

Phytochemical, Morphological, and Physiological Variation in Different Ajowan (Trachyspermum ammi L.) Populations as Affected by Salt Stress, Genotype × Year Interaction and Pollination System
 ,
, 

Abstract
:1. Introduction
2. Results
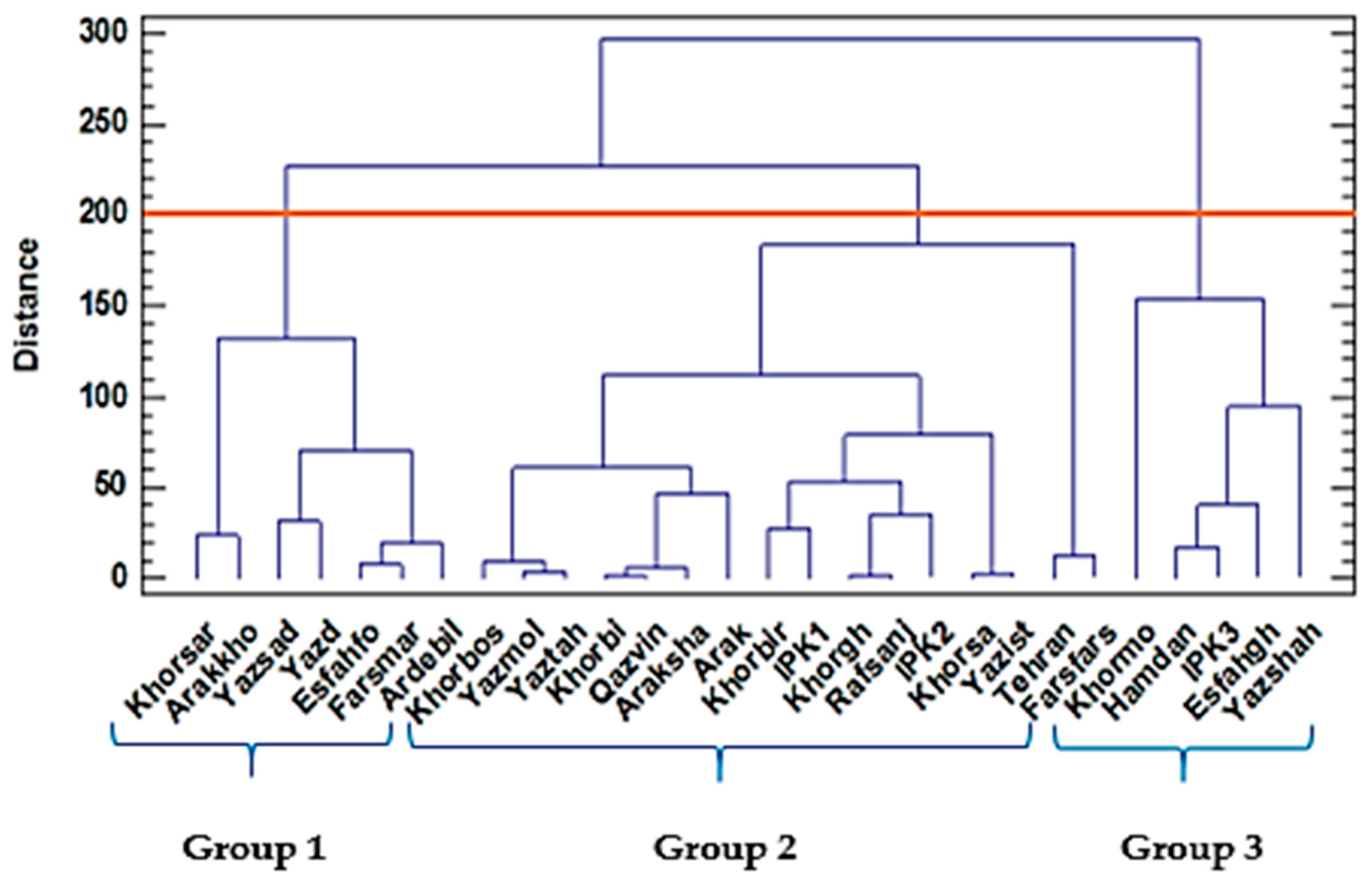
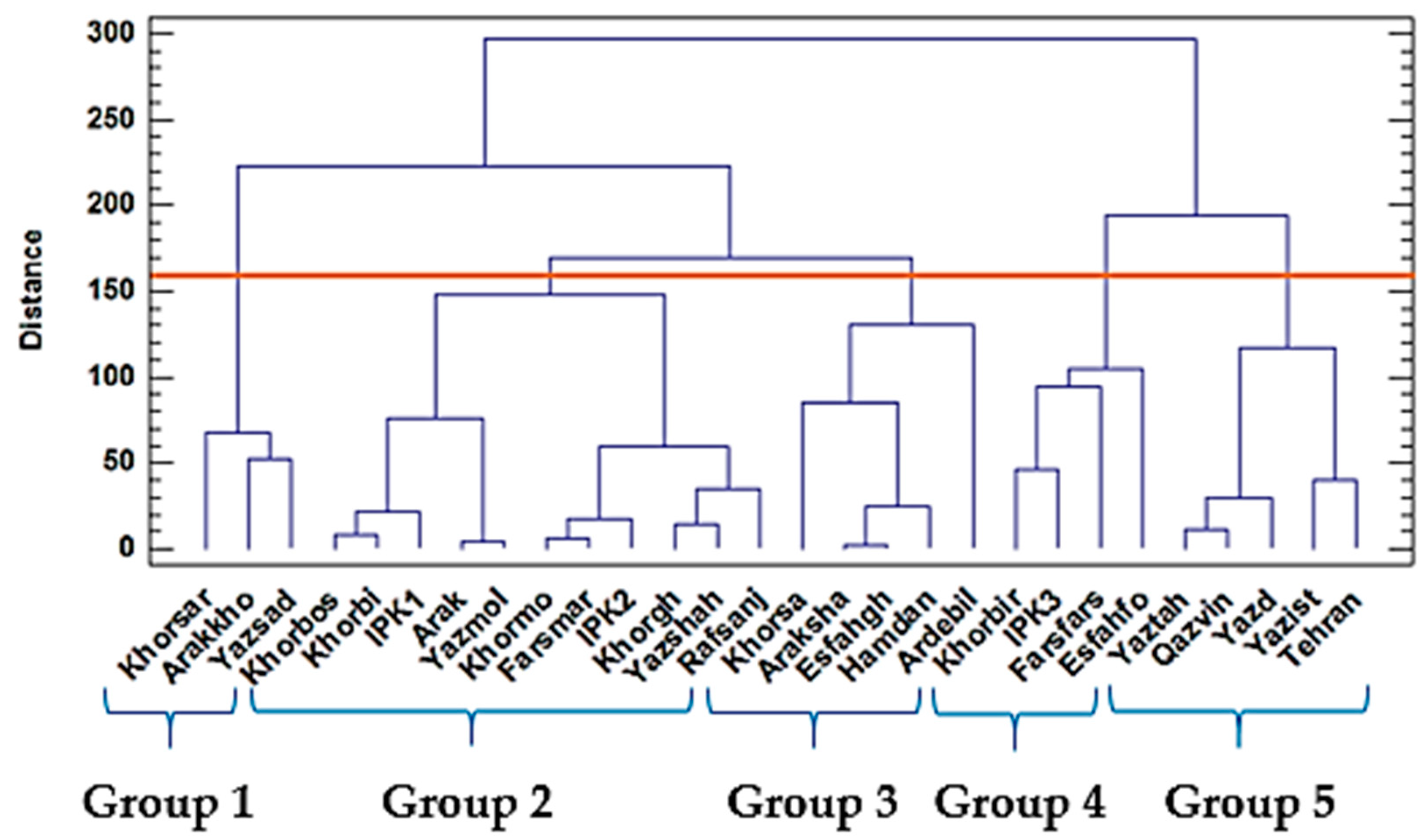
2.1. Two-Year Study of Morphological, Oil, and Yield-Related Traits of 28 Populations
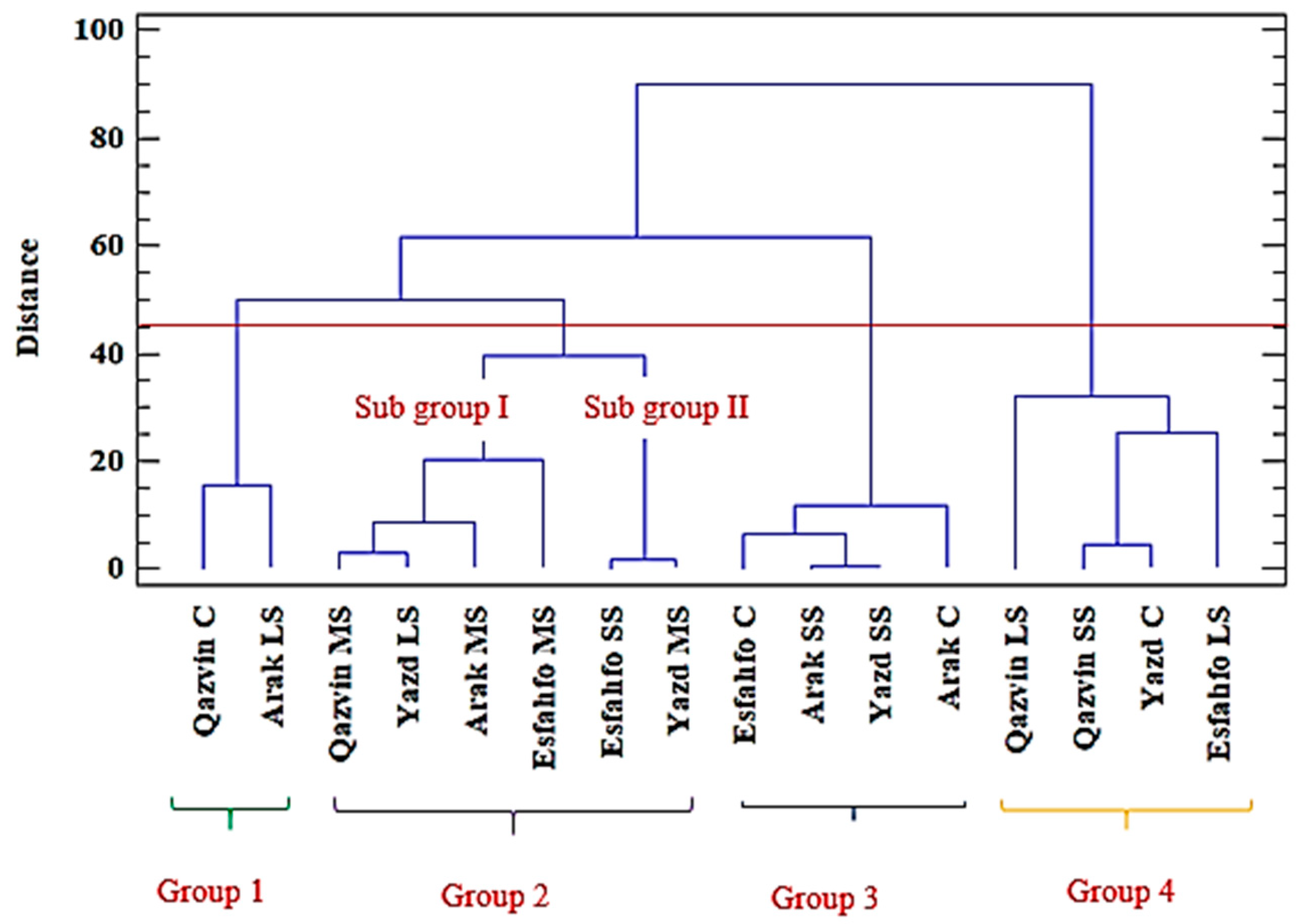
Hierarchical Cluster Analysis
2.2. Effects of Salt Stress on Morphophysiological, and Phytochemical Traits
2.2.1. Essential Oil Content
2.2.2. Essential Oil Composition
2.2.3. Yield-Related Traits
2.2.4. Total Phenolic and Flavonoid Contents
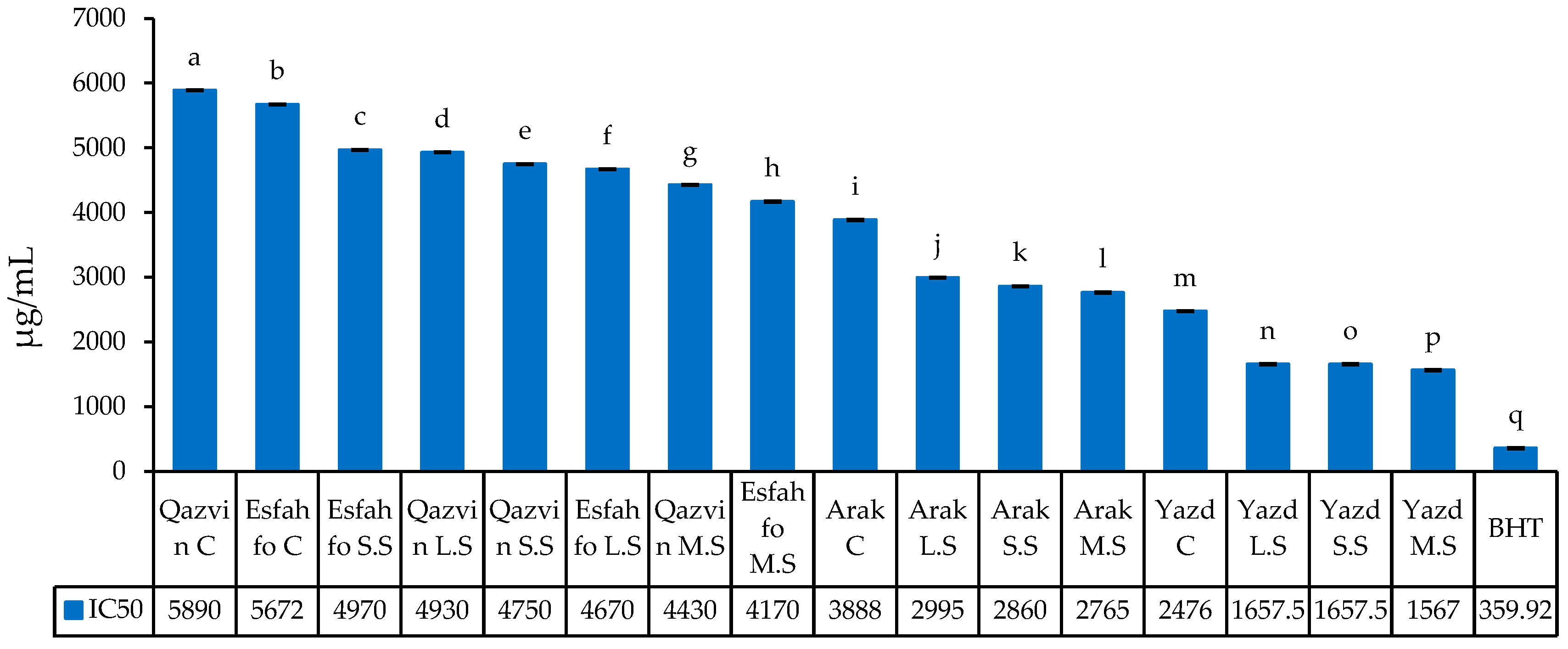
2.2.5. DPPH Radical Scavenging
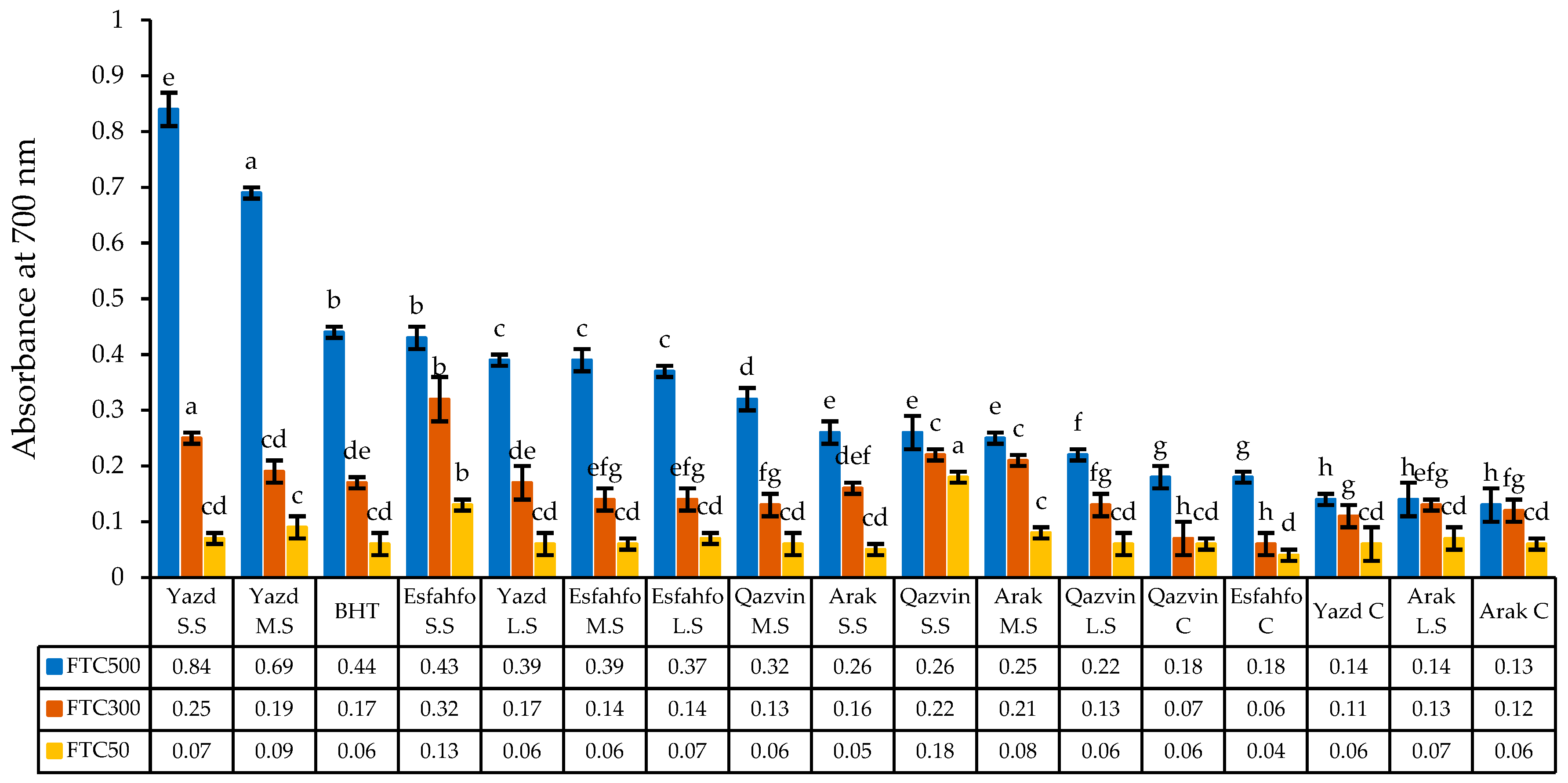
2.2.6. Reducing Power
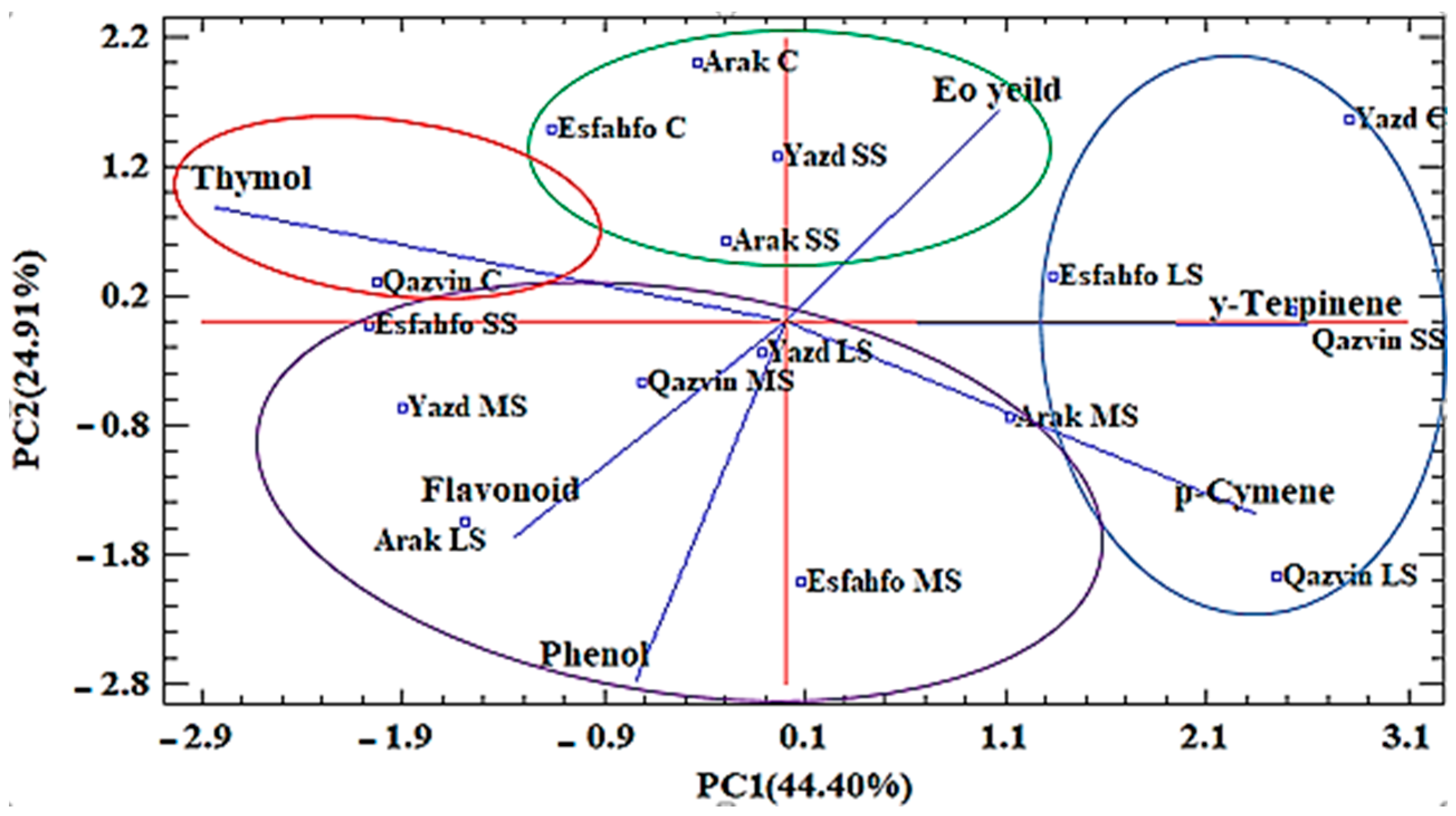
2.2.7. Cluster and Principal Component (PCA) Analysis
2.2.8. Correlations among the Components
2.2.9. Physiological Traits
Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2)
Antioxidant Enzymes Activity
Carotenoids
2.2.10. MANOVA Analysis
2.3. Morphological Traits in Self- and Open-Pollinated Populations
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Salt Stress Treatments
4.3. Essential Oil Extraction
4.4. GC/MS Analysis
4.5. Flavonoid and Phenolic Evaluation
4.6. Antioxidant Capacity
4.6.1. DPPH Assay
4.6.2. Reducing Power
4.7. Malondialdehyde (MDA) Content
4.8. Hydrogen Peroxide (H2O2) Content
4.9. Ascorbate Peroxidase (APX) Activity
4.10. Guaiacol Peroxidase (GPX) Activity
4.11. Protein Assay
4.12. Measurement of Chlorophyll Content
4.13. Self-Pollination Studies
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niazian, M.; Sadat-Noori, S.A.; Tohidfar, M.; Galuszka, P.; Mortazavian, S.M.M. Agrobacterium-mediated genetic transformation of ajowan Trachyspermum ammi (L.) Sprague: An important industrial medicinal plant. Ind. Crops Prod. 2019, 132, 29–40. [Google Scholar] [CrossRef]
- Rahimmalek, M.; Heidari, E.F.; Ehtemam, M.H.; Mohammadi, S. Essential oil variation in Iranian ajowan Trachyspermum ammi (L.) Sprague populations collected from different geographical regions in relation to climatic factors. Ind. Crops Prod. 2017, 95, 591–598. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Trindade, H. Review on essential oil, extracts composition, molecular and phytochemical properties of Thymus species in Iran. Ind. Crops Prod. 2019, 134, 89–99. [Google Scholar] [CrossRef]
- Mirniyam, G.; Rahimmalek, M.; Arzani, A.; Matkowski, A.; Gharibi, S.; Szumny, A. Changes in Essential Oil Composition, Polyphenolic Compounds and Antioxidant Capacity of Ajowan (Trachyspermum ammi L.) Populations in Response to Water Deficit. Foods 2022, 11, 3084. [Google Scholar] [CrossRef] [PubMed]
- Kiani, R.; Arzani, A.; MirmohammadyMaibody, S.A.M. Polyphenols, Flavonoids, and Antioxidant Activity Involved in Salt Tolerance in Wheat, Aegilops cylindrica and Their Amphidiploids. Front. Plant Sci. 2021, 12, 493. [Google Scholar] [CrossRef]
- Heidari, E.F.; Rahimmalek, M.; Mohammadi, S.; Ehtemam, M.H. Genetic structure and diversity of ajowan (Trachyspermum ammi) populations based on molecular, morphological markers, and volatile oil content. Ind. Crops Prod. 2016, 92, 186–196. [Google Scholar] [CrossRef]
- Goyal, S.; Chaturvedi, V.; Dhingra, G.; Tanwar, S.; Sharma, K.; Singh, S. Trachyspermum ammi: A review on traditional and modern pharmacological aspects. Biol. Sci. 2022, 2, 324–337. [Google Scholar] [CrossRef]
- Vafadar Shoshtari, Z.; Rahimmalek, M.; Sabzalian, M.R.; Hosseini, H. Essential oil and bioactive compounds variation in myrtle (Myrtus communis L.) as affected by seasonal variation and salt stress. Chem. Biodivers. 2017, 14, e1600365. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liang, C.; Chen, J.; Qi, X.; Liu, Y.; Li, W. The effects of salinity stress on morphological characteristics, mineral nutrient accumulation and essential oil yield and composition in Mentha canadensis (L.). Sci. Hortic. 2015, 197, 579–583. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris (L.) and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V.; Niazi, A.; Moghadam, A. Effect of salt stress on terpenoid biosynthesis in Salvia mirzayanii: From gene tometabolite. J. Hortic. Sci. Biotechnol. 2018, 94, 389–399. [Google Scholar] [CrossRef]
- Wani, I.A.; Verma, S.; Ahmad, P.; El-Serehy, H.A.; Hashim, M.J. Reproductive Biology of Rheum webbianum Royle, a Vulnerable Medicinal Herb From Alpines of North-Western Himalaya. Front. Plant Sci. 2022, 13, 699645. [Google Scholar] [CrossRef]
