

IFN-Inducible SerpinA5 Triggers Antiviral Immunity by Regulating STAT1 Phosphorylation and Nuclear Translocation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
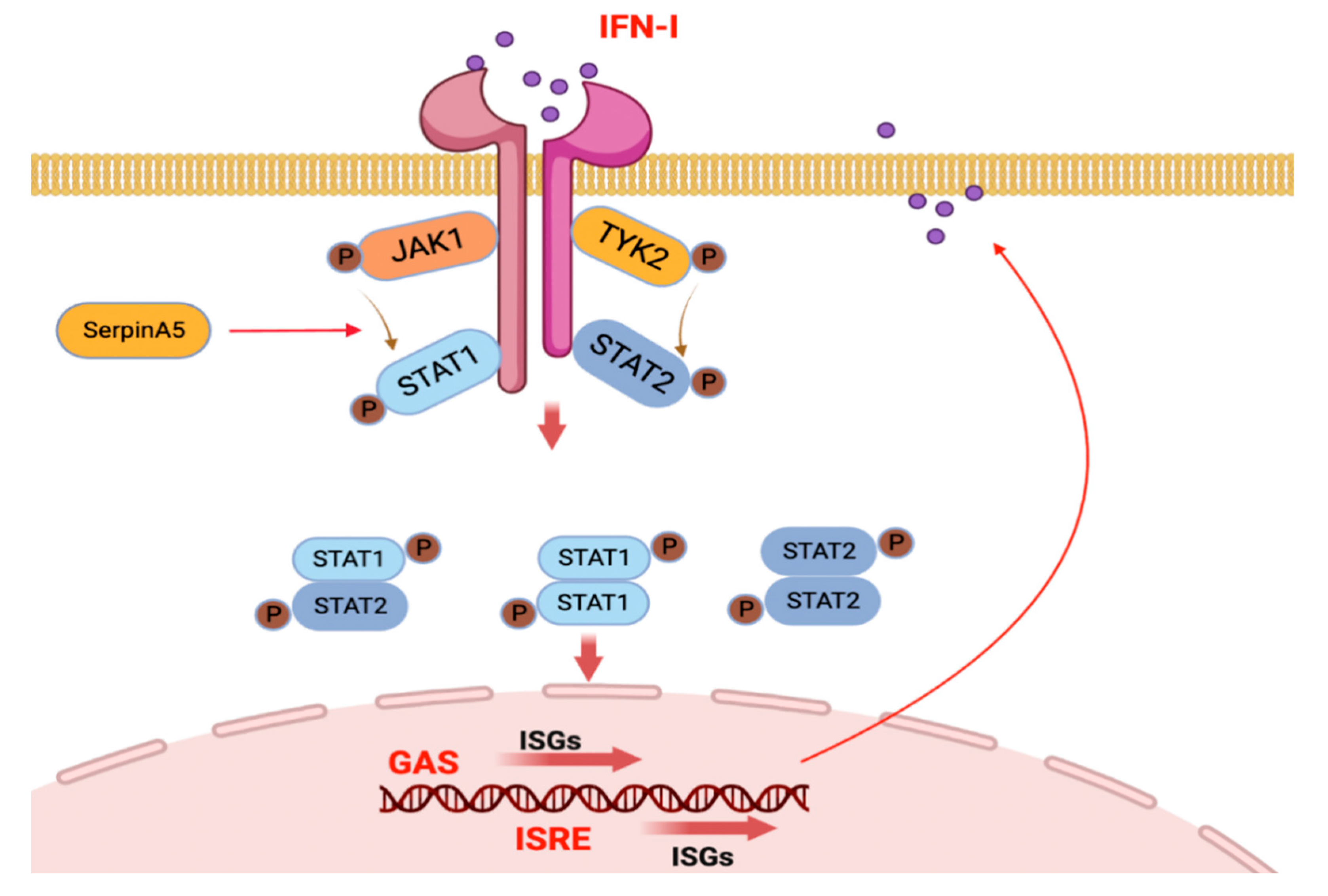
Abstract
:1. Introduction
2. Results
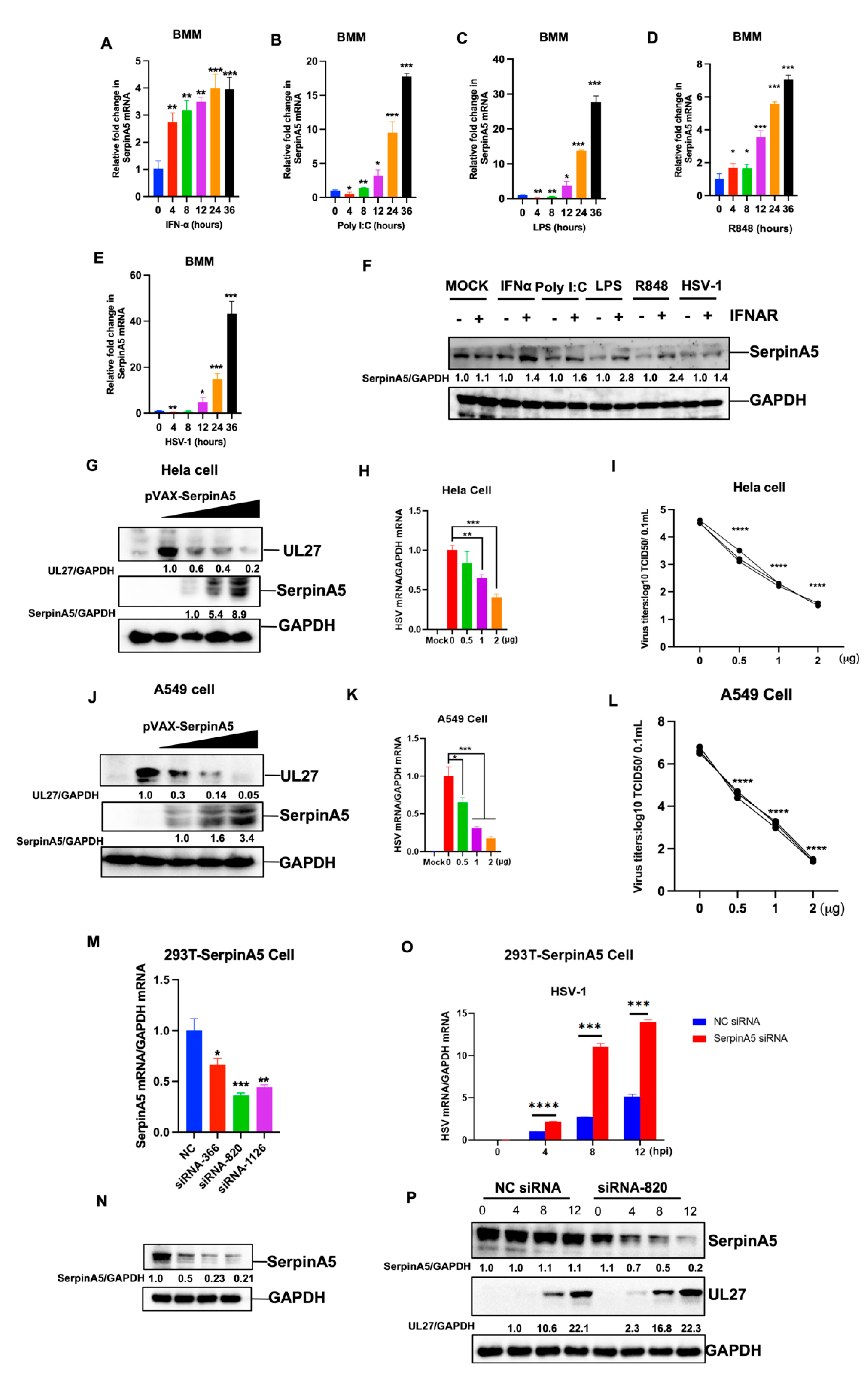
2.1. SerpinA5 Is an IFN-Stimulated Gene with Antiviral Function
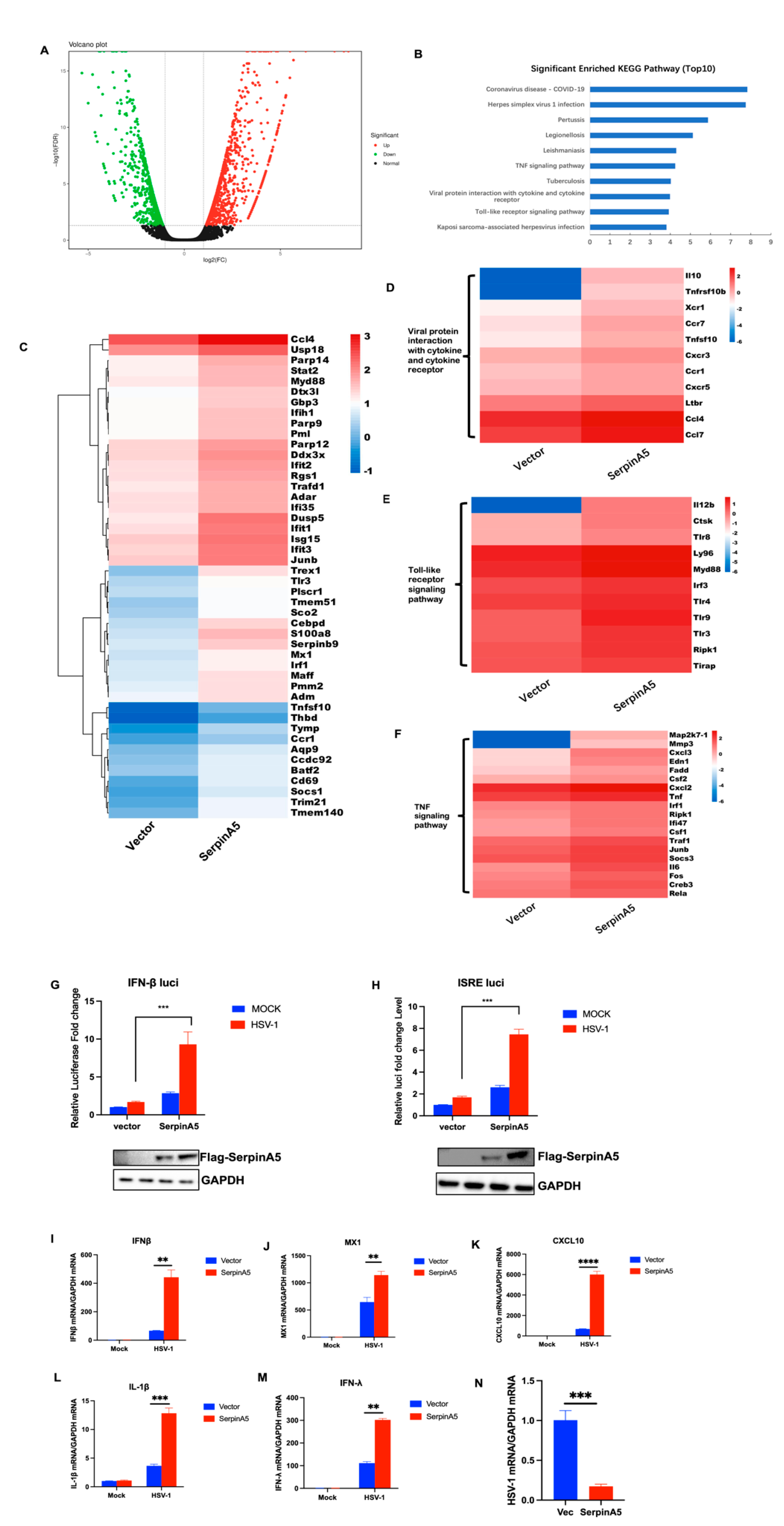
2.2. SerpinA5 Can Play the Antiviral Function through Modulating IFN Signaling Pathways
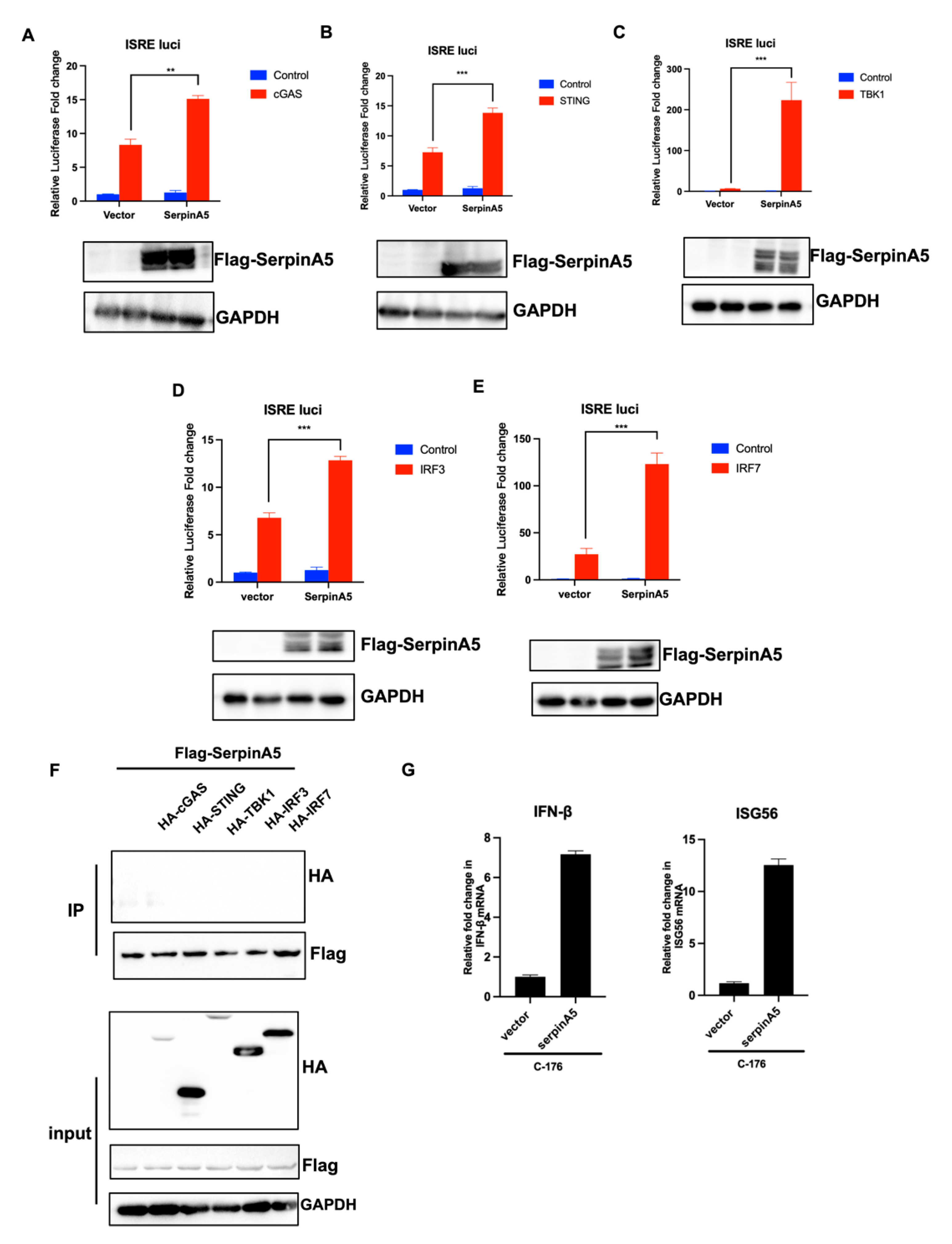
2.3. SerpinA5 Activated IFN Production Independent of cGAS-STING Signaling Pathway
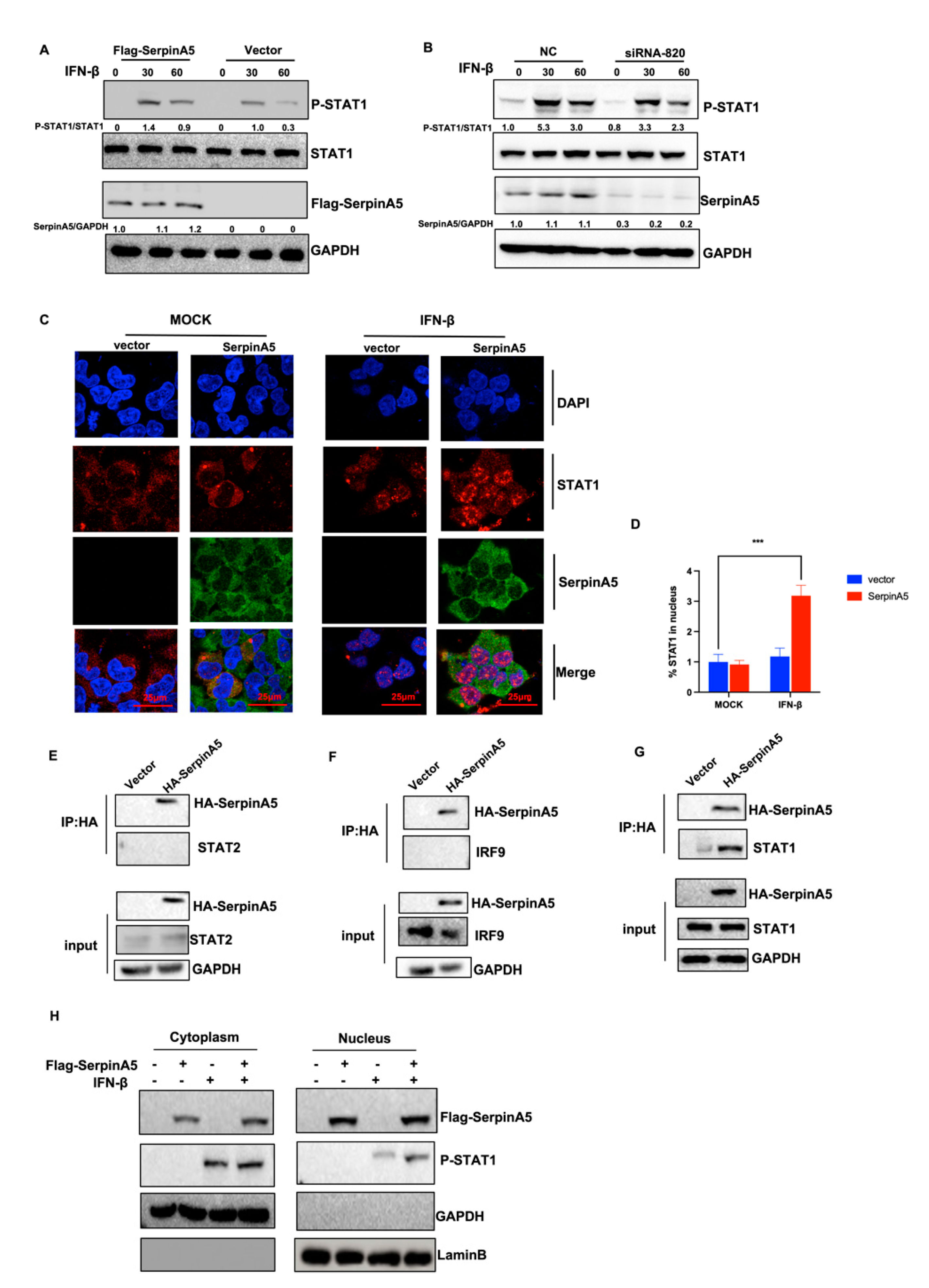
2.4. SerpinA5 Induced Antiviral Innate Immunity by Promoting STAT1 Phosphorylation and Nuclear Translocation
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Plasmid Constructs
4.3. Transient Transfection and siRNA Experiment
4.4. Western Blotting Analysis
4.5. Quantitative Real-Time PCR
4.6. RNA-Seq Library Preparation, Sequencing, and Data Processing
4.7. TCID50
4.8. Co-Iimmunoprecipitation
4.9. Luciferase Assay
4.10. Confocal Microscopy
4.11. Quantification and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akira, S. Pathogen recognition by innate immunity and its signaling. Proc. Jpn. Acad. Ser. B 2009, 85, 143–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [Green Version]
