
The Signaling Pathway of the ADP Receptor P2Y12 in the Immune System: Recent Discoveries and New Challenges
 , and
, and
Abstract
:1. Introduction
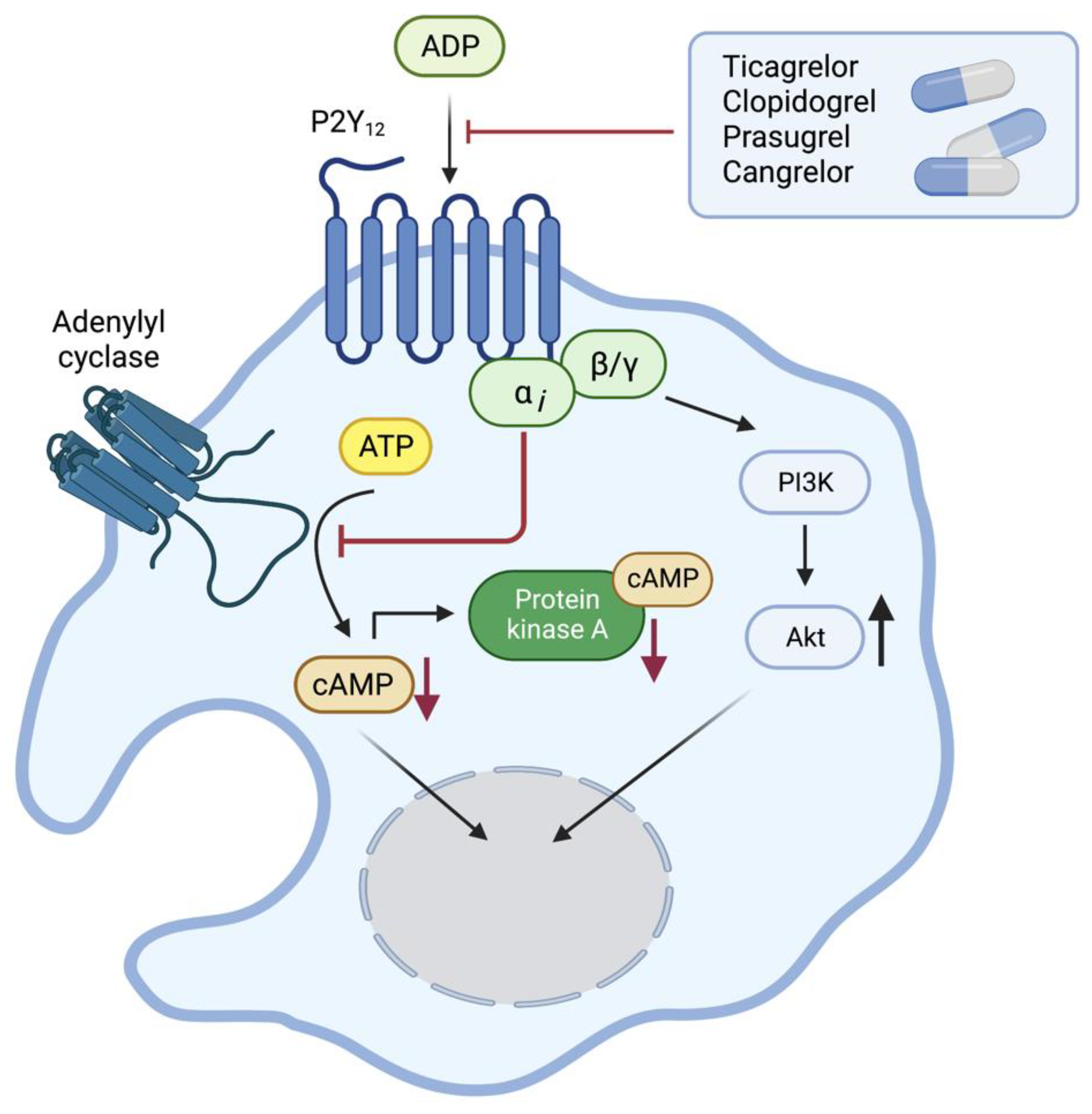
2. The ADP Receptor P2Y12
3. Expression of the ADP Receptor P2Y12 in the Immune System
3.1. Platelets
3.2. Monocytes and Macrophages
3.3. T Lymphocytes
3.4. Neutrophils and Eosinophils
3.5. Dendritic Cells
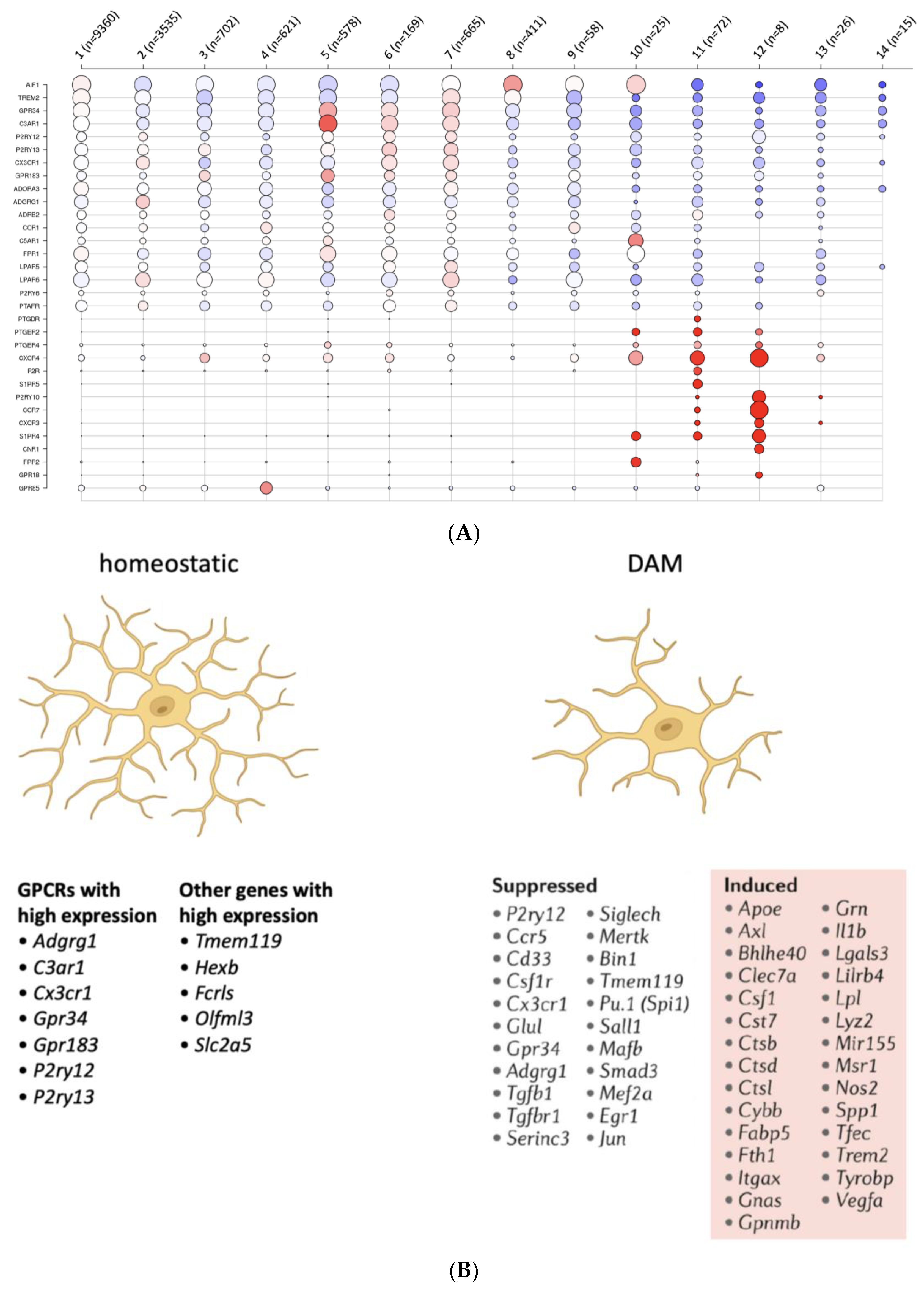
3.6. Microglia
3.7. Other Cells Relevant to Inflammation
4. Drugs Targeting P2Y12
5. Challenges in Studying the Receptor P2Y12
5.1. Reliable Antibody
5.2. Antagonist Specificity
5.3. Cell Purity
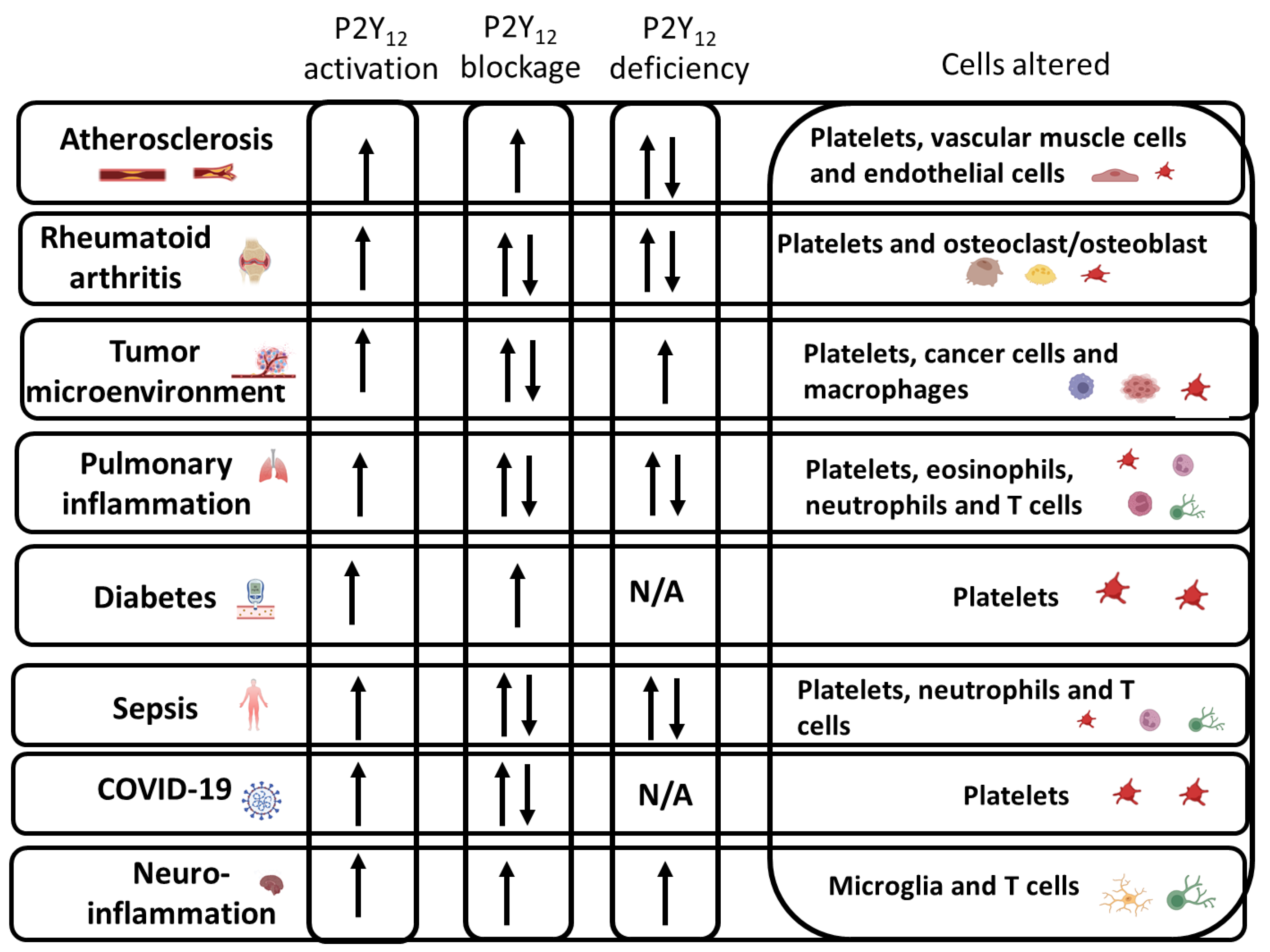
6. P2Y12 Activation in the Immune System during Inflammatory Conditions
6.1. Atherosclerosis
6.2. Rheumatoid Arthritis
6.3. Tumor Microenvironment
6.4. Inflammation in Diabetes
6.5. Pulmonary Inflammation and Asthma
6.6. Sepsis
6.7. COVID-19
6.8. Neuroinflammation
7. Sex-Related Differences in P2Y12 Activation
8. Age-Related Differences in P2Y12 Activation
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Murugappa, S.; Kunapuli, S.P. The role of ADP receptors in platelet function. Front. Biosci. 2006, 11, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Kahner, B.N.; Shankar, H.; Murugappan, S.; Prasad, G.L.; Kunapuli, S.P. Nucleotide receptor signaling in platelets. J. Thromb. Haemost. 2006, 4, 2317–2326. [Google Scholar] [CrossRef]
- Gomez Morillas, A.; Besson, V.C.; Lerouet, D. Microglia and Neuroinflammation: What Place for P2RY12? Int. J. Mol. Sci. 2021, 22, 1636. [Google Scholar] [CrossRef]
- Ben Addi, A.; Cammarata, D.; Conley, P.B.; Boeynaems, J.M.; Robaye, B. Role of the P2Y12 Receptor in the Modulation of Murine Dendritic Cell Function by ADP. J. Immunol. 2010, 185, 5900–5906. [Google Scholar] [CrossRef] [Green Version]
- Liverani, E.; Rico, M.C.; Garcia, A.E.; Kilpatrick, L.E.; Kunapuli, S.P. Prasugrel Metabolites Inhibit Neutrophil Functions. J. Pharmacol. Exp. Ther. 2013, 344, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Micklewright, J.J.; A Layhadi, J.; Fountain, S.J. P2Y12 receptor modulation of ADP-evoked intracellular Ca2+ signalling in THP-1 human monocytic cells. Br. J. Pharmacol. 2018, 175, 2483–2491. [Google Scholar] [CrossRef] [Green Version]
- Vemulapalli, H.; Albayati, S.; Patwa, V.C.; Tilley, D.G.; Tsygankov, A.Y.; Liverani, E. ADP exerts P2Y12-dependent and P2Y12 -independent effects on primary human T cell responses to stimulation. J. Cell Commun. Signal. 2020, 14, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jacobsen, S.E.W.; Bengtsson, A.; Erlinge, D. P2 receptor mRNA expression profiles in human lymphocytes, monocytes and CD34+ stem and progenitor cells. BMC Immunol. 2004, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlović, N.; Kopsida, M.; Gerwins, P.; Heindryckx, F. Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype. Int. J. Mol. Sci. 2020, 21, 8177. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Gao, M.; Luo, Y.; Gui, R.; Ji, H. Long non-coding RNA metallothionein 1 pseudogene 3 promotes p2y12 expression by sponging miR-126 to activate platelet in diabetic animal model. Platelets 2019, 30, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Albayati, S.; Vemulapalli, H.; Tsygankov, A.Y.; Liverani, E. P2Y12 antagonism results in altered interactions between platelets and regulatory T cells during sepsis. J. Leukoc. Biol. 2021, 110, 141–153. [Google Scholar] [CrossRef]
- Liverani, E.; Rico, M.C.; Tsygankov, A.Y.; Kilpatrick, L.E.; Kunapuli, S.P. P2Y12 Receptor Modulates Sepsis-Induced Inflammation. Arter. Thromb. Vasc. Biol. 2016, 36, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.-H.; Trinh, H.K.T.; Liu, J.-N.; Pham, L.D.; Park, S.M.; Park, H.-S.; Shin, Y.S. P2Y12 antagonist attenuates eosinophilic inflammation and airway hyperresponsiveness in a mouse model of asthma. J. Cell. Mol. Med. 2016, 20, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Trinh, H.K.T.; Nguyen, T.V.T.; Choi, Y.; Park, H.-S.; Shin, Y.S. The synergistic effects of clopidogrel with montelukast may be beneficial for asthma treatment. J. Cell. Mol. Med. 2019, 23, 3441–3450. [Google Scholar] [CrossRef]
- Garcia, A.E.; Mada, S.R.; Rico, M.C.; Cadena, R.A.D.; Kunapuli, S.P. Clopidogrel, a P2Y12 Receptor Antagonist, Potentiates the Inflammatory Response in a Rat Model of Peptidoglycan Polysaccharide-Induced Arthritis. PLoS ONE 2011, 6, e26035. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Floyd, D.H.; Hughes, A.; Xiang, J.; Schneider, J.G.; Uluckan, O.; Heller, E.; Deng, H.; Zou, W.; Craft, C.S.; et al. The ADP receptor P2RY12 regulates osteoclast function and pathologic bone remodeling. J. Clin. Investig. 2012, 122, 3579–3592. [Google Scholar] [CrossRef]
- Lou, N.; Takano, T.; Pei, Y.; Xavier, A.L.; Goldman, S.A.; Nedergaard, M. Purinergic receptor P2RY12-dependent microglial closure of the injured blood–brain barrier. Proc. Natl. Acad. Sci. USA 2016, 113, 1074–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, Z.; Hu, X.; Su, Q.; He, C.; Liu, J.; Ren, H.; Qian, M.; Liu, J.; Cui, S.; et al. Knockout of P2Y12 aggravates experimental autoimmune encephalomyelitis in mice via increasing of IL-23 production and Th17 cell differentiation by dendritic cells. Brain Behav. Immun. 2017, 62, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Jantzen, H.M.; Milstone, D.S.; Gousset, L.; Conley, P.B.; Mortensen, R.M. Impaired activation of murine platelets lacking G alpha(i2). J. Clin. Investig. 2001, 108, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kunapuli, S.P. P2Y12 receptor in platelet activation. Platelets 2011, 22, 54–58. [Google Scholar] [CrossRef]
- Quinton, T.M.; Murugappan, S.; Kim, S.; Jin, J.; Kunapuli, S.P. Different G protein-coupled signaling pathways are involved in α granule release from human platelets. J. Thromb. Haemost. 2004, 2, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Savi, P.; Zachayus, J.-L.; Delesque-Touchard, N.; Labouret, C.; Hervé, C.; Uzabiaga, M.-F.; Pereillo, J.-M.; Culouscou, J.-M.; Bono, F.; Ferrara, P.; et al. The active metabolite of Clopidogrel disrupts P2Y12 receptor oligomers and partitions them out of lipid rafts. Proc. Natl. Acad. Sci. USA 2006, 103, 11069–11074. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Katz, A.; Simon, M.I. Activation of phospholipase C beta 2 by the alpha and beta gamma subunits of trimeric GTP-binding protein. Proc. Natl. Acad. Sci. USA 1993, 90, 5297–5301. [Google Scholar] [CrossRef] [Green Version]
