
Spinal Cord Blood Vessels in Aged Mice Show Greater Levels of Hypoxia-Induced Vascular Disruption and Microglial Activation
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
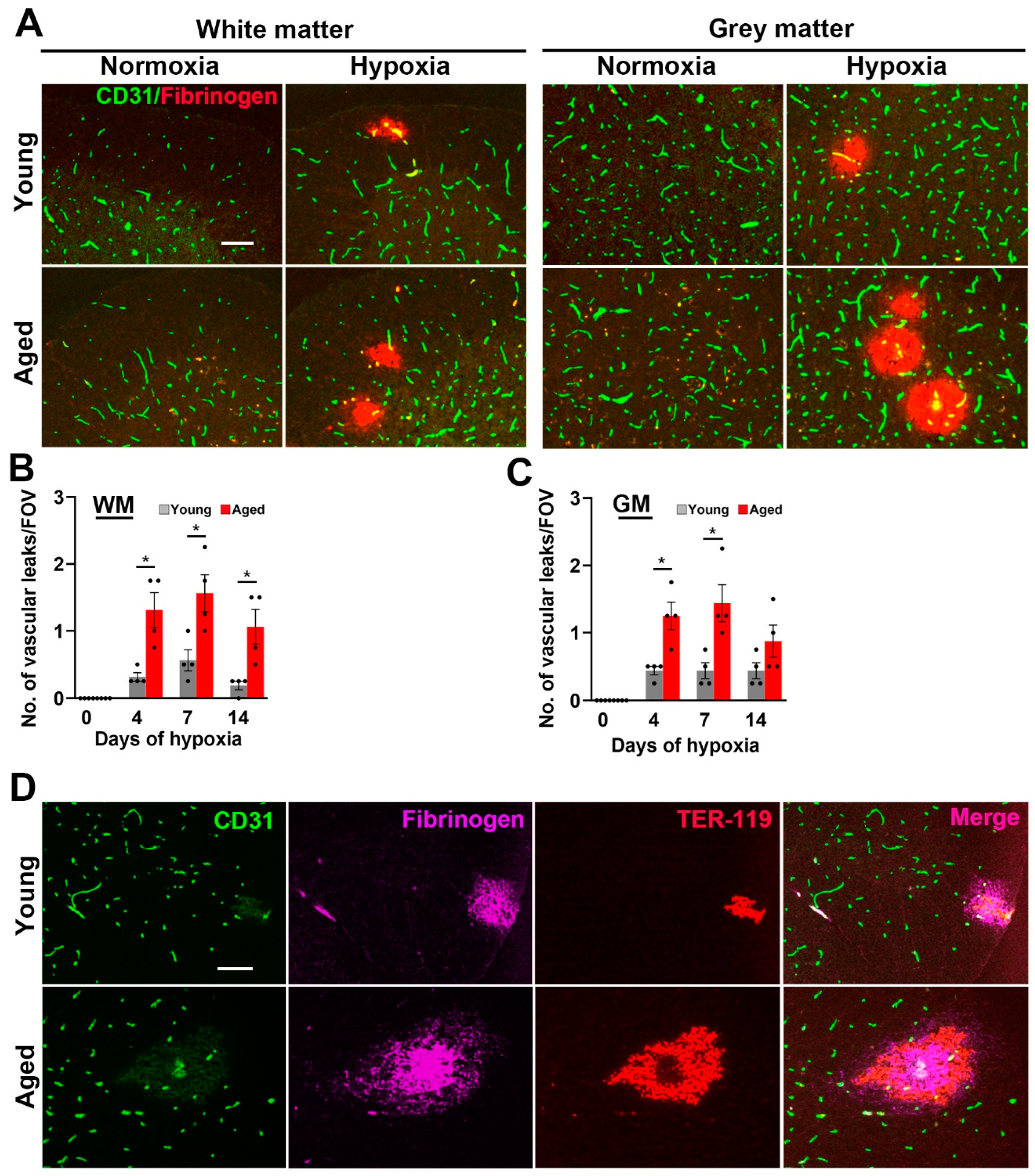
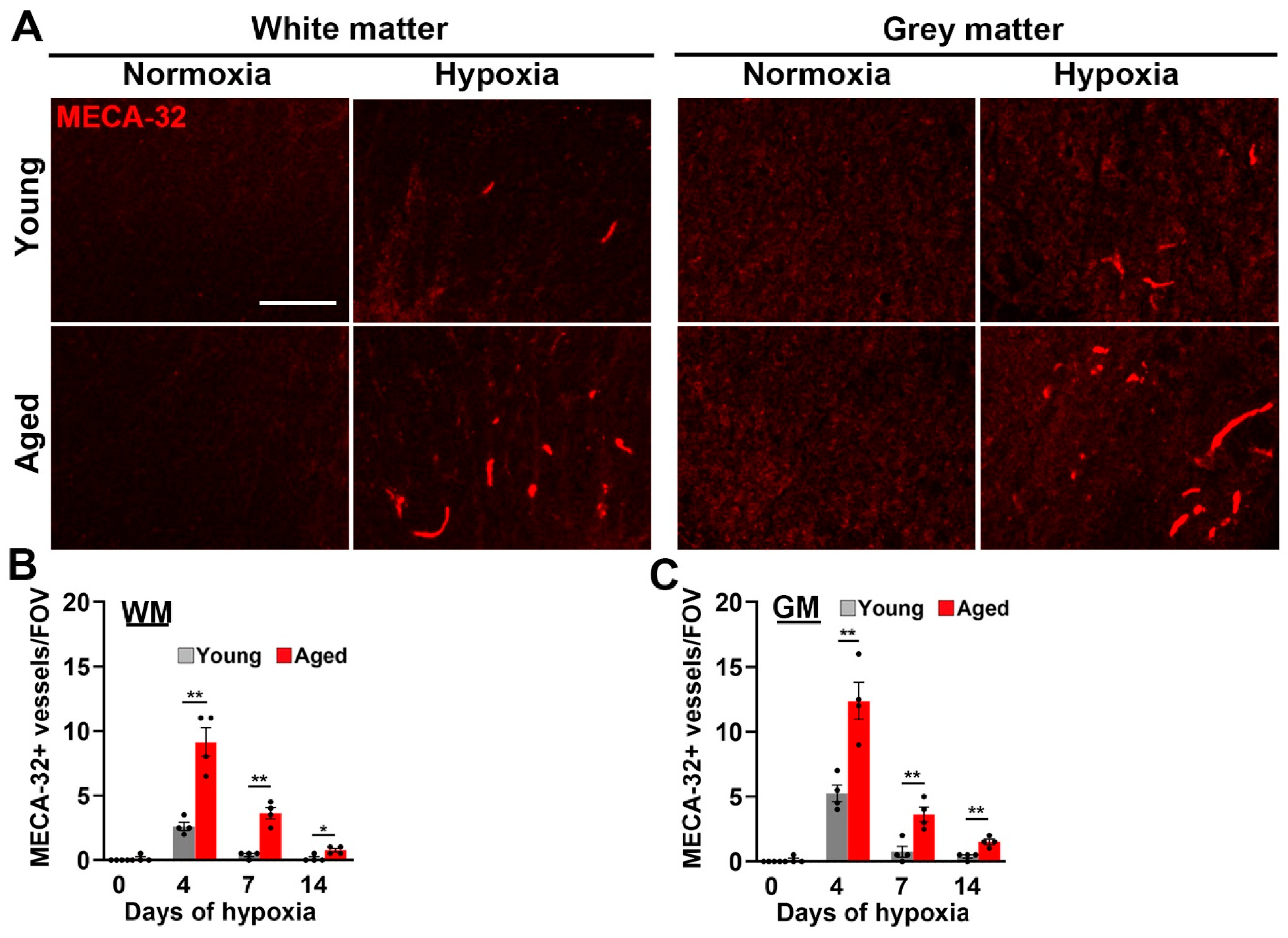
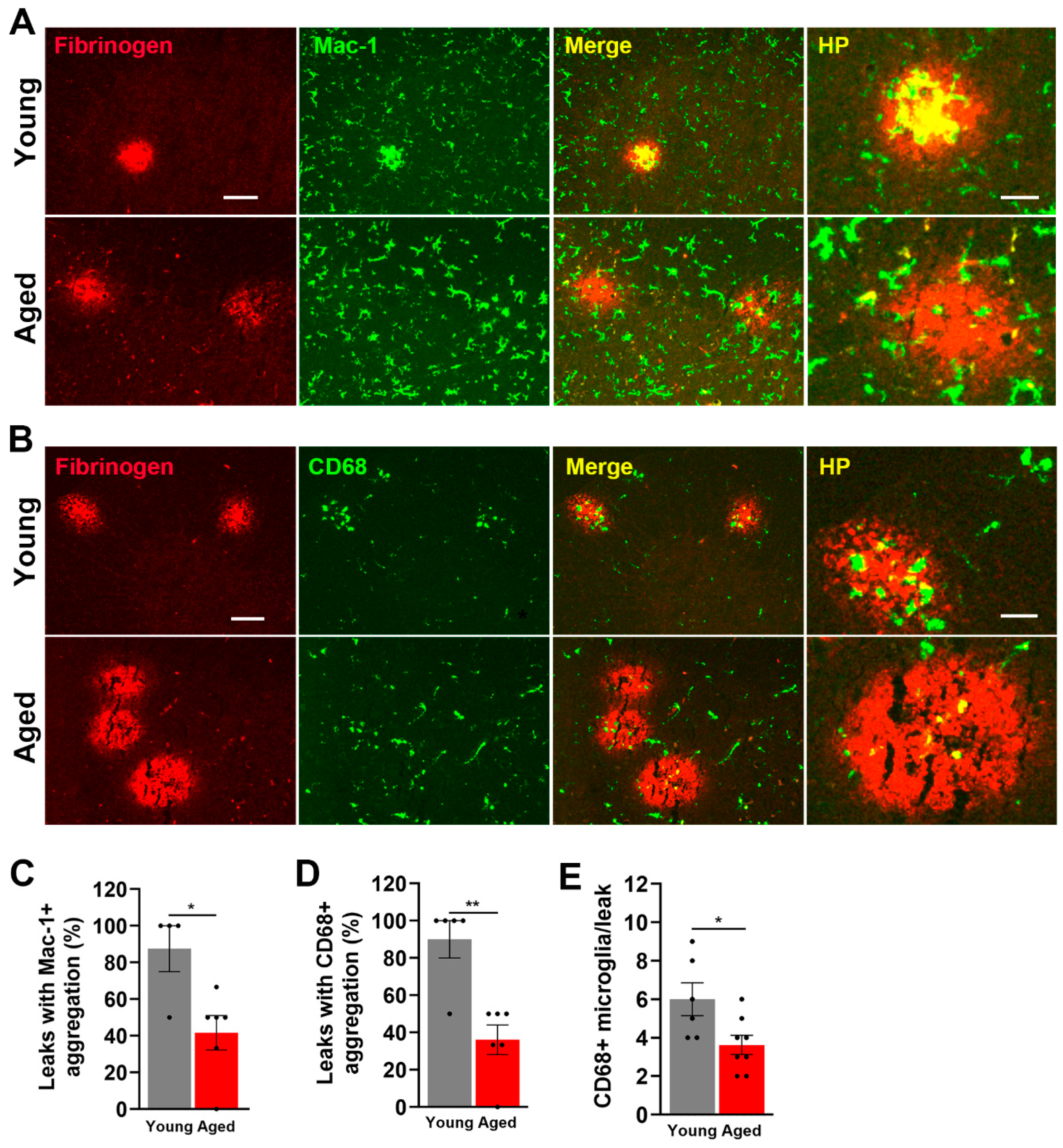
2.1. Hypoxia-Induced Vascular Disruption in the Spinal Cord Is Markedly Increased in Aged Mice
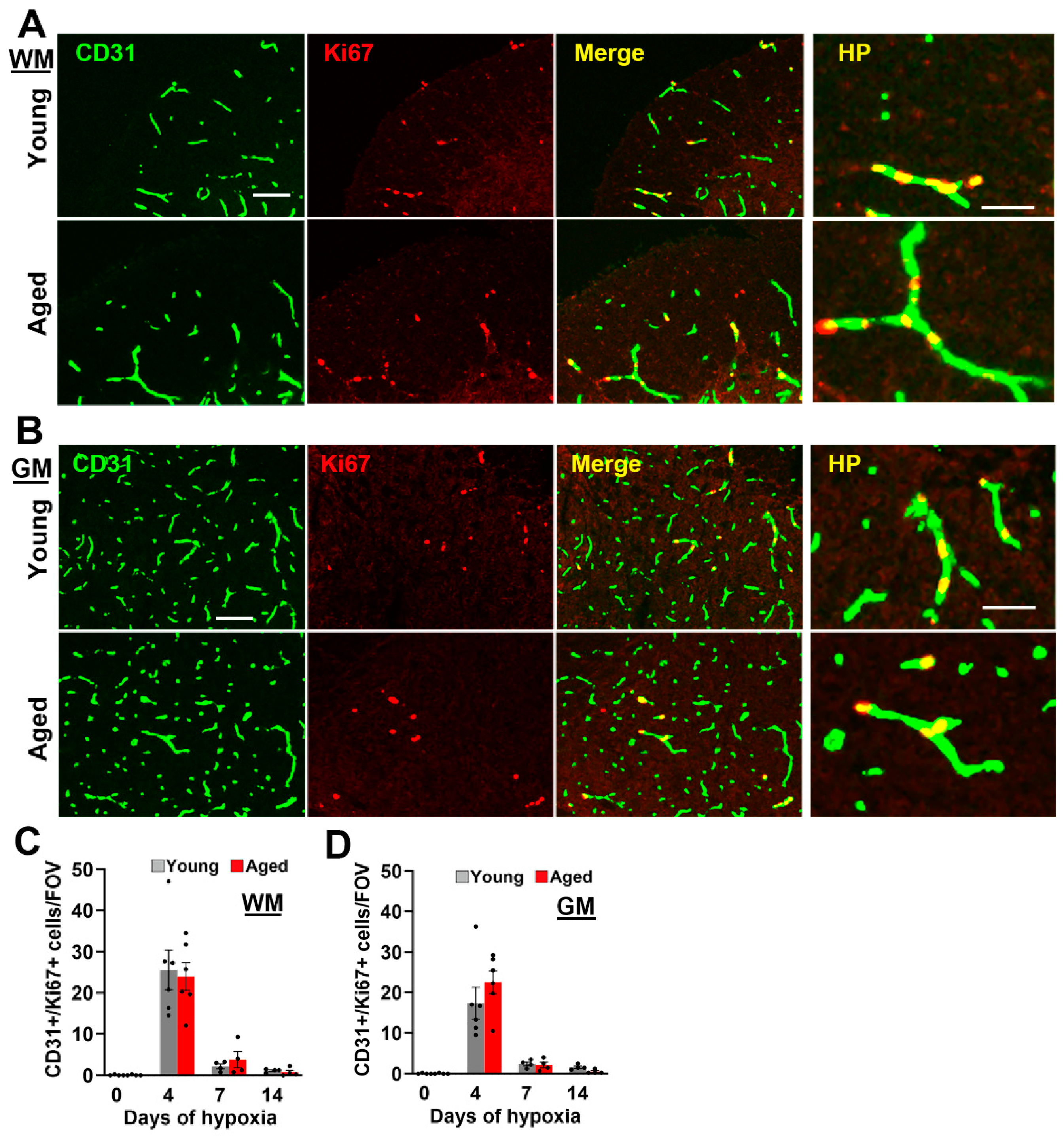
2.2. Age Has No Impact on Hypoxia-Induced Endothelial Proliferation in the Spinal Cord, but Vascular Maturation Is Delayed
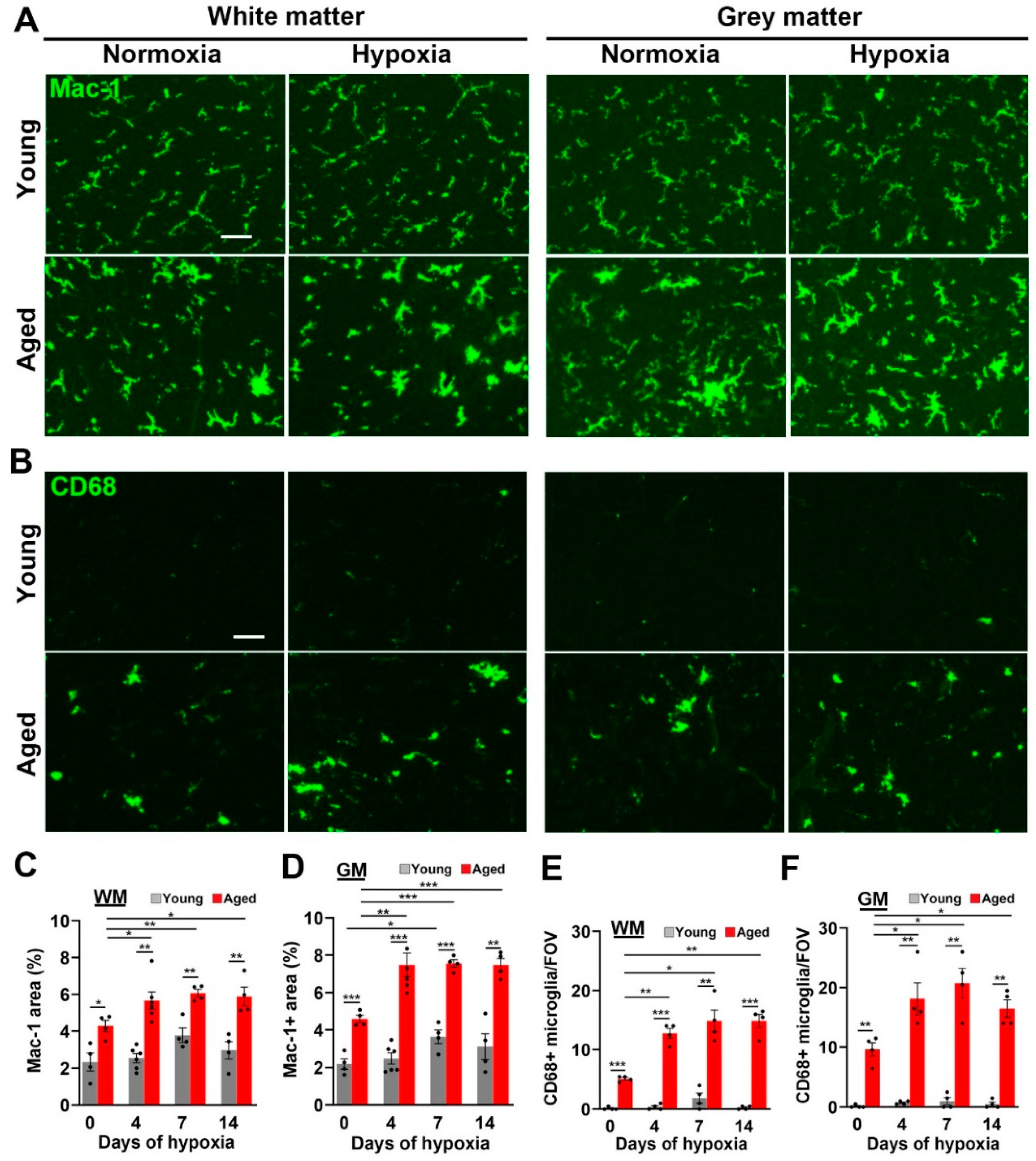
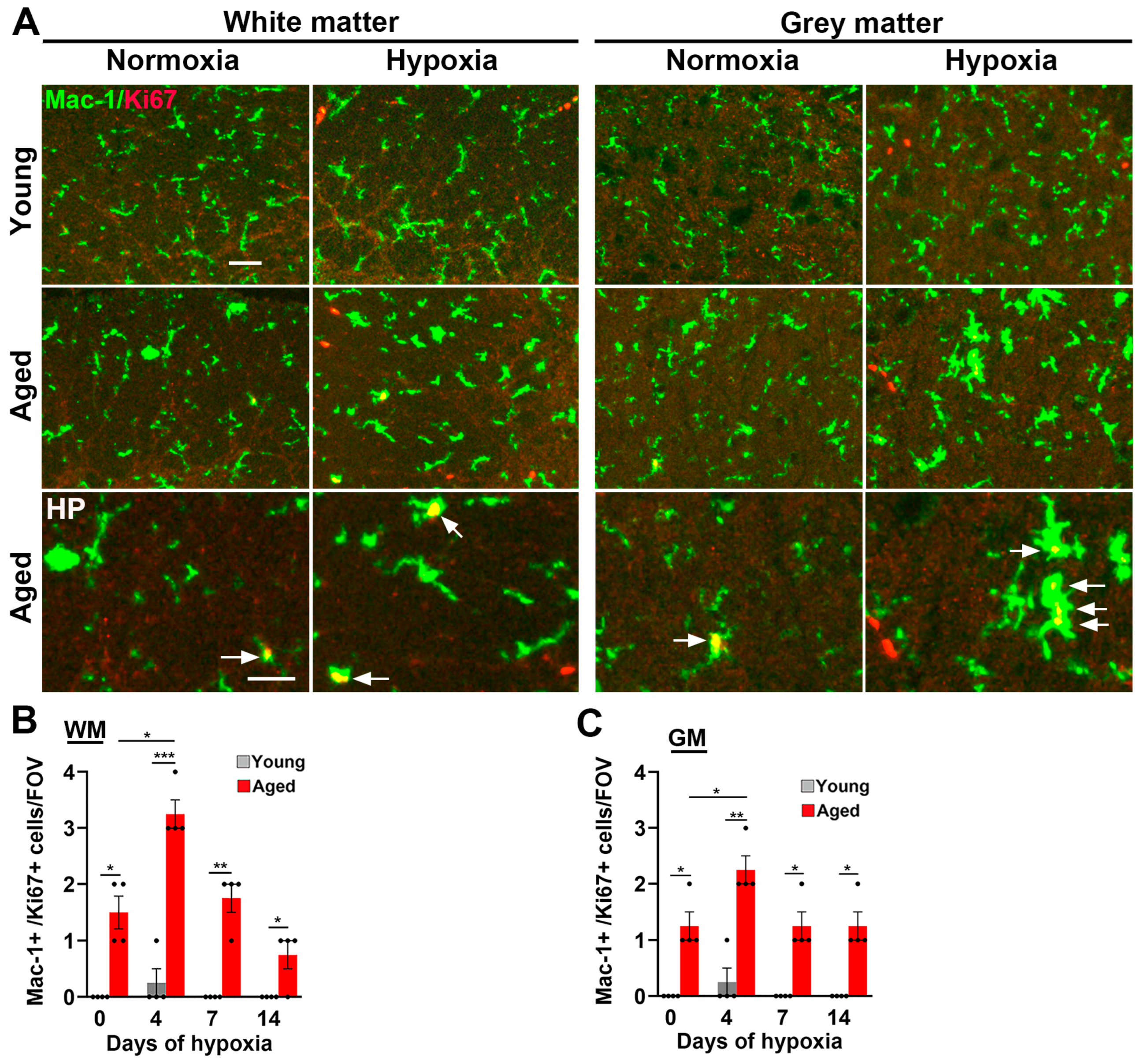
2.3. Microglial Activation in the Spinal Cord Is Greatly Increased by Age and CMH but, Paradoxically, the Microglial Vasculo-Protective Response Diminishes with Age
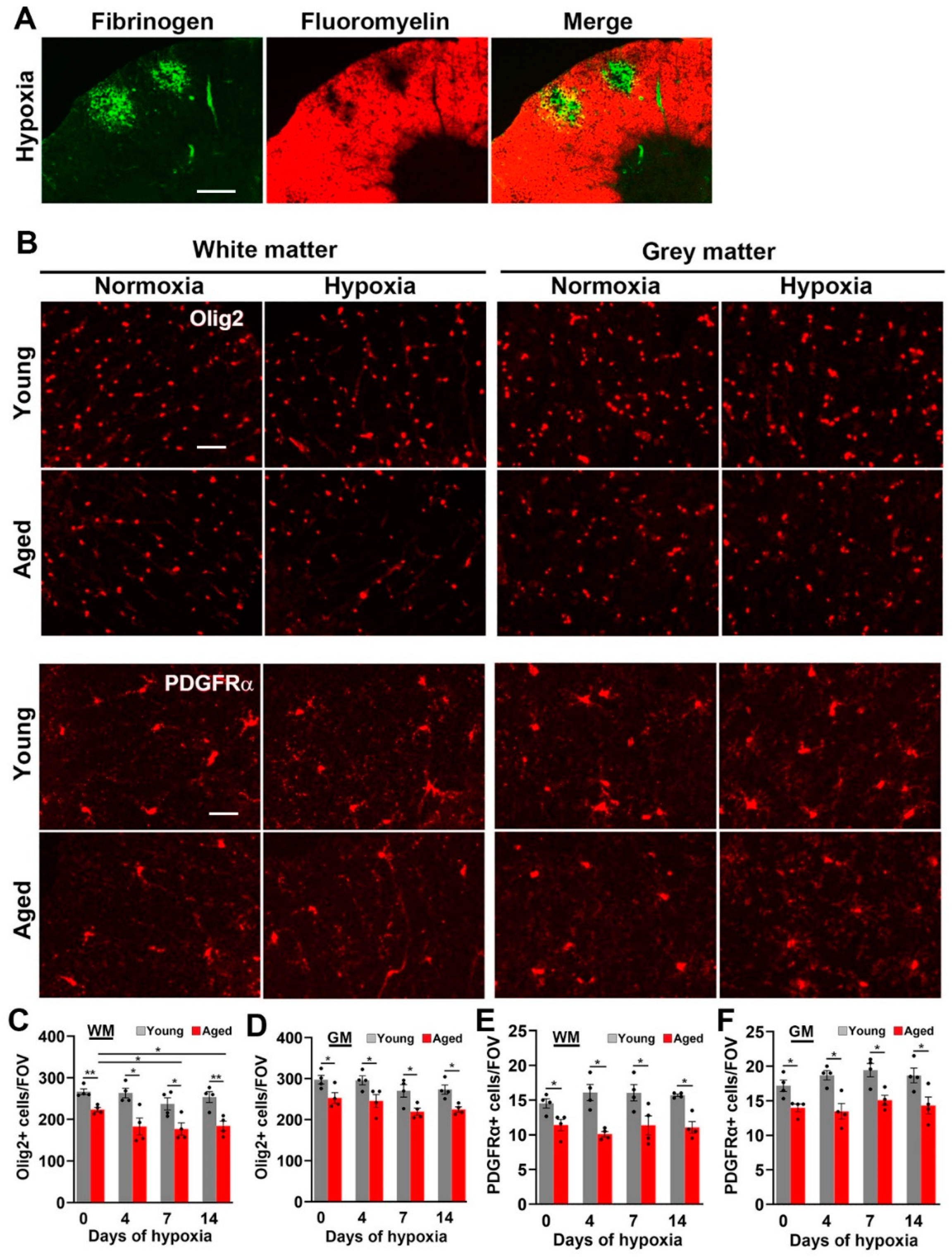
2.4. Hypoxia-Induced Vascular Disruption in the Spinal Cord Is Associated with Loss of White Matter Myelin and Oligodendrocytes
3. Discussion
3.1. The Impact of Aging on Spinal Cord Blood Vessels
3.2. Comparison of Vascular Responses in White Matter and Grey Matter
3.3. The Impact of CMH on Myelin and OLG in Young and Aged Spinal Cord
