
Interaction between Sulfate and Selenate in Tetraploid Wheat (Triticum turgidum L.) Genotypes

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Biomass Production and Chlorophyll Content
2.2. Sulfur and Selenium Concentrations
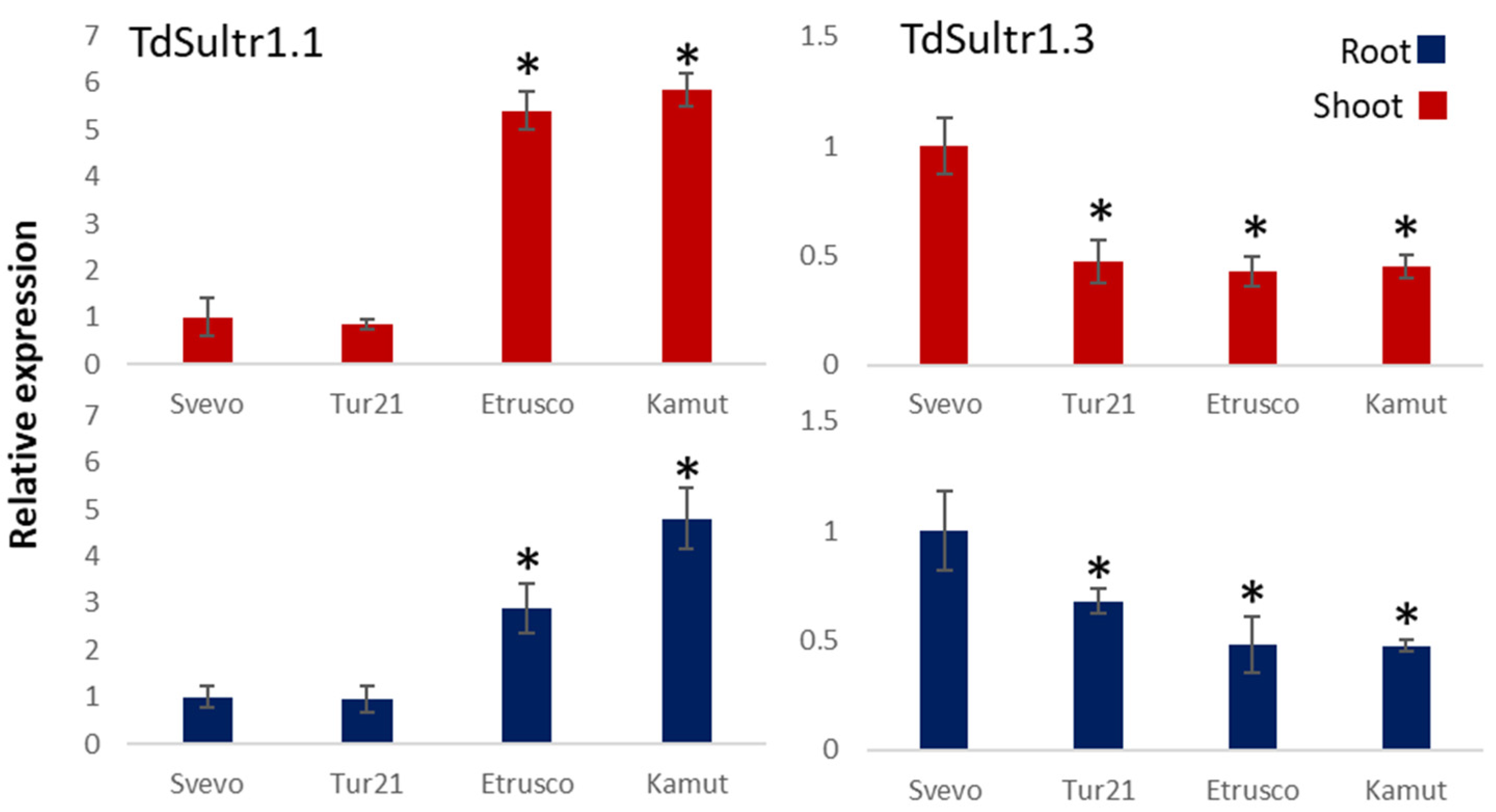
2.3. Expression of Genes Coding for High-Affinity Sulfate Transporters
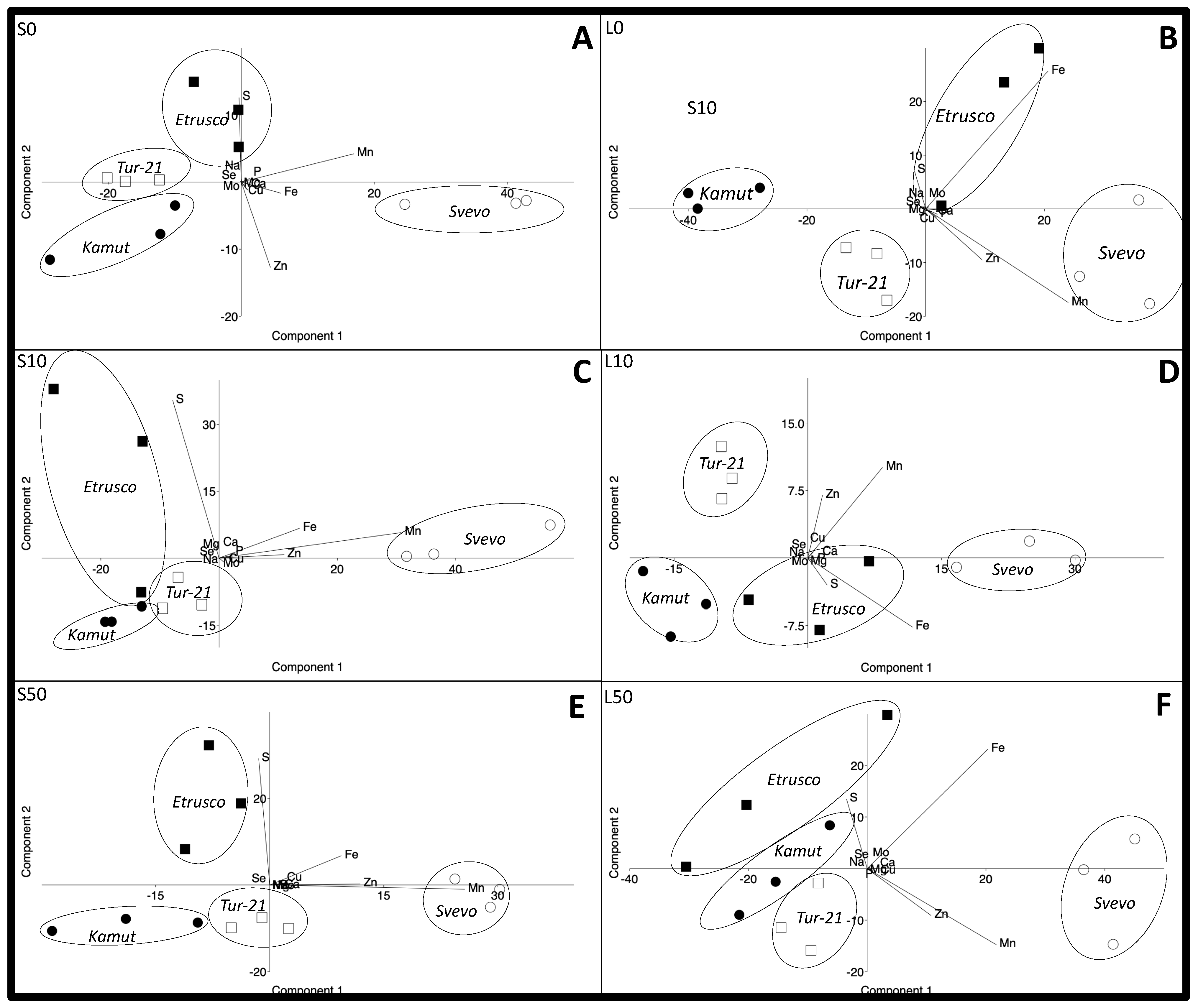
2.4. Principal Component Analysis (PCA) to Reveal Alterations within the Ionome of Plant Shoots
3. Discussion
4. Materials and Methods
4.1. Plant Growth
4.2. Chlorophyll Content
4.3. Analysis of Micro- and Macronutrient Concentrations
4.4. Total RNA Extraction and RT-PCR Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mora, M.; Durán, P.; Acuña, J.; Cartes, P.; Demanet, R.; Gianfreda, L. Improving selenium status in plant nutrition and quality. J. Soil Sci. Plant Nutr. 2015, 15, 486–503. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q.; Broadley, M. Selenium Biofortification. In Selenium in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.Q., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 231–255. [Google Scholar]
- Barchielli, G.; Capperucci, A.; Tanini, D. The Role of Selenium in Pathologies: An Updated Review. Antioxidants 2022, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; Pilon-Smits, E.A.H. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron. Soil Sci. 2014, 60, 1593–1624. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Modulation of Antioxidant Machinery and the Methylglyoxal Detoxification System in Selenium-Supplemented Brassica napus Seedlings Confers Tolerance to High Temperature Stress. Biol. Trace Elem. Res. 2014, 161, 297–307. [Google Scholar] [CrossRef]
- Floor, G.H.; Román-Ross, G. Selenium in volcanic environments: A review. Appl. Geochemistry 2012, 27, 517–531. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Na, G.N.; Lahner, B.; Lee, S.; Leustek, T.; Pickering, I.J.; Salt, D.E. Analysis of sulfur and selenium assimilation in Astragalus plants with varying capacities to accumulate selenium. Plant J. 2005, 42, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Hladun, K.R.; Parker, D.R.; Tran, K.D.; Trumble, J.T. Effects of selenium accumulation on phytotoxicity, herbivory, and pollination ecology in radish (Raphanus sativus L.). Environ. Pollut. 2013, 172, 70–75. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Shinmachi, F.; Buchner, P.; Stroud, J.L.; Parmar, S.; Zhao, F.J.; Mcgrath, S.P.; Hawkesford, M.J. Influence of sulfur deficiency on the expression of specific sulfate transporters and the distribution of sulfur, selenium, and molybdenum in wheat. Plant Physiol. 2010, 153, 327–336. [Google Scholar] [CrossRef] [Green Version]