- Mundell, S.J.; Barton, J.F.; Mayo-Martin, M.B.; Hardy, A.R.; Poole, A.W. Rapid resensitization of purinergic receptor function in human platelets. J. Thromb. Haemost. 2008, 6, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Hardy, A.R.; Conley, P.B.; Luo, J.; Benovic, J.L.; Poole, A.W.; Mundell, S.J. P2Y1 and P2Y12 receptors for ADP desensitize by distinct kinase-dependent mechanisms. Blood 2005, 105, 3552–3560. [Google Scholar] [CrossRef]
- Quinton, T.M.; Kim, S.; Jin, J.; Kunapuli, S.P. Lipid rafts are required in Galpha(i) signaling downstream of the P2Y12 receptor during ADP-mediated platelet activation. J. Thromb. Haemost. 2005, 3, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Nagy, B., Jr.; Jin, J.; Ashby, B.; Reilly, M.P.; Kunapuli, S.P. Contribution of the P2Y12 receptor-mediated pathway to platelet hyperreactivity in hypercholesterolemia. J. Thromb. Haemost. 2011, 9, 810–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, O.; El Ghannudi, S.; Hess, S.; Reydel, A.; Crimizade, U.; Jesel, L.; Radulescu, B.; Wiesel, M.L.; Gachet, C.; Ohlmann, P. The extent of P2Y12 inhibition by clopidogrel in diabetes mellitus patients with acute coronary syndrome is not related to glycaemic control: Roles of white blood cell count and body weight. Thromb. Haemost. 2012, 108, 338–348. [Google Scholar] [CrossRef]
- Remijn, J.A.; Wu, Y.-P.; Jeninga, E.H.; Ijsseldijk, M.J.; van Willigen, G.; de Groot, P.G.; Sixma, J.J.; Nurden, A.T.; Nurden, P. Role of ADP Receptor P2Y12 in Platelet Adhesion and Thrombus Formation in Flowing Blood. Arter. Thromb. Vasc. Biol. 2002, 22, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Goto, S.; Tamura, N.; Eto, K.; Ikeda, Y.; Handa, S. Functional significance of adenosine 5’-diphosphate receptor (P2Y(12)) in platelet activation initiated by binding of von Willebrand factor to platelet GP Ibalpha induced by conditions of high shear rate. Circulation 2002, 105, 2531–2536. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.A.; Moake, J.L.; McIntire, L.V. Blockade of adenosine diphosphate receptors P2Y12 and P2Y1 is required to inhibit platelet aggregation in whole blood under flow. Blood 2001, 98, 3340–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nergiz-Unal, R.; Cosemans, J.M.E.M.; Feijge, M.A.H.; Van Der Meijden, P.E.J.; Storey, R.F.; Van Giezen, J.J.J.; Egbrink, M.G.A.O.; Heemskerk, J.W.M.; Kuijpers, M.J.E. Stabilizing Role of Platelet P2Y12 Receptors in Shear-Dependent Thrombus Formation on Ruptured Plaques. PLoS ONE 2010, 5, e10130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendolicchio, G.L.; Zavalloni, D.; Bacci, M.; Corrada, E.; Marconi, M.; Lodigiani, C.; Presbitero, P.; Rota, L.; Ruggeri, Z.M. Variable effect of P2Y12 inhibition on platelet thrombus volume in flowing blood. J. Thromb. Haemost. 2010, 9, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, M.; Gachet, C. ADP Receptors and Clinical Bleeding Disorders. Arter. Thromb. Vasc. Biol. 1999, 19, 2281–2285. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, M.; Lecchi, A.; Lombardi, R.; Gachet, C.; Zighetti, M.L. Platelets from a patient heterozygous for the defect of P2CYC receptors for ADP have a secretion defect despite normal thromboxane A2 production and normal granule stores: Further evidence that some cases of platelet ‘primary secretion defect’ are heterozygous for a defect of P2CYC receptors. Arter. Thromb. Vasc. Biol. 2000, 20, E101–E106. [Google Scholar]
- Shah, S.A.; Page, C.P.; Pitchford, S.C. Platelet–Eosinophil Interactions as a Potential Therapeutic Target in Allergic Inflammation and Asthma. Front. Med. 2017, 4, 129. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.R.; Outteridge, S.N.; Ajjan, R.A.; Phoenix, F.; Sangha, G.K.; Faulkner, R.E.; Ecob, R.; Judge, H.M.; Khan, H.; West, L.E.; et al. Platelet P2Y12 Inhibitors Reduce Systemic Inflammation and Its Prothrombotic Effects in an Experimental Human Model. Arter. Thromb. Vasc. Biol. 2015, 35, 2562–2570. [Google Scholar] [CrossRef] [Green Version]
- Schedel, A.; Rolf, N. Genome-Wide Platelet RNA Profiling in Clinical Samples. Methods Mol. Biol. 2009, 496, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Gachet, C. P2Y12 receptors in platelets and other hematopoietic and non-hematopoietic cells. Purinergic Signal. 2012, 8, 609–619. [Google Scholar] [CrossRef]
- Foster, C.J.; Prosser, D.M.; Agans, J.M.; Zhai, Y.; Smith, M.D.; Lachowicz, J.E.; Zhang, F.L.; Gustafson, E.; Monsma, F.J., Jr.; Wiekowski, M.T.; et al. Molecular identification and characterization of the platelet ADP receptor targeted by thienopyridine antithrombotic drugs. J. Clin. Investig. 2001, 107, 1591–1598. [Google Scholar] [CrossRef]
- Tozaki-Saitoh, H.; Miyata, H.; Yamashita, T.; Matsushita, K.; Tsuda, M.; Inoue, K. P2Y12 receptors in primary microglia activate nuclear factor of activated T-cell signaling to induce C-C chemokine 3 expression. J. Neurochem. 2017, 141, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsawa, K.; Irino, Y.; Nakamura, Y.; Akazawa, C.; Inoue, K.; Kohsaka, S. Involvement of P2X4 and P2Y12 receptors in ATP-induced microglial chemotaxis. Glia 2007, 55, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Siegel, P.M.; Sander, L.; Fricke, A.; Stamm, J.; Wang, X.; Sharma, P.; Bassler, N.; Ying, Y.-L.; Olivier, C.B.; Eisenhardt, S.U.; et al. P2Y12 receptor blockers are anti-inflammatory drugs inhibiting both circulating monocytes and macrophages including THP-1 cells. Sci. Rep. 2021, 11, 17459. [Google Scholar] [CrossRef]
