
Impact of Solute Carrier Transporters in Glioma Pathology: A Comprehensive Review
 , , and
, , and
Abstract
:1. Introduction
2. Biochemical Characteristics and Physiological Role of Main SLC Family Members
2.1. Cation/Anion Transport
2.1.1. SLC1A5
2.1.2. SLC7A11
2.1.3. SLC8A2
2.1.4. SLC9A1
2.1.5. SLC11A1
2.1.6. SLC12A2
2.1.7. SLC16A1
2.1.8. SLC30A3
2.1.9. SLC39A1
2.2. Amino Acid Transport
2.2.1. SLC3A2
2.2.2. SLC7A5
2.2.3. SLC17A7
2.2.4. SLC22A5
2.3. Glucose Transport
SLC2A1
2.4. Neurotransmitter Transport
2.4.1. SLC6A2 (NET)
2.4.2. SLC6A4 (SERT)
2.4.3. SLC18A2
3. Implication of the Main SLC Family Members in Glioma Pathogenesis
3.1. SLC Implication in the Glioma Microenvironment Formation
3.2. SLC Implication in Glioma Proliferation and Progression
3.3. SLC Implication in Glioma Cell Metabolism
4. Therapeutic Targeting of SLCs

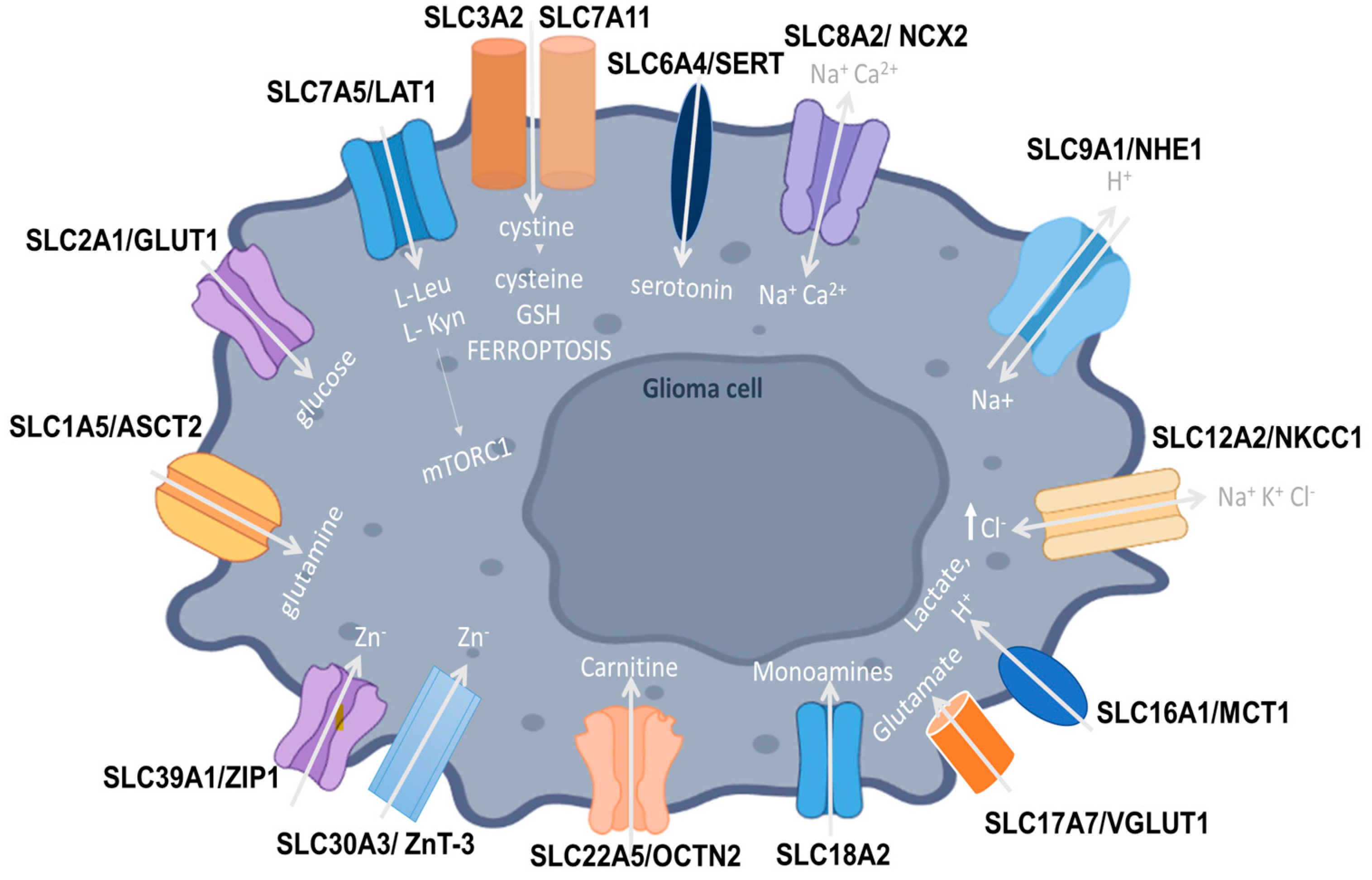{kind=link}
| SLC | Carrier Type | Role in Gliomas | Therapeutic Targeting | Clinical Trials | References |
|---|---|---|---|---|---|
| SLC1A5 (ASCT2) | Na+-dependent neutral amino acid transporter | Shifts tumor cell metabolism from glucose to glutamine pathways | [4,86,108] | ||
| SLC2A1 (GLUT1) | Glucose uptake transporter | Increased expression in tumor cells that facilitates higher glucose uptake | 2-DG potentiated the effects of radiation therapy in GB | I (NCT00096707) | [55] |
| SLC3A2 | Acts as a chaperone, facilitating the localization of the transporters to the plasma membrane | Negatively correlates with CD8+ T cell levels; therefore, it is associated with decreased cancer cell killing Transports essential amino acids and activates mTORC1, leading to tumor growth | DFMO and AMXT 1501 co-administration in DIPG and high-grade glioma increased cell death and improved survival | I/II (NCT05500508) | [60,108] |
| SLC6A2 (NET) | Norepinephrine transporter | NET-targeting 123I-metaiodobenzaguanidine (MIBG) therapy | [98] | ||
| SLC6A4 (SERT) | Monoamine serotonin transporter | TNF-α increases SERT-dependent serotonin uptake into glioma cells and activates the MAPK signaling pathway | [89] | ||
| SLC7A5 (LAT1) | Na+-independent antiporter, heterodimeric amino acid transporter | Angiogenesis and cancer cell invasion Facilitates amino acid influx and mTOR activity, aiding in tumor proliferation Evasion of immune surveillance | IV WP1066-loaded liposomes of Amphi-DOPA, combined with DC-targeted DNA vaccination in vivo Nanoparticles loaded with doxorubicin and coupled with L-Phe -3CDIT -HSVtk gene targeting through miR-145 over-expression -JPH203 LAT1 inhibitor -211At-PA -BNCT and BSH-polyR use | I (UMIN000016546) | [6,47] |
| SLC7A11 | Na+-independent, Cl−-dependent anionic L-cystine/L-glutamate antiporter | Removes lipid peroxides and inhibits apoptosis, aiding in tumor proliferation Immune cell regulation Resistance to chemotherapy | [8,9,64,109] | ||
| SLC8A2 (NCX2) | Na+/Ca2+ exchanger | Acts as a tumor suppressor, inhibiting tumor cell invasion Negatively regulates angiogenesis | [10,12] | ||
| SLC9A1 (NHE1) | Na+-H+ antiporter-1 | Promotes ECM remodeling and angiogenesis Related to the accumulation of tumor-associated macrophages | [13,15,16,17,110] | ||
| SLC11A1 (NRAMP1) | Antiporter, facilitating the influx of cations into the phagolysosome or cytosol | [19,20] | |||
| SLC12A2 (NKCC1) | Na-K-2Cl cotransporter-1 | Linked to GB cell proliferation Associated with aberrant and immature neuronal cells | Bumetanide reduced GB migration and invasion | [26,30,72,106] | |