- Bancerek, J.; Poss, Z.C.; Steinparzer, I.; Sedlyarov, V.; Pfaffenwimmer, T.; Mikulic, I.; Dölken, L.; Strobl, B.; Müller, M.; Taatjes, D.J.; et al. CDK8 Kinase Phosphorylates Transcription Factor STAT1 to Selectively Regulate the Interferon Response. Immunity 2013, 38, 250–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Gao, J.-S.; Guan, Y.-J.; McLane, K.E.; Yuan, Z.-L.; Ramratnam, B.; Chin, Y.E. Acetylation-Dependent Signal Transduction for Type I Interferon Receptor. Cell 2007, 131, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, J.; Wang, C.; Liu, Y.; Li, M.; Li, Y.; Li, R.; Han, Z.; Wang, J.; Chen, L.; et al. Kynurenine-3-monooxygenase (KMO) broadly inhibits viral infections via triggering NMDAR/Ca2+ influx and CaMKII/ IRF3-mediated IFN-beta production. PLoS Pathog. 2022, 18, e1010366. [Google Scholar] [CrossRef]
- Zhao, J.; Li, R.; Li, Y.; Chen, J.; Feng, F.; Sun, C. Broadly Antiviral Activities of TAP1 through Activating the TBK1-IRF3-Mediated Type I Interferon Production. Int. J. Mol. Sci. 2021, 22, 4668. [Google Scholar] [CrossRef]
- Wu, T.; Ma, F.; Ma, X.; Jia, W.; Pan, E.; Cheng, G.; Chen, L.; Sun, C. Regulating Innate and Adaptive Immunity for Controlling SIV Infection by 25-Hydroxycholesterol. Front. Immunol. 2018, 9, 2686. [Google Scholar] [CrossRef] [Green Version]
