGIL | |
| Temporin 1TGb | AVDLAKIANKVLSSLF | ||
| Temporin 1TGc | FLPVILPVIGKLLSGIL | ||
| R. tagoi okiensis | Temporin TOa | FLPILGKLLSGFL | |
| Temporin TOb | FLPILGKLLSGLL | ||
| R. temporaria | Temporin A | FLPLIGRVLSGIL | |
| Temporin B | LLPIVGNLLKSLL | ||
| Temporin C | ILPILGNLLNGLL | ||
| Temporin D | LLPIVGNLLNSLL | ||
| Temporin E | VLPIIGNLLNSLL | ||
| Temporin F | FLPLIGKVLSGIL | ||
| Temporin G | FFPVIGRILNGIL | ||
| Temporin H | LSPNLLKSLL | ||
| Temporin K | LPPNLLKSLL | ||
| Temporin L | FVQWFSKFLGRIL | ||
| R. tsushimensis | Temporin 1TSa | FLGALAKIISGIF | |
| Temporin 1TSb | FLPLLGNLLNGLL | ||
| Temporin 1TSc | FLPLLGNLLRGLL | ||
| Temporin 1TSd | FLPLLASLIGGML |
a C-terminal end is amidated.
Table 3.
Antimicrobial activity against selected microorganisms of temporin A.
| Microorganism | Antimicrobial Activity (µM) | Ref |
|---|---|---|
| E. coli D21 | LC: 11.9 | [37] |
| S. aureus Cowan 1 | LC: 2.3 | [37] |
| C. albicans ATCC 10261 | LC: 3.4 | [37] |
LC: lethal concentration.
Table 4.
Antimicrobial activity against selected microorganisms of peptide temporin B.
| Microorganism | Antimicrobial Activity | Ref |
|---|---|---|
| E. coli a | MIC: >100 μg/mL | [63] |
| S. aureus A170 | MIC 100: 25 μg/mL | [63] |
| C. albicans ATCC 10261 | LC: 4.0 μM | [37] |
a strain not indicated; MIC: minimum inhibitory concentration; LC: lethal concentration.
Table 5.
Antimicrobial activity against selected microorganisms of peptide temporin G.
| Microorganism | Antimicrobial Activity (µM) | Ref |
|---|---|---|
| S. aureus a | BC: 24 | [79] |
| C. albicans ATCC10231 | MIC: 16 | [80] |
a strain 1 as defined in [79]; BC: bactericidal concentration; MIC: minimum inhibitory concentration.
Table 6.
Antimicrobial activity against selected microorganisms of peptide temporin L.
| Microorganism | Antimicrobial Activity as MIC (µM) | Ref |
|---|---|---|
| E. coli D21 | 12 | [58] |
| S. aureus ATCC 25923 | 3 | [58] |
| C. albicans ATCC 10231 | 12 | [58] |
MIC: minimum inhibitory concentration.
Table 7.
Antimicrobial activity against selected microorganisms of peptides from Hylarana.
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC 25923 | Ref | E. coli ATCC 25922 | Ref | C. albicans ATCC 10231 | Ref | |
| Temporin GHa | 6.8 | [103] | 26.6 | [103] | 26.6 | [103] |
| Temporin GHb | 6.8 | [103] | 13.2 | [103] | 26.4 | [103] |
| Temporin GHc | 12.9 | [103] | 104 | [103] | 104 | [103] |
| Temporin GHd | 12.7 | [103] | 12.7 | [103] | 25.5 | [103] |
| Temporin PTa | 3.12 a | [104] | 25 b | [104] | n.d. | |
MIC: minimum inhibitory concentration; the peptide was tested on a: S.aureus USA300, b: E.coli K12; n.d.: not determined.
Table 8.
Antimicrobial activity against selected microorganisms of temporins from Lithobates.
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus | Ref | E. coli ATCC 25922 | Ref | C. albicans | Ref | |
| Temporin 1Cb | 140 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1Cd | 80 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1Ce | 100 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1AR | 15 a | [107] | 125 | [107] | N.A. c,e | [107] |
| Temporin CPa | ≥25 b | [108] | 12.5 | [108] | 25 d | [108] |
| Temporin CPb | 12.5 b | [108] | >50 | [108] | 50 d | [108] |
MIC: minimum inhibitory concentration; N.A.: not active at 150 μM; the peptide was tested a: on S. aureus NCTC 8325, b: on S. aureus USA300, c: on C. albicans ATCC90028, d: on C. albicans ATCC 10231, e: at 200 μM.
Table 9.
Antimicrobial activity against selected microorganisms of temporins from Odorrana.
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC25923 | Ref | E. coli ATCC25922 | Ref | C. albicans ATCC2002 | Ref | |
| Temporin HN1 | 37.5 | [110] | N.A. | [110] | 75 | [110] |
| Temporin HN2 | 4.8 | [110] | N.A. | [110] | 9.8 | [110] |
MIC: minimum inhibitory concentration; N.A.: not active at 150 μM.
Table 10.
Antimicrobial activity against selected microorganisms of temporins SH.
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC25923 | Ref | E. coli ATCC25922 | Ref | C. albicans ATCC90028 | Ref | |
| Temporin SHa | 3 | [113] | 10 | [113] | 16 | [113] |
| Temporin SHb | 58 | [113] | 231 | [113] | >116 | [113] |
| Temporin SHc | 10 | [113] | >161 | [113] | 20 | [113] |
| Temporin SHd | 6.25 | [114] | 5 | [114] | 100 | [114] |
| Temporin SHe | 3.12 | [115] | 25 | [115] | >100 | [115] |
| Temporin SHf | 12.5 | [116] | 25 | [116] | 50 | [116] |
MIC: minimum inhibitory concentration.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
D'Andrea, L.D.; Romanelli, A. Temporins: Multifunctional Peptides from Frog Skin. Int. J. Mol. Sci. 2023, 24, 5426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065426
AMA Style
D'Andrea LD, Romanelli A. Temporins: Multifunctional Peptides from Frog Skin. International Journal of Molecular Sciences. 2023; 24(6):5426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065426
Chicago/Turabian StyleD'Andrea, Luca Domenico, and Alessandra Romanelli. 2023. "Temporins: Multifunctional Peptides from Frog Skin" International Journal of Molecular Sciences 24, no. 6: 5426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065426
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.