

Simultaneous Application of Red and Blue Light Regulate Carbon and Nitrogen Metabolism, Induces Antioxidant Defense System and Promote Growth in Rice Seedlings under Low Light Stress
Abstract
:1. Introduction
2. Results
2.1. Morphogenesis
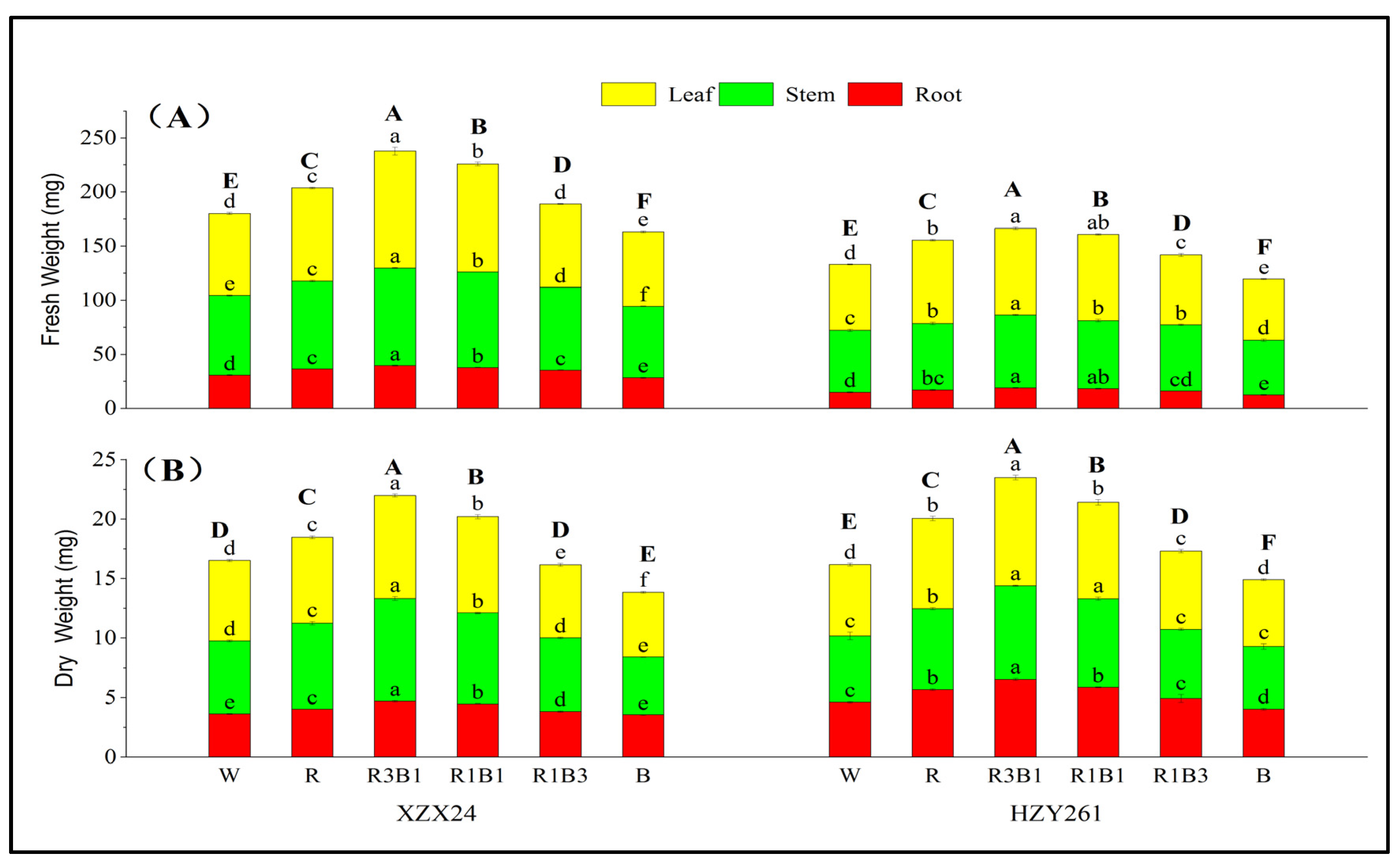
2.2. Fresh Weight, Dry Weight
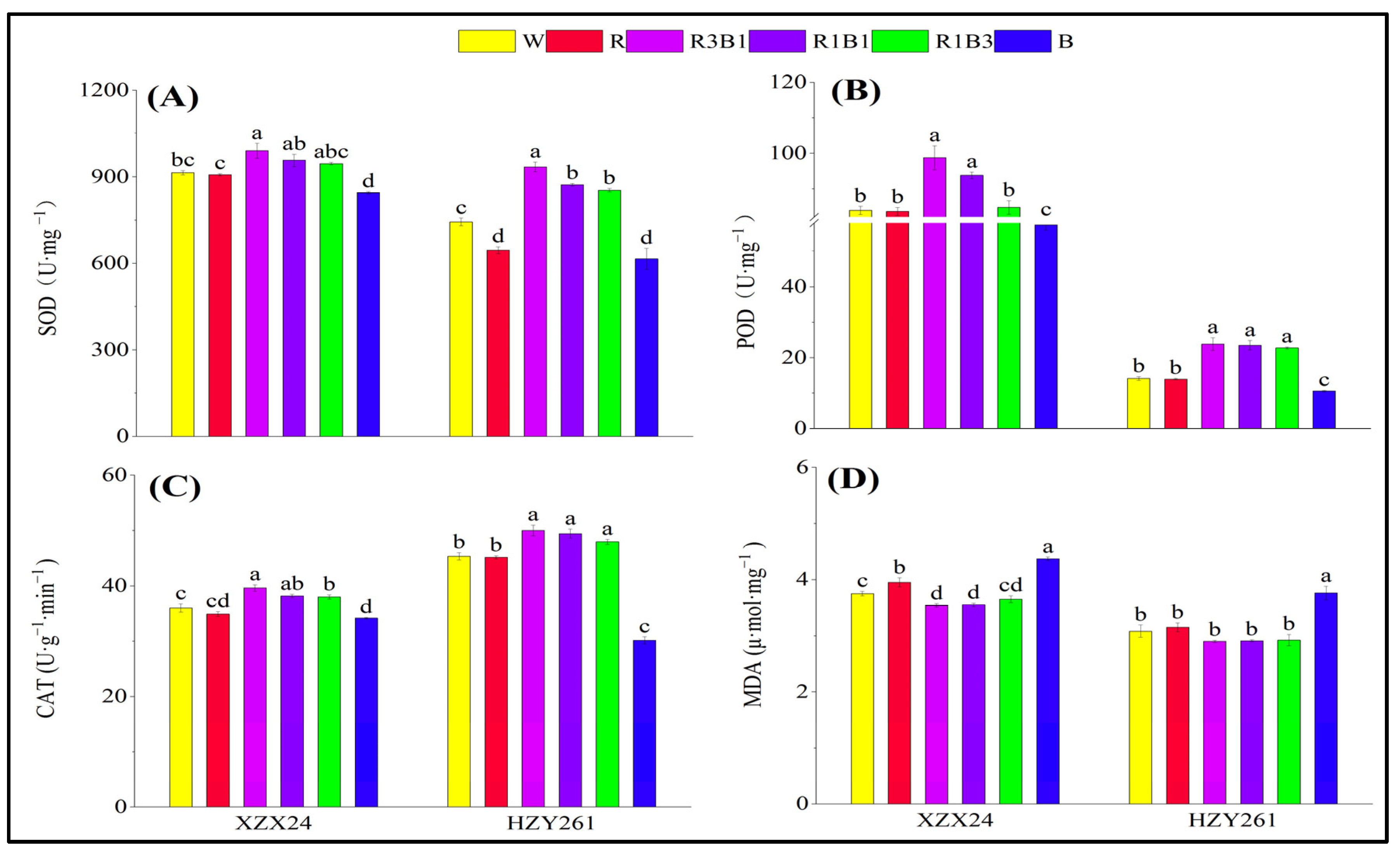
2.3. Antioxidant Enzyme Activities, MDA Content
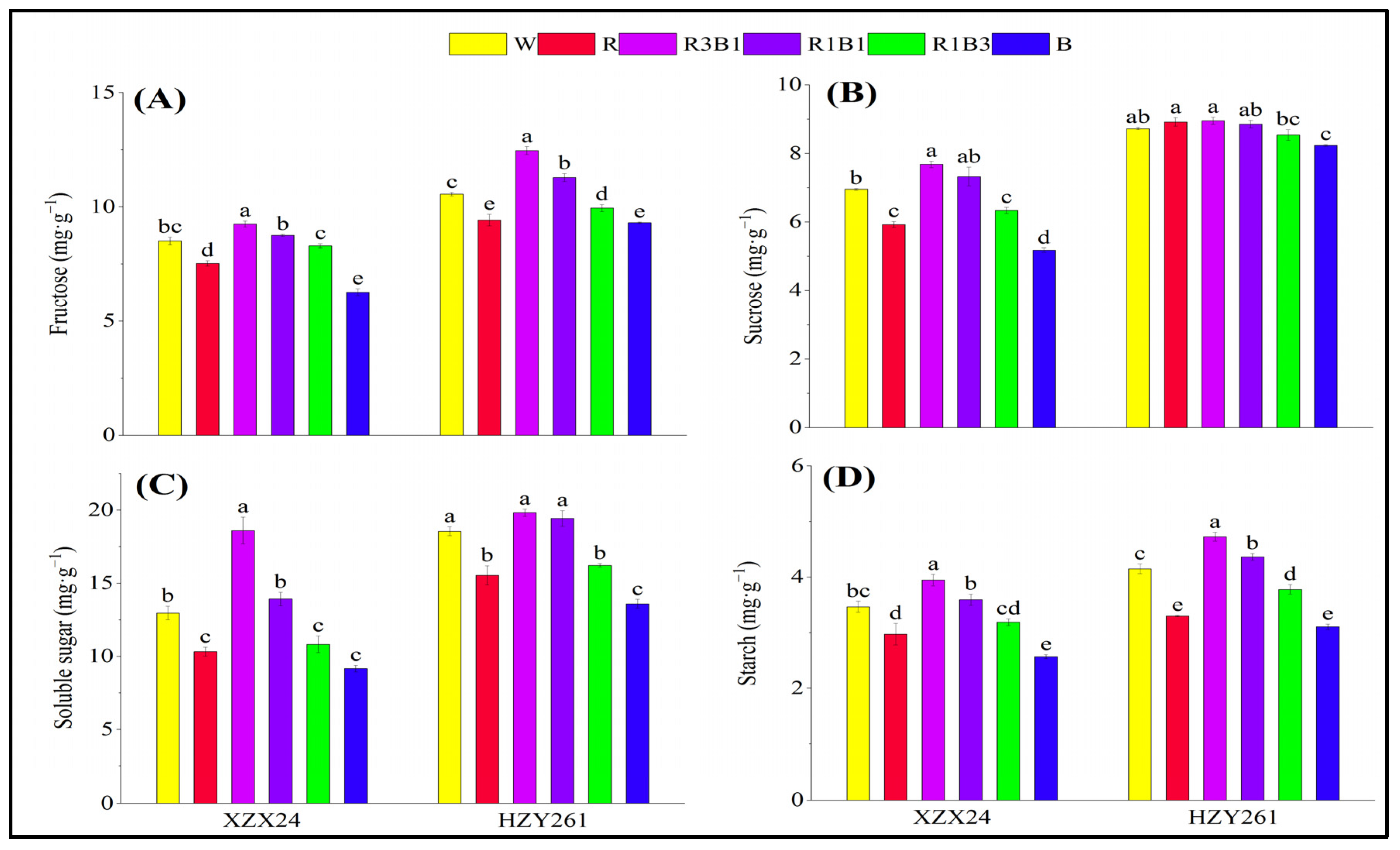
2.4. Carbon Metabolite
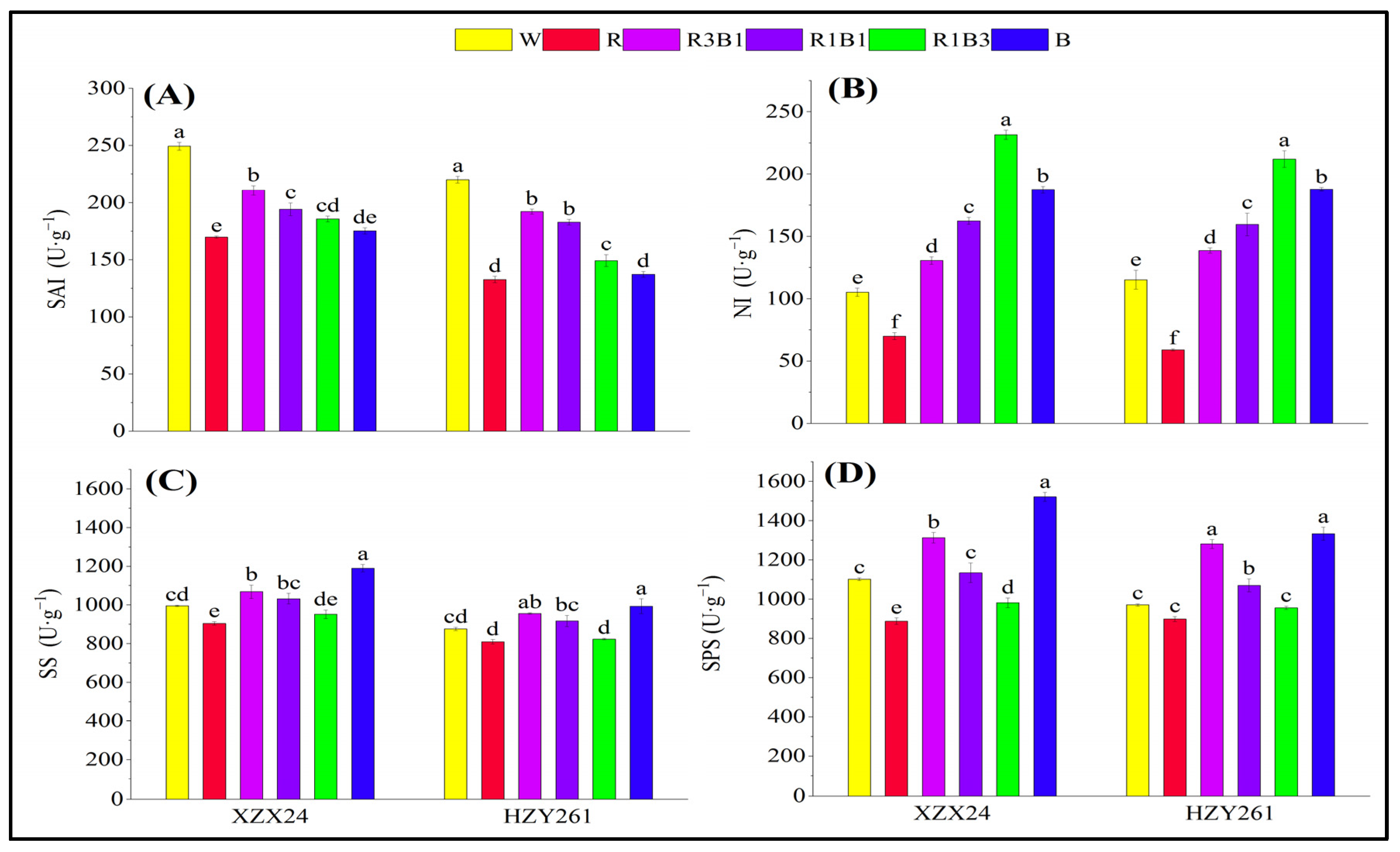
2.5. Soluble Acid Invertase (SAI), Neutral Invertase (NI), Sucrose Synthase (SS), Sucrose Phosphate Synthase (SPS)
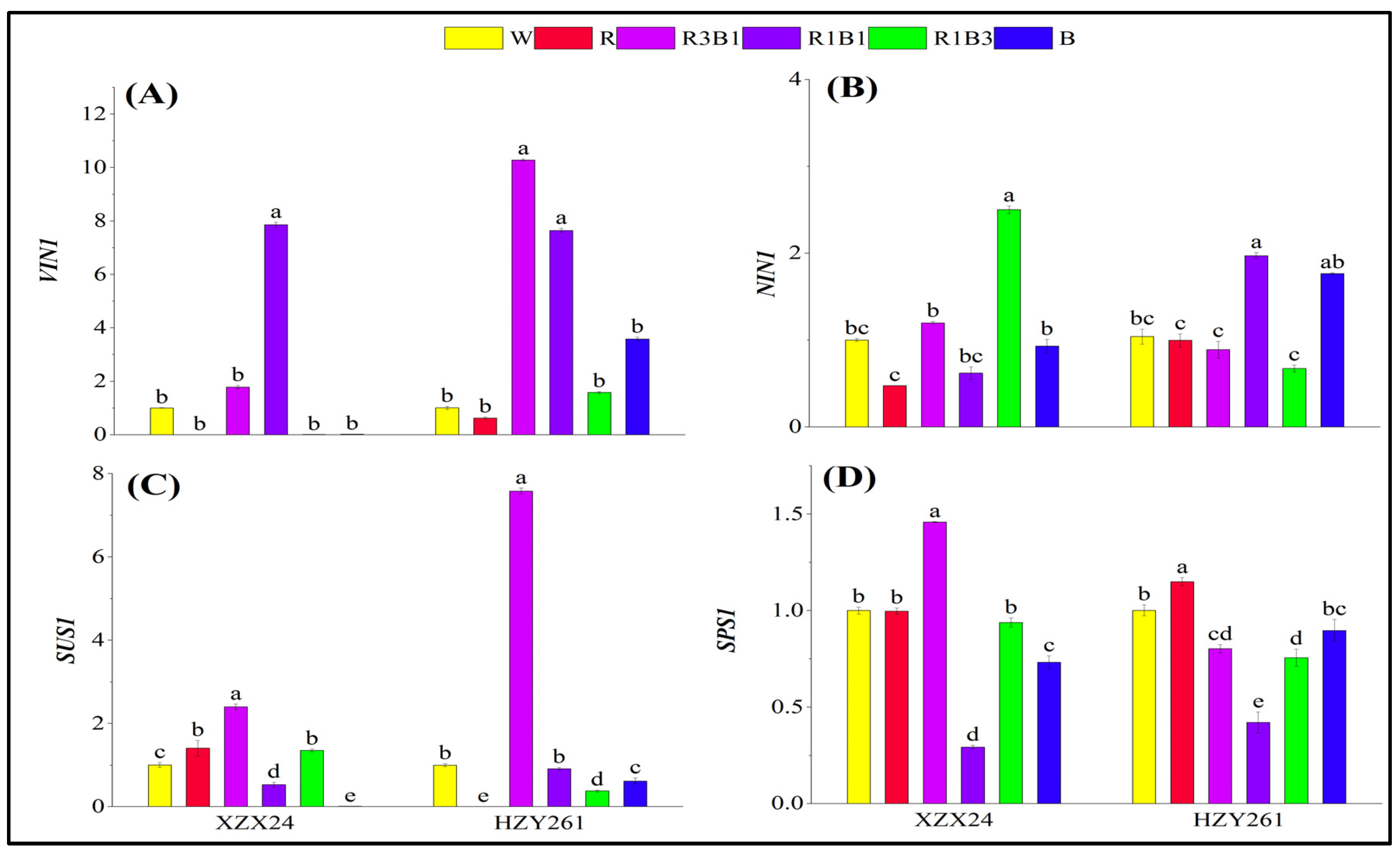
2.6. Key Gene Expression Related to Carbon Metabolism
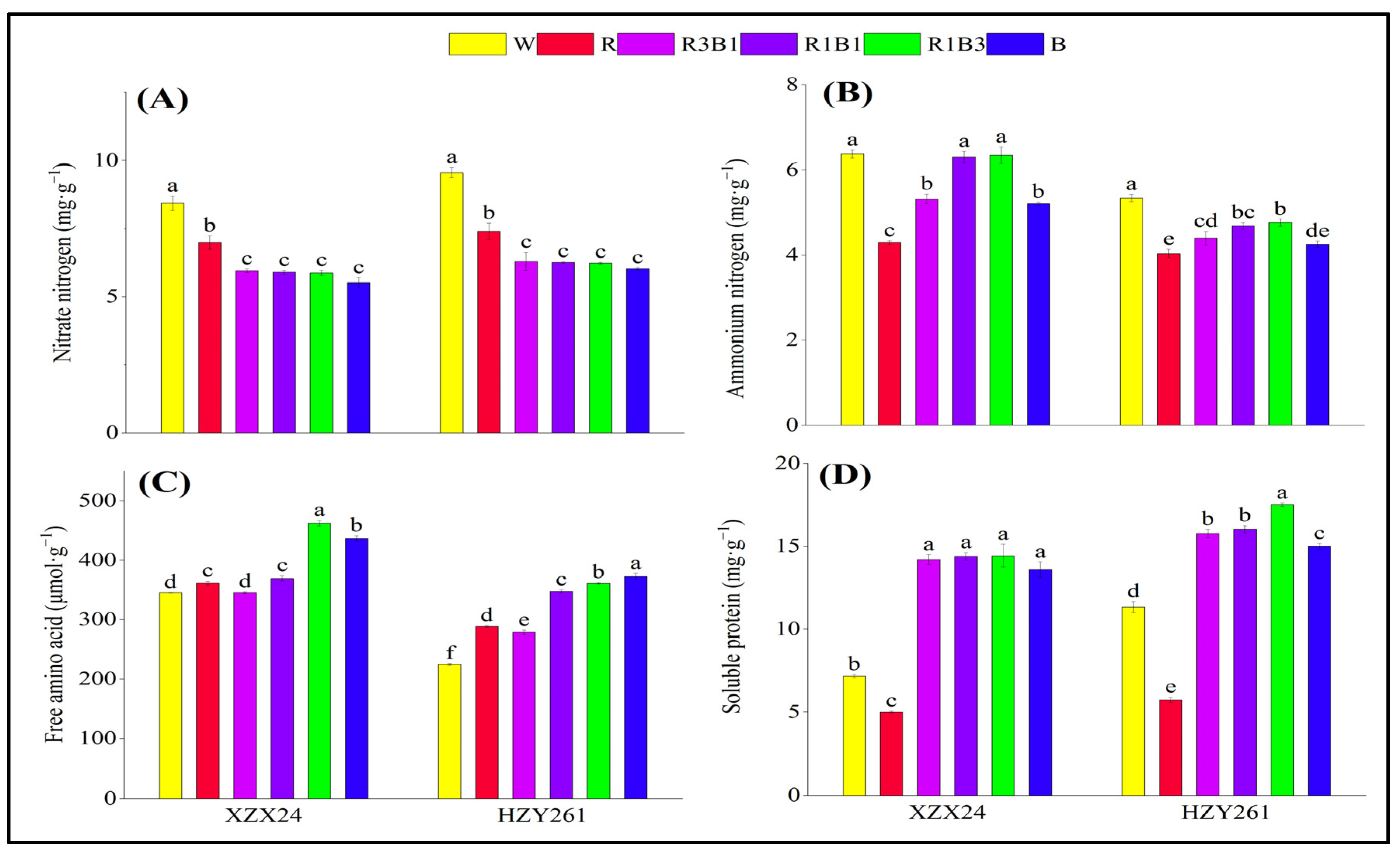
2.7. Nitrogen Metabolite
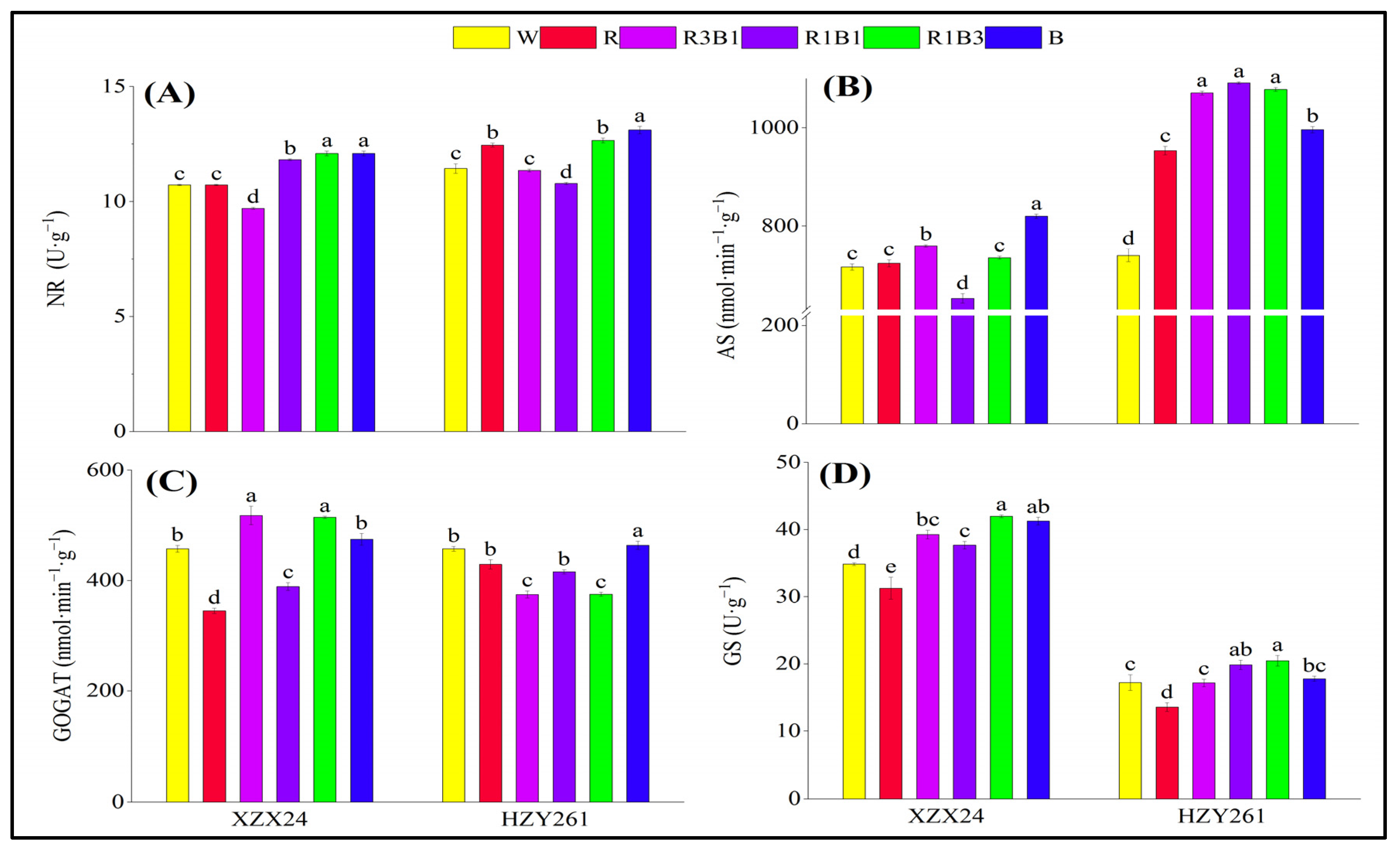
2.8. Nitrate Reductase (NR), Asparagine Synthetase (AS), Glutamate Synthetase (GOGAT), Glutamine Synthesis (GS)
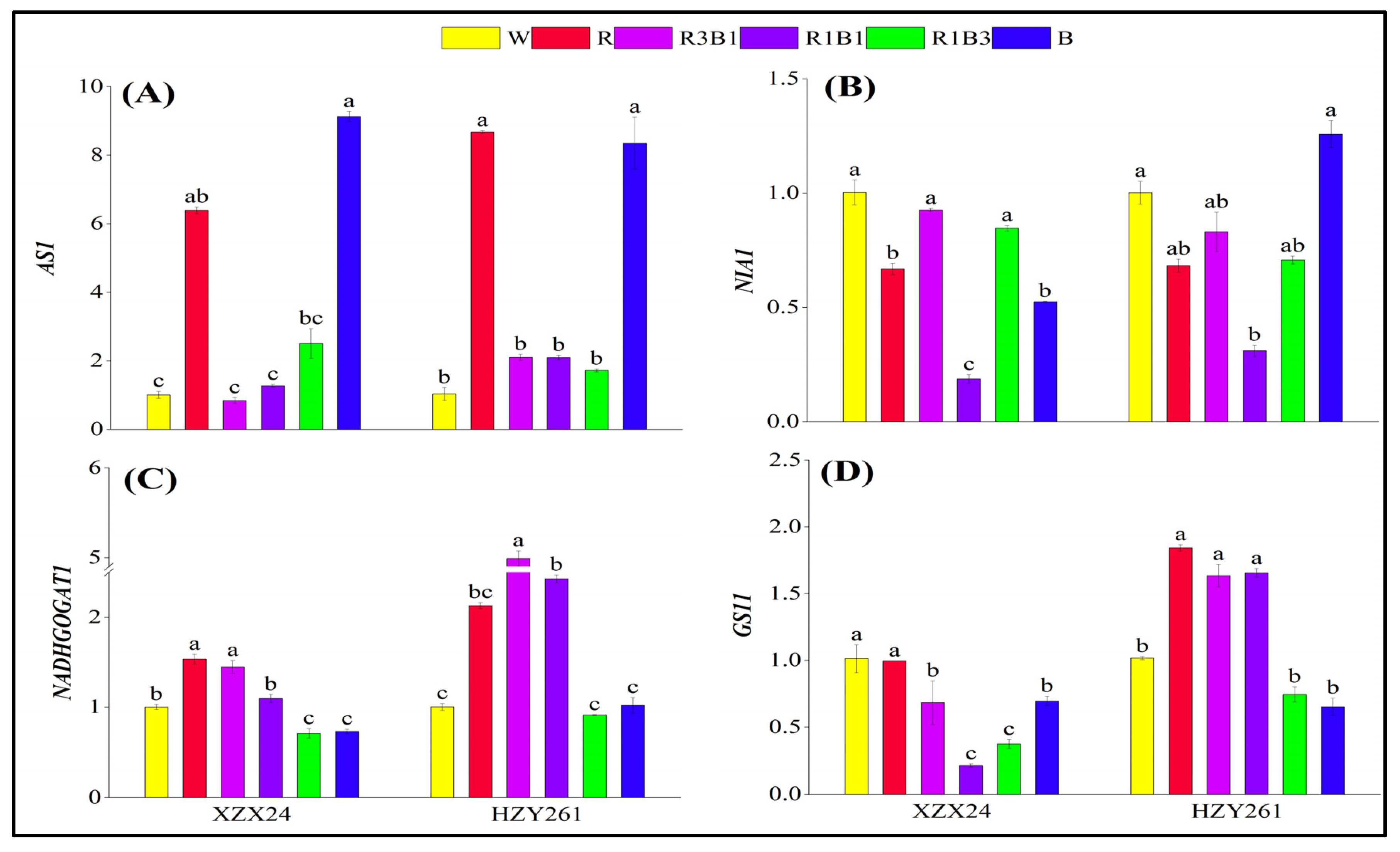
2.9. Key Gene Expression Related to Nitrogen Metabolism
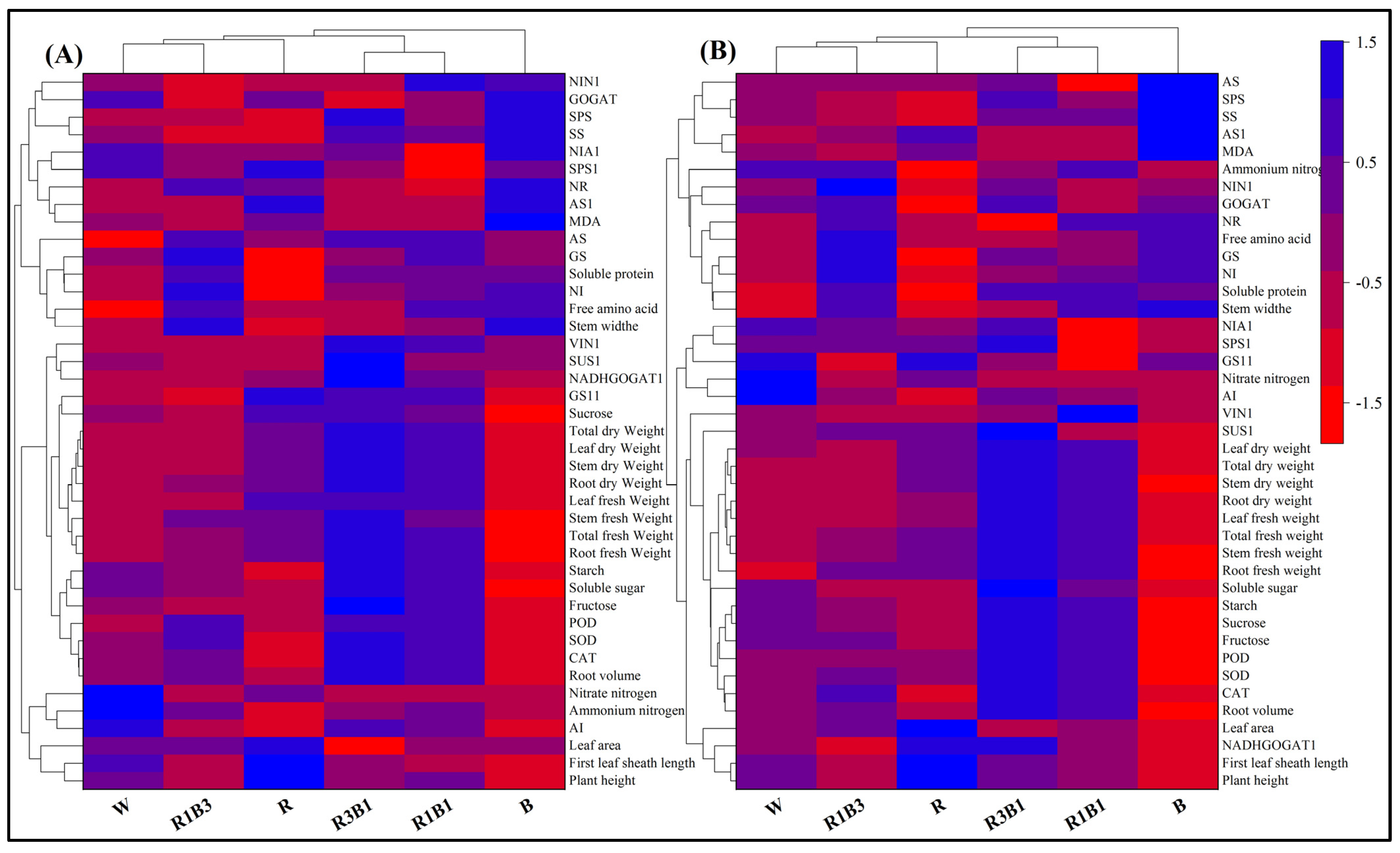
2.10. Heat Map Analysis
3. Discussion
3.1. Red and Blue Light Can Regulate the Early Morphogenesis in Rice
3.2. Red and Blue Light Can Regulate Carbon Metabolism of Rice Seedlings
3.3. Red and Blue Light Can Regulate Nitrogen Metabolism of Rice Seedlings
3.4. The Combination of Red Light and Blue Light Can Improve the Low Light Tolerance of Rice Seedlings
4. Materials and Methods
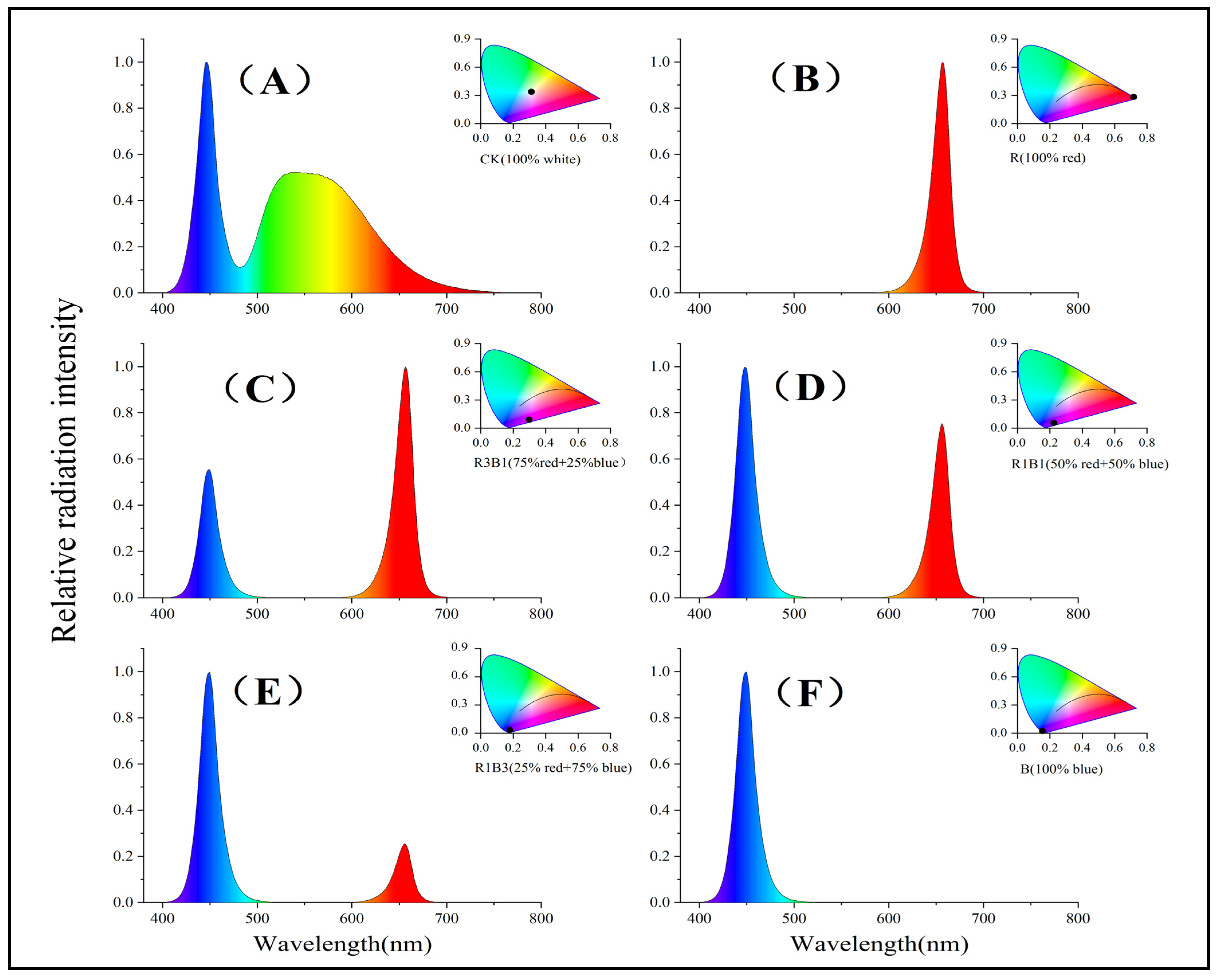
4.1. Plant Materials and Description of Experiment
4.2. Morphological Indicators
4.3. Antioxidant Enzyme Activities and MDA Content
4.4. Carbon Metabolite
4.5. Soluble Acid Invertase (SAI), Neutral Invertase (NI), Sucrose Synthase (SS), Sucrose Phosphate Synthase (SPS)
4.6. Nitrogen Metabolite
4.7. Nitrate Reductase (NR), Asparagine Synthetase (AS), Glutamate Synthetase (GOGAT), Glutamine Synthesis (GS)
4.8. Gene Expression of Carbon and Nitrogen Metabolism