| SLC16A1 (MCT1) | Proton-coupled monocarboxylate transporter | Increases glioma cell invasion and mitotic activity | [35,91] | ||
| SLC18A2 | Monoamine/neurotransmitter transporter | Promotes glioma formation | [82] | ||
| SLC22A5 (OCTN2) | Carnitine plasma membrane transporter | Correlated with FAO rates and carnitine transport Associated with glioma cell viability and reduced apoptosis | Combined SLC22A5 and CPT1 targeting Chemotherapy inhibited carnitine uptake and etomoxir | [50,95,107] | |
| SLC30A3 | Zinc transporter | Inhibits GB cell growth and cell cycle progression Inhibits EMT in GB Represses GB malignant phenotype | [83] | ||
| SLC39A1 | Zinc transporter | Promotes glioma cell proliferation Influences immune cell infiltration in the glioma TME | [45,84] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, C.; Tao, L.; Cao, X.; Chen, L. The Solute Carrier Transporters and the Brain: Physiological and Pharmacological Implications. Asian J. Pharm. Sci. 2020, 15, 131–144. [Google Scholar] [CrossRef]
- Hong, M. Biochemical Studies on the Structure–Function Relationship of Major Drug Transporters in the ATP-Binding Cassette Family and Solute Carrier Family. Adv. Drug Deliv. Rev. 2017, 116, 3–20. [Google Scholar] [CrossRef]
- Dvorak, V.; Wiedmer, T.; Ingles-Prieto, A.; Altermatt, P.; Batoulis, H.; Bärenz, F.; Bender, E.; Digles, D.; Dürrenberger, F.; Heitman, L.H.; et al. An Overview of Cell-Based Assay Platforms for the Solute Carrier Family of Transporters. Front. Pharmacol. 2021, 12, 722889. [Google Scholar] [CrossRef]
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The Human SLC1A5 (ASCT2) Amino Acid Transporter: From Function to Structure and Role in Cell Biology. Front. Cell Dev. Biol. 2018, 6, 96. [Google Scholar] [CrossRef]
- Lopes, C.; Pereira, C.; Medeiros, R. ASCT2 and LAT1 Contribution to the Hallmarks of Cancer: From a Molecular Perspective to Clinical Translation. Cancers 2021, 13, 203. [Google Scholar] [CrossRef]
- Scalise, M.; Galluccio, M.; Console, L.; Pochini, L.; Indiveri, C. The Human SLC7A5 (LAT1): The Intriguing Histidine/Large Neutral Amino Acid Transporter and Its Relevance to Human Health. Front. Chem. 2018, 6, 243. [Google Scholar] [CrossRef]
- Teixeira, E.; Silva, C.; Martel, F. The Role of the Glutamine Transporter ASCT2 in Antineoplastic Therapy. Cancer Chemother. Pharmacol. 2021, 87, 447–464. [Google Scholar] [CrossRef]
- Lin, W.; Wang, C.; Liu, G.; Bi, C.; Wang, X.; Zhou, Q.; Jin, H. SLC7A11/XCT in Cancer: Biological Functions and Therapeutic Implications. Am. J. Cancer Res. 2020, 10, 3106. [Google Scholar]
- Koppula, P.; Zhuang, L.; Gan, B. Cystine Transporter SLC7A11/XCT in Cancer: Ferroptosis, Nutrient Dependency, and Cancer Therapy. Protein Cell 2021, 12, 599–620. [Google Scholar] [CrossRef]
- Qu, M.; Yu, J.; Liu, H.; Ren, Y.; Ma, C.; Bu, X.; Lan, Q. The Candidate Tumor Suppressor Gene SLC8A2 Inhibits Invasion, Angiogenesis and Growth of Glioblastoma. Mol. Cells 2017, 40, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, L.; Serani, A.; Natale, S.; Tedeschi, V.; Guida, N.; Valsecchi, V.; Secondo, A.; Formisano, L.; Annunziato, L.; Molinaro, P. Identification and Characterization of the Promoter and Transcription Factors Regulating the Expression of Cerebral Sodium/Calcium Exchanger 2 (NCX2) Gene. Cell Calcium 2022, 102, 102542. [Google Scholar] [CrossRef]
- SLC8A2—Sodium/Calcium Exchanger 2—Homo sapiens (Human)|UniProtKB|UniProt. Available online: https://www.uniprot.org/uniprotkb/Q9UPR5/entry (accessed on 27 March 2023).