- Richardson, M.L.; Keathley, C.P.; Peterson, C.L. Breeding system of the critically endangered Lakela’s Mint and influence of plant height on pollinators and seed output. Popul. Ecol. 2016, 58, 277–284. [Google Scholar] [CrossRef]
- Mooney, E.H.; McGraw, J.B. Effects of self-pollination and outcrossing with cultivated plants in small natural populations of American ginseng, Panax quinquefolius (Araliaceae). Am. J. Bot. 2007, 94, 1677–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudeul, M.; Till-Bottraud, I. Reproductive ecology of the endangered alpine species Eryngium alpinum L. (Apiaceae): Phenology, gene dispersal and reproductive success. Ann. Bot. 2004, 93, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Davila, Y.C.; Wardle, G.M. Bee boys and fly girls: Do pollinators prefer male or female umbels in protandrous parsnip, Trachymene incisa (Apiaceae). Austral Ecol. 2007, 32, 798–807. [Google Scholar] [CrossRef]
- Salami, M.; Rahimmalek, M.; Ehtemam, M.H.; Sabzalian, M.R. Variation in bioactive contents and anatomical characteristics of different fennel (Foeniculum vulgare Mill.) populations as affected by self-pollination. J. Appl. Bot. Food Qual. 2016, 89, 38–48. [Google Scholar] [CrossRef]
- Nazem, V.; Sabzalian, M.R.; Saeidi, G.; Rahimmalek, M. Essential oil yield and composition and secondary metabolites in self-and open-pollinated populations of mint (Mentha spp.). Ind. Crops Prod. 2019, 130, 332–340. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elshafiey, E.H.; Shetaia, A.A.; El-Wahed, A.A.A.; Algethami, A.F.; Musharraf, S.G.; AlAjmi, M.F.; Zhao, C.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Overview of Bee Pollination and Its Economic Value for Crop Production. Insects 2021, 12, 688. [Google Scholar] [CrossRef]
- Yadav, N.; Shakya, P.; Kumar, A.; Gautam, R.D.; Chauhan, R.; Kumar, D.; Kumar, A.; Singh, S.; Singh, S. Investigation on pollination approaches, reproductive biology and essential oil variation during floral development in German chamomile (Matricaria chamomilla L.). Sci. Rep. 2022, 12, 15285. [Google Scholar] [CrossRef]
- Pandey, M.M.; Vijayakumar, M.; Rastogi, S.; Rawat, A.K.S. Phenolic content and antioxidant properties of selected Indian spices of Apiaceae. J. Herbs Spices Med. Plants 2012, 18, 246–256. [Google Scholar] [CrossRef]
- Zandi, M.; Khalesro, S.; Sharifi, Z. Improved yields and essential oil composition of ajowan (Trachyspermum ammi L.) and soil fertility properties in intercropping systems. Biol. Agric. Hortic. 2023, 39, 1–18. [Google Scholar] [CrossRef]
- Shafeiee, M.; Ehsanzadeh, P. Physiological and biochemical mechanisms of salinity tolerance in several fennel genotypes: Existence of clearly-expressed genotypic variations. Ind. Crops Prod. 2019, 132, 311–318. [Google Scholar] [CrossRef]
- Davazdahemami, S.; Allahdadi, M. Essential oil yield and composition of four annual plants (ajowan, dill, moldavian balm and black cumin) under saline irrigation. Food Ther. Health Care 2022, 4, 5. [Google Scholar] [CrossRef]
- Bairwa, R.; Sodha, R.S.; Rajawat, B.S. Trachyspermum ammi. Pharmacogn. Rev. 2012, 6, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Mirzahosseini, S.M.; Noori, S.A.S.; Amanzadeh, Y.; Javid, M.G.; Howyzeh, M.S. Phytochemical assessment of some native ajowan Therachyspermum ammi (L.) ecotypes in Iran. Ind. Crops Prod. 2017, 105, 142–147. [Google Scholar] [CrossRef]
- Neffati, M.; Sriti, J.; Hamdaoui, G.; Kchouk, M.E.; Marzouk, B. Salinity impact on fruit yield, essential oil composition and antioxidant activities of Coriandrum sativum fruit extracts. Food Chem. 2011, 124, 221–225. [Google Scholar] [CrossRef]
- Moein, M.R.; Zomorodian, K.; Pakshir, K.; Yavari, F.; Motamedi, M.; Zarshenas, M.M. Trachyspermum ammi L. sprague: Chemical composition of essential oil and antimicrobial activities of respective fractions. J. Evid. Based Complement. Altern. Med. 2015, 20, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Davazdahemami, S.; Sefidkon, F.; Jahansooz, M.R.; Mazaheri, D. Chemical Composition of the Essential Oils from Foliages and Seeds of ajowan (Trachyspermum ammi (L.) Sprague) in two planting dates (spring and summer). J. Essent. Oil Bear. Plants 2011, 14, 639–642. [Google Scholar] [CrossRef]
- Serraj, R.; Drevon, J.J. Effects of salinity and nitrogen source on growth and nitrogen fixation in alfalfa. J. Plant Nutr. 1998, 21, 1805–1818. [Google Scholar] [CrossRef]
- Esechie, H.A.; Al-Saidi, A.; Al-Khanjari, S. Effect of sodium chloride salinity on seedling emergence in chickpea. J. Agron. Crop Sci. 2002, 188, 155–160. [Google Scholar] [CrossRef]