- Moore, C.S.; Ase, A.R.; Kinsara, A.; Rao, V.T.; Michell-Robinson, M.; Leong, S.Y.; Butovsky, O.; Ludwin, S.K.; Séguéla, P.; Bar-Or, A.; et al. P2Y12 expression and function in alternatively activated human microglia. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, e80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloss, L.; Dollt, C.; Schledzewski, K.; Krewer, A.; Melchers, S.; Manta, C.; Sticht, C.; de la Torre, C.; Utikal, J.; Umansky, V.; et al. ADP secreted by dying melanoma cells mediates chemotaxis and chemokine secretion of macrophages via the purinergic receptor P2Y12. Cell Death Dis. 2019, 10, 760. [Google Scholar] [CrossRef] [Green Version]
- Isfort, K.; Ebert, F.; Bornhorst, J.; Sargin, S.; Kardakaris, R.; Pasparakis, M.; Bähler, M.; Schwerdtle, T.; Schwab, A.; Hanley, P.J. Real-time Imaging Reveals That P2Y2 and P2Y12 Receptor Agonists Are Not Chemoattractants and Macrophage Chemotaxis to Complement C5a Is Phosphatidylinositol 3-Kinase (PI3K)- and p38 Mitogen-activated Protein Kinase (MAPK)-independent. J. Biol. Chem. 2011, 286, 44776–44787. [Google Scholar] [CrossRef] [Green Version]
- Muniz, V.S.; Baptista-Dos-Reis, R.; Benjamim, C.F.; Mata-Santos, H.A.; Pyrrho, A.S.; Strauch, M.A.; Melo, P.A.; Vicentino, A.R.R.; Silva-Paiva, J.; Bandeira-Melo, C.; et al. Purinergic P2Y12 Receptor Activation in Eosinophils and the Schistosomal Host Response. PLoS ONE 2015, 10, e0139805. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Zhou, J.; Gao, Y.; Lai, W.; Yang, C.; Cai, Y.; Chen, S.; Du, C. Critical Role of P2Y12 Receptor in Regulation of Th17 Differentiation and Experimental Autoimmune Encephalomyelitis Pathogenesis. J. Immunol. 2017, 199, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.; Peebles, R.S., Jr. Prostacyclin Regulation of Allergic Inflammation. Biomedicines 2022, 10, 2862. [Google Scholar] [CrossRef]
- Harada, K.; Matsumoto, Y.; Umemura, K. Adenosine Diphosphate Receptor P2Y12-Mediated Migration of Host Smooth Muscle-Like Cells and Leukocytes in the Development of Transplant Arteriosclerosis. Transplantation 2011, 92, 148–154. [Google Scholar] [CrossRef]
- Aronoff, D.M.; Peres, C.M.; Serezani, C.H.; Ballinger, M.N.; Carstens, J.K.; Coleman, N.; Moore, B.B.; Peebles, R.S.; Faccioli, L.H.; Peters-Golden, M. Synthetic Prostacyclin Analogs Differentially Regulate Macrophage Function via Distinct Analog-Receptor Binding Specificities. J. Immunol. 2007, 178, 1628–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, R.A. Microglia orchestrate neuroinflammation. Elife 2022, 11, e81890. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Smith, J.; Engel, T. Differential Expression of the Metabotropic P2Y Receptor Family in the Cortex Following Status Epilepticus and Neuroprotection via P2Y1 Antagonism in Mice. Front. Pharmacol. 2019, 10, 1558. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.G.; Tang, T.M.; Mendsaikhan, A.; Tooyama, I.; Serrano, G.E.; Sue, L.I.; Beach, T.G.; Lue, L.-F. Patterns of Expression of Purinergic Receptor P2RY12, a Putative Marker for Non-Activated Microglia, in Aged and Alzheimer’s Disease Brains. Int. J. Mol. Sci. 2020, 21, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagas, L.D.S.; Sandre, P.C.; Ribeiro, N.C.A.R.E.; Marcondes, H.; Silva, P.O.; Savino, W.; Serfaty, C.A. Environmental Signals on Microglial Function during Brain Development, Neuroplasticity, and Disease. Int. J. Mol. Sci. 2020, 21, 2111. [Google Scholar] [CrossRef] [Green Version]
- Cserép, C.; Schwarcz, A.D.; Pósfai, B.; László, Z.I.; Kellermayer, A.; Környei, Z.; Kisfali, M.; Nyerges, M.; Lele, Z.; Katona, I.; et al. Microglial control of neuronal development via somatic purinergic junctions. Cell Rep. 2022, 40, 111369. [Google Scholar] [CrossRef]
- Illes, P.; Rubini, P.; Ulrich, H.; Zhao, Y.; Tang, Y. Regulation of Microglial Functions by Purinergic Mechanisms in the Healthy and Diseased CNS. Cells 2020, 9, 1108. [Google Scholar] [CrossRef]
- Olah, M.; Menon, V.; Habib, N.; Taga, M.F.; Ma, Y.; Yung, C.J.; Cimpean, M.; Khairallah, A.; Coronas-Samano, G.; Sankowski, R.; et al. Single cell RNA sequencing of human microglia uncovers a subset associated with Alzheimer’s disease. Nat. Commun. 2020, 11, 6129. [Google Scholar] [CrossRef]
- Preissler, J.; Grosche, A.; Lede, V.; Le Duc, D.; Krügel, K.; Matyash, V.; Szulzewsky, F.; Kallendrusch, S.; Immig, K.; Kettenmann, H.; et al. Altered microglial phagocytosis in GPR34-deficient mice. Glia 2015, 63, 206–215. [Google Scholar] [CrossRef]
- Hsiao, C.-C.; Sankowski, R.; Prinz, M.; Smolders, J.; Huitinga, I.; Hamann, J. GPCRomics of Homeostatic and Disease-Associated Human Microglia. Front. Immunol. 2021, 12, 674189. [Google Scholar] [CrossRef]
- Butovsky, O.; Weiner, H.L. Microglial signatures and their role in health and disease. Nat. Rev. Neurosci. 2018, 19, 622–635. [Google Scholar] [CrossRef]
- Suzuki, T.; Kohyama, K.; Moriyama, K.; Ozaki, M.; Hasegawa, S.; Ueno, T.; Saitoe, M.; Morio, T.; Hayashi, M.; Sakuma, H. Extracellular ADP augments microglial inflammasome and NF-κB activation via the P2Y12 receptor. Eur. J. Immunol. 2020, 50, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yu, C.; He, W. Emerging trends and hot spots of NLRP3 inflammasome in neurological diseases: A bibliometric analysis. Front. Pharmacol. 2022, 13, 952211. [Google Scholar] [CrossRef]
- Jing, F.; Zhang, Y.; Long, T.; He, W.; Qin, G.; Zhang, D.; Chen, L.; Zhou, J. P2Y12 receptor mediates microglial activation via RhoA/ROCK pathway in the trigeminal nucleus caudalis in a mouse model of chronic migraine. J. Neuroinflamm. 2019, 16, 217. [Google Scholar] [CrossRef] [Green Version]
- Wihlborg, A.-K.; Wang, L.; Braun, O.O.; Eyjolfsson, A.; Gustafsson, R.; Gudbjartsson, T.; Erlinge, D. ADP Receptor P2Y12 Is Expressed in Vascular Smooth Muscle Cells and Stimulates Contraction in Human Blood Vessels. Arter. Thromb. Vasc. Biol. 2004, 24, 1810–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, K.; Uehara, A. P2Y1, P2Y6, and P2Y12 receptors in rat splenic sinus endothelial cells: An immunohistochemical and ultrastructural study. Histochem. Cell Biol. 2011, 136, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Inhibiting P2Y12 receptor relieves LPS-induced inflammation and endothelial dysfunction. Immun. Inflamm. Dis. 2022, 10, e697. [Google Scholar] [CrossRef]
