
Astrocytes: Lessons Learned from the Cuprizone Model


Institute of Anatomy, Rostock University Medical Center, 18057 Rostock, Germany
Int. J. Mol. Sci. 2023, 24(22), 16420; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216420
Submission received: 12 October 2023
/
Revised: 6 November 2023
/
Accepted: 14 November 2023
/
Published: 16 November 2023
(This article belongs to the Collection Feature Papers in Molecular Neurobiology)
Abstract
:A diverse array of neurological and psychiatric disorders, including multiple sclerosis, Alzheimer’s disease, and schizophrenia, exhibit distinct myelin abnormalities at both the molecular and histological levels. These aberrations are closely linked to dysfunction of oligodendrocytes and alterations in myelin structure, which may be pivotal factors contributing to the disconnection of brain regions and the resulting characteristic clinical impairments observed in these conditions. Astrocytes, which significantly outnumber neurons in the central nervous system by a five-to-one ratio, play indispensable roles in the development, maintenance, and overall well-being of neurons and oligodendrocytes. Consequently, they emerge as potential key players in the onset and progression of a myriad of neurological and psychiatric disorders. Furthermore, targeting astrocytes represents a promising avenue for therapeutic intervention in such disorders. To gain deeper insights into the functions of astrocytes in the context of myelin-related disorders, it is imperative to employ appropriate in vivo models that faithfully recapitulate specific aspects of complex human diseases in a reliable and reproducible manner. One such model is the cuprizone model, wherein metabolic dysfunction in oligodendrocytes initiates an early response involving microglia and astrocyte activation, culminating in multifocal demyelination. Remarkably, following the cessation of cuprizone intoxication, a spontaneous process of endogenous remyelination occurs. In this review article, we provide a historical overview of studies investigating the responses and putative functions of astrocytes in the cuprizone model. Following that, we list previously published works that illuminate various aspects of the biology and function of astrocytes in this multiple sclerosis model. Some of the studies are discussed in more detail in the context of astrocyte biology and pathology. Our objective is twofold: to provide an invaluable overview of this burgeoning field, and, more importantly, to inspire fellow researchers to embark on experimental investigations to elucidate the multifaceted functions of this pivotal glial cell subpopulation.
1. Introduction
The central nervous system (CNS) comprises neurons and neuroglial cells. Neurons receive and facilitate nerve impulses across their membranes to the next neuron, thus forming a fine-tuned communication network. Indispensable for fast and energy-efficient action potential propagation are the myelin sheaths built up by extended and modified plasma membranes of a specialized glia cell population—the oligodendrocytes [1]. Disorders of CNS myelin form a large and growing list of neurological disorders in humans, ranging from the most common myelin disease, multiple sclerosis (MS), to rare genetic conditions, such as Niemann–Pick disease. It is of note that oligodendrocyte dysfunctions also appear to be involved in different neurodegenerative and psychiatric disorders, including, among them, Alzheimer’s and Parkinson’s disease, eating disorders, depression, and schizophrenia [2,3,4,5,6].
MS is an autoimmune-mediated inflammatory disorder of the CNS with a still largely unknown aetiology. Two main clinical MS courses can be defined: a relapsing–remitting and a chronic, progressive course. Relapsing–remitting MS (RRMS) is characterized by recurrent episodes of clearly delineated clinical impairment (i.e., relapsing) from which the patients can eventually recover completely (i.e., remission). A relapsing–remitting disease course often turns into a secondary progressive (SPMS) course after approximately 10–15 years. In this secondary phase of the disease, individual relapses occur less frequently, but there is a slow, more or less continuous progression of the clinical impairment. The main histopathological features of MS are peripheral immune cell recruitment (id est, mostly monocytes and lymphocytes), blood–brain barrier integrity loss, reactive gliosis, oligodendrocyte damage, and, most importantly, demyelination. Mechanistically, it is believed that the disruption of the function of myelin results from an immunologically specific interaction between autoimmune T-/B-lymphocytes and myelin antigens [7].
Although demyelination is still a distinct pathological entity in MS, it has become increasingly apparent in the last two decades that substantial axonal and neuronal losses are equally important features. This phenomenon is at least partially related to the failure of remyelination. Remyelination is a regenerative process of the CNS. It occurs in three consecutive steps: (i) activation of oligodendrocyte progenitor cells (OPCs)—a widespread population of multipotent progenitors, (ii) their enrichment within areas of demyelination through migration and proliferation, and, finally, (iii) their differentiation into new myelinating oligodendrocytes [8]. It is unclear why, in some MS patients, remyelination is widespread, while, in others, it is sparse. Thus, understanding why a relatively robust regenerative process may lose momentum is an essential prerequisite for developing an effective therapeutic approach for this disease.