- Michalska, A.; Blaszczyk, K.; Wesoly, J.; Bluyssen, H.A.R. A Positive Feedback Amplifier Circuit That Regulates Interferon (IFN)-Stimulated Gene Expression and Controls Type I and Type II IFN Responses. Front. Immunol. 2018, 9, 1135. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Geiger, M. Cell penetrating SERPINA5 (ProteinC inhibitor, PCI): More questions than answers. Semin. Cell Dev. Biol. 2017, 62, 187–193. [Google Scholar] [CrossRef]
- Heit, C.; Jackson, B.C.; McAndrews, M.; Wright, M.W.; Thompson, D.C.; Silverman, G.A.; Nebert, D.W. Update of the human and mouse SERPIN gene superfamily. Hum. Genom. 2013, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Barak, Y.; Lamed, R.; Bayer, E.A.; Morrison, M. The functional repertoire of prokaryote cellulosomes includes the serpin superfamily of serine proteinase inhibitors. Mol. Microbiol. 2006, 60, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.K.; Chung, K.-W.; Yang, S.K.; Park, M.J.; Min, H.S.; Kim, S.W.; Kang, H.S. DNA methylation of MAPK signal-inhibiting genes in papillary thyroid carcinoma. Anticancer Res. 2013, 33, 4833–4839. [Google Scholar]