- Gibberd, M.R.; Turner, N.C.; Storey, R. Influence of saline irrigation on growth, ion accumulation and partitioning, and leaf gas exchange of carrot (Daucus carota L.). Ann. Bot. 2002, 90, 715–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajar, A.S.; Zidan, M.A.; AlZahrani, H.S. Effect of salinity stress on the germination, growth and some physiological activities of black cumin (Nigella sativa L). Arab. Gulf J. Sci. Res. 1996, 14, 445–454. [Google Scholar]
- Dastan, D.; Salehi, P.; Maroofi, H. Chemical composition, antioxidant, and antimicrobial activities on Laserpitium carduchorum Hedge & Lamond essential oil and extracts during various growing stages. Chem. Biodivers. 2016, 13, 1397–1403. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V. Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. S. Afr. J. Bot. 2014, 93, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Ghassemi-Golezani, K.; Abdoli, S. Physiological and Biochemical Responses of Medicinal Plants to Salt Stress. In Environmental Challenges and Medicinal Plants. Environmental Challenges and Solutions; Aftab, T., Ed.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Lopez-Berenguer, C.; Martinez-Ballesta, M.; Del, C.; Moreno, D.A.; Carvajal, M.; Garcia-Viguera, C. Growing hardier crops for better health: Salinity tolerance and the nutritional value of broccoli. J. Agric. Food Chem. 2009, 57, 572–578. [Google Scholar] [CrossRef]
- Hichem, H.; Mounir, D. Differential responses of two maize Zea mays (L.) varieties to salt stress: Changes on polyphenols composition of foliage and oxidative damages. Ind. Crops Prod. 2009, 30, 144–151. [Google Scholar] [CrossRef]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Chauhan, B.; Kumar, G.; Ali, M. A review on phytochemical constituents and activities of Trachyspermum ammi (L.) Sprague fruits. Am. J. Pharmtech. Res. 2012, 2, 329–340. [Google Scholar]
- Moukhles, A.; Ellaghdach, A.; Driss, A.B.; El Amrani, M.A.; Aghmiz, A.; Mansour, A.I. Chemical profile and in vitro Antibacterial potential of Essential Oils and Hydrolat Extracts from Aerial Parts of Three Wild species of Moroccan Thymus. Sci. Afr. 2022, 18, e01434. [Google Scholar] [CrossRef]
- Ang, L.Z.P.; Hashim, R.; Sulaiman, S.F.; Coulibaly, A.Y.; Sulaiman, O.; Kawamura, F.; Salleh, K.M. In vitro antioxidant and antidiabetic activites of Gluta torquata. Ind. Crops Prod. 2015, 76, 755–760. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Arzani, A. Essential oil composition, total phenolic, flavonoid contents, and antioxidant activity of Thymus species collected from different regions of Iran. Food Chem. 2017, 220, 153–161. [Google Scholar] [CrossRef]
- Akbarian, A.; Rahimmalek, M.; Sabzalian, M.R. Variation in essential oil yield and composition of Dorema aucheri Boiss., an endemic Medicinal plant collected from wild populations in natural habitats. Chem. Biodivers. 2016, 13, 1756–1766. [Google Scholar] [CrossRef] [PubMed]
- Afshari, M.; Rahimmalek, M. Variation in essential oil composition, bioactive compounds, anatomical and antioxidant activity of Achillea aucheri, an endemic species of Iran, at different phenological stages. Chem. Biodivers. 2018, 15, e1800319. [Google Scholar] [CrossRef]
- Mohammadi, S.; Saharkhiz, M.J.; Javanmardi, J. Evaluation of interaction effects of spermidine and salinity on physiological and morphology trait of catnip Nepeta cataria (L.). Z. Arznei-Gewürzpflanzen 2017, 22, 104–109. [Google Scholar]
- Crocoll, C.; Asbach, J.; Novak, J.; Gershenzon, J.; Degenhardt, J. Terpene synthases of oregano Origanum vulgare (L.) and their roles in the pathway and regulation of terpene biosynthesis. Plant Mol. Biol. 2010, 73, 587–603. [Google Scholar] [CrossRef]
- Sarfaraz, D.; Rahimmalek, M.; Sabzalian, M.R.; Gharibi, S.; Matkowski, A.; Szumny, A. Essential Oil Composition and Antioxidant Activity of Oregano and Marjoram as Affected by Different Light-Emitting Diodes. Molecules 2023, 28, 3714. [Google Scholar] [CrossRef]
- Bazrafshan, A.H.; Ehsanzadeh, P. Evidence for differential lipid peroxidation and antioxidant enzyme activities in Sesamum indicum L. genotypes under NaCl salinity. J. Agric. Sci. Technol. 2016, 18, 202–222. [Google Scholar]
- Elkahoui, S.; Hernandez, J.A.; Abdelly, C.; Ghrir, R.; Limam, F. Effects of salt on lipid peroxidation and antioxidant enzyme activities of Catharanthus roseus suspension cells. Plant Sci. 2005, 168, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Santini, R.; De Lima, J.P.; Gratao, P.L.; Camargo, A.F.M. Evaluation of growth and oxidative stress as indicative of salinity tolerance by the invasive tropical aquatic macrophyte tanner grass. Hydrobiologia 2022, 849, 1261–1271. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Shannon, M.C. Adaptation of plants to salinity. Adv. Agron. 1998, 60, 75–120. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Khanahmadi, F.; Rahimmalek, M.; Sabzalian, M.R.; Tohidi, B. Variation of essential oil content and composition, phenolics, and yield related traits using different pollination systems in populations of Thymus species. Chem. Biodivers. 2021, 18, e2100097. [Google Scholar] [CrossRef]