- Malhotra, K.; Katsanos, A.H.; Bilal, M.; Ishfaq, M.F.; Goyal, N.; Tsivgoulis, G. Cerebrovascular Outcomes with Proton Pump Inhibitors and Thienopyridines: A Systematic Review and Meta-Analysis. Stroke 2018, 49, 312–318. [Google Scholar] [CrossRef]
- Bhavaraju, K.; Mayanglambam, A.; Rao, A.K.; Kunapuli, S.P. P2Y(12) antagonists as antiplatelet agents-Recent developments. Curr. Opin. Drug Discov. Dev. 2010, 13, 497–506. [Google Scholar]
- Pradhan, A.; Tiwari, A.; Caminiti, G.; Salimei, C.; Muscoli, S.; Sethi, R.; Perrone, M.A. Ideal P2Y12 Inhibitor in Acute Coronary Syndrome: A Review and Current Status. Int. J. Environ. Res. Public Health 2022, 19, 8977. [Google Scholar] [CrossRef]
- Angiolillo, D.J.; Rollini, F.; Storey, R.F.; Bhatt, D.L.; James, S.; Schneider, D.J.; Sibbing, D.; So, D.Y.F.; Trenk, D.; Alexopoulos, D.; et al. International Expert Consensus on Switching Platelet P2Y12 Receptor–Inhibiting Therapies. Circulation 2017, 136, 1955–1975. [Google Scholar] [CrossRef] [PubMed]
- Gelbenegger, G.; Jilma, B. Clinical pharmacology of antiplatelet drugs. Expert Rev. Clin. Pharmacol. 2022, 15, 1177–1197. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.-A.; Pereira, N. Pharmacogenomic Impact of CYP2C19 Variation on Clopidogrel Therapy in Precision Cardiovascular Medicine. J. Pers. Med. 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuszynski, D.S.; Lauver, D.A. Pleiotropic effects of clopidogrel. Purinergic Signal. 2022, 18, 253–265. [Google Scholar] [CrossRef]
- Wiviott, S.D.; Antman, E.M.; Braunwald, E. Prasugrel. Circulation 2010, 122, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Wiviott, S.D.; White, H.D.; Ohman, E.M.; Fox, K.A.; Armstrong, P.W.; Prabhakaran, D.; Hafley, G.; Lokhnygina, Y.; Boden, W.E.; Hamm, C.; et al. Prasugrel versus clopidogrel for patients with unstable angina or non-ST-segment elevation myocardial infarction with or without angiography: A secondary, prespecified analysis of the TRILOGY ACS trial. Lancet 2013, 382, 605–613. [Google Scholar] [CrossRef]
- Wongsalap, Y.; Ungsriwong, S.; Kumtep, W.; Saokaew, S.; Senthong, V.; Kengkla, K. Efficacy and Safety of Low-Dose Prasugrel Versus Clopidogrel in Patients with Acute Coronary Syndrome Undergoing Percutaneous Coronary Intervention: A Systematic Review and Meta-analysis. Cardiovasc. Drugs Ther. 2022, 36, 991–1000. [Google Scholar] [CrossRef]
- Lam, A.S.M.; Yan, B.P.Y.; Lee, V.W.Y. Efficacy and safety comparing prasugrel/ticagrelor and clopidogrel in Hong Kong post-acute coronary syndrome patients–A 10-year cohort study. Clin. Cardiol. 2021, 44, 1072–1079. [Google Scholar] [CrossRef]
- Jeger, R.V.; Pfisterer, M.; Vogt, D.R.; Galatius, S.; Abildgaard, U.; Naber, C.; Alber, H.; Eberli, F.; Kurz, D.J.; Pedrazzini, G.; et al. Competing risks of major bleeding and thrombotic events with prasugrel-based dual antiplatelet therapy after stent implantation—An observational analysis from BASKET-PROVE II. PLoS ONE 2019, 14, e0210821. [Google Scholar] [CrossRef] [Green Version]
- Amsterdam, E.A.; Wenger, N.K.; Brindis, R.G.; Casey, D.E., Jr.; Ganiats, T.G.; Holmes, D.R., Jr.; Jaffe, A.S.; Jneid, H.; Kelly, R.F.; Kontos, M.C.; et al. 2014 AHA/ACC Guideline for the Management of Patients with Non-ST-Elevation Acute Coronary Syndromes: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Am. Coll. Cardiol. 2014, 64, e139–e228. [Google Scholar] [CrossRef] [Green Version]
- Gurbel, P.A.; Bliden, K.P.; Butler, K.; Antonino, M.J.; Wei, C.; Teng, R.; Rasmussen, L.; Storey, R.F.; Nielsen, T.; Eikelboom, J.W.; et al. Response to Ticagrelor in Clopidogrel Nonresponders and Responders and Effect of Switching Therapies: The RESPOND study. Circulation 2010, 121, 1188–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zhang, C.; Zhao, J.; Xu, X.; Dang, H.; Xiao, Q.; Li, Y.; Hou, H. Effects of clopidogrel, prasugrel and ticagrelor on prevention of stent thrombosis in patients underwent percutaneous coronary intervention: A network meta-analysis. Clin. Cardiol. 2021, 44, 488–494. [Google Scholar] [CrossRef]
- Huang, B.; Qian, Y.; Xie, S.; Ye, X.; Chen, H.; Chen, Z.; Zhang, L.; Xu, J.; Hu, H.; Ma, S.; et al. Ticagrelor inhibits the NLRP3 inflammasome to protect against inflammatory disease independent of the P2Y12 signaling pathway. Cell. Mol. Immunol. 2021, 18, 1278–1289. [Google Scholar] [CrossRef]
- Triska, J.; Maitra, N.; Deshotels, M.R.; Haddadin, F.; Angiolillo, D.J.; Vilahur, G.; Jneid, H.; Atar, D.; Birnbaum, Y. A Comprehensive Review of the Pleiotropic Effects of Ticagrelor. Cardiovasc. Drugs Ther. 2022, 1–23. [Google Scholar] [CrossRef]
- Badolia, R.; Manne, B.K.; Dangelmaier, C.; Chernoff, J.; Kunapuli, S.P. Gq-mediated Akt translocation to the membrane: A novel PIP3-independent mechanism in platelets. Blood 2015, 125, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Chang, L.; Zhang, Y.; Zhai, L.; Zhang, S.; Qi, Z.; Yan, H.; Yan, Y.; Luo, X.; Zhang, S.; et al. Platelets Express Activated P2Y12 Receptor in Patients with Diabetes Mellitus. Circulation 2017, 136, 817–833. [Google Scholar] [CrossRef]
- Judge, H.M.; Buckland, R.J.; Jakubowski, J.A.; Storey, R.F. Cangrelor inhibits the binding of the active metabolites of clopidogrel and prasugrel to P2Y12 receptors in vitro. Platelets 2016, 27, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marnat, G.; Finistis, S.; Delvoye, F.; Sibon, I.; Desilles, J.-P.; Mazighi, M.; Gariel, F.; Consoli, A.; Rosso, C.; Clarençon, F.; et al. Safety and Efficacy of Cangrelor in Acute Stroke Treated with Mechanical Thrombectomy: Endovascular Treatment of Ischemic Stroke Registry and Meta-analysis. AJNR Am. J. Neuroradiol. 2022, 43, 410–415. [Google Scholar] [CrossRef] [PubMed]
- El Aouni, M.C.; Magro, E.; Abdelrady, M.; Nonent, M.; Gentric, J.C.; Ognard, J. Safety and Efficacy of Cangrelor among Three Antiplatelet Regimens during Stent-Assisted Endovascular Treatment of Unruptured Intracranial Aneurysm: A Single-Center Retrospective Study. Front. Neurol. 2022, 13, 727026. [Google Scholar] [CrossRef]