- White, P.J. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Elselvier Ltd: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Parker, D.R.; Page, A.L.; Bell, P.F. Contrasting Selenate-Sulfate Interactions in Selenium-Accumulating and Nonaccumulating Plant Species. Soil Sci. Soc. Am. J. 1992, 56, 1818–1824. [Google Scholar] [CrossRef]
- White, P.J.; Bowen, H.C.; Marshall, B.; Broadley, M.R. Extraordinarily High Leaf Selenium to Sulfur Ratios Define ‘Se-accumulator’ Plants. Ann. Bot. 2007, 100, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, G.; Ortiz-Monasterio, I.; Stangoulis, J.; Graham, R. Selenium concentration in wheat grain: Is there sufficient genotypic variation to use in breeding? Plant Soil 2005, 269, 369–380. [Google Scholar] [CrossRef]
- Galinha, C.; Sánchez-Martínez, M.; Pacheco, A.M.G.; Freitas, M.d.C.; Coutinho, J.; Maçãs, B.; Almeida, A.S.; Pérez-Corona, M.T.; Madrid, Y.; Wolterbeek, H.T. Characterization of selenium-enriched wheat by agronomic biofortification. J. Food Sci. Technol. 2014, 52, 4236–4245. [Google Scholar] [CrossRef] [Green Version]
- Erenstein, O.; Jaleta, M.; Mottaleb, K.A.; Sonder, K.; Donovan, J.; Braun, H.-J. Global Trends in Wheat Production, Consumption and Trade. In Wheat Improvement; Springer: Cham, Switzerland, 2022; pp. 47–66. [Google Scholar]
- Whanger, P.D. Selenocompounds in Plants and Animals and their Biological Significance. J. Am. Coll. Nutr. 2002, 21, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Thomson, C.D. Assessment of requirements for selenium and adequacy of selenium status: A review. Eur. J. Clin. Nutr. 2004, 58, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Connorton, J.M.; Balk, J.; Miller, A.J.; Sanders, D.; Uauy, C. Biofortification of wheat grain with iron and zinc: Integrating novel genomic resources and knowledge from model crops. Front. Plant Sci. 2014, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Frittelli, A.; Botticella, E.; Palombieri, S.; Masci, S.; Celletti, S.; Fontanella, M.C.; Astolfi, S.; De Vita, P.; Volpato, M.; Sestili, F. The suppression of TdMRP3 genes reduces the phytic acid and increases the nutrient accumulation in durum wheat grain. Front. Plant Sci. 2023, 14, 46. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Selenium: Significance, and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef]