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, D.; Luo, C.; Xiang, J.; Cai, S. Evaluation and Application of Reanalyzed Combined Data under Extreme Climate Conditions: A Case Study of a Typical Flood Event in the Jinsha River. Atmosphere 2022, 13, 263. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, F.; Zhou, L.; Hu, Z.; Li, Y. Regional changes of climate extremes and its effect on rice yield in Jiangsu province, southeast China. Environ. Earth Sci. 2018, 77, 106. [Google Scholar] [CrossRef]
- Wang, H.; Zhong, L.; Fu, X.; Huang, S.; Fu, H.; Shi, X.; Chen, X. Physiological and Transcriptomic Analyses Reveal the Mechanisms of Compensatory Growth Ability for Early Rice after Low Temperature and Weak Light Stress. Plants 2022, 11, 2523. [Google Scholar] [CrossRef] [PubMed]
- Budiastuti, M.T.S.; Purnomo, D.; Supriyono; Pujiasmanto, B.; Setyaningrum, D. Effect of light intensity on growth, yield and indigo content of Indigofera tinctoria L. Earth Environ. Sci. 2021, 724, 012085. [Google Scholar] [CrossRef]
- Morello, V.; Vincent, D.B.; Wu, N.; Bo-Sen, W.; MacPherson, S.; Lefsrud, M. Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa. Plants 2022, 11, 2982. [Google Scholar] [CrossRef]
- Zha, L.; Liu, W. Effects of light quality, light intensity, and photoperiod on growth and yield of cherry radish grown under red plus blue LEDs. Hortic. Environ. Biotechnol. 2018, 59, 511–518. [Google Scholar] [CrossRef]
- Kumar, A.; Panda, D.; Biswal, M.; Dey, P.; Behera, L.; Baig, M.J.; Sharma, S. Low light stress influences resistant starch content and Glycemic index of rice (O. sativa L). Starch 2019, 71, 1800216. [Google Scholar] [CrossRef]
- Yangxuan, L.; Pan, T.; Tang, Y.; Yong, Z.; Liu, Z.; Penghui, L.; Songhu, W. Proteomic analysis of rice subjected to low light stress and overexpression of OsGAPB increases the stress tolerance. Rice 2020, 13. [Google Scholar] [CrossRef]
- Li, D.; Yu, F.; Zhang, Y.; Hu, K.; Dai, D.; Song, S.; Sheng, Y. Integrative analysis of different low-light-tolerant cucumber lines in response to low-light stress. Front. Plant Sci. 2022, 13, 1093859. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Fu, X.; Gao, Z.; Liu, H.; Tan, J.; Mo, Z. Light and water treatment during the early grain filling stage regulates yield and aroma formation in aromatic rice. Sci. Rep. 2020, 10, 14830. [Google Scholar] [CrossRef]