- Yang, H.; Wahlmüller, F.C.; Sarg, B.; Furtmüller, M.; Geiger, M. A+-Helix of Protein C Inhibitor (PCI) Is a Cell-penetrating Peptide That Mediates Cell Membrane Permeation of PCI. J. Biol. Chem. 2015, 290, 3081–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmström, E.; Mörgelin, M.; Malmsten, M.; Johansson, L.; Norrby-Teglund, A.; Shannon, O.; Schmidtchen, A.; Meijers, J.C.M.; Herwald, H. Protein C Inhibitor—A Novel Antimicrobial Agent. PLoS Pathog. 2009, 5, e1000698. [Google Scholar] [CrossRef] [Green Version]
- Van Raemdonck, G.; Zegels, G.; Coen, E.; Vuylsteke, B.; Jennes, W.; Van Ostade, X. Increased Serpin A5 levels in the cervicovaginal fluid of HIV-1 exposed seronegatives suggest that a subtle balance between serine proteases and their inhibitors may determine susceptibility to HIV-1 infection. Virology 2014, 458–459, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [Green Version]
- Au-Yeung, N.; Mandhana, R.; Horvath, C.M. Transcriptional regulation by STAT1 and STAT2 in the interferon JAK-STAT pathway. Jak-Stat 2013, 2, e23931. [Google Scholar] [CrossRef] [Green Version]
- Cheon, H.; Holvey-Bates, E.G.; Schoggins, J.W.; Forster, S.; Hertzog, P.; Imanaka, N.; Rice, C.M.; Jackson, M.W.; Junk, D.J.; Stark, G.R. IFNβ-dependent increases in STAT1, STAT2, and IRF9 mediate resistance to viruses and DNA damage. EMBO J. 2013, 32, 2751–2763. [Google Scholar] [CrossRef] [Green Version]