- Dovlatova, N.L.; Jakubowski, J.A.; Sugidachi, A.; Heptinstall, S. The reversible P2Y12 antagonist cangrelor influences the ability of the active metabolites of clopidogrel and prasugrel to produce irreversible inhibition of platelet function. J. Thromb. Haemost. 2008, 6, 1153–1159. [Google Scholar] [CrossRef]
- Entezami, P.; Dalfino, J.C.; Boulos, A.S.; Yamamoto, J.; Holden, D.N.; Field, N.C.; Rock, A.K.; Najera, E.; Paul, A.R. Use of intravenous cangrelor in the treatment of ruptured and unruptured cerebral aneurysms: An updated single-center analysis and pooled analysis of current studies. J. Neurointerv. Surg. 2022. [Google Scholar] [CrossRef]
- Franchi, F.; Rollini, F.; Rivas, A.; Wali, M.; Briceno, M.; Agarwal, M.; Shaikh, Z.; Nawaz, A.; Silva, G.; Been, L.; et al. Platelet Inhibition With Cangrelor and Crushed Ticagrelor in Patients With ST-Segment–Elevation Myocardial Infarction Undergoing Primary Percutaneous Coronary Intervention. Circulation 2019, 139, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, S.; Iwasaka, H.; Hasegawa, A.; Oyama, M.; Imatomi, R.; Uchida, T.; Noguchi, T. Adenosine Diphosphate Receptor Antagonist Clopidogrel Sulfate Attenuates LPS-Induced Systemic Inflammation in a Rat Model. Shock 2011, 35, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, X.-M.; Fang, L.; Jennings, N.L.; Su, Y.; Samson, A.L.; Kiriazis, H.; Wang, X.-F.; Shan, L.; Sturgeon, S.A.; et al. Novel Role of Platelets in Mediating Inflammatory Responses and Ventricular Rupture or Remodeling Following Myocardial Infarction. Arter. Thromb. Vasc. Biol. 2011, 31, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Hackert, T.; Sperber, R.; Hartwig, W.; Fritz, S.; Schneider, L.; Gebhard, M.-M.; Werner, J. P-Selectin Inhibition Reduces Severity of Acute Experimental Pancreatitis. Pancreatology 2009, 9, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Goth, C.K.; Petäjä-Repo, U.E.; Rosenkilde, M.M. G Protein-Coupled Receptors in the Sweet Spot: Glycosylation and other Post-translational Modifications. ACS Pharmacol. Transl. Sci. 2020, 3, 237–245. [Google Scholar] [CrossRef]
- Calebiro, D.; Koszegi, Z.; Lanoiselée, Y.; Miljus, T.; O’Brien, S.L. G protein-coupled receptor-G protein interactions: A single-molecule perspective. Physiol. Rev. 2021, 101, 857–906. [Google Scholar] [CrossRef]
- von Kügelgen, I.; Hoffmann, K. Pharmacology and structure of P2Y receptors. Neuropharmacology 2016, 104, 50–61. [Google Scholar] [CrossRef]
- Pérez-Sen, R.; Gómez-Villafuertes, R.; Ortega, F.; Gualix, J.; Delicado, E.G.; Miras-Portugal, M.T. An Update on P2Y13 Receptor Signalling and Function. Adv. Exp. Med. Biol. 2017, 1051, 139–168. [Google Scholar] [CrossRef]
- Marteau, F.; Le Poul, E.; Communi, D.; Communi, D.; Labouret, C.; Savi, P.; Boeynaems, J.-M.; Gonzalez, N.S. Pharmacological Characterization of the Human P2Y13 Receptor. Mol. Pharmacol. 2003, 64, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Olivecrona, G.; Götberg, M.; Olsson, M.L.; Winzell, M.S.; Erlinge, D. ADP Acting on P2Y13 Receptors Is a Negative Feedback Pathway for ATP Release from Human Red Blood Cells. Circ. Res. 2005, 96, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaduzzaman, M.; Lavasani, S.; Rahman, M.; Zhang, S.; Braun, O.Ö.; Jeppsson, B.; Thorlacius, H. Platelets support pulmonary recruitment of neutrophils in abdominal sepsis. Crit. Care Med. 2009, 37, 1389–1396. [Google Scholar] [CrossRef]
- Liverani, E.; Rico, M.C.; Yaratha, L.; Tsygankov, A.Y.; Kilpatrick, L.E.; Kunapuli, S.P. LPS-induced systemic inflammation is more severe in P2Y12 null mice. J. Leukoc. Biol. 2014, 95, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabouel, Y.; Magnenat, S.; Delabranche, X.; Gachet, C.; Hechler, B. Platelet P2Y12 Receptor Deletion or Pharmacological Inhibition does not Protect Mice from Sepsis or Septic Shock. TH Open 2021, 5, e343–e352. [Google Scholar] [CrossRef] [PubMed]
- Gopalan, C.; Erik, K. Biology of Cardiovascular and Metabolic Diseases; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Wolf, D.; Ley, K. Immunity and Inflammation in Atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Al-Abdouh, A.; Abusnina, W.; Mhanna, M.; Radideh, Q.; Alzu’Bi, H.; Abu Rmilah, A.; Jabri, A.; Barbarawi, M.; Obeidat, K.; Alabduh, T.; et al. P2Y12 Inhibitors versus Aspirin Monotherapy for Long-term Secondary Prevention of Atherosclerotic Cardiovascular Disease Events: A Systematic Review and Meta-analysis. Curr. Probl. Cardiol. 2022, 47, 101292. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, Y.; Liu, T.; Niu, X. Comparison of aspirin and P2Y12 inhibitors for secondary prevention of ischaemic stroke: A systematic review and meta-analysis. Curr. Clin. Pharmacol. 2023, 18, 270–283. [Google Scholar] [CrossRef]
- Ganbaatar, B.; Fukuda, D.; Salim, H.M.; Nishimoto, S.; Tanaka, K.; Higashikuni, Y.; Hirata, Y.; Yagi, S.; Soeki, T.; Sata, M. Ticagrelor, a P2Y12 antagonist, attenuates vascular dysfunction and inhibits atherogenesis in apolipoprotein-E-deficient mice. Atherosclerosis 2018, 275, 124–132. [Google Scholar] [CrossRef]
- Heim, C.; Gebhardt, J.; Ramsperger-Gleixner, M.; Jacobi, J.; Weyand, M.; Ensminger, S.M. Clopidogrel significantly lowers the development of atherosclerosis in ApoE-deficient mice in vivo. Heart Vessel. 2016, 31, 783–794. [Google Scholar] [CrossRef]
- Li, D.; Wang, Y.; Zhang, L.; Luo, X.; Li, J.; Chen, X.; Niu, H.; Wang, K.; Sun, Y.; Wang, X.; et al. Roles of Purinergic Receptor P2Y, G Protein–Coupled 12 in the Development of Atherosclerosis in Apolipoprotein E–Deficient Mice. Arter. Thromb. Vasc. Biol. 2012, 32, e81–e89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanker, G.; Kontos, J.L.; Eckman, D.M.; Wesley-Farrington, D.; Sane, D.C. Nicotine upregulates the expression of P2Y12 on vascular cells and megakaryoblasts. J. Thromb. Thrombolysis 2006, 22, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Rauch, B.H.; Rosenkranz, A.C.; Ermler, S.; Böhm, A.; Driessen, J.; Fischer, J.W.; Sugidachi, A.; Jakubowski, J.A.; Schrör, K. Regulation of Functionally Active P2Y12 ADP Receptors by Thrombin in Human Smooth Muscle Cells and the Presence of P2Y12 in Carotid Artery Lesions. Arter. Thromb. Vasc. Biol. 2010, 30, 2434–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.; Pi, S.-L.; Baral, S.; Xia, Y.-P.; He, Q.-W.; Li, Y.-N.; Jin, H.-J.; Li, M.; Wang, M.-D.; Mao, L.; et al. P2Y12 Promotes Migration of Vascular Smooth Muscle Cells Through Cofilin Dephosphorylation During Atherogenesis. Arter. Thromb. Vasc. Biol. 2017, 37, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Yu, C.; Pi, S.; Mao, L.; Hu, B. The role of P2Y12 receptor in ischemic stroke of atherosclerotic origin. Cell. Mol. Life Sci. 2019, 76, 341–354. [Google Scholar] [CrossRef]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The etiology of rheumatoid arthritis. J. Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- McInnes, I.B.P.; Schett, G.P. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef] [Green Version]