- Counillon, L.; Pouysségur, J.; Reithmeier, R.A.F. The Na+/H+ Exchanger NHE-1 Possesses N- and O-Linked Glycosylation Restricted to the First N-Terminal Extracellular Domain. Biochemistry 1994, 33, 10463–10469. [Google Scholar] [CrossRef] [PubMed]
- Haworth, R.S.; Frohlich, O.; Fliegel, L. Multiple Carbohydrate Moieties on the Na+/H+ Exchanger. Biochem. J. 1993, 289, 637–640. [Google Scholar] [CrossRef] [PubMed]
- Denker, S.P.; Barber, D.L. Cell Migration Requires Both Ion Translocation and Cytoskeletal Anchoring by the Na-H Exchanger NHE1. J. Cell Biol. 2002, 159, 1087–1096. [Google Scholar] [CrossRef]
- Denker, S.P.; Huang, D.C.; Orlowski, J.; Furthmayr, H.; Barber, D.L. Direct Binding of the Na–H Exchanger NHE1 to ERM Proteins Regulates the Cortical Cytoskeleton and Cell Shape Independently of H+ Translocation. Mol. Cell 2000, 6, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.; Stock, C.M.; Dieterich, P.; Jensen, B.H.; Pedersen, L.B.; Satir, P.; Schwab, A.; Christensen, S.T.; Pedersen, S.F. The Na+/H+ Exchanger NHE1 Is Required for Directional Migration Stimulated via PDGFR-Alpha in the Primary Cilium. J. Cell Biol. 2009, 185, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhao, P.; Xue, J.; Gu, X.Q.; Sun, X.; Yao, H.; Haddad, G.G. Na+ Channel Expression and Neuronal Function in the Na+/H+ Exchanger 1 Null Mutant Mouse. J. Neurophysiol. 2003, 89, 229–236. [Google Scholar] [CrossRef]
- Cellier, M.F.M. Cell-Type Specific Determinants of NRAMP1 Expression in Professional Phagocytes. Biology 2013, 2, 233. [Google Scholar] [CrossRef]
- Gruenheid, S.; Pinner, E.; Desjardins, M.; Gros, P. Natural Resistance to Infection with Intracellular Pathogens: The Nramp1 Protein Is Recruited to the Membrane of the Phagosome. J. Exp. Med. 1997, 185, 717–730. [Google Scholar] [CrossRef]
- Goswami, T.; Bhattacharjee, A.; Babal, P.; Searle, S.; Moore, E.; Li, M.; Blackwell, J.M. Natural-Resistance-Associated Macrophage Protein 1 Is an H+/Bivalent Cation Antiporter. Biochem. J. 2001, 354, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Philpott, D.J.; Sorbara, M.T.; Robertson, S.J.; Croitoru, K.; Girardin, S.E. NOD Proteins: Regulators of Inflammation in Health and Disease. Nat. Rev. Immunol. 2014, 14, 9–23. [Google Scholar] [CrossRef]
- Thorsen, F.; Afione, S.; Huszthy, P.C.; Tysnes, B.B.; Svendsen, A.; Bjerkvig, R.; Kotin, R.M.; Lønning, P.E.; Hoover, F. Adeno-Associated Virus (AAV) Serotypes 2, 4 and 5 Display Similar Transduction Profiles and Penetrate Solid Tumor Tissue in Models of Human Glioma. J. Gene Med. 2006, 8, 1131–1140. [Google Scholar] [CrossRef]
- Xu, Y.F.; Zhang, Y.Q.; Xu, X.M.; Song, G.X. Papillomavirus Virus-like Particles as Vehicles for the Delivery of Epitopes or Genes. Arch. Virol. 2006, 151, 2133–2148. [Google Scholar] [CrossRef]
- Zhang, X.; Godbey, W.T. Viral Vectors for Gene Delivery in Tissue Engineering. Adv. Drug Deliv. Rev. 2006, 58, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Koumangoye, R.; Bastarache, L.; Delpire, E. NKCC1: Newly Found as a Human Disease-Causing Ion Transporter. Function 2021, 2, zqaa028. [Google Scholar] [CrossRef] [PubMed]
- Piechotta, K.; Lu, J.; Delpire, E. Cation Chloride Cotransporters Interact with the Stress-Related Kinases Ste20-Related Proline-Alanine-Rich Kinase (SPAK) and Oxidative Stress Response 1 (OSR1). J. Biol. Chem. 2002, 277, 50812–50819. [Google Scholar] [CrossRef]
- Dowd, B.F.X.; Forbush, B. PASK (Proline-Alanine-Rich STE20-Related Kinase), a Regulatory Kinase of the Na-K-Cl Cotransporter (NKCC1). J. Biol. Chem. 2003, 278, 27347–27353. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, K.B.E.; England, R.; Delpire, E. Volume Sensitivity of Cation-Cl- Cotransporters Is Modulated by the Interaction of Two Kinases: Ste20-Related Proline-Alanine-Rich Kinase and WNK4. Am. J. Physiol. Cell Physiol. 2006, 290, C134–C142. [Google Scholar] [CrossRef] [PubMed]
- Vitari, A.C.; Thastrup, J.; Rafiqi, F.H.; Deak, M.; Morrice, N.A.; Karlsson, H.K.R.; Alessi, D.R. Functional Interactions of the SPAK/OSR1 Kinases with Their Upstream Activator WNK1 and Downstream Substrate NKCC1. Biochem. J. 2006, 397, 223–231. [Google Scholar] [CrossRef]
- Darman, R.B.; Flemmer, A.; Forbush, B. Modulation of Ion Transport by Direct Targeting of Protein Phosphatase Type 1 to the Na-K-Cl Cotransporter. J. Biol. Chem. 2001, 276, 34359–34362. [Google Scholar] [CrossRef]