- Blaszczyk, K.; Olejnik, A.; Nowicka, H.; Ozgyin, L.; Chen, Y.-L.; Chmielewski, S.; Kostyrko, K.; Wesoly, J.; Balint, B.L.; Lee, C.-K.; et al. STAT2/IRF9 directs a prolonged ISGF3-like transcriptional response and antiviral activity in the absence of STAT1. Biochem. J. 2015, 466, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, O.H.; Heinzel, T. Phosphorylation–acetylation switch in the regulation of STAT1 signaling. Mol. Cell. Endocrinol. 2010, 315, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Song, Q.; Huang, W.; Lin, Y.; Wang, X.; Wang, C.; Willard, B.; Zhao, C.; Nan, J.; Holvey-Bates, E.; et al. A virus-induced conformational switch of STAT1-STAT2 dimers boosts antiviral defenses. Cell Res. 2020, 31, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Heeb, M.J.; España, F.; Geiger, M.; Collen, D.; Stump, D.C.; Griffin, J.H. Immunological identity of heparin-dependent plasma and urinary protein C inhibitor and plasminogen activator inhibitor-3. J. Biol. Chem. 1987, 262, 15813–15816. [Google Scholar] [CrossRef]
- Papareddy, P.; Kalle, M.; Bhongir, R.K.; Mörgelin, M.; Malmsten, M.; Schmidtchen, A. Antimicrobial Effects of Helix D-derived Peptides of Human Antithrombin III. J. Biol. Chem. 2014, 289, 29790–29800. [Google Scholar] [CrossRef] [Green Version]