- Vassiliou, E.; Awoleye, O.; Davis, A.; Mishra, S. Anti-Inflammatory and Antimicrobial Properties of Thyme Oil and Its Main Constituents. Int. J. Mol. Sci. 2023, 24, 6936. [Google Scholar] [CrossRef]
- Asif, H.M.; Hashmi, H.A.S. Bioactive Compounds of Ajwain (Trachyspermum ammi [L.] Sprague). In Bioactive Compounds in Underutilized Vegetables and Legumes; Reference Series in Phytochemistry; Murthy, H.N., Paek, K.Y., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–18. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech. F. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kim, J.-Y. Understanding Wheat Starch Metabolism in Properties, Environmental Stress Condition, and Molecular Approaches for Value-Added Utilization. Plants 2021, 10, 2282. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Azevedo Neto, A.D.; Prisco, J.T.; Eneas-Filho, J.; De Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. J. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Shahid, S.; Ali, Q.; Ali, S.; Al-Misned, F.A.; Maqbool, S. Water Deficit Stress Tolerance Potential of Newly Developed Wheat Genotypes for Better Yield Based on Agronomic Traits and Stress Tolerance Indices: Physio-Biochemical Responses, Lipid Peroxidation and Antioxidative Defense Mechanism. Plants 2022, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lee, H. Salinity-Triggered Responses in Plant Apical Meristems for Developmental Plasticity. Int. J. Mol. Sci. 2023, 24, 6647. [Google Scholar] [CrossRef] [PubMed]
- Hammami, H.; Saadatian, B.; Aliverdi, A. Geographical variation in breaking the seed dormancy of Persian cumin (Carum carvi L.) ecotypes and their physiological responses to salinity and drought stresses. Ind. Crops Prod. 2018, 124, 600–606. [Google Scholar] [CrossRef]
- Nourimand, M.; Mohsenzadeh, S.; Jaime Teixeira da Silva, A. Physiological responses of fennel seedling to four environmental stresses. Iran J. Sci. Technol. 2012, 36, 37–46. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Biotechnol. Appl. Biochem. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Baharfar, R.; Azimi, R.; Mohseni, M. Antioxidant and antibacterial activity of flavonoid-, polyphenol-and anthocyanin-rich extracts from Thymus kotschyanus boiss & hohen aerial parts. J. Food Sci. Technol. 2015, 52, 6777–6783. [Google Scholar] [CrossRef] [Green Version]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H.; Talebi, M. Total phenolic content and antioxidant activity of three Iranian endemic Achillea species. Ind. Crops Prod. 2013, 50, 154–158. [Google Scholar] [CrossRef]
- Taulavuori, E.; Hellstrom, E.; Taulavuori, K.; Laine, K. Comparison of two methods used to analyse lipid peroxidation from Vaccinium myrtillus (L.) during snow removal, reacclimation and cold acclimation. J. Exp. Bot. 2001, 52, 2375–2380. [Google Scholar] [CrossRef] [Green Version]
- Genc, Y.; Mcdonald, G.K.; Tester, M. Reassessment of tissue Na+ concentration as a criterion for salinity tolerance in bread wheat. Plant Cell Environ. 2007, 30, 1486–1498. [Google Scholar] [CrossRef]
- Dazy, M.; Beraud, E.; Cotelle, S.; Meux, E.; Masfaraud, J.F.; Ferard, J.F. Antioxidant enzyme activities as affected by trivalent and hexavalent chromium species in Fontinalis antipyretica Hedw. Chemosphere 2008, 73, 281–290. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; pp. 764–775. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Means of Square | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | df | Plant Height | No. of Flowering Branches | Inflorescence Diameter | No. of Umbels | No. of Umbellule per Inflorescence | No. of Flowers per Umbel | No. of Seeds per Umbel | Crown Cover Diameter | One Thousand Seed Weight | Seed Yield per Plant | Essential Oil Yield |
| Year (Y) | 1 | 24396.01 ** | 2648.14 ** | 60.84 ** | 785321.11 ** | 1618.82 ** | 840793.21 ** | 336,3172 ns | 357,643.06 ** | 0.0370 ** | 386,539.88 ** | 1.040 ** |
| Rep (year) | 4 | 195.21 | 23.70 | 0.40 | 9048.53 | 5.86 | 3063.36 | 12,253.47 | 228,663.37 | 0.0006 | 4356.98 | 0.045 |
| G | 27 | 381.72 ** | 110.35 ** | 0.80 ** | 25,217.35 ** | 16.58 ** | 21,926.23 ** | 87,704.90 ** | 596,830.26 ** | 0.0500 ** | 10,394.04 ** | 6.360 ** |
| Y × G | 27 | 323.70 ** | 130.90 ** | 0.92 ** | 24389.70 ** | 23.23 ** | 23,900.90 ** | 95,603.80 ** | 4386.02 ns | 0.0001 ns | 10,673.01 ** | 0.036 ns |
