
Structural and Genetic Identification of the O-Antigen from an Escherichia coli Isolate, SD2019180, Representing a Novel Serogroup

Abstract
:1. Introduction
2. Results and Discussion
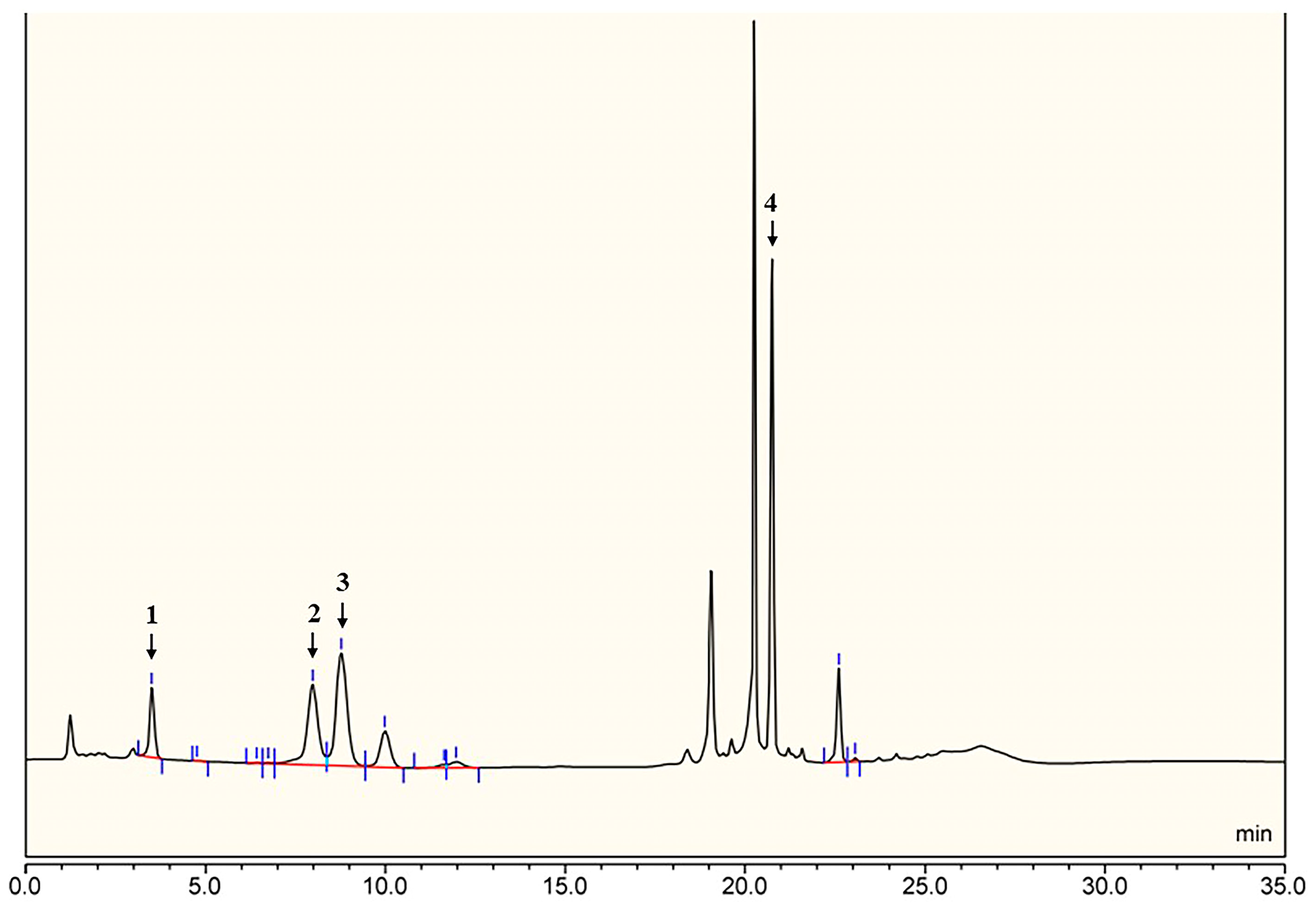
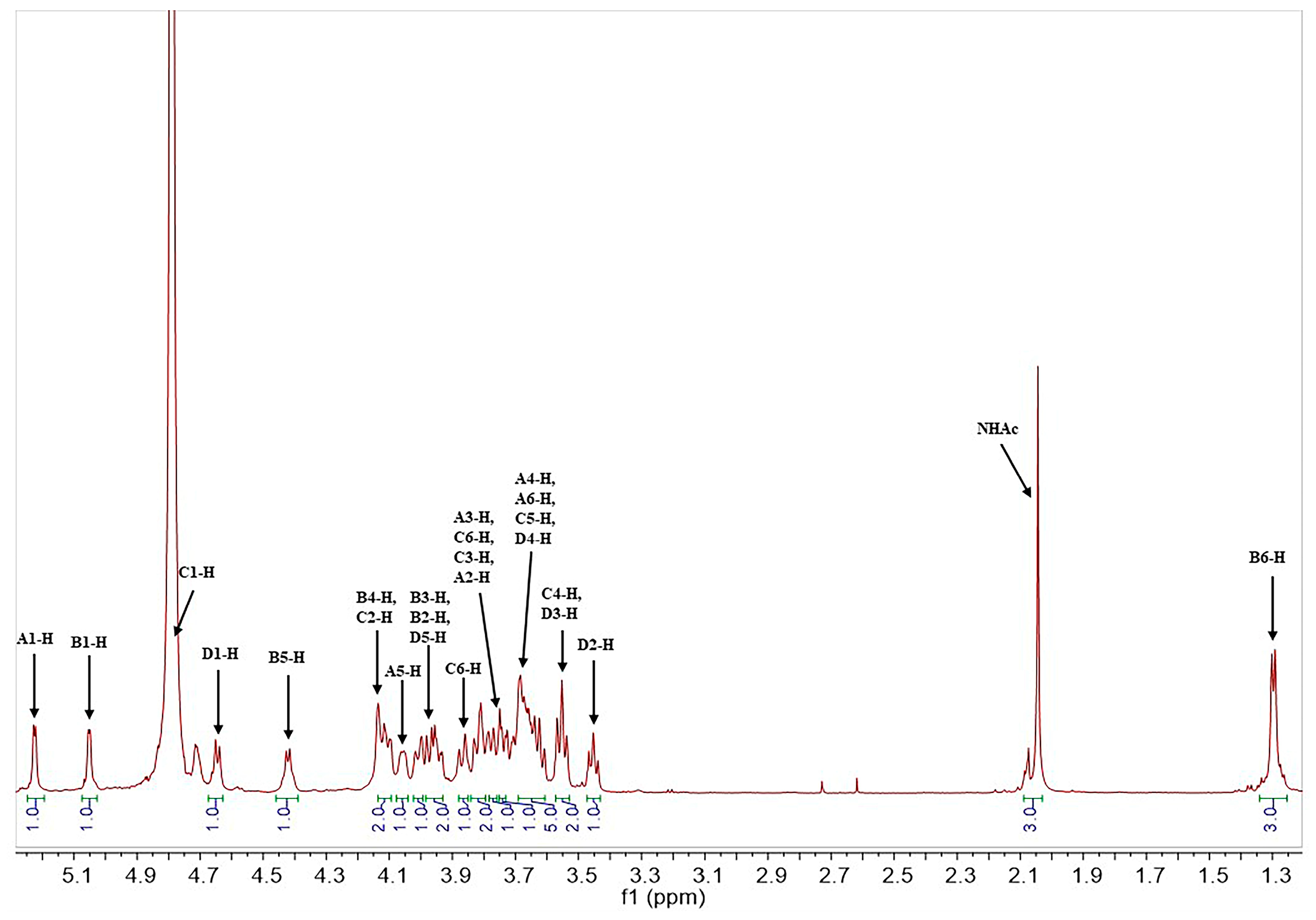
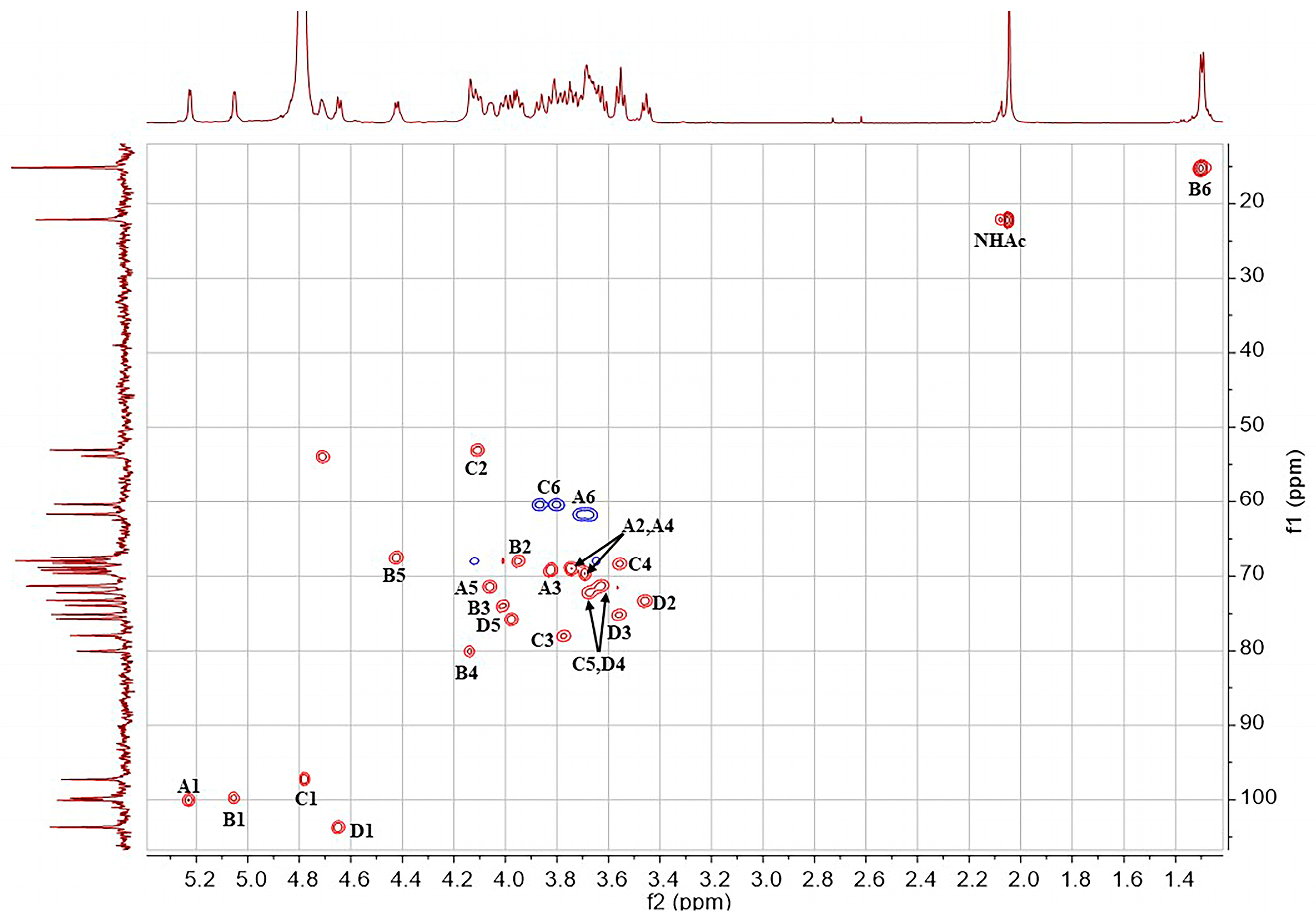
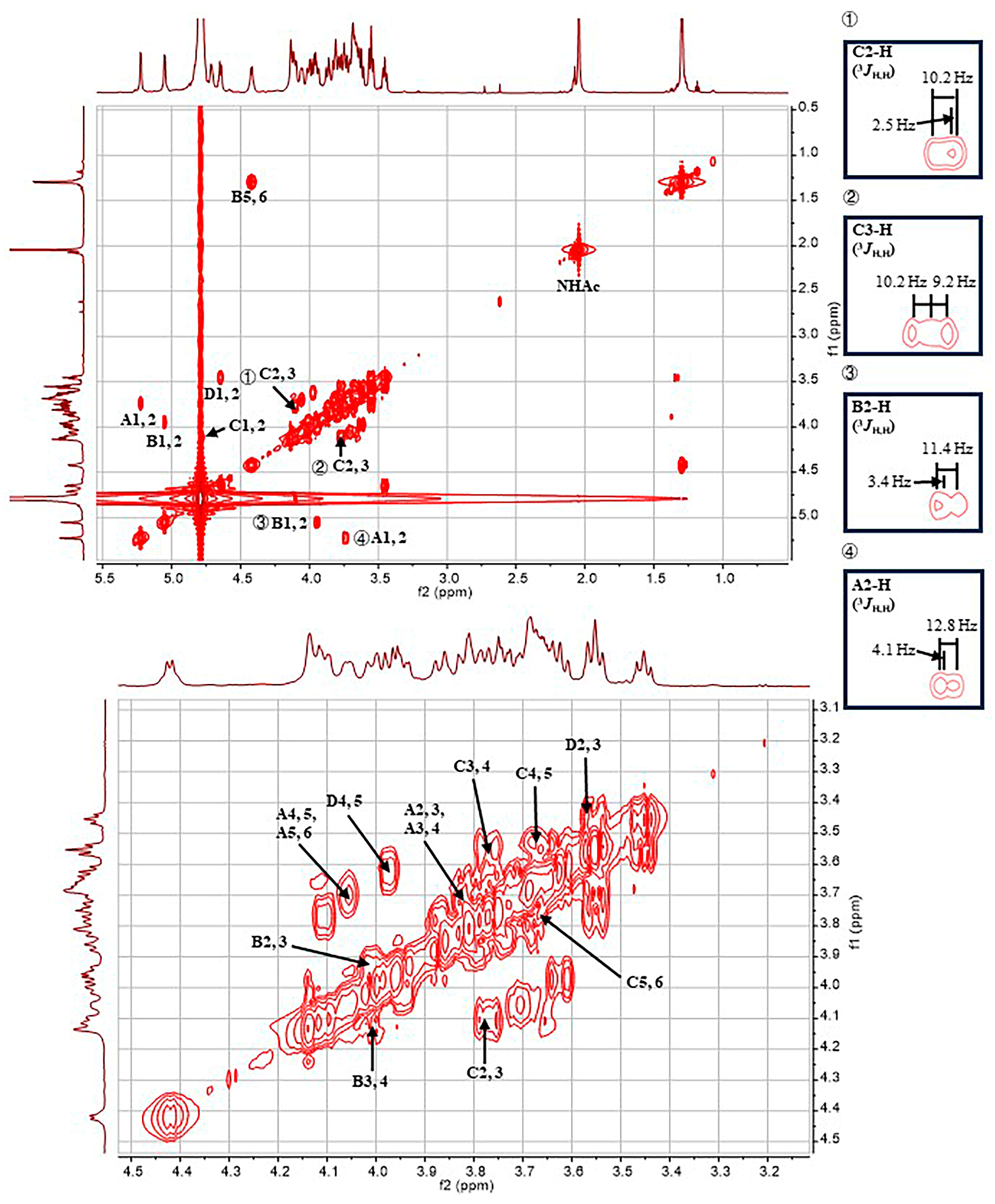
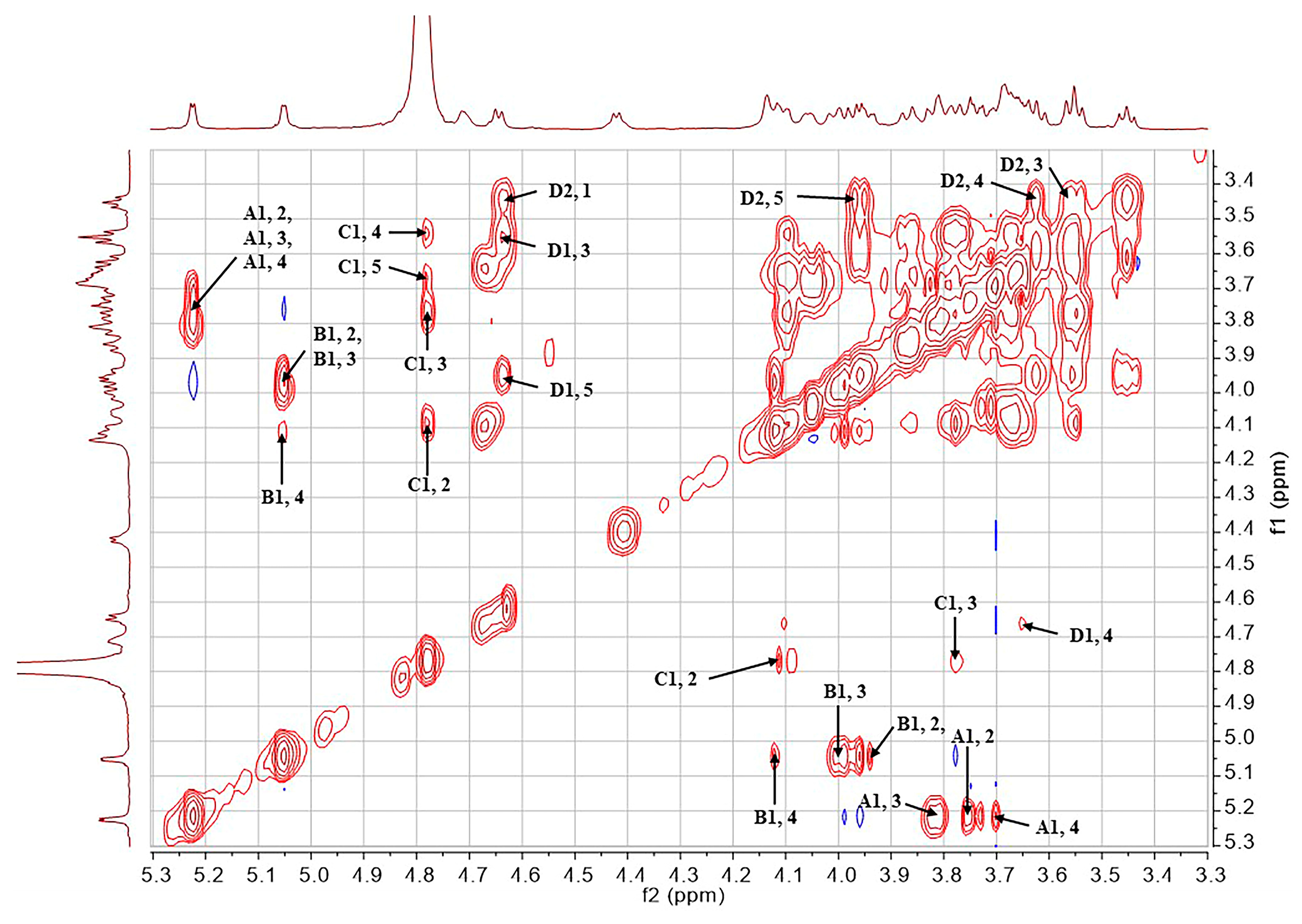
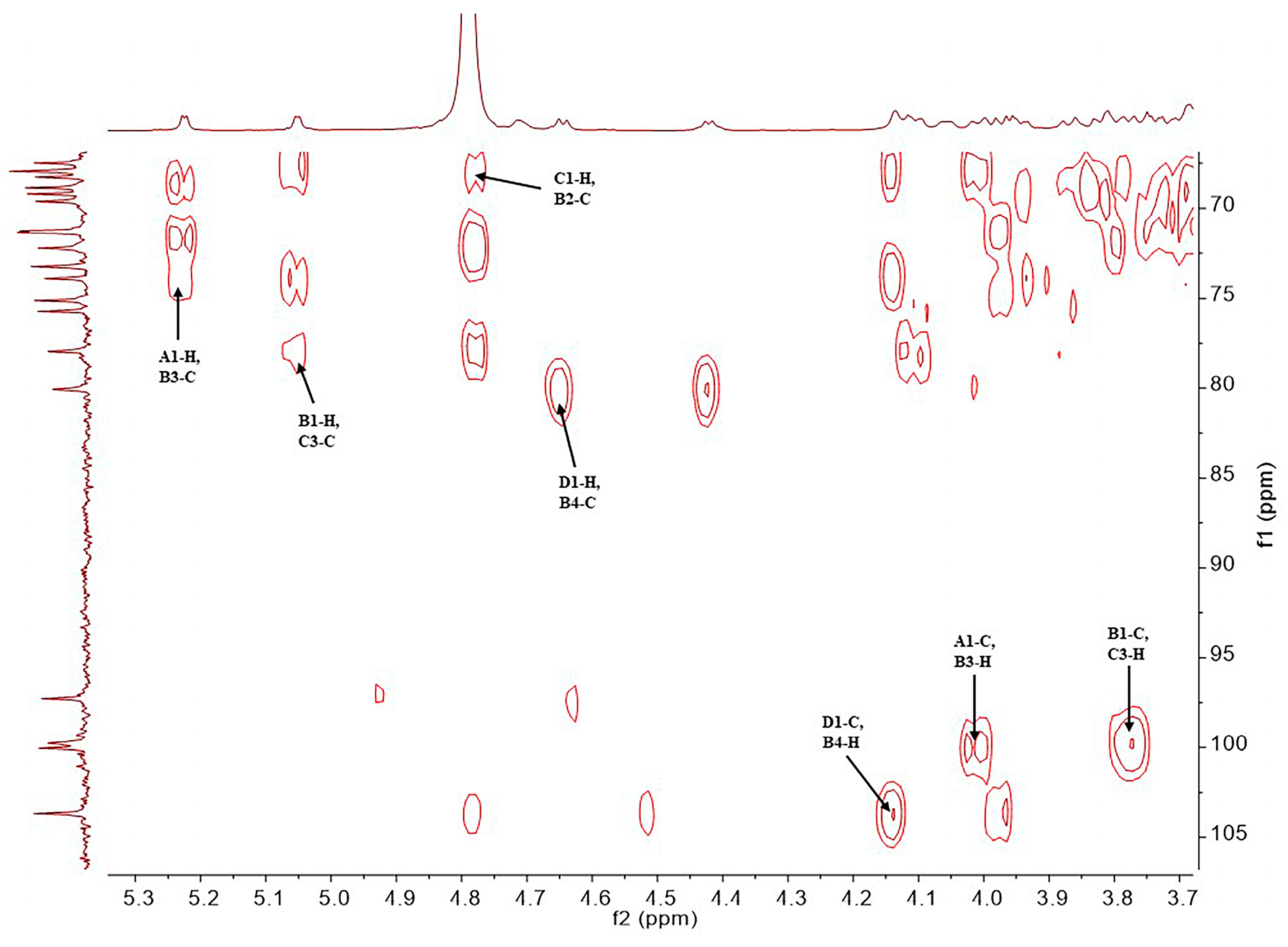
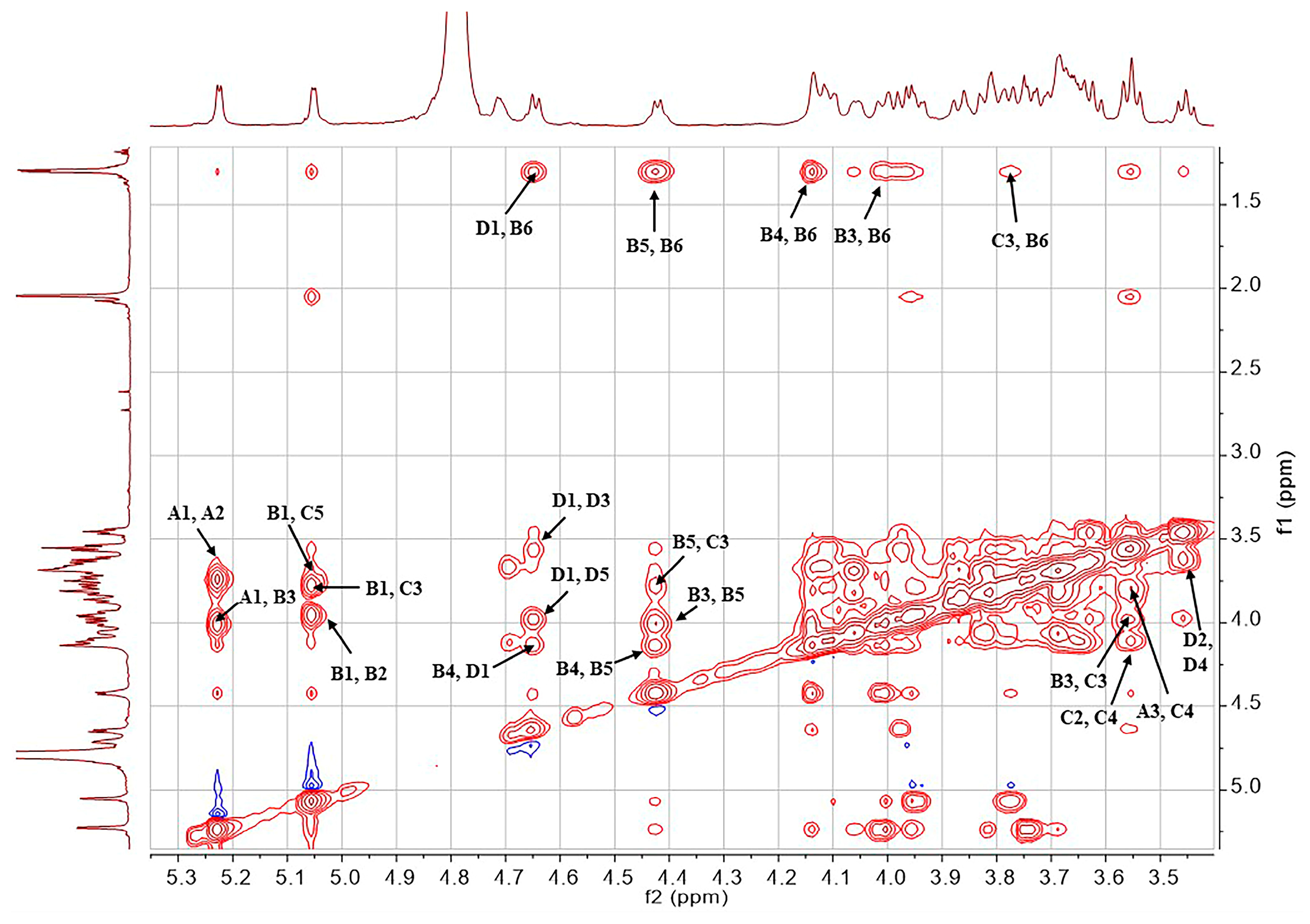
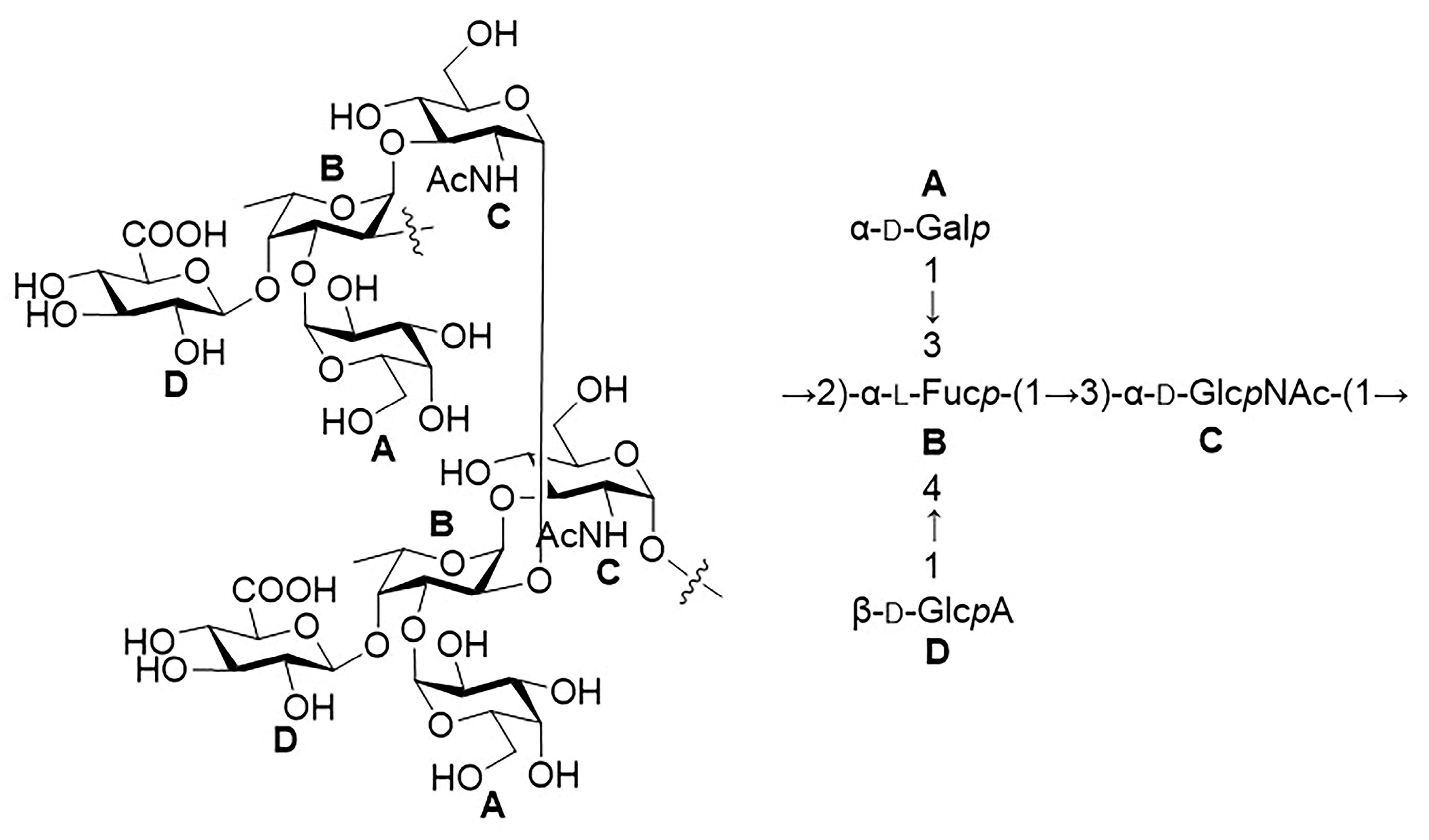
2.1. Structural Determination of SD2019180 O-Antigen
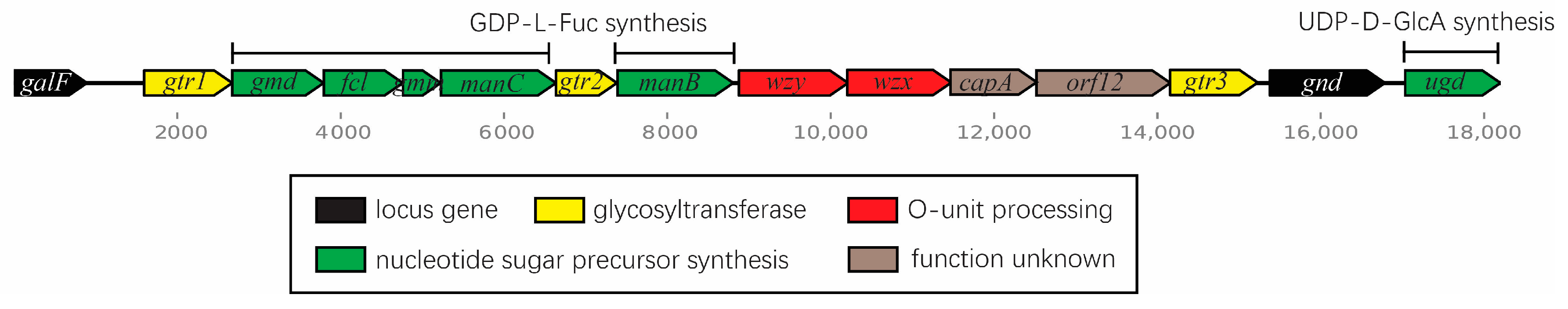
2.2. The SD2019180 O-AGC Corresponds to the Identified O-Antigen Structure
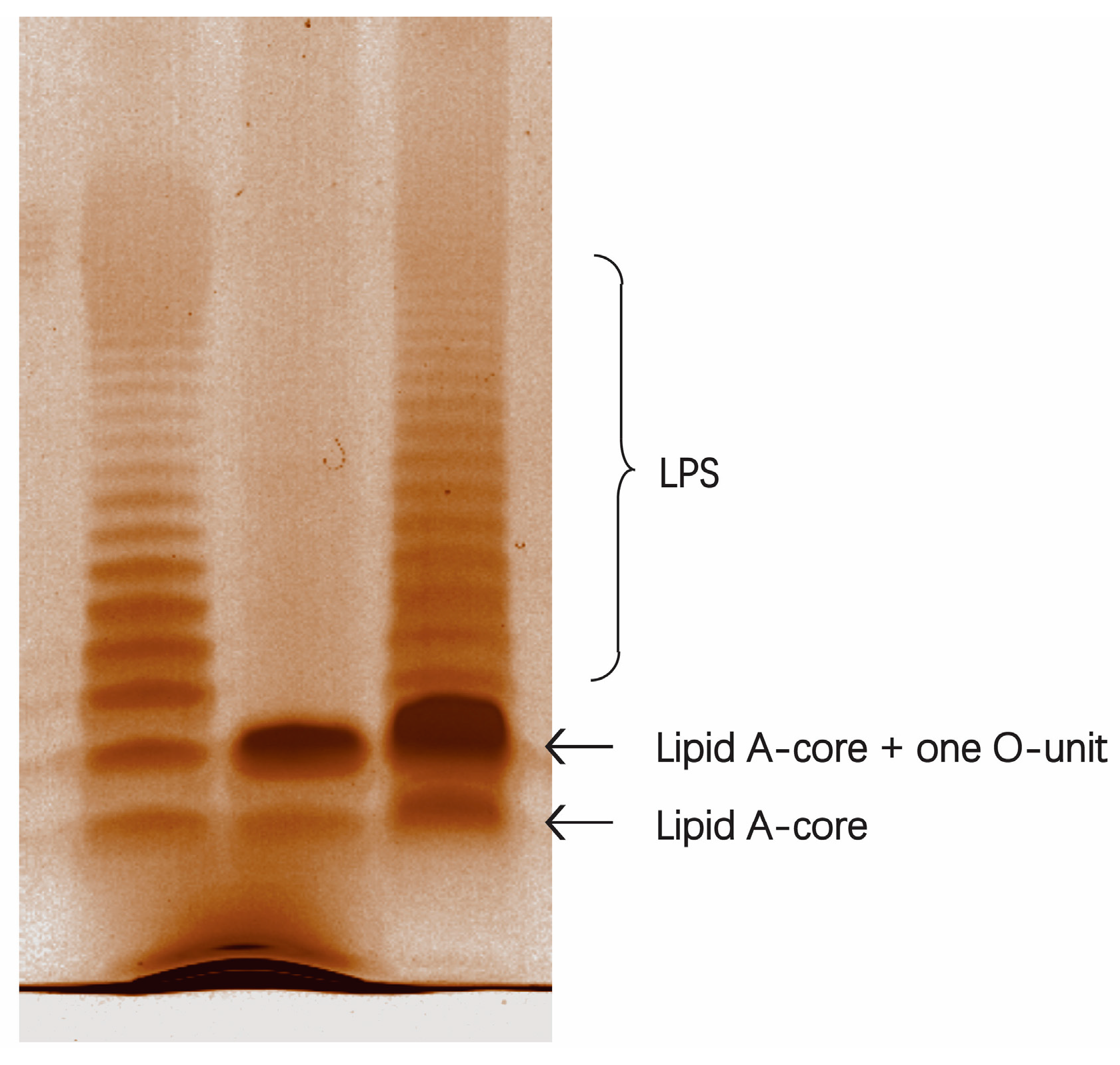
2.3. A Deletion and Complementation Test Confirmed the Role of SD2019180 O-AGC
3. Materials and Methods
3.1. LPS and O-Specific Polysaccharide Extraction
3.2. Monosaccharide Analysis
3.3. NMR Analysis
3.4. Bacterial Strains, Plasmids, and Growth Conditions
3.5. Genome Sequencing and Annotation
3.6. SDS-PAGE Analysis of LPS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okuda, S.; Sherman, D.J.; Silhavy, T.J.; Ruiz, N.; Kahne, D. Lipopolysaccharide transport and assembly at the outer membrane: The PEZ model. Nat. Rev. Microbiol. 2016, 14, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Merino, S.; Gonzalez, V.; Tomás, J.M. The first sugar of the repeat units is essential for the Wzy polymerase activity and elongation of the O-antigen lipopolysaccharide. Future Microbiol. 2016, 11, 903–918. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Reeves, P.R. The variation of O antigens in gram-negative bacteria. Subcell. Biochem. 