- Liu, Q.; Bo-cong, C.; Jia-qing, M.; Gao, J.; Wu, X. Effects of Low Light on Agronomic and Physiological Characteristics of Rice Including Grain Yield and Quality. Rice Sci. 2014, 21, 243–251. [Google Scholar] [CrossRef]
- Kharshiing, E.V.; Mawphlang, O.I.L.; Lama, V.; Bhattacharjee, R.; Sahoo, L. Manipulation of light environment for optimising photoreceptor activity towards enhancing plant traits of agronomic and horticultural importance in crops. J. Hortic. Sci. Biotechnol. 2022, 97, 535–551. [Google Scholar] [CrossRef]
- Paradiso, R.; Simona, P. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Di, Q.H.; Li, J.; Yufen, D.; Wei, M.; Shi, Q.; Li, Y.; Fengjuan, Y. Combination of Red and Blue Lights Improved the Growth and Development of Eggplant (Solanum melongena L.) Seedlings by Regulating Photosynthesis. J. Plant Growth Regul. 2020, 40, 1477–1492. [Google Scholar] [CrossRef]
- Lee, H.; Gyu, H.I.; Cheong, E.J. Effect of different treatments and light quality on Ulmus pumila L. germination and seedling growth. For. Sci. Technol. 2021, 17, 162–168. [Google Scholar] [CrossRef]
- Cioć, M.; Dziurka, M.; Pawłowska, B. Changes in Endogenous Phytohormones of Gerbera jamesonii Axillary Shoots Multiplied under Different Light Emitting Diodes Light Quality. Molecules 2022, 27, 1804. [Google Scholar] [CrossRef]
- Mao, R.; Guo, S. Performance of the mixed LED light quality on the growth and energy efficiency of Arthrospira platensis. Appl. Microbiol. Biotechnol. 2018, 102, 5245–5254. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmuelling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef]
- Han, P.; Dong-xue, Z.; Rong-jun, G.; Rong-rong, Y.; Shi-gang, S.; Shi-ru, J.; Yi-kai, W. ROS Is a Factor Regulating the Increased Polysaccharide Production by Light Quality in the Edible Cyanobacterium Nostoc flagelliforme. J. Agric. Food Chem. 2019, 67, 2235–2244. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of Light Quality on Growth and Phytonutrient Accumulation of Herbs under Controlled Environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Frąszczak, B.; Kula-Maximenko, M. The Biometric Parameters of Microgreen Crops Grown under Various Light Conditions. Agriculture 2022, 12, 576. [Google Scholar] [CrossRef]