| Residual | 108 | 99.51 | 8.62 | 0.31 | 2260.86 | 4.39 | 3868.95 | 15475 | 36513 | 0.0003 | 1517.52 | 0.0531 |
| Genotype | Height (cm) | No. of Flowering Branches | Inflorescence Diameter | No. of Umbels | No. of Umbellule per Inflorescence | No. of Flowers per Umbel | No. of Seeds per Umbel | Crown Cover Diameter (cm2) | One Thousand Seed Weight (g) | Seed Yield per Plant | Essential Oil Yield (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| 1 | 87 | 109.83 | 14.5 | 18.1 | 3.91 | 5.5 | 58 | 105.2 | 13 | 18.5 | 219.83 | 337.5 | 439.67 | 675 | 1462.7 | 1896.9 | 0.72 | 0.75 | 31.93 | 80.31 | 4.35 | 4.51 |
| 2 | 84.1 | 107.83 | 14.16 | 11.6 | 3.16 | 3.7 | 76 | 262.7 | 11.83 | 17.1 | 172.67 | 164.5 | 345.33 | 329 | 351.5 | 569.8 | 0.87 | 0.92 | 52.48 | 86.37 | 4.35 | 4.47 |
| 3 | 91.6 | 115.83 | 12.66 | 43 | 3 | 4.2 | 78.67 | 389.3 | 10.833 | 21 | 139.5 | 471.5 | 279 | 943 | 741.2 | 1056 | 0.76 | 0.79 | 27.81 | 286.8 | 3.63 | 4.13 |
| 4 | 77.1 | 139.67 | 14.66 | 18.3 | 2.83 | 4.5 | 102.83 | 199 | 13.16 | 20 | 180.5 | 232.8 | 361 | 465.7 | 449.1 | 697.1 | 0.58 | 0.61 | 21.43 | 61.94 | 3.01 | 3.16 |
| 5 | 77.6 | 101.33 | 12.66 | 15 | 2.66 | 4.5 | 82.5 | 167 | 13.16 | 15 | 198.33 | 205.3 | 396.67 | 410.7 | 278.6 | 489.1 | 0.8 | 0.82 | 41.31 | 58.44 | 4.36 | 4.43 |
| 6 | 100.5 | 104.5 | 20.83 | 26.1 | 2.75 | 4.1 | 273.39 | 135 | 13 | 19.5 | 197.83 | 349.1 | 395.67 | 698.3 | 688 | 987.9 | 0.72 | 0.75 | 72.45 | 71.39 | 4.09 | 4.16 |
| 7 | 88.33 | 107.33 | 12.5 | 15.6 | 3.41 | 4.4 | 140 | 162.8 | 13.16 | 15.8 | 192.67 | 280 | 385.33 | 560 | 778.7 | 1097.4 | 0.7 | 0.73 | 32.09 | 67.61 | 2.79 | 2.9 |
| 8 | 87.33 | 108.67 | 10.5 | 16.8 | 3.5 | 4.1 | 57.5 | 127.6 | 13.66 | 20.6 | 219.83 | 264 | 439.67 | 528 | 1548.7 | 1906.2 | 0.92 | 0.96 | 43.38 | 63.6 | 5.23 | 5.2 |
| 9 | 76 | 129.83 | 10 | 10.5 | 2.16 | 4.1 | 65.5 | 147.3 | 12 | 18.8 | 137.5 | 275.6 | 275 | 551.3 | 563.9 | 833.2 | 0.83 | 0.86 | 15.53 | 71.9 | 5.08 | 5.13 |
| 10 | 67.83 | 116 | 11.16 | 14.5 | 2.65 | 4 | 88.33 | 342 | 12.83 | 18.5 | 204.33 | 324 | 408.67 | 648 | 491.9 | 755.7 | 0.77 | 0.8 | 34.7 | 187.27 | 4.61 | 4.66 |
| 11 | 95.16 | 109.5 | 14 | 10.5 | 3.38 | 4.3 | 143 | 413.3 | 16 | 18.5 | 273.17 | 362.1 | 546.33 | 724.3 | 458.3 | 705.6 | 0.73 | 0.75 | 58.53 | 228.05 | 2.78 | 2.98 |
| 12 | 85.66 | 112.66 | 12.33 | 12.5 | 3.13 | 3.8 | 132.5 | 173.3 | 12 | 20.3 | 180.67 | 275.1 | 361.33 | 550.3 | 669.6 | 969.5 | 0.91 | 0.94 | 39.11 | 91.9 | 4.61 | 4.69 |
| 13 | 85.16 | 104.66 | 14 | 25.5 | 2.86 | 5.2 | 110.83 | 204.1 | 12 | 22.6 | 172.83 | 360.5 | 345.67 | 721 | 626.4 | 905.1 | 0.85 | 0.87 | 32.19 | 128.71 | 4.03 | 4.15 |
| 14 | 74.5 | 105.33 | 10 | 28 | 3.08 | 4.8 | 81.33 | 194.5 | 15.16 | 15.6 | 193.67 | 273.3 | 387.33 | 546.7 | 771.1 | 1059.9 | 0.96 | 0.98 | 20.46 | 106.39 | 4.72 | 4.85 |
| 15 | 84 | 91.5 | 7.66 | 19.5 | 3.91 | 3.8 | 61.5 | 154.8 | 13.83 | 16.3 | 282.67 | 236 | 565.33 | 472 | 606.7 | 886.2 | 0.63 | 0.66 | 22.19 | 51.56 | 1.62 | 1.74 |
| 16 | 72.66 | 107.16 | 12.16 | 11.5 | 2.83 | 4.5 | 130.25 | 250.5 | 11.83 | 23.8 | 171.17 | 470 | 342.33 | 940 | 537 | 832.8 | 0.58 | 0.61 | 23.46 | 127.19 | 3.43 | 3.53 |
| 17 | 75.5 | 111.83 | 7.83 | 13.1 | 3.25 | 4.8 | 29.83 | 235.8 | 15.83 | 20.6 | 253.67 | 404.8 | 507.33 | 809.7 | 354.4 | 584.9 | 0.92 | 0.96 | 14.16 | 177.6 | 5.51 | 5.58 |
| 18 | 103.33 | 118.83 | 17.66 | 15.3 | 3.75 | 3.7 | 145.65 | 372.1 | 13.5 | 16.6 | 263.33 | 211.3 | 526.67 | 422.7 | 357.9 | 585.5 | 0.7 | 0.72 | 69.33 | 114.77 | 4.23 | 4.37 |
| 19 | 75.5 | 100 | 10.5 | 20.5 | 3.4 | 4.1 | 86 | 326.8 | 13.83 | 29.6 | 217.5 | 614 | 435 | 1228 | 694.9 | 1021.4 | 0.74 | 0.77 | 24.11 | 315.02 | 5.32 | 5.52 |
| 20 | 77.33 | 97.83 | 10.83 | 25.5 | 2.91 | 5.1 | 102.33 | 188.1 | 14.33 | 22.3 | 192 | 510.1 | 384 | 1020.3 | 417.5 | 672 | 0.78 | 0.8 | 25.71 | 159.55 | 4.73 | 4.94 |
| 21 | 76.5 | 95.83 | 8 | 18.5 | 2.25 | 5.5 | 56.33 | 80.6 | 11.5 | 20.5 | 125.5 | 444.3 | 251 | 888.7 | 644.8 | 954.6 | 0.71 | 0.74 | 9.85 | 53.59 | 2.07 | 2.28 |
| 22 | 66.16 | 112.66 | 6.16 | 42.6 | 3.25 | 4.1 | 43.5 | 550.3 | 13.83 | 20.1 | 205.67 | 238.8 | 411.33 | 477.7 | 625.3 | 896.1 | 0.67 | 0.7 | 12.13 | 171.82 | 3.8 | 3.91 |
| 23 | 83.16 | 104.5 | 8.66 | 15.3 | 3.25 | 3.8 | 44.09 | 134.5 | 14 | 16.5 | 238.83 | 397.6 | 477.67 | 795.3 | 358.7 | 612.5 | 0.75 | 0.78 | 20.09 | 74.06 | 4.34 | 4.4 |
| 24 | 63.66 | 88.5 | 7.66 | 27.6 | 2.33 | 3.9 | 61 | 399.8 | 9.33 | 24.6 | 114 | 493.3 | 228 | 986.7 | 432.9 | 848.8 | 0.7 | 0.73 | 10.61 | 290.74 | 1.2 | 2 |
| 25 | 87.66 | 93.33 | 13 | 21.6 | 3.08 | 3.3 | 163.67 | 344 | 12 | 17 | 175.67 | 302.8 | 351.33 | 605.7 | 755.2 | 1098.7 | 0.72 | 0.75 | 40.72 | 118.15 | 3.53 | 3.67 |