- El-Hefnawy, S.M.; Kasemy, Z.A.; Eid, H.A.; Elmadbouh, I.; Mostafa, R.G.; Omar, T.A.; Kasem, H.E.; Ghonaim, E.M.; Ghonaim, M.M.; Salehet, A.A. Potential impact of serpin peptidase inhibitor clade (A) member 4 SERPINA4 (rs2093266) and SERPINA5 (rs1955656) genetic variants on COVID-19 induced acute kidney injury. Meta. Gene. 2022, 32, 101023. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, X.; Jin, J.-O.; Lu, H.; Tan, Z. Recent 5-year Findings and Technological Advances in the Proteomic Study of HIV-associated Disorders. Genom. Proteom. Bioinform. 2017, 15, 110–120. [Google Scholar] [CrossRef]
- Zhang, X.; Groen, K.; Morten, B.C.; Reinhardt, L.S.; Campbell, H.G.; Braithwaite, A.W.; Bourdon, J.-C.; Avery-Kiejda, K.A. Effect of p53 and its N-terminally truncated isoform, Delta40p53, on breast cancer migration and invasion. Mol. Oncol. 2022, 16, 447–465. [Google Scholar] [CrossRef]
- Baumgärtner, P.; Geiger, M.; Zieseniss, S.; Malleier, J.; Huntington, J.A.; Hochrainer, K.; Bielek, E.; Stoeckelhuber, M.; Lauber, K.; Scherfeld, D.; et al. Phosphatidylethanolamine critically supports internalization of cell-penetrating protein C inhibitor. J. Cell Biol. 2007, 179, 793–804. [Google Scholar] [CrossRef]