- Poblaciones, M.J.; Rodrigo, S.; Santamaría, O.; Chen, Y.; McGrath, S.P. Agronomic selenium biofortification in Triticum durum under Mediterranean conditions: From grain to cooked pasta. Food Chem. 2014, 146, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Stadlober, M.; Sager, M.; Irgolic, K.J. Effects of selenate supplemented fertilisation on the selenium level of cereals—Identification and quantification of selenium compounds by HPLC-ICP-MS. Food Chem. 2001, 73, 357–366. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Pilon-Smits, E.A.H.; Zhao, F.J.; Williams, P.N.; Meharg, A.A. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation. Trends Plant Sci. 2009, 14, 436–442. [Google Scholar] [CrossRef]
- Raina, M.; Sharma, A.; Nazir, M.; Kumari, P.; Rustagi, A.; Hami, A.; Bhau, B.S.; Zargar, S.M.; Kumar, D. Exploring the new dimensions of selenium research to understand the underlying mechanism of its uptake, translocation, and accumulation. Physiol. Plant. 2021, 171, 882–895. [Google Scholar] [CrossRef] [PubMed]
- Bouain, N.; Krouk, G.; Lacombe, B.; Rouached, H. Getting to the Root of Plant Mineral Nutrition: Combinatorial Nutrient Stresses Reveal Emergent Properties. Trends Plant Sci. 2019, 24, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium speciation from food source to metabolites: A critical review. Anal. Bioanal. Chem. 2006, 385, 1304–1323. [Google Scholar] [CrossRef] [PubMed]
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Boldrin, K.V.F.; Ávila, F.W.; Guilherme, L.R.G. Soil and foliar application of selenium in rice biofortification. J. Food Compos. Anal. 2013, 31, 238–244. [Google Scholar] [CrossRef]
- Boldrin, P.F.; de Figueiredo, M.A.; Yang, Y.; Luo, H.; Giri, S.; Hart, J.J.; Faquin, V.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Selenium promotes sulfur accumulation and plant growth in wheat (Triticum aestivum). Physiol. Plant. 2016, 158, 80–91. [Google Scholar] [CrossRef]
- Guerrero, B.; Llugany, M.; Palacios, O.; Valiente, M. Dual effects of different selenium species on wheat. Plant Physiol. Biochem. 2014, 83, 300–307. [Google Scholar] [CrossRef]