2010, 53, 123–152. [Google Scholar]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Plainvert, C.; Bidet, P.; Peigne, C.; Barbe, V.; Medigue, C.; Denamur, E.; Bingen, E.; Bonacorsi, S. A new O-antigen gene cluster has a key role in the virulence of the Escherichia coli meningitis clone O45:K1:H7. J. Bacteriol. 2007, 189, 8528–8536. [Google Scholar] [CrossRef] [PubMed]
- Caboni, M.; Pedron, T.; Rossi, O.; Goulding, D.; Pickard, D.; Citiulo, F.; MacLennan, C.A.; Dougan, G.; Thomson, N.R.; Saul, A.; et al. An O antigen capsule modulates bacterial pathogenesis in Shigella sonnei. PLoS Pathog. 2015, 11, e1004749. [Google Scholar] [CrossRef]
- Sarkar, S.; Ulett, G.C.; Totsika, M.; Phan, M.D.; Schembri, M.A. Role of capsule and O antigen in the virulence of uropathogenic Escherichia coli. PLoS ONE 2014, 9, e94786. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiao, H.; Zhang, X.; Zhang, Y.; Sun, N.; Yang, Y.; Wei, Y.; Hu, B.; Guo, X. Two Enteropathogenic Escherichia coli Strains Representing Novel Serotypes and Investigation of Their Roles in Adhesion. J. Microbiol. Biotechnol. 2021, 31, 1191–1199. [Google Scholar] [CrossRef]
- Samuel, G.; Reeves, P. Biosynthesis of O-antigens: Genes and pathways involved in nucleotide sugar precursor synthesis and O-antigen assembly. Carbohydr. Res. 2003, 338, 2503–2519. [Google Scholar] [CrossRef]
- Keenleyside, W.J.; Whitfield, C. A novel pathway for O-polysaccharide biosynthesis in Salmonella enterica serovar Borreze. J. Biol. Chem. 1996, 271, 28581–28592. [Google Scholar] [CrossRef]