3.4. Microglia in Aged Spinal Cord Show Greatly Enhanced Activation but Reduced Vasculo-Protection
4. Materials and Methods
4.1. Animals
4.2. Chronic Hypoxia Model
4.3. Immunohistochemistry and Antibodies
4.4. Image Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ballabh, P.; Braun, A.; Nedergaard, M. The blood-brain barrier: An overview. Structure, regulation and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [PubMed]
- Huber, J.D.; Egleton, R.D.; Davis, T.P. Molecular physiology and pathophysiology of tight junctions in the blood-brain barrier. Trends Neourosci. 2001, 24, 719–725. [Google Scholar]
- LaManna, J.C.; Vendel, L.M.; Farrell, R.M. Brain adaptation to chronic hypobaric hypoxia in rats. J. Appl. Physiol. 1992, 72, 2238–2243. [Google Scholar]
- Li, L.; Welser, J.V.; Dore-Duffy, P.; Del Zoppo, G.J.; LaManna, J.C.; Milner, R. In the hypoxic central nervous system, endothelial cell proliferation is followed by astrocyte activation, proliferation, and increased expression of the α6β4 integrin and dystroglycan. Glia 2010, 58, 1157–1167. [Google Scholar] [PubMed] [Green Version]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar]
- del Zoppo, G.J.; Milner, R. Integrin-matrix interactions in the cerebral microvasculature. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1966–1975. [Google Scholar]
- Halder, S.K.; Milner, R. A critical role for microglia in maintaining vascular integrity in the hypoxic spinal cord. Proc. Natl. Acad. Sci. USA 2019, 116, 26029–26037. [Google Scholar]
- Janzer, R.C.; Raff, M.C. Astrocytes induce blood-brain barrier properties in endothelial cells. Nature 1987, 325, 253–257. [Google Scholar] [PubMed]
- Davies, D.C. Blood-brain barrier breakdown in septic encephalopathy and brain tumours. J. Anat. 2002, 200, 639–646. [Google Scholar] [PubMed]
- Gay, D.; Esiri, M. Blood-Brain Barrier Damage in Acute Multiple Sclerosis Plaques. Brain 1991, 114, 557–572. [Google Scholar]
- Oudega, M. Spinal cord injury and repair: Role of blood vessel loss and endogenous angiogenesis. Adv. Wound Care 2010, 1, 335–340. [Google Scholar]
- Waters, S.; Swanson, M.E.V.; Dieriks, B.V.; Zhang, Y.B.; Grimsey, N.L.; Murray, H.C.; Turner, C.; Waldvogel, H.J.; Faull, R.L.M.; An, J.; et al. Blood-spinal cord barrier leakage is independent of motor neuron pathology in ALS. Acta Neuropathol. Commun. 2021, 9, 144. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.K.; Kant, R.; Milner, R. Chronic mild hypoxia promotes profound vascular remodeling in spinal cord blood vessels, preferentially in white matter, via an α5β1 integrin-mediated mechanism. Angiogenesis 2018, 21, 251–266. [Google Scholar] [PubMed]
- Findley, L.J.; Wilhoit, S.C.; Suratt, P.M. Apnea duration and hypoxemia during REM sleep in patients with obstructive sleep apnea. Chest 1985, 87, 432–436. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factors: Roles in cardiovascular disease progression, prevention, and treatment. Cardiovasc. Res. 2023, 119, 371–380. [Google Scholar] [CrossRef]
- Waddell, J.A.; Emerson, P.A.; Gunstone, R.F. Hypoxia in bronchial asthma. Br. Med. J. 1967, 2, 402–404. [Google Scholar] [CrossRef] [Green Version]
- Yaffe, K.; Laffan, A.M.; Harrison, S.L.; Redline, S.; Spira, A.P.; Ensrud, K.E.; Ancoli-Israel, S.; Stone, K.L. Sleep-disordered breathing, hypoxia, and risk of mild cognitive impairment and dementia in older women. JAMA 2011, 306, 613–619. [Google Scholar]
- Yohannes, A.M.; Chen, W.; Moga, A.M.; Leroi, I.; Connolly, M.J. Cognitive Impairment in Chronic Obstructive Pulmonary Disease and Chronic Heart Failure: A Systematic Review and Meta-analysis of Observational Studies. J. Am. Med. Dir. Assoc. 2017, 18, 451. [Google Scholar]