| 26 | 98 | 104.66 | 10.16 | 12.5 | 3.33 | 3.3 | 60.67 | 76 | 14.16 | 15.5 | 136.5 | 164.6 | 273 | 329.3 | 434.8 | 733.9 | 0.71 | 0.76 | 11.6 | 19.3 | 4.04 | 4.18 |
| 27 | 85.5 | 87.83 | 11.66 | 22.3 | 4.08 | 4.6 | 103.25 | 111.6 | 13.16 | 16.1 | 187 | 291.1 | 374 | 582.3 | 560.4 | 839.4 | 0.73 | 0.76 | 23.89 | 46.9 | 4.33 | 4.44 |
| 28 | 98.66 | 114.66 | 15.16 | 21 | 3.66 | 4.6 | 99.75 | 258.1 | 16.16 | 20.8 | 243.67 | 497.3 | 487.33 | 994.7 | 932.1 | 1289.8 | 0.74 | 0.75 | 41.16 | 247.64 | 4.17 | 4.39 |
| LSD | 14.72 | 17.79 | 4.76 | 4.8 | 1.08 | 0.6 | 22.38 | 107.7 | 3.72 | 3.1 | 93.47 | 109.5 | 186.94 | 219 | 288.2 | 335.5 | 0.03 | 0.03 | 33.72 | 83.64 | 0.38 | 0.36 |
| Species | TFC (mg QE g−1 DW) | TPC (mg TAE g−1 DW) | ESO (%) | No. of Seeds per Plant | MAD | H2O2 | GPX | APX | Pro | Chla | Chlb | Car |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Qazvin (C) | 3.50 k | 124.32 h | 2.16 o | 48.98 g | 5.36 a | 2.40 a | 0.22 a | 0.007 d | 0.23 ab | 0.07 c | 0.031 c | 0.041 d |
| Qazvin (LS) | 4.22 g | 157.32 b | 3.11 l | 52.55 e | 0.86 d | 1.94 b | 0.067 b | 0.02 b | 0.23 a | 0.18 a | 0.06 a | 0.08 b |
| Qazvin (MS) | 4.37 f | 135.15 d | 3.53 j | 29.22 o | 1.81 d | 1.59 c | 0.075 b | 0.006 c | 0.23 a | 0.22 b | 0.063 b | 0.104 b |
| Qazvin (SS) | 3.49 k | 119.26 i | 4.26 d | 28.59p | 4.35 a | 1.95 a | 0.277 a | 0.014 a | 0.238 a | 0.136 b | 0.040 a | 0.076 b |
| Esfahfo (C) | 3.99 h | 64.85 m | 3.22 k | 63.79 c | 1.46 d | 2.20 b | 0.11 c | 0.016 b | 0.24 a | 0.17 b | 0.037 b c | 0.077 c |
| Esfahfo (LS) | 3.76 j | 77.32 l | 3.74 i | 67.34 b | 3.62 b | 2.12 a | 0.261 a | 0.01 c | 0.24 a | 0.13 b | 0.04 a | 0.07 c |
| Esfahfo (MS) | 5.35 b | 127.44 f | 2.64 n | 49.50 f | 3.10 c | 1.97 a | 0.014 c | 0.011 a | 0.24 a | 0.36 a | 0.089 a | 0.155 a |
| Esfahfo (SS) | 5.00 c | 133.58 e | 3.77 h | 38.01 l | 2.28 d | 1.52 d | 0.012 d | 0.010 b | 0.243 a | 0.179 a | 0.040 a | 0.092 a |
| Arak (C) | 4.26 g | 81.86 k | 4.77 a | 47.66 h | 2.51 b | 0.91 d | 0.18 b | 0.025 a | 0.23 b | 0.19 b | 0.065 ab | 0.099 b |
| Arak (LS) | 4.42 f | 183.83 a | 3.01 m | 40.46 ,j | 3.70 a | 0.45 d | 0.022 c | 0.025 a | 0.23 a | 0.23 a | 0.05 a | 0.10 a |
| Arak (MS) | 4.71 d | 125.99 g | 4.22 e | 39.94 k | 3.84 a | 1.54 c | 0.089 a | 0.010 b | 0.24 a | 0.14 c | 0.070 ab | 0.083 b |
| Arak (SS) | 4.40 f | 99.13 j | 4.10 f | 34.10 m | 3.30 c | 1.73 b | 0.208 b | 0.014 a | 0.240 a | 0.177 a | 0.048 a | 0.078 b |
| Yazd (C) | 3.48 k | 61.76 n | 4.61 b | 68.75 a | 1.69 c | 1.66 c | 0.17 b | 0.014 c | 0.23 b | 0.34 a | 0.080 a | 0.154 a |
| Yazd (LS) | 4.50 e | 147.00 c | 4.39 c | 59.13 d | 2.81 c | 1.41 c | 0.078 b | 0.002 d | 0.23 a | 0.05 c | 0.03 a | 0.03 d |
| Yazd (MS) | 5.94 a | 133.58 e | 4.10 f | 45.00 i | 3.13 b | 1.80 b | 0.077 a b | 0.011 a | 0.24 a | 0.11 c | 0.023 c | 0.044 c |
| Yazd (SS) | 3.92 i | 77.32 l | 3.97 g | 30.74 n | 3.50 b | 1.60 c | 0.125 c | 0.008 c | 0.240 a | 0.137 b | 0.025 b | 0.072 b |
| Population | Arak | Esfahfo | Qazvin | Yazd | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compounds (%) | *RIa | C | LS | MS | SS | C | LS | MS | SS | C | LS | MS | SS | C | LS | MS | SS |
| α-Thujene | 927 | 0.36 | 0.93 | 0.59 | 0.57 | 0.31 | 0.9 | 1.15 | 0.72 | 0.42 | 0.58 | 1.15 | 1.04 | 0.94 | 0.82 | 0.87 | 0.61 |
| α-Pinene | 941 | 0.15 | 0.27 | 0.16 | 0.17 | 0.11 | 0.23 | 0.30 | 0.22 | 0.13 | 0.16 | 0.31 | 0.27 | 0.23 | 0.21 | 0.21 | 0.17 |
| Sabinene | 972 | 0.18 | 0.38 | 0.24 | 0.22 | 0.21 | 0.35 | 0.48 | 0.31 | 0.25 | 0.21 | 0.68 | 0.42 | 0.58 | 0.40 | 0.44 | 0.24 |
| β-Pinene | 979 | 1.01 | 1.00 | 0.35 | 0.46 | 0.66 | 0.42 | 0.46 | 0.78 | 0.43 | 0.32 | 0.81 | 0.43 | 0.25 | 0.34 | 0.20 | 0.41 |
| Myrcene | 996 | 0.35 | 1.02 | 0.72 | 0.63 | 0.47 | 1.16 | 1.43 | 1.00 | 0.48 | 0.88 | 1.44 | 1.28 | 1.19 | 1.10 | 1.17 | 0.87 |
| α-Terpinene | 1017 | 0.38 | 0.75 | 0.53 | 0.66 | 0.19 | 0.74 | 0.85 | 0.67 | 0.34 | 0.67 | 0.94 | 0.84 | 0.77 | 0.74 | 0.82 | 0.62 |
| p-Cymene | 1025 | 18.74 | 22.18 | 25.42 | 22.34 | 20.32 | 26.16 | 25.71 | 19.21 | 18.80 | 25.67 | 23.09 | 25.10 | 24.06 | 21.73 | 20.55 | 22.8 |
| β-Thujone | 1110 | 0.81 | 1.11 | 0.79 | 0.95 | 0.74 | 0.18 | 0.24 | 0.34 | 0.77 | 1.04 | 1.14 | 1.13 | 1.06 | 0.88 | 0.96 | 1.00 |
| γ-Terpinene | 1057 | 26.89 | 21.94 | 26.71 | 24.27 | 23.1 | 22.75 | 25.51 | 21.71 | 22.85 | 32.81 | 23.02 | 30.83 | 31.82 | 25.55 | 22.00 | 24.33 |
| Pulegone | 1246 | 0 | 0 | 0 | 0 | 0.71 | 0.08 | 0.03 | 0.02 | 0 | 0 | 0 | 0 | 0.02 | 0.01 | 0.03 | 0.07 |
| Terpinene-4-ol | 1181 | 0.21 | 0.23 | 0.31 | 0.27 | 0.29 | 0.35 | 0.12 | 0.12 | 0.31 | 0.27 | 0.13 | 0.30 | 0.27 | 0.21 | 0.30 | 0.34 |
| Thymol | 1290 | 50.16 | 50.5 | 40.08 | 47.92 | 52.2 | 37.63 | 42.11 | 51.58 | 54.29 | 32.7 | 48.67 | 36.44 | 36.79 | 44.92 | 49.83 | 47.83 |