- Graham, T.; Yorio, N.; Zhang, P.; Massa, G.; Wheeler, R. Early seedling response of six candidate crop species to increasing levels of blue light. Life Sci. Space Res. 2019, 21, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Fantini, E.; Facella, P. Cryptochromes in the field: How blue light influences crop development. Physiol. Plant. 2020, 169, 336–346. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Simlat, M.; Ptak, A.; Skrzypek, E.; Moś, M.; Warchoł, M.; Patrycja, Ś. The effect of light quality on seed germination, seedling growth and selected biochemical properties of Stevia rebaudiana Bertoni. Sci. Hortic. 2016, 211, 295–304. [Google Scholar] [CrossRef]
- Jae, W.S.; Shiva, R.B.; Shin, Y.K.; Lee, J.G. The Influence of Red and Blue Light Ratios on Growth Performance, Secondary Metabolites, and Antioxidant Activities of Centella asiatica (L.) Urban. Horticulturae 2022, 8, 601. [Google Scholar] [CrossRef]
- Stefański, P.; Siedlarz, P.; Matysik, P.; Rybka, K. Usefulness of LED lightings in cereal breeding on example of wheat, barley and oat seedlings. Int. J. Agric. Biol. Eng. 2019, 12, 32–37. [Google Scholar] [CrossRef]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding Blue to Red Supplemental Light Increases Biomass and Yield of Greenhouse-Grown Tomatoes, but Only to an Optimum. Front. Plant Sci. 2018, 9, 2002. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, M.; Kim, J.A.; Mi-Jeong, J.; Lee, S.I. Blue and red light upregulate α-expansin 1 (EXPA1) in transgenic Brassica rapa and its overexpression promotes leaf and root growth in Arabidopsis. Plant Growth Regul. 2020, 91, 75–87. [Google Scholar] [CrossRef]
- Fan, C.; Manivannan, A.; Hao, W. Light Quality-Mediated Influence of Morphogenesis in Micropropagated Horticultural Crops: A Comprehensive Overview. BioMed Res. Int. 2022, 2022, 4615079. [Google Scholar] [CrossRef]