- Hassler, O. Blood supply to human spinal cord. A microangiographic study. Arch. Neurol. 1966, 15, 302–307. [Google Scholar]
- Turnbull, I.M.; Brieg, A.; Hassler, O. Blood supply of cervical spinal cord in man. A microangiographic cadaver study. J. Neurosurg. 1966, 24, 951–965. [Google Scholar] [PubMed]
- Engelhardt, B.; Conley, F.K.; Butcher, E.C. Cell adhesion molecules on vessels during neuroinflammation in the mouse central nervous system. J. Neuroimmunol. 1994, 51, 199–208. [Google Scholar] [PubMed]
- Hallman, R.; Mayer, D.N.; Berg, E.L.; Broermann, R.; Butcher, E.C. Novel mouse endothelial cell surface marker is suppressed during differentiation of the blood brain barrier. Dev. Dyn. 1995, 202, 325–332. [Google Scholar]
- Halder, S.K.; Milner, R. Exaggerated hypoxic vascular breakdown in aged brain due to reduced microglial vasculo-protection. Aging Cell 2022, 21, e13720. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.K.; Milner, R. Mild hypoxia triggers transient blood-brain barrier disruption: A fundamental protective role for microglia. Acta Neuropathol. Commun. 2020, 8, 175. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.G.; Lue, L.F. Immune phenotypes of microglia in human neurodegenerative disease: Challenges to detecting microglial polarization in human brains. Alzheimers Res. Ther. 2015, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, F.; Burke, K.; Kantor, C.; Miller, R.H.; Yang, Y. Cyclin-dependent kinase 5 mediates adult OPC maturation and myelin repair through modulation of Akt and GsK-3β signaling. J. Neurosci. 2014, 34, 10415–10429. [Google Scholar] [CrossRef] [PubMed]
- Farrall, A.J.; Wardlaw, J.M. Blood-brain barrier: Ageing and microvascular disease—A systematic review and meta-analysis. Neurobiol. Aging 2009, 30, 337–352. [Google Scholar]
- Senatorov, V.V.J.; Friedman, A.R.; DZ, M.; Ofer, J.; Saar-Ashkenazy, R.; Charbash, A.; Jahan, N.; Chin, G.; Mihaly, E.; Lin, J.M.; et al. Blood-brain barrier dysfunction in aging induces hyperactivation of TGFβ signaling and chronic yet reversible neural dysfunction. Sci. Transl. Med. 2019, 11, 521. [Google Scholar]
- Winkler, E.A.; Sengillo, J.D.; Bell, R.D.; Wang, J.; Zlokovic, B.V. Blood-spinal cord barrier pericyte reductions contribute to increased capillary permeability. J. Cereb. Blood Flow Metab. 2012, 32, 1841–1852. [Google Scholar] [CrossRef] [Green Version]