- Boilard, E.; Nigrovic, P.A.; Larabee, K.; Watts, G.F.M.; Coblyn, J.S.; Weinblatt, M.E.; Massarotti, E.M.; Remold-O’Donnell, E.; Farndale, R.W.; Ware, J.; et al. Platelets Amplify Inflammation in Arthritis via Collagen-Dependent Microparticle Production. Science 2010, 327, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garshick, M.S.; Rosenthal, P.B.; Luttrell-Williams, E.; Cronstein, B.N.; Berger, J.S. Ticagrelor added to methotrexate improves rheumatoid arthritis disease severity. Rheumatology 2021, 60, 5473–5475. [Google Scholar] [CrossRef]
- Garcia, A.E.; Rico, M.C.; Liverani, E.; Cadena, R.A.D.; Bray, P.F.; Kunapuli, S.P. Erosive Arthritis and Hepatic Granuloma Formation Induced by Peptidoglycan Polysaccharide in Rats Is Aggravated by Prasugrel Treatment. PLoS ONE 2013, 8, e69093. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S. Imaging the Tumor Microenvironment. Cancer J. 2015, 21, 174–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The Tumor Microenvironment: A Milieu Hindering and Obstructing Antitumor Immune Responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- Ballerini, P.; Dovizio, M.; Bruno, A.; Tacconelli, S.; Patrignani, P. P2Y12 Receptors in Tumorigenesis and Metastasis. Front. Pharmacol. 2018, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.R.; Chauhan, M.; Shah, C.; Ring, A.; Thomas, A.L.; Goodall, A.H.; Adlam, D. The TICONC (Ticagrelor-Oncology) Study: Implications of P2Y12 Inhibition for Metastasis and Cancer-Associated Thrombosis. JACC Cardio Oncol. 2020, 2, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.; Bachelot-Loza, C.; Nesseler, N.; Gaussem, P.; Gouin-Thibault, I. P2Y12 Inhibition beyond Thrombosis: Effects on Inflammation. Int. J. Mol. Sci. 2020, 21, 1391. [Google Scholar] [CrossRef] [Green Version]
- Gareau, A.J.; Brien, C.; Gebremeskel, S.; Liwski, R.S.; Johnston, B.; Bezuhly, M. Ticagrelor inhibits platelet–tumor cell interactions and metastasis in human and murine breast cancer. Clin. Exp. Metastasis 2018, 35, 25–35. [Google Scholar] [CrossRef]
- Kaufmann, C.C.; Lyon, A.R.; Wojta, J.; Huber, K. Is P2Y12 inhibitor therapy associated with an increased risk of cancer? Eur. Heart J.—Cardiovasc. Pharmacother. 2019, 5, 100–104. [Google Scholar] [CrossRef]
- Forouhi, N.G.; Wareham, N.J. Epidemiology of diabetes. Medicine 2010, 38, 602–606. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.-A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. Rev. 2019, 14, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, L.; Thomson, G.J.A.; Adams, R.C.M.; Nell, T.A.; Laubscher, W.A.; Pretorius, E. Platelet activity and hypercoagulation in type 2 diabetes. Cardiovasc. Diabetol. 2018, 17, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuno, H.; Tokuda, H.; Ishisaki, A.; Zhou, Y.; Kitajima, Y.; Kozawa, O. P2Y12 Receptors Play a Significant Role in the Development of Platelet Microaggregation in Patients with Diabetes. J. Clin. Endocrinol. Metab. 2005, 90, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, H.; Li, L.; Yang, R.; Li, G.; Liu, S.; Schmalzing, G.; Nie, H.; Liang, S. Study of the Involvement of the P2Y12 Receptor in Chronic Itching in Type 2 Diabetes Mellitus. Mol. Neurobiol. 2022, 59, 1604–1618. [Google Scholar] [CrossRef] [PubMed]
- Zaccardi, F.; Pitocco, D.; Willeit, P.; Laukkanen, J.A. Efficacy and safety of P2Y12 inhibitors according to diabetes, age, gender, body mass index and body weight: Systematic review and meta-analyses of randomized clinical trials. Atherosclerosis 2015, 240, 439–445. [Google Scholar] [CrossRef]
- Zuo, L.; Otenbaker, N.P.; Rose, B.A.; Salisbury, K.S. Molecular mechanisms of reactive oxygen species-related pulmonary inflammation and asthma. Mol. Immunol. 2013, 56, 57–63. [Google Scholar] [CrossRef]
- Yang, I.V.; Lozupone, C.A.; Schwartz, D.A. The environment, epigenome, and asthma. J. Allergy Clin. Immunol. 2017, 140, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Santana, P.T.; Luna-Gomes, T.; Rangel-Ferreira, M.V.; Tamura, A.S.; Da Graça, C.L.A.L.; Machado, M.N.; Zin, W.A.; Takiya, C.M.; Faffe, D.S.; Coutinho-Silva, R. P2Y12 Receptor Antagonist Clopidogrel Attenuates Lung Inflammation Triggered by Silica Particles. Front. Pharmacol. 2020, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Paruchuri, S.; Tashimo, H.; Feng, C.; Maekawa, A.; Xing, W.; Jiang, Y.; Kanaoka, Y.; Conley, P.; Boyce, J.A. Leukotriene E4–induced pulmonary inflammation is mediated by the P2Y12 receptor. J. Exp. Med. 2009, 206, 2543–2555. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.W.; Han, S.-T.; Gunderson, K.A.; Busse, W.W.; Jarjour, N.N.; Mosher, D.F. Platelet Activation, P-Selectin, and Eosinophil β1-Integrin Activation in Asthma. Am. J. Respir. Crit. Care Med. 2012, 185, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Sexton, T.R.; Zhang, G.; Macaulay, T.E.; Callahan, L.A.; Charnigo, R.; Vsevolozhskaya, O.A.; Li, Z.; Smyth, S. Ticagrelor Reduces Thromboinflammatory Markers in Patients With Pneumonia. JACC Basic Transl. Sci. 2018, 3, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Waresi, M.; Zhang, W.; Han, L.; Zhao, Y.; Chen, Y.; Zhou, P.; Chang, L.; Pan, G.; Wu, B.; et al. NOD2-mediated P2Y12 upregulation increases platelet activation and thrombosis in sepsis. Biochem. Pharmacol. 2021, 194, 114822. [Google Scholar] [CrossRef]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nat. Rev. Immunol. 2020, 20, 269–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.X.; Tyagi, T.; Jain, K.; Gu, V.W.; Lee, S.H.; Hwa, J.M.; Kwan, J.M.; Krause, D.S.; Lee, A.I.; Halene, S.; et al. Thrombocytopathy and endotheliopathy: Crucial contributors to COVID-19 thromboinflammation. Nat. Rev. Cardiol. 2021, 18, 194–209. [Google Scholar] [CrossRef]