- Gagnon, K.B.; Delpire, E. Multiple Pathways for Protein Phosphatase 1 (PP1) Regulation of Na-K-2Cl Cotransporter (NKCC1) Function: The N-Terminal Tail of the Na-K-2Cl Cotransporter Serves as a Regulatory Scaffold for Ste20-Related Proline/Alanine-Rich Kinase (SPAK) AND PP1. J. Biol. Chem. 2010, 285, 14115–14121. [Google Scholar] [CrossRef]
- Delpire, E.; Gagnon, K.B. Na+ -K+ -2Cl− Cotransporter (NKCC) Physiological Function in Nonpolarized Cells and Transporting Epithelia. Compr. Physiol. 2018, 8, 871–901. [Google Scholar] [CrossRef]
- Garcia, C.K.; Goldstein, J.L.; Pathak, R.K.; Anderson, R.G.W.; Brown, M.S. Molecular Characterization of a Membrane Transporter for Lactate, Pyruvate, and Other Monocarboxylates: Implications for the Cori Cycle. Cell 1994, 76, 865–873. [Google Scholar] [CrossRef]
- Garcia, C.K.; Li, X.; Luna, J.; Francke, U. CDNA Cloning of the Human Monocarboxylate Transporter 1 and Chromosomal Localization of the SLC16A1 Locus to 1p13.2-P12. Genomics 1994, 23, 500–503. [Google Scholar] [CrossRef]
- Felmlee, M.A.; Jones, R.S.; Rodriguez-Cruz, V.; Follman, K.E.; Morris, M.E. Monocarboxylate Transporters (SLC16): Function, Regulation, and Role in Health and Disease. Pharmacol. Rev. 2020, 72, 466–485. [Google Scholar] [CrossRef]
- Berrios, C.; Padi, M.; Keibler, M.A.; Park, D.E.; Molla, V.; Cheng, J.; Lee, S.M.; Stephanopoulos, G.; Quackenbush, J.; DeCaprio, J.A. Merkel Cell Polyomavirus Small T Antigen Promotes Pro-Glycolytic Metabolic Perturbations Required for Transformation. PLoS Pathog. 2016, 12, e1006020. [Google Scholar] [CrossRef]
- Le Floch, R.; Chiche, J.; Marchiq, I.; Naïken, T.; Ilk, K.; Murray, C.M.; Critchlow, S.E.; Roux, D.; Simon, M.P.; Pouysségur, J. CD147 Subunit of Lactate/H+ Symporters MCT1 and Hypoxia-Inducible MCT4 Is Critical for Energetics and Growth of Glycolytic Tumors. Proc. Natl. Acad. Sci. USA 2011, 108, 16663–16668. [Google Scholar] [CrossRef]
- Payen, V.L.; Hsu, M.Y.; Rädecke, K.S.; Wyart, E.; Vazeille, T.; Bouzin, C.; Porporato, P.E.; Sonveaux, P. Monocarboxylate Transporter MCT1 Promotes Tumor Metastasis Independently of Its Activity as a Lactate Transporter. Cancer Res. 2017, 77, 5591–5601. [Google Scholar] [CrossRef]
- Sprowl-Tanio, S.; Habowski, A.N.; Pate, K.T.; McQuade, M.M.; Wang, K.; Edwards, R.A.; Grun, F.; Lyou, Y.; Waterman, M.L. Lactate/Pyruvate Transporter MCT-1 Is a Direct Wnt Target That Confers Sensitivity to 3-Bromopyruvate in Colon Cancer. Cancer Metab. 2016, 4, 20. [Google Scholar] [CrossRef]
- Doherty, J.R.; Yang, C.; Scott, K.E.N.; Cameron, M.D.; Fallahi, M.; Li, W.; Hall, M.A.; Amelio, A.L.; Mishra, J.K.; Li, F.; et al. Blocking Lactate Export by Inhibiting the Myc Target MCT1 Disables Glycolysis and Glutathione Synthesis. Cancer Res. 2014, 74, 908–920. [Google Scholar] [CrossRef]
- Cole, T.B.; Wenzel, H.J.; Kafer, K.E.; Schwartzkroin, P.A.; Palmiter, R.D. Elimination of Zinc from Synaptic Vesicles in the Intact Mouse Brain by Disruption of the ZnT3 Gene. Proc. Natl. Acad. Sci. USA 1999, 96, 1716–1721. [Google Scholar] [CrossRef]
- Patrushev, N.; Seidel-Rogol, B.; Salazar, G. Angiotensin II Requires Zinc and Downregulation of the Zinc Transporters ZnT3 and ZnT10 to Induce Senescence of Vascular Smooth Muscle Cells. PLoS ONE 2012, 7, e33211. [Google Scholar] [CrossRef]
- Mott, D.D.; Dingledine, R. Unraveling the Role of Zinc in Memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3103–3104. [Google Scholar] [CrossRef]
- Yu, D.; Chen, Y.; Luo, M.; Peng, Y.; Yi, S. Upregulated Solute Carrier SLC39A1 Promotes Gastric Cancer Proliferation and Indicates Unfavorable Prognosis. Genet. Res. 2022, 2022, 1256021. [Google Scholar] [CrossRef]
- Cappoli, N.; Jenkinson, M.D.; Dello Russo, C.; Dickens, D. LAT1, a Novel Pharmacological Target for the Treatment of Glioblastoma. Biochem. Pharmacol. 2022, 201, 115103. [Google Scholar] [CrossRef]
- Onishi, Y.; Hiraiwa, M.; Kamada, H.; Iezaki, T.; Yamada, T.; Kaneda, K.; Hinoi, E. Hypoxia Affects Slc7a5 Expression through HIF-2α in Differentiated Neuronal Cells. FEBS openbio 2019, 9, 241–247. [Google Scholar] [CrossRef]
- SLC17A7 Solute Carrier Family 17 Member 7 [Homo sapiens (Human)]—Gene—NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/57030 (accessed on 8 April 2023).