- Doucette, J.R.; Jiao, R.; Nazarali, A.J. Age-related and cuprizone-induced changes in myelin and transcription factor gene expression and in oligodendrocyte cell densities in the rostral corpus callosum of mice. Cell. Mol. Neurobiol. 2010, 30, 607–629. [Google Scholar] [CrossRef]
- Kanaan, A.; Farahani, R.; Douglas, R.M.; Lamanna, J.C.; Haddad, G.G. Effect of chronic continuous or intermittent hypoxia and reoxygenation on cerebral capillary density and myelination. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1105–R1114. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, N.; Pham, L.D.; Hayakawa, K.; Matsuzaki, T.; Seo, J.H.; Magnain, C.; Ayata, C.; Kim, K.W.; Boas, D.; Lo, E.H.; et al. Age-related decline in oligodendrogenesis retards white matter repair in mice. Stroke 2013, 44, 2573–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, F.J.; Zhao, C.; Penderis, J.; Franklin, R.J. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J. Neurosci. 2002, 22, 2451–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagne, A.; Nikolakopoulou, A.M.; Zhao, Z.; Sagare, A.P.; Si, G.; Lazic, D.; Barnes, S.R.; Daianu, M.; Ramanathan, A.; Go, A.; et al. Pericyte degeneration causes white matter dysfunction in the mouse central nervous system. Nat. Med. 2018, 24, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Boroujerdi, A.; Welser-Alves, J.; Milner, R. Matrix metalloproteinase-9 mediates post-hypoxic vascular pruning of cerebral blood vessels by degrading laminin and claudin-5. Angiogenesis 2015, 18, 255–264. [Google Scholar] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halder, S.K.; Milner, R. Spinal Cord Blood Vessels in Aged Mice Show Greater Levels of Hypoxia-Induced Vascular Disruption and Microglial Activation. Int. J. Mol. Sci. 2023, 24, 11235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241411235
Halder SK, Milner R. Spinal Cord Blood Vessels in Aged Mice Show Greater Levels of Hypoxia-Induced Vascular Disruption and Microglial Activation. International Journal of Molecular Sciences. 2023; 24(14):11235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241411235
Chicago/Turabian StyleHalder, Sebok K., and Richard Milner. 2023. "Spinal Cord Blood Vessels in Aged Mice Show Greater Levels of Hypoxia-Induced Vascular Disruption and Microglial Activation" International Journal of Molecular Sciences 24, no. 14: 11235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241411235