- Guan, W.; Cai, L.; Fang, J.; Wu, B.; George Wang, P. Enzymatic synthesis of UDP-GlcNAc/UDP-GalNAc analogs using N-acetylglucosamine 1-phosphate uridyltransferase (GlmU). Chem. Commun. 2009, 6976–6978. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.S.; Taylor, V.L.; Islam, S.T.; Hao, Y.; Kocincova, D. Genetic and Functional Diversity of Pseudomonas aeruginosa Lipopolysaccharide. Front. Microbiol. 2011, 2, 118. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Furevi, A.; Perepelov, A.V.; Guo, X.; Cao, H.; Wang, Q.; Reeves, P.R.; Knirel, Y.A.; Wang, L.; Widmalm, G. Structure and genetics of Escherichia coli O antigens. FEMS Microbiol. Rev. 2020, 44, 655–683. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shpirt, A.M.; Guo, X.; Shashkov, A.S.; Zhuang, Y.; Wang, L.; Knirel, Y.A.; Liu, B. Identification serologically, chemically and genetically of two Escherichia coli strains as candidates for new O serogroups. Microbiology (Reading) 2015, 161, 1790–1796. [Google Scholar] [CrossRef]
- Iguchi, A.; von Mentzer, A.; Kikuchi, T.; Thomson, N.R. An untypeable enterotoxigenic Escherichia coli represents one of the dominant types causing human disease. Microb. Genom. 2017, 3, e000121. [Google Scholar] [CrossRef]
- Lang, C.; Hiller, M.; Konrad, R.; Fruth, A.; Flieger, A. Whole-Genome-Based Public Health Surveillance of Less Common Shiga Toxin-Producing Escherichia coli Serovars and Untypeable Strains Identifies Four Novel O Genotypes. J. Clin. Microbiol. 2019, 57, e00768-19. [Google Scholar] [CrossRef]
- Wang, J.; Xu, Y.; Qin, C.; Hu, J.; Yin, J.; Guo, X. Structural Determination and Genetic Identification of the O-Antigen from an Escherichia coli Strain, LL004, Representing a Novel Serogroup. Int. J. Mol. Sci. 2021, 22, 12746. [Google Scholar] [CrossRef]
- Zhao, G.; Perepelov, A.V.; Senchenkova, S.N.; Shashkov, A.S.; Feng, L.; Li, X.; Knirel, Y.A.; Wang, L. Structural relation of the antigenic polysaccharides of Escherichia coli O40, Shigella dysenteriae type 9, and E. coli K47. Carbohydr. Res. 2007, 342, 1275–1279. [Google Scholar] [CrossRef]