- Park, Y.G.; Jeong, B.R. Both the Quality and Positioning of the Night Interruption Light are Important for Flowering and Plant Extension Growth. J. Plant Growth Regul. 2020, 39, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Bercel, T.L.; Kranz, S.A. Effects of spectral light quality on the growth, productivity, and elemental ratios in differently pigmented marine phytoplankton species. J. Appl. Phycol. 2022, 34, 185–202. [Google Scholar] [CrossRef]
- Naif, A.E.; Yousef, A.F.; Lin, K.; Zhang, X.; Ali, M.M.; Lamlom, S.F.; Kalaji, H.M.; Kowalczyk, K.; Xu, Y. Photosynthetic performance of rocket (Eruca sativa. Mill.) grown under different regimes of light intensity, quality, and photoperiod. PLoS ONE 2021, 16, e0257745. [Google Scholar] [CrossRef]
- Riichi, O.; Ichiro, T.; Chow, W.S. The effect of different spectral light quality on the photoinhibition of Photosystem I in intact leaves. Photosynth. Res. 2021, 149, 83–92. [Google Scholar] [CrossRef]
- Kumiko, O.; Yoshifumi, U.; Makio, Y.; Jian-Ren, S.; Ryo, N.; Seiji, A. Adaptation of light-harvesting and energy-transfer processes of a diatom Phaeodactylum tricornutum to different light qualities. Photosynth. Res. 2020, 146, 227–234. [Google Scholar] [CrossRef]
- Hamdani, S.; Khan, N.; Perveen, S.; Qu, M.; Jiang, J.; Govindjee; Zhu, X.-G. Changes in the photosynthesis properties and photoprotection capacity in rice (Oryza sativa) grown under red, blue, or white light. Photosynth. Res. 2019, 139, 107–121. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, Y.; Liu, W. Light Quality Affected the Growth and Root Organic Carbon and Autotoxin Secretions of Hydroponic Lettuce. Plants 2020, 9, 1542. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, H.; Deng, J.; Zhang, Y.; Li, Y.; Huang, J.; Chen, F. Coordinating Carbon Metabolism and Cell Cycle of Chlamydomonasreinhardtii with Light Strategies under Nitrogen Recovery. Microorganisms 2021, 9, 2480. [Google Scholar] [CrossRef]
- Tarakanov, I.G.; Tovstyko, D.A.; Lomakin, M.P.; Shmakov, A.S.; Sleptsov, N.N.; Shmarev, A.N.; Ivlev, A.A. Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants. Plants 2022, 11, 441. [Google Scholar] [CrossRef]