- Du, X.; Li, J.; Li, M.; Yang, X.; Qi, Z.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. Research Progress on the Role of Type i Vesicular Glutamate Transporter (VGLUT1) in Nervous System Diseases. Cell Biosci. 2020, 10, 26. [Google Scholar] [CrossRef]
- Wang, C.; Uray, I.P.; Mazumdar, A.; Mayer, J.A.; Brown, P.H. SLC22A5/OCTN2 Expression in Breast Cancer Is Induced by Estrogen via a Novel Intronic Estrogen-Response Element (ERE). Breast Cancer Res. Treat. 2012, 134, 101–115. [Google Scholar] [CrossRef]
- Zhou, X.; Ringseis, R.; Wen, G.; Eder, K. The Pro-Inflammatory Cytokine Tumor Necrosis Factor α Stimulates Expression of the Carnitine Transporter OCTN2 (Novel Organic Cation Transporter 2) and Carnitine Uptake via Nuclear Factor-ΚB in Madin-Darby Bovine Kidney Cells. J. Dairy Sci. 2015, 98, 3840–3848. [Google Scholar] [CrossRef]
- Ingoglia, F.; Visigalli, R.; Rotoli, B.M.; Barilli, A.; Riccardi, B.; Puccini, P.; Milioli, M.; Di Lascia, M.; Bernuzzi, G.; Dall’Asta, V. Human Macrophage Differentiation Induces OCTN2-Mediated L-Carnitine Transport through Stimulation of MTOR-STAT3 Axis. J. Leukoc. Biol. 2017, 101, 665–674. [Google Scholar] [CrossRef]
- Di San Filippo, C.A.; Ardon, O.; Longo, N. Glycosylation of the OCTN2 Carnitine Transporter: Study of Natural Mutations Identified in Patients with Primary Carnitine Deficiency. Biochim. Biophys. Acta 2011, 1812, 312–320. [Google Scholar] [CrossRef]
- Czeredys, M.; Samluk, Ł.; Michalec, K.; Tułodziecka, K.; Skowronek, K.; Nałȩcz, K.A. Caveolin-1—A Novel Interacting Partner of Organic Cation/Carnitine Transporter (Octn2): Effect of Protein Kinase C on This Interaction in Rat Astrocytes. PLoS ONE 2013, 8, e82105. [Google Scholar] [CrossRef]
- Chen, C.; Pore, N.; Behrooz, A.; Ismail-Beigi, F.; Maity, A. Regulation of Glut1 MRNA by Hypoxia-Inducible Factor-1. Interaction between H-Ras and Hypoxia. J. Biol. Chem. 2001, 276, 9519–9525. [Google Scholar] [CrossRef]
- SLC6A2 Solute Carrier Family 6 Member 2 [Homo sapiens (Human)]—Gene—NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6530#summary (accessed on 21 May 2023).
- SLC18A2 Solute Carrier Family 18 Member A2 [Homo sapiens (Human)]—Gene—NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6571 (accessed on 21 May 2023).
- Anagnostakis, F.; Piperi, C. Targeting Options of Tumor-Associated Macrophages (TAM) Activity in Gliomas. Curr. Neuropharmacol. 2022, 21, 457–470. [Google Scholar] [CrossRef]
- Strepkos, D.; Markouli, M.; Klonou, A.; Piperi, C.; Papavassiliou, A.G. Insights in the Immunobiology of Glioblastoma. J. Mol. Med. 2020, 98, 1–10. [Google Scholar] [CrossRef]
- He, J.; Liu, D.; Liu, M.; Tang, R.; Zhang, D. Characterizing the Role of SLC3A2 in the Molecular Landscape and Immune Microenvironment across Human Tumors. Front. Mol. Biosci. 2022, 9, 961410. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Liu, X.-H.; Zeng, X.-Y.; Long, Q.-Y.; Liu, Y.-H.; Mao, Q. Evaluation of CD98 Light Chain-LAT1 as a Potential Marker of Cancer Stem-like Cells in Glioblastoma. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119303. [Google Scholar] [CrossRef]
- Cai, L.; Kirchleitner, S.V.; Zhao, D.; Li, M.; Tonn, J.C.; Glass, R.; Kälin, R.E. Glioblastoma Exhibits Inter-Individual Heterogeneity of Tspo and Lat1 Expression in Neoplastic and Parenchymal Cells. Int. J. Mol. Sci. 2020, 21, 612. [Google Scholar] [CrossRef]
- Ni, R.; Li, Z.; Li, L.; Peng, D.; Ming, Y.; Li, L.; Liu, Y. Rethinking glutamine metabolism and the regulation of glutamine addiction by oncogenes in cancer. Front Oncol. 2023, 13, 1143798. [Google Scholar] [CrossRef]
- Koppula, P.; Zhang, Y.; Zhuang, L.; Gan, B. Amino Acid Transporter SLC7A11/ XCT at the Crossroads of Regulating Redox Homeostasis and Nutrient Dependency of Cancer. Cancer Commun. 2018, 38, 1–13. [Google Scholar] [CrossRef]
- Bay, C.; Bajraktari-Sylejmani, G.; Haefeli, W.E.; Burhenne, J.; Weiss, J.; Sauter, M. Functional Characterization of the Solute Carrier LAT-1 (SLC7A5/SLC2A3) in Human Brain Capillary Endothelial Cells with Rapid UPLC-MS/MS Quantification of Intracellular Isotopically Labelled L-Leucine. Int. J. Mol. Sci. 2022, 23, 3637. [Google Scholar] [CrossRef]