- Fish, E.N.; Platanias, L.C. Interferon Receptor Signaling in Malignancy: A Network of Cellular Pathways Defining Biological Outcomes. Mol. Cancer Res. 2014, 12, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Levy, D.E.; Darnell, J.E., Jr. STATs: Transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 2002, 3, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, S.; Takeuchi, K.; Chihara, K.; Honjoh, C.; Kato, Y.; Yoshiki, H.; Hotta, H.; Sada, K. STAT1 is essential for the inhibition of hepatitis C virus replication by interferon-lambda but not by interferon-alpha. Sci. Rep. 2016, 6, 38336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, M.; Pardo–Saganta, A.; Alvarez–Asiain, L.; Di Scala, M.; Qian, C.; Prieto, J.; Avila, M.A. Nuclear α1-Antichymotrypsin Promotes Chromatin Condensation and Inhibits Proliferation of Human Hepatocellular Carcinoma Cells. Gastroenterology 2013, 144, 818–828.e4. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; He, Y.; Zhao, J.; Luo, K.; Wen, Z.; Zhang, Y.; Li, M.; Cui, Y.; Liu, Z.; Wang, C.; et al. Exacerbated AIDS Progression by PD-1 Blockade during Therapeutic Vaccination in Chronically Simian Immunodeficiency Virus-Infected Rhesus Macaques after Interruption of Antiretroviral Therapy. J. Virol. 2022, 96, e0178521. [Google Scholar] [CrossRef]