- Zheng, L.; Ceusters, J.; Marie-Christine, V.L. Light quality affects light harvesting and carbon sequestration during the diel cycle of crassulacean acid metabolism in Phalaenopsis. Photosynth. Res. 2019, 141, 195–207. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Y.; Piao, F.; Sun, Z. Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Org. 2018, 134, 231–240. [Google Scholar] [CrossRef]
- Chen, X.-L.; Qi-chang, Y. Effects of intermittent light exposure with red and blue light emitting diodes on growth and carbohydrate accumulation of lettuce. Sci. Hortic. 2018, 234, 220–226. [Google Scholar] [CrossRef]
- Liu, J.; Liu, W. Regulation of accumulation and metabolism circadian rhythms of starch and sucrose in two leaf-color lettuces by red:blue ratios of LED continuous light. Environ. Exp. Bot. 2022, 196. [Google Scholar] [CrossRef]
- Tang, Z.; Yu, J.; Xie, J.; Lyu, J.; Feng, Z.; Dawuda, M.M.; Hu, L. Physiological and Growth Response of Pepper (Capsicum annum L.) Seedlings to Supplementary Red/Blue Light Revealed through Transcriptomic Analysis. Agronomy 2019, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zha, L.; Liu, W.; Zhou, C.; Shao, M.; Yang, Q. LED Light Quality of Continuous Light before Harvest Affects Growth and AsA Metabolism of Hydroponic Lettuce Grown under Increasing Doses of Nitrogen. Plants 2021, 10, 176. [Google Scholar] [CrossRef]
- He, J.; Gan, J.H.S.; Qin, L. Productivity, photosynthetic light-use efficiency, nitrogen metabolism and nutritional quality of C4 halophyte Portulaca oleracea L. grown indoors under different light intensities and durations. Front. Plant Sci. 2023, 14, 1106394. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Q.; Xin, Y.; Mei, Z.; Gao, A.; Liu, W.; Wang, N. Analyses of the photosynthetic characteristics, chloroplast ultrastructure, and transcriptome of apple (Malus domestica) grown under red and blue lights. BMC Plant Biol. 2021, 21, 1–14. [Google Scholar] [CrossRef]