| Carvacrol | 1315 | 0.65 | 0.85 | 0.63 | 0.62 | 0.63 | 0.45 | 0.82 | 0.98 | 0.66 | 0.45 | 0.78 | 0.77 | 0.75 | 1.06 | 0.91 | 0.55 |
| Total | - | 99.80 | 98.75 | 98.94 | 99.08 | 99.99 | 98.19 | 92.42 | 97.66 | 99.46 | 99.36 | 98.56 | 98.85 | 98.73 | 98.15 | 98.11 | 99.84 |
| Seed Constitute | PC1 | PC2 |
|---|---|---|
| p-cymene | 0.465 | 0.091 |
| y-terpinene | 0.537 | −0.016 |
| Thymol | −0.568 | −0.087 |
| Essential oil yield | 0.204 | 0.451 |
| Total phenolic content | −0.147 | −0.004 |
| Total flavonoid content | −0.295 | 0.368 |
| DPPH radical scavenging | −0.0001 | −0.640 |
| Reducing power (500 mg/L) | −0.139 | 0.483 |
| Eigenvalue | 2.696 | 1.956 |
| Percent of variance | 33.706 | 24.455 |
| Cumulative percentage | 33.706 | 58.161 |
| Traits | ||||||||
|---|---|---|---|---|---|---|---|---|
| p-Cymene | 1 | 0.37 n.s | −0.870 n.s | −0.28 n.s | −0.643 n.s | −0869 n.s | 0.947 * | 0.007 n.s |
| γ-Terpinene | 0.796 n.s | 1 | −0.733 n.s | −0.188 n.s | 0.210 n.s | 0.114 n.s | 0.390 n.s | 0.003 n.s |
| Thymol | −0.93 n.s | −0.961 * | 1 | 0.076 n.s | 0.475 n.s | 0.580 n.s | −0.773 n.s | −0.225 n.s |
| Essential oil yield | 0.430 n.s | 0.804 n.s | −0.669 n.s | 1 | −0.429 n.s | 0.062 n.s | −0.573 n.s | 0.953 n.s |
| Total phenolic content | −0.642 n.s | −0.56 n.s | 0.614 n.s | −0.707 n.s | 1 | 0.874 n.s | −0.401 n.s | −0.597 n.s |
| Total flavonoid content | −0.524 n.s | −0.219 n.s | 0.383 n.s | 0.403 n.s | −0.289 n.s | 1 | −0.758 n.s | −0.152 n.s |
| DPPH a | −0.780 n.s | −0.990 ** | 0.919 n.s | −0.872 n.s | 0.574 n.s | 0.093 n.s | 1 | −0.309 n.s |
| Reducing power | −0.165 n.s | −0.712 n.s | 0.499 n.s | −0.914 n.s | 0.361 n.s | −0.407 n.s | 0.799 n.s | 1 |
| Traits | ||||||||
|---|---|---|---|---|---|---|---|---|
| p-Cymene | 1 | 0.944 * | −0.927 n.s | 0.969 n.s | −0.268 n.s | −0.949 n.s | −0.088 n.s | −0.270 n.s |
| γ-Terpinene | 0.931 n.s | 1 | −0.998 n.s | 0.905 n.s | 0.032 n.s | −0.888 n.s | 0.221 n.s | −0.388 n.s |
| Thymol | −0.916 n.s | −0.994 ** | 1 | −0.887 n.s | −0.079 n.s | 0.869 n.s | −0.267 n.s | 0.405 n.s |
| Essential oil yield | −0.461 n.s | −0.154 n.s | 0.179 n.s | 1 | −0.187 n.s | −0.843 n.s | −0.028 n.s | −0.463 n.s |
| Total phenolic content | −0.815 n.s | −0.939 n.s | 0.970 * | 0.138 n.s | 1 | 0.404 n.s | 0.980 n.s | −0.637 n.s |
| Total flavonoid content | −0.472 n.s | −0.359 n.s | 0.257 n.s | −0.006 n.s | 0.019 n.s | 1 | 0.219 n.s | −0.026 n.s |
| DPPH a | 0.584 n.s | 0.265 n.s | −0.211 n.s | −0.732 n.s | −0.017 n.s | −0.668 n.s | 1 | −0.663 n.s |
| Reducing power | −0.549 n.s | −0.516 n.s | 0.419 n.s | −0.142 n.s | 0.204 n.s | 0.969 * | −0.527 n.s | 1 |
| Sources of Variation | df | DPPH | RP | TFC | TPC | NCP | ESO | Thymol | γ-Terpinene | p-Cymene |
|---|---|---|---|---|---|---|---|---|---|---|
| Gen | 3 | 3,499,789 ** | 0.115 ** | 1.03 ** | 2882.8 ** | 663.74 ** | 3.27 ** | 39.73 ** | 40.08 ** | 3.05 ** |
| Salt | 3 | 27,167,123 ** | 0.323 ** | 3.51 ** | 8003.4 ** | 919.11 ** | 0.44 ** | 97.89 ** | 10.20 ** | 30.41 ** |
| Gen × Salt | 9 | 66623 ** | 0.015 ** | 0.82 ** | 2543.8 ** | 344.43 ** | 1.27 ** | 181.4 ** | 47.95 ** | 22.06 ** |
| Error | 32 | 1875.01 | 0.001 | 0.002 | 0.018 | 128.42 | 0.033 | 0.081 | 0.181 | 0.006 |
| Coeff Var | - | 1.16 | 13.37 | 1.21 | 0.11 | 24.37 | 4.91 | 0.63 | 1.68 | 0.35 |
| Sources of Variation | df | MAD | H2O2 | GPX | APX | Pro | Chla | Chlb | Car | |
| Gen | 3 | 1.24 ** | 1.73 ** | 0.008 ** | 0.0002 ** | 0.00003 ** | 0.01 ** | 0.001 ** | 0.002 ** | |
| Salt | 3 | 0.97 ** | 0.22 ** | 0.031 ** | 0.00008 ** | 0.00004 ** | 0.01 ** | 0.001 ** | 0.001 ** | |
| Gen × Salt | 9 | 5.99 ** | 0.49 ** | 0.024 ** | 0.0001 ** | 0.00001 ** | 0.028 ** | 0.001 ** | 0.005 ** | |
| Error | 32 | 0.056 | 0.029 | 0.0002 | 0.0000004 | 0.00002 | 0.0004 | 0.0003 | 0.0006 | |
| Coeff Var | - | 8.05 | 10.26 | 11.81 | 5.19 | 2.04 | 11.47 | 32.08 | 8.24 | |
| Sources of Variation | df | Value |
|---|---|---|
| Gen | ||
| Wilks’ Lambda | 51 | 0.0000006 ** |
| Pillai’s Trace | 51 | 2.9457172 ** |
| Hotelling-Lawley Trace | 51 | 1133.2316064 ** |
| Stress | ||
| Wilks’ Lambda | 51 | 0.00000650 ** |
| Pillai’s Trace | 51 | 2.88597805 ** |
| Hotelling-Lawley Trace | 51 | 380.54326555 ** |
| Accession Code | No. of Umbels | No. of Umbellules per Inflorescence | No. of Flowers per Umbels | ||||||
|---|---|---|---|---|---|---|---|---|---|
| S | OP | ID | S | OP | ID | S | OP | ID | |
| Esfahfo | 71.67 | 58.33 | 13.33 | 12.33 | 12.67 | −0.33 | 163 | 140.67 | 22.33 |
| Yazd | 69 | 86 | −17 | 18.33 | 15 | 3.33 | 222.33 | 213.67 | 8.67 |
| Qazvin | 65.33 | 40.33 | 25 | 17.67 | 13.33 | 4.33 | 229.33 | 192 | 37.33 |
| Arak | 52.67 | 153.33 | −100.67 | 16 | 16.67 | −0.67 | 242.67 | 228.67 | 14 |