- Zaid, Y.; Puhm, F.; Allaeys, I.; Naya, A.; Oudghiri, M.; Khalki, L.; Limami, Y.; Zaid, N.; Sadki, K.; Ben El Haj, R.; et al. Platelets Can Associate With SARS-CoV-2 RNA and Are Hyperactivated in COVID-19. Circ. Res. 2020, 127, 1404–1418. [Google Scholar] [CrossRef] [PubMed]
- Hottz, E.D.; Bozza, P.T. Platelet-leukocyte interactions in COVID-19: Contributions to hypercoagulability, inflammation, and disease severity. Res. Pract. Thromb. Haemost. 2022, 6, e12709. [Google Scholar] [CrossRef]
- Santoro, F.; Nuñez-Gil, I.J.; Vitale, E.; Viana-Llamas, M.C.; Reche-Martinez, B.; Romero-Pareja, R.; Feltez Guzman, G.; Fernandez Rozas, I.; Uribarri, A.; Becerra-Muñoz, V.M.; et al. Antiplatelet therapy and outcome in COVID-19: The Health Outcome Predictive Evaluation Registry. Heart 2022, 108, 130–136. [Google Scholar] [CrossRef]
- Bradbury, C.A.; Lawler, P.R.; Stanworth, S.J.; McVerry, B.J.; McQuilten, Z.; Higgins, A.M.; Mouncey, P.R.; Al-Beidh, F.; Rowan, K.M.; Berry, L.R.; et al. Effect of Antiplatelet Therapy on Survival and Organ Support–Free Days in Critically Ill Patients with COVID-19: A Randomized Clinical Trial. JAMA J. Am. Med. Assoc. 2022, 327, 1247–1259. [Google Scholar] [CrossRef]
- Khalaji, A.; Behnoush, A.H.; Peiman, S. Aspirin and P2Y12 inhibitors in treating COVID-19. Eur. J. Intern. Med. 2022, 110, 101–103. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 2019, 10, 1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, B.; Baber, U.; Cohen, D.J.; Sartori, S.; Sharma, S.K.; Angiolillo, D.J.; Farhan, S.; Goel, R.; Zhang, Z.; Briguori, C.; et al. Sex Differences among Patients with High Risk Receiving Ticagrelor with or without Aspirin after Percutaneous Coronary Intervention: A Subgroup Analysis of the TWILIGHT Randomized Clinical Trial. JAMA Cardiol. 2021, 6, 1032–1041. [Google Scholar] [CrossRef]
- Schreuder, M.M.; Badal, R.; Boersma, E.; Kavousi, M.; Roos-Hesselink, J.; Versmissen, J.; Visser, L.E.; Roeters van Lennep, J.E. Efficacy and Safety of High Potent P2Y12 Inhibitors Prasugrel and Ticagrelor in Patients with Coronary Heart Disease Treated with Dual Antiplatelet Therapy: A Sex-Specific Systematic Review and Meta-Analysis. J. Am. Heart Assoc. 2020, 9, e014457. [Google Scholar] [CrossRef]
- Husted, S.; James, S.K.; Bach, R.G.; Becker, R.C.; Budaj, A.; Heras, M.; Himmelmann, A.; Horrow, J.; Katus, H.A.; Lassila, R.; et al. The efficacy of ticagrelor is maintained in women with acute coronary syndromes participating in the prospective, randomized, PLATelet inhibition and patient Outcomes (PLATO) trial. Eur. Heart J. 2014, 35, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Ranucci, M.; Aloisio, T.; Di Dedda, U.; Menicanti, L.; de Vincentiis, C.; Baryshnikova, E.; Surgical and Clinical Outcome REsearch (SCORE) Group. Gender-based differences in platelet function and platelet reactivity to P2Y12 inhibitors. PLoS ONE 2019, 14, e0225771. [Google Scholar] [CrossRef]
- Waissi, F.; Dekker, M.; Bank, I.E.M.; Korporaal, S.J.A.; Urbanus, R.T.; de Borst, G.J.; Pasterkamp, G.; Scholtens, A.M.; Grobbee, D.E.; Mosterd, A.; et al. Sex differences in flow cytometry–based platelet reactivity in stable outpatients suspected of myocardial ischemia. Res. Pract. Thromb. Haemost. 2020, 4, 879–885. [Google Scholar] [CrossRef]
- Waissi, F.; Dekker, M.; Timmerman, N.; Hoogeveen, R.M.; Van Bennekom, J.; Dzobo, K.E.; Schnitzler, J.G.; Pasterkamp, G.; Grobbee, D.E.; De Borst, G.J.; et al. Elevated Lp(a) (Lipoprotein[a]) Levels Increase Risk of 30-Day Major Adverse Cardiovascular Events in Patients Following Carotid Endarterectomy. Stroke 2020, 51, 2972–2982. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.M.; El-Aal, S.A.A.; El-Abhar, H.S.; Ahmed, K.A.; Awny, M.M. Repositioning of Ticagrelor: Renoprotection mediated by modulating renin-angiotensin system, inflammation, autophagy and galectin-3. Eur. J. Pharmacol. 2022, 918, 174793. [Google Scholar] [CrossRef]
- Rahman, M.; Gustafsson, D.; Wang, Y.; Thorlacius, H.; Braun, O.Ö. Ticagrelor reduces neutrophil recruitment and lung damage in abdominal sepsis. Platelets 2014, 25, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Birnbaum, G.D.; Perez-Polo, J.R.; Nanhwan, M.K.; Nylander, S.; Birnbaum, Y. Ticagrelor Protects the Heart Against Reperfusion Injury and Improves Remodeling after Myocardial Infarction. Arter. Thromb. Vasc. Biol. 2015, 35, 1805–1814. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Gao, C.-M.; Xie, N.; Wang, X.-Q.; Ma, Y.-Q. Effect of ticagrelor on acute kidney injury in septic rats and its underlying mechanism. Exp. Ther. Med. 2021, 21, 475. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.-Z.; Ma, Q.-Q.; Wu, J.; Liu, R.; Wang, F.; Bai, J.; Yang, X.-J.; Fu, Q.; Wei, P. Comparison of the Effects of Ticagrelor and Clopidogrel on Inflammatory Factors, Vascular Endothelium Functions and Short-Term Prognosis in Patients with Acute ST-Segment Elevation Myocardial Infarction Undergoing Emergency Percutaneous Coronary Intervention: A Pilot Study. Cell. Physiol. Biochem. 2018, 48, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.; Reineri, S.; Piranda, D.; Pietrapiana, D.; Lova, P.; Bertoni, A.; Graziani, A.; Defilippi, P.; Canobbio, I.; Torti, M.; et al. Nongenomic effects of 17β-estradiol in human platelets: Potentiation of thrombin-induced aggregation through estrogen receptor β and Src kinase. Blood 2005, 105, 115–121. [Google Scholar] [CrossRef]