- Zhou, X.; Huang, J.; Gan, Y.; Li, Z.; Su, L.; He, Z.; Zheng, W. Transcriptome Mechanisms of Tomato Seedlings Induced by Low-Red to Far-Red Light Ratio under Calcium Nitrate Stress. Int. J. Mol. Sci. 2023, 24, 3738. [Google Scholar] [CrossRef]
- Chen, R.; Hu, X.; Chen, D.; Song, T.; Wang, W.; Lv, M.; Hansen, N.C. Nitrogen Modulates the Effects of Short-Term Heat, Drought and Combined Stresses after Anthesis on Photosynthesis, Nitrogen Metabolism, Yield, and Water and Nitrogen Use Efficiency of Wheat. Water 2022, 14, 1407. [Google Scholar] [CrossRef]
- Ren, M.; Mao, G.; Zheng, H.; Wang, W.; Tang, Q. Growth changes of tomato seedlings responding to sodium salt of α-naphthalene acetic acid and potassium salt of fulvic acid. Sci. Rep. 2023, 13, 4024. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Konishi, N.; Ma, J.F. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytol. 2021, 232, 1778–1792. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatments | Plant Height (cm) | First Leaf Sheath Length (cm) | Stem Width (mm) | Leaf Area (mm2) | Root Volume (mm3) |
|---|---|---|---|---|---|---|
| XZX24 | W | 24.36 ± 0.38 b | 5.46 ± 0.11 b | 0.99 ± 0.02 cd | 973.09 ± 16.55 c | 204.64 ± 2.02 c |
| R | 30.98 ± 0.21 a | 6.84 ± 0.21 a | 0.95 ± 0.02 d | 1169.46 ± 6.57 a | 186.12 ± 0.83 d | |
| R3B1 | 24.26 ± 0.36 b | 5.50 ± 0.16 b | 1.04 ± 0.02 c | 925.41 ± 5.28 cd | 268.04 ± 3.04 a | |
| R1B1 | 23.46 ± 0.70 b | 5.28 ± 0.07 b | 1.22 ± 0.02 b | 946.09 ± 12.53 c | 259.41 ± 4.68 a | |
| R1B3 | 19.26 ± 0.54 c | 4.64 ± 0.04 c | 1.22 ± 0.01 b | 1036.45 ± 31.09 b | 231.24 ± 3.14 b | |
| B | 16.50 ± 0.25 d | 4.30 ± 0.08 c | 1.28 ± 0.01 a | 871.43 ± 30.7 d | 156.32 ± 2.55 e | |
| HZY261 | W | 24.18 ± 0.84 b | 5.84 ± 0.29 b | 0.86 ± 0.02 bc | 901.13 ± 6.83 ab | 146.68 ± 1.94 c |
| R | 28.16 ± 0.97 a | 6.74 ± 0.15 a | 0.82 ± 0.01 c | 937.32 ± 5.10 a | 127.52 ± 3.41 d | |
| R3B1 | 23.58 ± 0.29 b | 4.88 ± 0.15 c | 0.84 ± 0.01 c | 812.18 ± 1.77 c | 210.77 ± 3.90 a | |
| R1B1 | 22.52 ± 0.82 bc | 4.20 ± 0.30 d | 0.92 ± 0.01 b | 865.44 ± 6.77 b | 195.13 ± 3.78 b | |
| R1B3 | 21.08 ± 0.38 cd | 4.08 ± 0.12 d | 1.04 ± 0.04 a | 900.61 ± 15.23 ab | 185.32 ± 6.74 b | |