- Cescutti, P.; Toffanin, R.; Pollesello, P.; Sutherland, I.W. Structural determination of the acidic exopolysaccharide produced by a Pseudomonas sp. strain 1.15. Carbohydr. Res. 1999, 315, 159–168. [Google Scholar] [CrossRef]
- Perry, M.B.; MacLean, L.L. Structural characterization of the antigenic O-chain of the lipopolysaccharide of Escherichia coli serotype O65. Carbohydr. Res. 1999, 322, 57–66. [Google Scholar] [CrossRef]
- Vinogradov, E.; Maclean, L.L.; Perry, M.B. Structural determination of the O-antigenic polysaccharide of enteropathogenic Escherichia coli O103:H2. Can. J. Microbiol. 2010, 56, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Shashkov, A.S.; Lipkind, G.M.; Knirel, Y.A.; Kochetkov, N.K. Stereochemical factors determining the effects of glycosylation on the 13C chemical shifts in carbohydrates. Magn. Reson. Chem. 1988, 26, 735–747. [Google Scholar] [CrossRef]
- Leslie, M.R.; Parolis, H.; Parolis, L.A.S.; Petersen, B.O. The capsular antigen of Escherichia coli O9:K33:H−: A polysaccharide containing both pyruvate and O-acetyl groups. Carbohydr. Res. 1998, 309, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Senchenkova, S.N.; Shashkov, A.S.; Knirel, Y.A.; Esteve, C.; Alcaide, E.; Merino, S.; Tomas, J.M. Structure of a polysaccharide from the lipopolysaccharide of Vibrio vulnificus clinical isolate YJ016 containing 2-acetimidoylamino-2-deoxy-l-galacturonic acid. Carbohydr. Res. 2009, 344, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Rosano, C.; Bisso, A.; Izzo, G.; Tonetti, M.; Sturla, L.; De Flora, A.; Bolognesi, M. Probing the catalytic mechanism of GDP-4-keto-6-deoxy-d-mannose Epimerase/Reductase by kinetic and crystallographic characterization of site-specific mutants. J. Mol. Biol. 2000, 303, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, G.; Andrianopoulos, K.; Hobbs, M.; Reeves, P.R. Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colanic acid. J. Bacteriol. 1996, 178, 4885–4893. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, J.; Vigeant, K.A.; Tatar, L.D.; Valvano, M.A. Functional characterization and membrane topology of Escherichia coli WecA, a sugar-phosphate transferase initiating the biosynthesis of enterobacterial common antigen and O-antigen lipopolysaccharide. J. Bacteriol. 2007, 189, 2618–2628. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Beness, A.M.; Saier, M.H. Computer-based analyses of the protein constituents of transport systems catalysing export of complex carbohydrates in bacteria. Microbiology (Reading) 1997, 143, 2685–2699. [Google Scholar] [CrossRef]
- Collins, R.F.; Kargas, V.; Clarke, B.R.; Siebert, C.A.; Clare, D.K.; Bond, P.J.; Whitfield, C.; Ford, R.C. Full-length, Oligomeric Structure of Wzz Determined by Cryoelectron Microscopy Reveals Insights into Membrane-Bound States. Structure 2017, 25, 806–815.e3. [Google Scholar] [CrossRef]
- Robbins, P.W.; Uchida, T. Studies on the chemical basis of the phage conversion of O-antigens in the E-group Salmonellae. Biochemistry 1962, 1, 323–335. [Google Scholar] [CrossRef]
- Fu, C.Y.; Ren, L.; Liu, W.J.; Sui, Y.; Nong, Q.N.; Xiao, Q.H.; Li, X.Q.; Cao, W. Structural characteristics of a hypoglycemic polysaccharide from Fructus Corni. Carbohydr. Res. 2021, 506, 108358. [Google Scholar] [CrossRef]
- Hugo, E.G.; Vadim, K.; Abraham, N. NMR chemical shifts of common laboratory solvents as trace impurities. J. Org. Chem. 1997, 62, 7512–7515. [Google Scholar]