- Guan, X.; Luo, L.; Begum, G.; Kohanbash, G.; Song, Q.; Rao, A.; Amankulor, N.; Sun, B.; Sun, D.; Jia, W. Elevated Na/H Exchanger 1 (SLC9A1) Emerges as a Marker for Tumorigenesis and Prognosis in Gliomas 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. J. Exp. Clin. Cancer Res. 2018, 37, 255. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Liu, Y.; Wang, S.; Sun, M.; Zhang, Z.; Zheng, X.; Li, J.; Li, Y. The Positive Feedback Loop of NHE1-ERK Phosphorylation Mediated by BRAFV600E Mutation Contributes to Tumorigenesis and Development of Glioblastoma. Biochem. Biophys. Res. Commun. 2022, 588, 1–7. [Google Scholar] [CrossRef]
- Granja, S.; Tavares-Valente, D.; Queirós, O.; Baltazar, F. Value of PH Regulators in the Diagnosis, Prognosis and Treatment of Cancer. Semin. Cancer Biol. 2017, 43, 17–34. [Google Scholar] [CrossRef]
- Aronica, E.; Boer, K.; Redeker, S.; Spliet, W.G.M.; van Rijen, P.C.; Troost, D.; Gorter, J.A. Differential Expression Patterns of Chloride Transporters, Na+-K+-2Cl--Cotransporter and K+-Cl--Cotransporter, in Epilepsy-Associated Malformations of Cortical Development. Neuroscience 2007, 145, 185–196. [Google Scholar] [CrossRef]
- Pallud, J.; Le Van Quyen, M.; Bielle, F.; Pellegrino, C.; Varlet, P.; Labussiere, M.; Cresto, N.; Dieme, M.J.; Baulac, M.; Duyckaerts, C.; et al. Cortical GABAergic Excitation Contributes to Epileptic Activities around Human Glioma. Sci. Transl. Med. 2014, 6, 244ra89. [Google Scholar] [CrossRef]
- Cong, D.; Zhu, W.; Kuo, J.S.; Hu, S.; Sun, D. Ion Transporters in Brain Tumors. Curr. Med. Chem. 2015, 22, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Garzon-Muvdi, T.; Schiapparelli, P.; ap Rhys, C.; Guerrero-Cazares, H.; Smith, C.; Kim, D.H.; Kone, L.; Farber, H.; Lee, D.Y.; An, S.S.; et al. Regulation of Brain Tumor Dispersal by NKCC1 through a Novel Role in Focal Adhesion Regulation. PLoS Biol. 2012, 10, e1001320. [Google Scholar] [CrossRef]
- Damanskienė, E.; Balnytė, I.; Valančiūtė, A.; Alonso, M.M.; Preikšaitis, A.; Stakišaitis, D. The Different Temozolomide Effects on Tumorigenesis Mechanisms of Pediatric Glioblastoma PBT24 and SF8628 Cell Tumor in CAM Model and on Cells In Vitro. Int. J. Mol. Sci. 2022, 23, 2001. [Google Scholar] [CrossRef]
- Algharabil, J.; Kintner, D.B.; Wang, Q.; Begum, G.; Clark, P.A.; Yang, S.S.; Lin, S.H.; Kahle, K.T.; Kuo, J.S.; Sun, D. Inhibition of Na+-K+-2Cl−-Cotransporter Isoform 1 Accelerates Temozolomidemediated Apoptosis in Glioblastoma Cancer Cells. Cell. Physiol. Biochem. 2012, 30, 33–48. [Google Scholar] [CrossRef]
- Gagnon, K.B.E.; England, R.; Delpire, E. Characterization of SPAK and OSR1, Regulatory Kinases of the Na-K-2Cl Cotransporter. Mol. Cell Biol. 2006, 26, 689–698. [Google Scholar] [CrossRef]
- Haas, B.R.; Sontheimer, H. Inhibition of the Sodium-Potassium-Chloride Cotransporter Isoform-1 Reduces Glioma Invasion. Cancer Res. 2010, 70, 5597–5606. [Google Scholar] [CrossRef]
- Lin, B.; Lee, H.; Yoon, J.G.; Madan, A.; Wayner, E.; Tonning, S.; Hothi, P.; Schroeder, B.; Ulasov, I.; Foltz, G.; et al. Global Analysis of H3K4me3 and H3K27me3 Profiles in Glioblastoma Stem Cells and Identification of SLC17A7 as a Bivalent Tumor Suppressor Gene. Oncotarget 2015, 6, 5369. [Google Scholar] [CrossRef]
- Hervás-Corpión, I.; Navarro-Calvo, J.; Martín-Climent, P.; Iriarte-Gahete, M.; Geribaldi-Doldán, N.; Castro, C.; Valor, L.M. Defining a Correlative Transcriptional Signature Associated with Bulk Histone H3 Acetylation Levels in Adult Glioblastomas. Cells 2023, 12, 374. [Google Scholar] [CrossRef] [PubMed]
- Markouli, M.; Strepkos, D.; Papavassiliou, K.A.; Papavassiliou, A.G.; Piperi, C. Bivalent Genes Targeting of Glioma Heterogeneity and Plasticity. Int. J. Mol. Sci. 2021, 22, 540. [Google Scholar] [CrossRef] [PubMed]
- Klonou, A.; Korkolopoulou, P.; Giannopoulou, A.I.; Kanakoglou, D.S.; Pampalou, A.; Gargalionis, A.N.; Sarantis, P.; Mitsios, A.; Sgouros, S.; Papavassiliou, A.G.; et al. Histone H3K9 Methyltransferase SETDB1 Overexpression Correlates with Pediatric High-Grade Gliomas Progression and Prognosis. J. Mol. Med. 2023, 101, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Feyissa, A.M.; Carrano, A.; Wang, X.; Allen, M.; Ertekin-Taner, N.; Dickson, D.W.; Jentoft, M.E.; Rosenfeld, S.S.; Tatum, W.O.; Ritaccio, A.L.; et al. Analysis of Intraoperative Human Brain Tissue Transcriptome Reveals Putative Risk Genes and Altered Molecular Pathways in Glioma-Related Seizures. Epilepsy Res. 2021, 173, 106618. [Google Scholar] [CrossRef]