| B | 19.32 ± 0.85 d | 3.92 ± 0.14 d | 1.06 ± 0.02 a | 867.78 ± 25.80 b | 108.09 ± 0.89 e |
| Gene Name | Accession No. | Up-Primer (5′-3′) | Down-Primer (5′-3′) |
|---|---|---|---|
| OsActin3 | Os03g0718100 | CCACTATGTTCCCTGGCATT | GTACTCAGCCTTGGCAATCC |
| GS11 | Os02g0735200 | CACCAACAAGAGGCACAATG | ACTCCCACTGTCCTGGCAT |
| NADHGOGAT1 | Os01g0681900 | GTGCAGCCTGTTGCAGCATAA | CGGCATTTCACCATGCAAATC |
| SUS1 | Os03g0401300 | CATCTCAGGCTGAGACTCTGA | CAAATTCAATCGACCTTACTT |
| SPS1 | Os01g0919400 | TAGCAATGGGAAGCTGGTCT | GATCTGCTCCAGCTTGTTCC |
| AS1 | Os03g0291500 | TCGCAGGCGAAGAGGGCTCACGTCCTC | AGCGGGGAGACGATGGCGAGACGCTGC |
| NIA1 | Os08g0468100 | TTACAAGGACAACCGCGTCC | GGCGTATCCCTTCATGGTGT |
| NIN1 | Os03g0314800 | TGCCACTCAAGATATGCTACC | CCACAAGATTGCCAAAATAACC |
| VIN1 | Os04g0535600 | TGGAGCAGCAGCATACAGC | CGGATGTAAGCAGAGTTCAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, M.; Liu, S.; Mao, G.; Tang, C.; Gai, P.; Guo, X.; Zheng, H.; Wang, W.; Tang, Q. Simultaneous Application of Red and Blue Light Regulate Carbon and Nitrogen Metabolism, Induces Antioxidant Defense System and Promote Growth in Rice Seedlings under Low Light Stress. Int. J. Mol. Sci. 2023, 24, 10706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310706
Ren M, Liu S, Mao G, Tang C, Gai P, Guo X, Zheng H, Wang W, Tang Q. Simultaneous Application of Red and Blue Light Regulate Carbon and Nitrogen Metabolism, Induces Antioxidant Defense System and Promote Growth in Rice Seedlings under Low Light Stress. International Journal of Molecular Sciences. 2023; 24(13):10706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310706
Chicago/Turabian StyleRen, Maofei, Shanzhen Liu, Guiling Mao, Chengzhu Tang, Panpan Gai, Xiaoli Guo, Huabin Zheng, Weiqin Wang, and Qiyuan Tang. 2023. "Simultaneous Application of Red and Blue Light Regulate Carbon and Nitrogen Metabolism, Induces Antioxidant Defense System and Promote Growth in Rice Seedlings under Low Light Stress" International Journal of Molecular Sciences 24, no. 13: 10706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241310706