- Feng, F.; Hao, H.; Zhao, J.; Li, Y.; Zhang, Y.; Li, R.; Wen, Z.; Wu, C.; Li, M.; Li, P.; et al. Shell-mediated phagocytosis to reshape viral-vectored vaccine-induced immunity. Biomaterials 2021, 276, 121062. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Liu, Y.; Liu, X.; Zhao, J.; Lang, B.; Wu, F.; Wen, Z.; Sun, C. IFN-Inducible SerpinA5 Triggers Antiviral Immunity by Regulating STAT1 Phosphorylation and Nuclear Translocation. Int. J. Mol. Sci. 2023, 24, 5458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065458
Wang C, Liu Y, Liu X, Zhao J, Lang B, Wu F, Wen Z, Sun C. IFN-Inducible SerpinA5 Triggers Antiviral Immunity by Regulating STAT1 Phosphorylation and Nuclear Translocation. International Journal of Molecular Sciences. 2023; 24(6):5458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065458
Chicago/Turabian StyleWang, Congcong, Yajie Liu, Xinglai Liu, Jin Zhao, Bing Lang, Fan Wu, Ziyu Wen, and Caijun Sun. 2023. "IFN-Inducible SerpinA5 Triggers Antiviral Immunity by Regulating STAT1 Phosphorylation and Nuclear Translocation" International Journal of Molecular Sciences 24, no. 6: 5458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065458