- Kraboth, Z.; Kajtár, B.; Gálik, B.; Gyenesei, A.; Miseta, A.; Kalman, B. Involvement of the Catecholamine Pathway in Glioblastoma Development. Cells 2021, 10, 549. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Z.; Dong, Y.; Kong, L. Epigenetic Targeting of SLC30A3 by HDAC1 Is Related to the Malignant Phenotype of Glioblastoma. IUBMB Life 2021, 73, 784–799. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, J.; He, S.; Xiao, B.; Peng, X. SLC39A1 Contribute to Malignant Progression and Have Clinical Prognostic Impact in Gliomas. Cancer Cell Int. 2020, 20, 573. [Google Scholar] [CrossRef]
- Markouli, M.; Strepkos, D.; Papavassiliou, K.A.; Papavassiliou, A.G.; Piperi, C. Crosstalk of Epigenetic and Metabolic Signaling Underpinning Glioblastoma Pathogenesis. Cancers 2022, 14, 2655. [Google Scholar] [CrossRef]
- Han, L.; Zhou, J.; Li, L.; Wu, X.; Shi, Y.; Cui, W.; Zhang, S.; Hu, Q.; Wang, J.; Bai, H.; et al. SLC1A5 Enhances Malignant Phenotypes through Modulating Ferroptosis Status and Immune Microenvironment in Glioma. Cell Death Dis. 2022, 13, 1071. [Google Scholar] [CrossRef]
- Syafruddin, S.E.; Nazarie, W.F.W.M.; Moidu, N.A.; Soon, B.H.; Mohtar, M.A. Integration of RNA-Seq and Proteomics Data Identifies Glioblastoma Multiforme Surfaceome Signature. BMC Cancer 2021, 21, 850. [Google Scholar] [CrossRef]
- Huang, Q.; Chen, L.; Liang, J.; Huang, Q.; Sun, H. Neurotransmitters: Potential Targets in Glioblastoma. Cancers (Basel) 2022, 14, 3970. [Google Scholar] [CrossRef]
- Malynn, S.; Campos-Torres, A.; Moynagh, P.; Haase, J. The Pro-Inflammatory Cytokine TNF-α Regulates the Activity and Expression of the Serotonin Transporter (SERT) in Astrocytes. Neurochem. Res. 2013, 38, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Kiuchi, Y.; Nemoto, M.; Oyamada, H.; Ohno, M.; Funahashi, H.; Shioda, S.; Oguchi, K. Regulation of Serotonin Transporter Gene Expression in Human Glial Cells by Growth Factors. Eur. J. Pharmacol. 2001, 417, 69–76. [Google Scholar] [CrossRef]
- Lin, H.H.; Tsai, W.C.; Tsai, C.K.; Chen, S.H.; Huang, L.C.; Hueng, D.Y.; Hung, K.C. Overexpression of Cell-Surface Marker SLC16A1 Shortened Survival in Human High-Grade Gliomas. J. Mol. Neurosci. 2021, 71, 1614–1621. [Google Scholar] [CrossRef]
- Ghosh, D.; Ulasov, I.V.; Chen, L.P.; Harkins, L.E.; Wallenborg, K.; Hothi, P.; Rostad, S.; Hood, L.; Cobbs, C.S. TGFβ-Responsive HMOX1 Expression Is Associated with Stemness and Invasion in Glioblastoma Multiforme. Stem Cells 2016, 34, 2276–2289. [Google Scholar] [CrossRef]
- Ghosh, D.; Funk, C.C.; Caballero, J.; Shah, N.; Rouleau, K.; Earls, J.C.; Soroceanu, L.; Foltz, G.; Cobbs, C.S.; Price, N.D.; et al. A Cell-Surface Membrane Protein Signature for Glioblastoma. Cell Syst. 2017, 4, 516–529. [Google Scholar] [CrossRef]
- Fang, J.; Quinones, Q.J.; Holman, T.L.; Morowitz, M.J.; Wang, Q.; Zhao, H.; Sivo, F.; Maris, J.M.; Wahl, M.L. The H+-Linked Monocarboxylate Transporter (MCT1/SLC16A1): A Potential Therapeutic Target for High-Risk Neuroblastoma. Mol. Pharmacol. 2006, 70, 2108–2115. [Google Scholar] [CrossRef]
- Juraszek, B.; Nałąecz, K.A. SLC22A5 (OCTN2) Carnitine Transporter—Indispensable for Cell Metabolism, a Jekyll and Hyde of Human Cancer. Molecules 2020, 25, 14. [Google Scholar] [CrossRef]
- Shah, S.S.; Rodriguez, G.A.; Musick, A.; Walters, W.M.; de Cordoba, N.; Barbarite, E.; Marlow, M.M.; Marples, B.; Prince, J.S.; Komotar, R.J.; et al. Targeting Glioblastoma Stem Cells with 2-Deoxy-D-Glucose (2-DG) Potentiates Radiation-Induced Unfolded Protein Response (UPR). Cancers 2019, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Gamble, L.D.; Upton, D.H.; Ung, C.; Yu, D.M.T.; Ehteda, A.; Pandher, R.; Mayoh, C.; Hébert, S.; Jabado, N.; et al. Dual Targeting of Polyamine Synthesis and Uptake in Diffuse Intrinsic Pontine Gliomas. Nat. Commun. 2021, 12, 971. [Google Scholar] [CrossRef] [PubMed]
- Pandit-Taskar, N.; Modak, S. Norepinephrine Transporter as a Target for Imaging and Therapy. J. Nucl. Med. 2017, 58, 39S. [Google Scholar] [CrossRef]
- Bhunia, S.; Vangala, V.; Bhattacharya, D.; Ravuri, H.G.; Kuncha, M.; Chakravarty, S.; Sistla, R.; Chaudhuri, A. Large Amino Acid Transporter 1 Selective Liposomes of l -DOPA Functionalized Amphiphile for Combating Glioblastoma. Mol. Pharm. 2017, 14, 3834–3847. [Google Scholar] [CrossRef]