- Zhao, D.; Feng, F.; Yuan, F.; Su, J.; Cheng, Y.; Wu, H.; Song, K.; Nie, B.; Yu, L.; Zhang, F. Simultaneous determination of 13 carbohydrates using high-performance anion-exchange chromatography coupled with pulsed amperometric detection and mass spectrometry. J. Sep. Sci. 2017, 40, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, T.; Wang, Q.; Huang, J.; Zhu, Y.; Liu, X.; Liu, R.; Yang, B.; Zhou, K. Vibrio cholerae senses human enteric alpha-defensin 5 through a CarSR two-component system to promote bacterial pathogenicity. Commun. Biol. 2022, 5, 559. [Google Scholar]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- MacLean, L.L.; Perry, M.B. Structural characterization of the serotype O:5 O-polysaccharide antigen of the lipopolysaccharide of Escherichia coli O:5. Biochem. Cell Biol. 1997, 75, 199–205. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | Chemical Shifts (ppm) | ||||||
|---|---|---|---|---|---|---|---|
| 1-H/1-C | 2-H/2-C | 3-H/3-C | 4-H/4-C | 5-H/5-C | 6-H/6-C | NAc (C=O) | |
| A α-d-Galp 1JH1,C1 171.5 Hz | 5.23 (3JH,H 4.1 Hz)/100.0 | 3.74 (3JH,H 12.8, 4.1 Hz)/68.9 | 3.82 (3JH,H 12.8 Hz)/69.2 | 3.69 (singlet)/69.6 | 4.06 (3JH,H 6.4 Hz)/71.3 | 3.68, 3.70/61.7 | - |
| B α-l-Fucp 1JH1,C1 171.9 Hz | 5.05 (3JH,H 3.4 Hz)/99.8 | 3.95 (3JH,H 11.4, 3.4 Hz)/67.9 | 4.02 (3JH,H 11.4 Hz)/73.9 | 4.14 (singlet)/80.1 | 4.42 (3JH,H 6.5 Hz)/67.5 | 1.30 (3JH,H 6.5 Hz)/15.2 | - |
| C α-d-GlcpNAc 1JH1,C1 172.1 Hz | 4.78/97.3 | 4.10 (3JH,H 10.2, 2.5 Hz)/53.1 | 3.77 (3JH,H 10.2, 9.2 Hz)/78.0 | 3.55 (3JH,H 9.2, 9.2 Hz)/68.3 | 3.67/72.2 | 3.80, 3.87/60.4 | 2.04/22.2 (174.5) |