- Dong, Y.; Silbermann, M.; Speiser, A.; Forieri, I.; Linster, E.; Poschet, G.; Allboje Samami, A.; Wanatabe, M.; Sticht, C.; Teleman, A.A.; et al. Sulfur availability regulates plant growth via glucose-TOR signaling. Nat. Commun. 2017, 8, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astolfi, S.; Celletti, S.; Vigani, G.; Mimmo, T.; Cesco, S. Interaction Between Sulfur and Iron in Plants. Front. Plant Sci. 2021, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Y.; Hu, H.; Hu, J.; Xiang, M.; Yang, Q. Comparative proteomics analysis of the responses to selenium in selenium-enriched alfalfa (Medicago sativa L.) leaves. Plant Physiol. Biochem. 2021, 165, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Zeka, D.; Fetahu, S.; Rusinovci, I.; Kaul, H.P. Selenium supply affects chlorophyll concentration and biomass production of maize (Zea mays L.). Bodenkultur 2018, 69, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Zhao, Z.; Lv, C.; Zhang, Z.; Yuan, L.; Liu, X. Effects of sulfur application on selenium uptake and seed selenium speciation in soybean (Glycine max L.) grown in different soil types. Ecotoxicol. Environ. Saf. 2021, 209, 111790. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; McGrath, S.P.; Zhao, F. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef]
- Davidian, J.-C.; Kopriva, S. Regulation of sulfate uptake and assimilation—The same or not the same? Mol. Plant 2010, 3, 314–325. [Google Scholar] [CrossRef]
- Takahashi, H.; Buchner, P.; Yoshimoto, N.; Hawkesford, M.J.; Shiu, S.-H. Evolutionary Relationships and Functional Diversity of Plant Sulfate Transporters. Front. Plant Sci. 2012, 2, 119. [Google Scholar] [CrossRef] [Green Version]
- El Kassis, E.; Cathala, N.; Rouached, H.; Fourcroy, P.; Berthomieu, P.; Terry, N.; Davidian, J.-C. Characterization of a Selenate-Resistant Arabidopsis Mutant. Root Growth as a Potential Target for Selenate Toxicity. Plant Physiol. 2007, 143, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Ciaffi, M.; Paolacci, A.R.; Celletti, S.; Catarcione, G.; Kopriva, S.; Astolfi, S. Transcriptional and physiological changes in the S assimilation pathway due to single or combined S and Fe deprivation in durum wheat (Triticum durum L.) seedlings. J. Exp. Bot. 2013, 64, 1663–1675. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.Y.; Hale, B.A. Differential accumulation of Cd in durum wheat cultivars: Uptake and retranslocation as sources of variation. J. Exp. Bot. 2004, 55, 2571–2579. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.S.; Römheld, V.; Marschner, H. Role of the root apoplasm for iron acquisition by wheat plants. Plant Physiol. 1991, 97, 1302–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celletti, S.; Pii, Y.; Mimmo, T.; Cesco, S.; Astolfi, S. The characterization of the adaptive responses of durum wheat to different Fe availability highlights an optimum Fe requirement threshold. Plant Physiol. Biochem. 2016, 109, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Camerlengo, F.; Sestili, F.; Silvestri, M.; Colaprico, G.; Margiotta, B.; Ruggeri, R.; Lupi, R.; Masci, S.; Lafiandra, D. Production and molecular characterization of bread wheat lines with reduced amount of α-type gliadins. BMC Plant Biol. 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestili, F.; Pagliarello, R.; Zega, A.; Saletti, R.; Pucci, A.; Botticella, E.; Masci, S.; Tundo, S.; Moscetti, I.; Foti, S.; et al. Enhancing grain size in durum wheat using RNAi to knockdown GW2 genes. Theor. Appl. Genet. 2019, 132, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | S (mg g−1 DW) | Se (μg g−1 DW) | ||
|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | ||
| Svevo | S0 | 2.09 ± 0.09 c | 3.20 ± 0.15 c | 0 ± 0 d | 0.40 ± 0 bc |
| S10 | 2.12 ± 0.04 c | 7.34 ± 1.55 c | 0 ± 0 d | 0.64 ± 0.01 ab | |
| S50 | 6.0 ± 0.88 c | 9.60 ± 0.45 c | 1.15 ± 0.01 cd | 0.69 ± 0.09 a | |
| Tur-21 | S0 | 1.73 ± 0.01 c | 2.79 ± 0.28 c | 1.79 ± 0.37 cd | 0.65 ± 0.12 a |
| S10 | 8.39 ± 0.41 c | 5.49 ± 0.17 c | 3.09 ± 0.47 bc | 0.55 ± 0.11 abc | |
| S50 | 29.16 ± 1.53 bc | 5.39 ± 1.24 c | 1.93 ± 0.04 cd | 0.64 ± 0.07 ab | |
| Kamut | S0 | 8.79 ± 1.05 c | 2.76 ± 0.05 c | 1.70 ± 0.32 cd | 0.50 ± 0.07 abc |
| S10 | 21.47 ± 9.88 bc | 4.18 ± 0.41 c | 2.46 ± 0.45 c | 0.47 ± 0.07 abc | |
| S50 | 3.18 ± 0.51 c | 2.93 ± 0.07 c | 2.32 ± 0.03 c | 0.59 ± 0.08 abc | |
| Etrusco | S0 | 11.46 ± 3.61 c | 18.45 ± 1.34 b | 6.33 ± 2.05 a | 0.36 ± 0.01 c |
| S10 | 48.99 ± 28.82 b | 49.24 ± 6.91 a | 3.22 ± 1.24 bc | 0.67 ± 0.12 a | |
| S50 | 100.97 ± 12.45 a | 25.31 ± 5.38 b | 4.97 ± 0.29 ab | 0.66 ± 0.08 a | |
| Source of variation | |||||
| Genotype | 1.62 × 10−10 *** | 1.074 × 10−17 *** | 5.79779 × 10−11 *** | 0.146132 - | |
| Treatment | 6.91 × 10−7 *** | 1.541 × 10−8 *** | 0.416611378 - | 0.000155 *** | |