- Cormerais, Y.; Pagnuzzi-Boncompagni, M.; Schrötter, S.; Giuliano, S.; Tambutté, E.; Endou, H.; Wempe, M.F.; Pagès, G.; Pouysségur, J.; Picco, V. Inhibition of the Amino-Acid Transporter LAT1 Demonstrates Anti-Neoplastic Activity in Medulloblastoma. J. Cell Mol. Med. 2019, 23, 2711–2718. [Google Scholar] [CrossRef]
- Watabe, T.; Kaneda-Nakashima, K.; Shirakami, Y.; Liu, Y.; Ooe, K.; Teramoto, T.; Toyoshima, A.; Shimosegawa, E.; Nakano, T.; Kanai, Y.; et al. Targeted Alpha Therapy Using Astatine (211At)-Labeled Phenylalanine: A Preclinical Study in Glioma Bearing Mice. Oncotarget 2020, 11, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, S.I.; Kawabata, S.; Hiramatsu, R.; Kuroiwa, T.; Suzuki, M.; Kondo, N.; Ono, K. Boron Neutron Capture Therapy for Malignant Brain Tumors. Neurol. Med. Chir. 2016, 56, 361. [Google Scholar] [CrossRef]
- Ohnishi, K.; Misawa, M.; Sikano, N.; Nakai, K.; Suzuki, M. Enhancement of Cancer Cell-Killing Effects of Boron Neutron Capture Therapy by Manipulating the Expression of L-Type Amino Acid Transporter 1. Radiat. Res. 2021, 196, 17–22. [Google Scholar] [CrossRef]
- Fujimura, A.; Yasui, S.; Igawa, K.; Ueda, A.; Watanabe, K.; Hanafusa, T.; Ichikawa, Y.; Yoshihashi, S.; Tsuchida, K.; Kamiya, A.; et al. In Vitro Studies to Define the Cell-Surface and Intracellular Targets of Polyarginine-Conjugated Sodium Borocaptate as a Potential Delivery Agent for Boron Neutron Capture Therapy. Cells 2020, 9, 2149. [Google Scholar] [CrossRef]
- Luo, L.; Wang, J.; Ding, D.; Hasan, M.N.; Yang, S.-S.; Lin, S.-H.; Schreppel, P.; Sun, B.; Yin, Y.; Erker, T.; et al. Role of NKCC1 Activity in Glioma K+ Homeostasis and Cell Growth: New Insights With the Bumetanide-Derivative STS66. Front. Physiol. 2020, 11, 911. [Google Scholar] [CrossRef]
- Luo, L.; Guan, X.; Begum, G.; Ding, D.; Gayden, J.; Hasan, M.N.; Fiesler, V.M.; Dodelson, J.; Kohanbash, G.; Hu, B.; et al. Blockade of Cell Volume Regulatory Protein NKCC1 Increases TMZ-Induced Glioma Apoptosis and Reduces Astrogliosis. Mol. Cancer Ther. 2020, 19, 1550–1561. [Google Scholar] [CrossRef]
- Juraszek, B.; Czarnecka-Herok, J.; Nałęcz, K.A. Glioma Cells Survival Depends Both on Fatty Acid Oxidation and on Functional Carnitine Transport by SLC22A5. J. Neurochem. 2021, 156, 642–657. [Google Scholar] [CrossRef]
- Nachef, M.; Ali, A.K.; Almutairi, S.M.; Lee, S.H. Targeting SLC1A5 and SLC3A2/SLC7A5 as a Potential Strategy to Strengthen Anti-Tumor Immunity in the Tumor Microenvironment. Front. Immunol. 2021, 12, e624324. [Google Scholar] [CrossRef]
- Jyotsana, N.; Ta, K.T.; DelGiorno, K.E. The Role of Cystine/Glutamate Antiporter SLC7A11/XCT in the Pathophysiology of Cancer. Front. Oncol. 2022, 12, 858462. [Google Scholar] [CrossRef]
- Lagana, A.; Vadnais, J.; Le, P.U.; Nguyen, T.N.; Laprade, R.; Nabi, I.R.; Noel, J. Regulation of the Formation of Tumor Cell Pseudopodia by the Na(+)/H(+) Exchanger NHE1. J. Cell Sci. 2000, 113 Pt 20, 3649–3662. [Google Scholar] [CrossRef]
- Pardridge, W.M. Blood-Brain Barrier Endogenous Transporters as Therapeutic Targets: A New Model for Small Molecule CNS Drug Discovery. Expert Opin. Ther. Targets 2015, 19, 1059–1072. [Google Scholar] [CrossRef]
- Tashima, T. Intriguing Possibilities and Beneficial Aspects of Transporter-Conscious Drug Design. Bioorg. Med. Chem. 2015, 23, 4119–4131. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anagnostakis, F.; Kokkorakis, M.; Markouli, M.; Piperi, C. Impact of Solute Carrier Transporters in Glioma Pathology: A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 9393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119393
Anagnostakis F, Kokkorakis M, Markouli M, Piperi C. Impact of Solute Carrier Transporters in Glioma Pathology: A Comprehensive Review. International Journal of Molecular Sciences. 2023; 24(11):9393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119393
Chicago/Turabian StyleAnagnostakis, Filippos, Michail Kokkorakis, Mariam Markouli, and Christina Piperi. 2023. "Impact of Solute Carrier Transporters in Glioma Pathology: A Comprehensive Review" International Journal of Molecular Sciences 24, no. 11: 9393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119393