| D β-d-GlcpA 1JH1,C1 161.2 Hz | 4.65 (3JH,H 7.6 Hz)/103.7 | 3.45 (3JH,H 8.6, 7.6 Hz)/73.3 | 3.57 (3JH,H 9.2, 8.6 Hz)/75.2 | 3.63/71.3 | 3.98/75.8 | -/169.0 | - |
| orf No. | Gene Name | Position of Gene | G+C Content (%) | Similar Protein(s), Strain(s) (Genbank Accession No.) | %Identical/%Similar (Total No. of aa) | Putative Function of the Protein |
|---|---|---|---|---|---|---|
| 1 | galF | 1.894 | 51.78 | GalU regulator GalF, [Escherichia coli] (EEW6173515.1) | 100/100 (297) | GalU regulator GalF |
| 2 | gtr1 | 1594.2673 | 31.01 | Glycosyltransferase family 4 protein, [Escherichia coli] (EII8714459.1) | 54/74 (359) | Glycosyltransferase |
| 3 | gmd | 2674.3792 | 42.6 | GDP-mannose 4,6-dehydratase, [Escherichia coli] (WP_096321002.1) | 99/100 (372) | GDP-mannose 4,6-dehydratase |
| 4 | fcl | 3796.4761 | 39.23 | GDP-l-fucose synthase, [Escherichia coli] (WP_044695018.1) | 99/100 (321) | GDP-l-fucose synthase |
| 5 | gmm | 4764.5225 | 37.22 | GDP-mannose mannosyl hydrolase, [Shigella sonnei] (EGD4982807.1) | 99/100 (153) | GDP-mannose mannosyl hydrolase |
| 6 | manC | 5231.6637 | 41.43 | mannose-1-phosphate guanylyltransferase/mannose-6-phosphate isomerase, [Shigella sonnei] (EGD4982806.1) | 99/99 (468) | mannose-1-phosphate guanylyltransferase/mannose-6-phosphate isomerase |
| 7 | gtr2 | 6637.7383 | 34.27 | Glycosyltransferase, [Escherichia coli] (MCB6245792.1) | 99/99 (248) | Glycosyltransferase |
| 8 | manB | 7389.8807 | 36.01 | Phosphomannomutase, [Escherichia coli] (WP_054486156.1) | 100/100 (472) | Phosphomannomutase |
| 9 | wzy | 8876.10213 | 32.21 | O-antigen polysaccharide polymerase Wzy, [Escherichia coli] (HCK1104133.1) | 90/98 (445) | O-antigen polymerase |
| 10 | wzx | 10,203.11477 | 30.03 | Oligosaccharide flippase family protein, [Escherichia coli] (EFC0723458.1) | 98/99 (424) | Flippase |