| Genotype*Treatment | 2.2 × 10−7 *** | 2.417 × 10−9 *** | 0.000473439 *** | 0.002185 ** | |
| Genotype | Treatment | S (mg g−1 DW) | Se (μg g−1 DW) | ||
|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | ||
| Svevo | L0 | 1.61 ± 0.01 c | 2.76 ± 0.09 f | 0 ± 0 d | 0 ± 0 d |
| L10 | 6.37 ± 1.20 c | 10.21 ± 0.34 cd | 1.77± 0.08 c | 0.98 ± 0.12 c | |
| L50 | 1.76 ± 0.23 c | 3.22 ± 0.66 f | 3.036 ± 0.10 bc | 4.09 ± 0.18 a | |
| Tur-21 | L0 | 1.73 ± 0.04 c | 5.61 ± 0.89 ef | 2.55 ± 0.04 c | 0.81 ± 0.046 c |
| L10 | 10.09 ± 1.46 c | 3.63 ± 1.03 ef | 2.95 ± 0.16 bc | 1.03 ± 0.13 c | |
| L50 | 10.01 ± 2.78 c | 7.41 ± 0.57 de | 2.56 ± 0.69 c | 3.17 ± 0.13 b | |
| Kamut | L0 | 2.19 ± 0.03 c | 11.75 ± 3.06 c | 1.34 ± 0.02 cd | 0.58 ± 0.01 c |
| L10 | 3.29 ± 1.32 c | 2.74 ± 0.42 f | 2.59 ± 0.23 c | 1.10 ± 0.16 c | |
| L50 | 5.25 ± 0.53 c | 3.55 ± 0.25 ef | 4.48 ± 0.39 b | 3.45 ± 0.18 b | |
| Etrusco | L0 | 41.11 ± 3.09 b | 11.93 ± 2.18 c | 7.064 ± 1.68 a | 1.35 ± 0.29 c |
| L10 | 76.26 ± 20.64 a | 18.40 ± 0.99 b | 2.50 ± 0.64 c | 1.15 ± 0.18 c | |
| L50 | 98.76 ± 29.23 a | 23.45 ± 2.10 a | 6.99 ± 0.30 a | 3.68 ± 0.43ab | |
| Source of variation | |||||
| Genotype | 2.06 × 10−13 *** | 1.3555 × 10−16 *** | 3.08 × 10−12 *** | 0.002313 ** | |
| Treatment | 0.001401 ** | 0.06291953 - | 1.11 × 10−7 *** | 1.44 × 10−21 *** | |
| Genotype*Treatment | 0.00158 ** | 1.83 × 10−11 *** | 6.97 × 10−9 *** | 3.04 × 10−6 *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppa, E.; Celletti, S.; Sestili, F.; Mimmo, T.; Garcia Molina, M.D.; Cesco, S.; Astolfi, S. Interaction between Sulfate and Selenate in Tetraploid Wheat (Triticum turgidum L.) Genotypes. Int. J. Mol. Sci. 2023, 24, 5443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065443
Coppa E, Celletti S, Sestili F, Mimmo T, Garcia Molina MD, Cesco S, Astolfi S. Interaction between Sulfate and Selenate in Tetraploid Wheat (Triticum turgidum L.) Genotypes. International Journal of Molecular Sciences. 2023; 24(6):5443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065443
Chicago/Turabian StyleCoppa, Eleonora, Silvia Celletti, Francesco Sestili, Tanja Mimmo, Maria Dolores Garcia Molina, Stefano Cesco, and Stefania Astolfi. 2023. "Interaction between Sulfate and Selenate in Tetraploid Wheat (Triticum turgidum L.) Genotypes" International Journal of Molecular Sciences 24, no. 6: 5443. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065443