| Accession Code | Height (cm) | No. of Flowering Branches | Inflorescence Diameter (cm) | ||||||
| S | OP | ID | S | OP | ID | S | OP | ID | |
| Esfahfo | 109.67 | 117.67 | −8 | 9.33 | 8 | 1.33 | 2.33 | 2.83 | −0.5 |
| Yazd | 107 | 107 | 0 | 7.33 | 6.67 | 0.67 | 2.67 | 2.67 | 0 |
| Qazvin | 122.67 | 100 | 22.67 | 12.67 | 8 | 4.67 | 3.17 | 2.5 | 0.67 |
| Arak | 104.33 | 112 | −7.67 | 10.67 | 22 | −11.33 | 2.77 | 3.83 | −1.07 |
| Accession Code | No. of Seeds per Umbel | Essential Oil Yield (%) | Crown Cover Diameter (cm2) | ||||||
| S | OP | ID | S | OP | ID | S | OP | ID | |
| Esfahfo | 326 | 281.33 | 44.67 | 1.5 | 5.32 | −3.82 | 596.6 | 1227.22 | −630.62 |
| Yazd | 444.67 | 427.33 | 17.33 | 4.54 | 5.51 | −0.96 | 558.4 | 429.66 | 128.74 |
| Qazvin | 458.67 | 384 | 74.67 | 2.07 | 4.73 | −2.65 | 1452.77 | 727.96 | 724.82 |
| Arak | 485.33 | 457.33 | 28 | 3.01 | 5.08 | −2.07 | 928.92 | 858.27 | 70.65 |
| No | Accession Number | Location | Accession Code | Geographical Region | Latitude | Longitude | Altitude (m) |
|---|---|---|---|---|---|---|---|
| 1 | 37,477 | Nahadjan, Khorasan, Iran | Khorsar | East | 32°32′ N | 59°47′ E | 1816 |
| 2 | 38,913 | Boshruieh, Khorasan, Iran | Khorbos | East | 33°53′ N | 57°27′ E | 880 |
| 3 | 38,924 | Birjand, Khorasan, Iran | Khorbir | East | 32°53′ N | 59°13′ E | 1461 |
| 4 | 38,929 | Sarbisheh, Khorasan, Iran | Khorsa | East | 32°34′ N | 59°48′ E | 1827 |
| 5 | 37,492 | Boztanj, Khorasan, Iran | Khorbi | East | 32°51′ N | 59°12′ E | 1458 |
| 6 | 37,483 | Mohammadieh, Khorasan, Iran | Khormo | East | 32°55′ N | 59°13′ E | 1460 |
| 7 | 37,529 | Ghayen, Khorasan, Iran | Khorgh | East | 33°43′ N | 59°10′ E | 1455 |
| 8 | 15,226 | Khomein, Markazi, Iran | Arakkho | West | 33°38′ N | 50°4′ E | 1811 |
| 9 | 14,743 | Arak, Markazi, Iran | Arak | West | 34°5′ N | 49°42′ E | 1735 |
| 10 | 14,492 | Shazand, Markazi, Iran | Araksha | West | 33°56′ N | 49°24′ E | 1914 |
| 11 | 14,322 | Hamedan, Hamedan, Iran | Hamdan | West | 34°47′ N | 48°30′ E | 1818 |
| 12 | 37,251 | Mollasadra, Yazd, Iran | Yazmol | Center | 31°50′ N | 54°22′ E | 1242 |
| 13 | 31,831 | Markaztahghighat, Yazd Iran | Yaztah | Center | 31°54′ N | 54°16′ E | 1213 |
| 14 | 33,683 | Saduq, Yazd, Iran | Yazsad | Center | 32°1′ N | 53°28′ E | 2091 |
| 15 | 15484 | Shahedieh, Yazd, Iran | Yazshah | Center | 31°56′ N | 54°16′ E | 1193 |
| 16 | 15,864 | Sadooqi, Yazd, Iran | Yazist | Center | 31°52′ N | 54°20′ E | 1228 |
| 17 | 1085 | Yazd, Yazd, Iran | Yazd | Center | 31°53′ N | 54°21′ E | 1215 |
| 18 | 4077 | Ghahderijan, Isfahan, Iran | Esfahgh | Center | 32°34′ N | 51°26′ E | 1615 |
| 19 | 943 | Fozveh, Isfahan, Iran | Esfahfo | Center | 32°36′ N | 51°26′ E | 1615 |
| 20 | 20,055 | Qazvin, Qazvin, Iran | Qazvin | North | 36°16′ N | 49°59′ E | 1305 |
| 21 | 906 | Tehran, Tehran, Iran | Tehran | North | 35°41′ N | 51°23′ E | 1168 |
| 22 | 10,569 | Ardabil, Ardabil, Iran | Ardebil | Northwest | 38°16′ N | 48°18′ E | 1332 |
| 23 | 17,902 | Marvdasht, Fars, Iran | Farsmar | South | 29°52′ N | 52°49′ E | 1600 |
| 24 | 17,861 | Shiraz, Fars, Iran | Farsfars | South | 29°35′ N | 52°35′ E | 1508 |
| 25 | 23,011 | Rafsanjan, Kerman, Iran | Rafsanj | South | 30°21′ N | 56°0′ E | 1545 |
| 26 | - | Genebank IPK | IPK1 | - | - | - | - |
| 27 | - | Genebank IPK | IPK2 | - | - | - | - |
| 28 | - | Genebank IPK | IPK3 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirniyam, G.; Rahimmalek, M.; Arzani, A.; Yavari, P.; Sabzalian, M.R.; Ehtemam, M.H.; Szumny, A. Phytochemical, Morphological, and Physiological Variation in Different Ajowan (Trachyspermum ammi L.) Populations as Affected by Salt Stress, Genotype × Year Interaction and Pollination System. Int. J. Mol. Sci. 2023, 24, 10438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310438
Mirniyam G, Rahimmalek M, Arzani A, Yavari P, Sabzalian MR, Ehtemam MH, Szumny A. Phytochemical, Morphological, and Physiological Variation in Different Ajowan (Trachyspermum ammi L.) Populations as Affected by Salt Stress, Genotype × Year Interaction and Pollination System. International Journal of Molecular Sciences. 2023; 24(13):10438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310438
Chicago/Turabian StyleMirniyam, Gita, Mehdi Rahimmalek, Ahmad Arzani, Parisa Yavari, Mohammad R. Sabzalian, Mohammad Hossein Ehtemam, and Antoni Szumny. 2023. "Phytochemical, Morphological, and Physiological Variation in Different Ajowan (Trachyspermum ammi L.) Populations as Affected by Salt Stress, Genotype × Year Interaction and Pollination System" International Journal of Molecular Sciences 24, no. 13: 10438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310438