| 11 | capA | 11,467.12522 | 30.96 | CapA family protein, [Escherichia coli] (WP_205849603.1) | 99/99 (351) | CapA |
| 12 | orf12 | 12,519.14159 | 29.06 | Hypothetical protein, [Escherichia coli] (EIP2350151.1) | 99/100 (546) | Hypothetical protein |
| 13 | gtr3 | 14,156.15229 | 31.28 | Glycosyltransferase family 4 protein, [Escherichia coli] (EFN6163013.1) | 99/99 (357) | Glycosyltransferase |
| 14 | gnd | 15,380.16786 | 50.31 | NADP-dependent phosphogluconate dehydrogenase, [Escherichia coli] (WP_251885442.1) | 100/100 (468) | Phosphogluconate dehydrogenase |
| 15 | ugd | 17,035.18201 | 43.87 | UDP-glucose 6-dehydrogenase, [Escherichia coli] (WP_042048836.1) | 99/100 (338) | UDP-glucose 6-dehydrogenase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Qin, C.; Xu, Y.; Yin, J.; Hu, J.; Guo, X. Structural and Genetic Identification of the O-Antigen from an Escherichia coli Isolate, SD2019180, Representing a Novel Serogroup. Int. J. Mol. Sci. 2023, 24, 15040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015040
Wang J, Qin C, Xu Y, Yin J, Hu J, Guo X. Structural and Genetic Identification of the O-Antigen from an Escherichia coli Isolate, SD2019180, Representing a Novel Serogroup. International Journal of Molecular Sciences. 2023; 24(20):15040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015040
Chicago/Turabian StyleWang, Jing, Chunjun Qin, Yujuan Xu, Jian Yin, Jing Hu, and Xi Guo. 2023. "Structural and Genetic Identification of the O-Antigen from an Escherichia coli Isolate, SD2019180, Representing a Novel Serogroup" International Journal of Molecular Sciences 24, no. 20: 15040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015040