- Nealen, M.L.; Vijayan, K.V.; Bolton, E.; Bray, P.F. Human Platelets Contain a Glycosylated Estrogen Receptor β. Circ. Res. 2001, 88, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Reineri, S.; Bertoni, A.; Sanna, E.; Baldassarri, S.; Sarasso, C.; Zanfa, M.; Canobbio, I.; Torti, M.; Sinigaglia, F. Membrane lipid rafts coordinate estrogen-dependent signaling in human platelets. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuis, M.; Severin, S.; Noirrit-Esclassan, E.; Arnal, J.-F.; Payrastre, B.; Valéra, M.-C. Effects of Estrogens on Platelets and Megakaryocytes. Int. J. Mol. Sci. 2019, 20, 3111. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Kwon, J.-A.; Cho, S.-A.; Jarrar, Y.B.; Shin, J.-G. Effects of testosterone and 17β-oestradiol on expression of the G protein-coupled receptor P2Y12 in megakaryocytic DAMI cells. Platelets 2012, 23, 579–585. [Google Scholar] [CrossRef]
- Ngo, A.T.P.; Sheriff, J.; Rocheleau, A.D.; Bucher, M.; Jones, K.R.; Sepp, A.-L.I.; Malone, L.E.; Zigomalas, A.; Maloyan, A.; Bahou, W.F.; et al. Assessment of neonatal, cord, and adult platelet granule trafficking and secretion. Platelets 2020, 31, 68–78. [Google Scholar] [CrossRef]
- Li, J.S.; Yow, E.; Berezny, K.Y.; Bokesch, P.M.; Takahashi, M.; Graham, T.P., Jr.; Sanders, S.P.; Sidi, D.; Bonnet, D.; Ewert, P.; et al. Dosing of Clopidogrel for Platelet Inhibition in Infants and Young Children: Primary results of the Platelet Inhibition in Children On cLOpidogrel (PICOLO) trial. Circulation 2008, 117, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Fujisaki, T.; Kuno, T.; Ando, T.; Briasoulis, A.; Takagi, H.; Bangalore, S. Potent P2Y12 inhibitors versus Clopidogrel in elderly patients with acute coronary syndrome: Systematic review and meta-analysis. Am. Heart J. 2021, 237, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Capranzano, P.; Angiolillo, D.J. Tailoring P2Y12 Inhibiting Therapy in Elderly Patients with Myocardial Infarction Undergoing Primary Percutaneous Coronary Intervention. J. Am. Heart Assoc. 2019, 8, e014000. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Immune Cell Type | P2Y12 mRNA | P2Y12 Protein | Functional Studies | Signaling | Discrepancy of Studies | Key References |
|---|---|---|---|---|---|---|
| Platelets | Detected | Detected | Platelet aggregation and secretion | AKT phosphorylation and decrease in cAMP | Consistency | [20,21,38,39,40] |
| Monocytes | Detected | Detected | Migration | Ca2+ mobilization | Not consistently detected in monocytes | [6,36,37,41,42,43,44,45,46] |
| Macrophages | Detected | Detected | Migration | Ca2+ mobilization | Consistency | [6,36,37,41,42,43,44,45,46] |
| Neutrophils/eosinophils | Not detected in neutrophils, no studies in eosinophils | Detected in eosinophils, not neutrophils | No studies | No studies | Consistency | [5,36,47] |
| T lymphocytes | Detected | Detected | Migration, differentiation (Th17 and Tregs), and cytokine secretion | No conclusive studies | Consistency | [7,11,18,48,49,50] |
| Dendritic cells | Detected | Detected | Endocytosis, Ag-presenting functions, IL-23 production | Ca2+ mobilization | Consistency | [4,18,49] |
| Microglia | Detected | Detected | Migration | PI3K, decrease in cAMP | Consistency | [41,42] |
| Natural killer cells | No studies | No studies | No studies | No studies | N/A | N/A |
| B lymphocytes | No studies | No studies | No studies | No studies | N/A | N/A |
| Challenges | Suggested Solutions |
|---|---|
| Reliable antibody |
|
| Antagonist specificity |
|
| Cell purity |
|
| Inflammatory Disease | Animal Models | P2Y12 Antagonist | Outcome | Mechanism | Patients | Key References |
|---|---|---|---|---|---|---|
| Atherosclerosis | Clopidogrel Prasugrel Ticagrelor | Decrease plaque size and inflammatory molecules. | Blocking P2Y12 prevents platelet hyperactivation | Blocking P2Y12 appears to be beneficial. | [100,101,102,103,104] | |
| Rheumatoid arthritis | Peptidoglycan polysaccharide (PG-PS)-induced arthritis model (rats) | Clopidogrel Ticagrelor P2Y12 KO mice | Not established | Unknown | Blocking P2Y12 appears to be beneficial. | [15,16,105,106,107] |
| Tumor microenvironment | In vitro Murine model of breast cancer Murine model of pancreatic cancer | Ticagrelor Prasugrel | Mostly decrease cancer growth and risk of thrombosis but not established | Decrease platelet activation and cancer growth | Blocking P2Y12 appears to be beneficial. | [6,9,86,87,88,89,90,91,92] |
| Diabetes | Rats Mice—high glucose and high-fat diet | Ticagrelor ticlopidine | Improvements in Cardiovascular diseases | Increase P2Y12 | Blocking P2Y12 appears to be beneficial. | [68,103,104,105,106,108] |
| Pulmonary inflammation and asthma | OVA-induced asthma | Clopidogrel Ticagrelor | Decreased lung damage and diminished eosinophil infiltration. | Decrease cytokine secretion and eosinophil activation | Blocking P2Y12 appears to be beneficial. | [4,5,6,7,65,67,105,106,107,109,110,111,112,113,114,115,116] |
| Sepsis | Cecal ligation and double puncture (CLP) LPS-induced inflammation | Clopidogrel Ticagrelor P2Y12 KO mice | Not established | P-selectin surface expression T-cell differentiation | The effects of blocking P2Y12 are still unclear | [5,11,12,83,94,102,103,104,108] |
| COVID-19 | N/A | Clopidogrel Prasugrel Ticagrelor | Not established | Unknown | Only patients | [86,89,91] |
| Neuro-inflammation | Experimental autoimmune encephalomyelitis (EAE) | Clopidogrel P2Y12 KO mice | Ameliorate EAE. | Th17 differentiation microglial chemotaxis | Only animals | [15,16,41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Entsie, P.; Kang, Y.; Amoafo, E.B.; Schöneberg, T.; Liverani, E. The Signaling Pathway of the ADP Receptor P2Y12 in the Immune System: Recent Discoveries and New Challenges. Int. J. Mol. Sci. 2023, 24, 6709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076709
Entsie P, Kang Y, Amoafo EB, Schöneberg T, Liverani E. The Signaling Pathway of the ADP Receptor P2Y12 in the Immune System: Recent Discoveries and New Challenges. International Journal of Molecular Sciences. 2023; 24(7):6709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076709
Chicago/Turabian StyleEntsie, Philomena, Ying Kang, Emmanuel Boadi Amoafo, Torsten Schöneberg, and Elisabetta Liverani. 2023. "The Signaling Pathway of the ADP Receptor P2Y12 in the Immune System: Recent Discoveries and New Challenges" International Journal of Molecular Sciences 24, no. 7: 6709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24076709