2. Glia Cells
Following the classical textbooks, neuronal cells can be divided into neurons and glial cells; the latter, based on their morphology, are subdivided into big (macroglia) and small (microglia) ones. While microglia constitute the innate immune cells of the CNS, macroglia are a diverse cell population, including oligodendrocytes, ependymal cells, pericytes, and astrocytes.
Astrocytes are highly heterogeneous in form and function, including the protoplasmic astrocytes residing in the grey matter, fibrous astrocytes residing in the white matter, velate astrocytes, which are localized in brain regions where small neurons are densely packed (e.g., the olfactory bulb or the granular layer of the cerebellar cortex), radial glia cells with essential guidance functions for neurons during development, the cerebellar Bergmann glia [9,10], the retinal Müller glia [11,12], pituicytes that are localized in the neurohypophysis, Gomori-astrocytes, which are prominent in the arcuate nucleus of the hypothalamus [13], and perivascular astrocytes, whose endfeet connect with blood vessels and are fundamental for the establishment of the blood–brain barriers. Furthermore, the literature describes specialized astrocytes, which are observed only in the brain of higher primates, including interlaminar astrocytes [14] and varicose projection astrocytes [15].
Astrocytes assume various essential functions, much like assistants in a theatre play. For example, astrocytes structure neuronal networks and shield neurons from each other with their extensions. They, thus, function like set designers creating the backdrop of the stage. They are assistant directors, ensuring that the “actors” (the neurons) are where they need to be and interacting with the right colleagues at the right time [16,17]. Like oligodendrocytes [18], astrocytes supply neurons with metabolites and dispose of their waste products; thus, they are responsible for both the pantry and catering service, as well as sanitary facilities and waste disposal [19,20,21]. Furthermore, astrocytes maintain the finely tuned balance of ions and transmitters. Like props, they are responsible for ensuring that the performers have their lines and costumes ready and that unneeded materials are cleared away [22,23]. Finally, astrocytes receive signals from neurons and signal back to them, influencing synaptic transmission and being capable of modifying it. They are the prompters assisting the actors on the stage (adopted from dasGehirn.info (https://www.dasgehirn.info/grundlagen/glia/astrozyten-die-heimlichen-stars-des-gehirns?gclid=CjwKCAiA9dGqBhAqEiwAmRpTCzqdOdy2G26KS9fL-T4EQuAFkhV72cF7i1-9c0j4PRTz6yytgrLYYxoCy2kQAvD_BwE (accessed on 15 November 2023)) and [24]).
Several excellent reviews have been written addressing the putative relevance of astrocytes in demyelinating disorders, including MS [25,26,27]. In this work, we will focus on astrocytic changes and their putative function in the so-called “cuprizone model,” which is frequently used to study mechanisms of oligodendrocyte degeneration, demyelination, and remyelination. Although the tabular listing of published works, which we provide at the end of this manuscript, claims to be comprehensive, we would like to focus in more detail on only a few works with groundbreaking insights for the field of research. The next chapter gives a brief overview of the most frequently used pre-clinical MS animal models, followed by a brief outline of the mechanistic and histo-pathological characteristics of the cuprizone model.
3. Multiple Sclerosis Animal Models
To study the distinct aspects of MS pathology, several different animal models exist, and they are essential for assessing the impacts of novel therapeutic approaches. One category consists of the experimental autoimmune encephalomyelitis (EAE) models induced by injecting a CNS antigen combined with Freund’s and other adjuvants to trigger an immune response. From a motor–behavioral perspective, most of the EAE models lead to ascending paralysis in mice, with the severity and clinical course depending on factors like the immunization peptide and mouse strain used. If, for example, C56BL6 mice are immunized with the MOG35–55 (myelin oligodendrocyte glycoprotein) peptide, the animals develop a chronic disease course. In contrast, a relapsing–remitting disease course develops if SJL-mice are immunized with the PLP139–151 (proteolipid protein) peptide. Histopathologically, these models are characterized by multifocal inflammatory lesions, which can mainly be found in the spinal cord, brainstem, and cerebellum.
Another group of commonly used animal models to understand MS disease pathology and to develop new therapeutic strategies are the toxin-mediated demyelination models. In these models, the toxins can be either administered via focal injection into the brain parenchyma (usually, lysophosphatidylcholine (LPC) is used) or provided to the animal per os (usually, cuprizone). LPC injection into the brain parenchyma causes rapid disruption of cellular membranes, myelin sheath disintegration, and focal demyelination within 1–2 weeks, followed by natural remyelination [28,29]. Reduced macrophage/microglial activation and myelin clearance post LPC injection in T-cell-deficient nude mice, along with inhibited remyelination in Rag-1-deficient mice lacking both B and T cells, indicate that the LPC model captures aspects of MS’s autoimmune component [30].
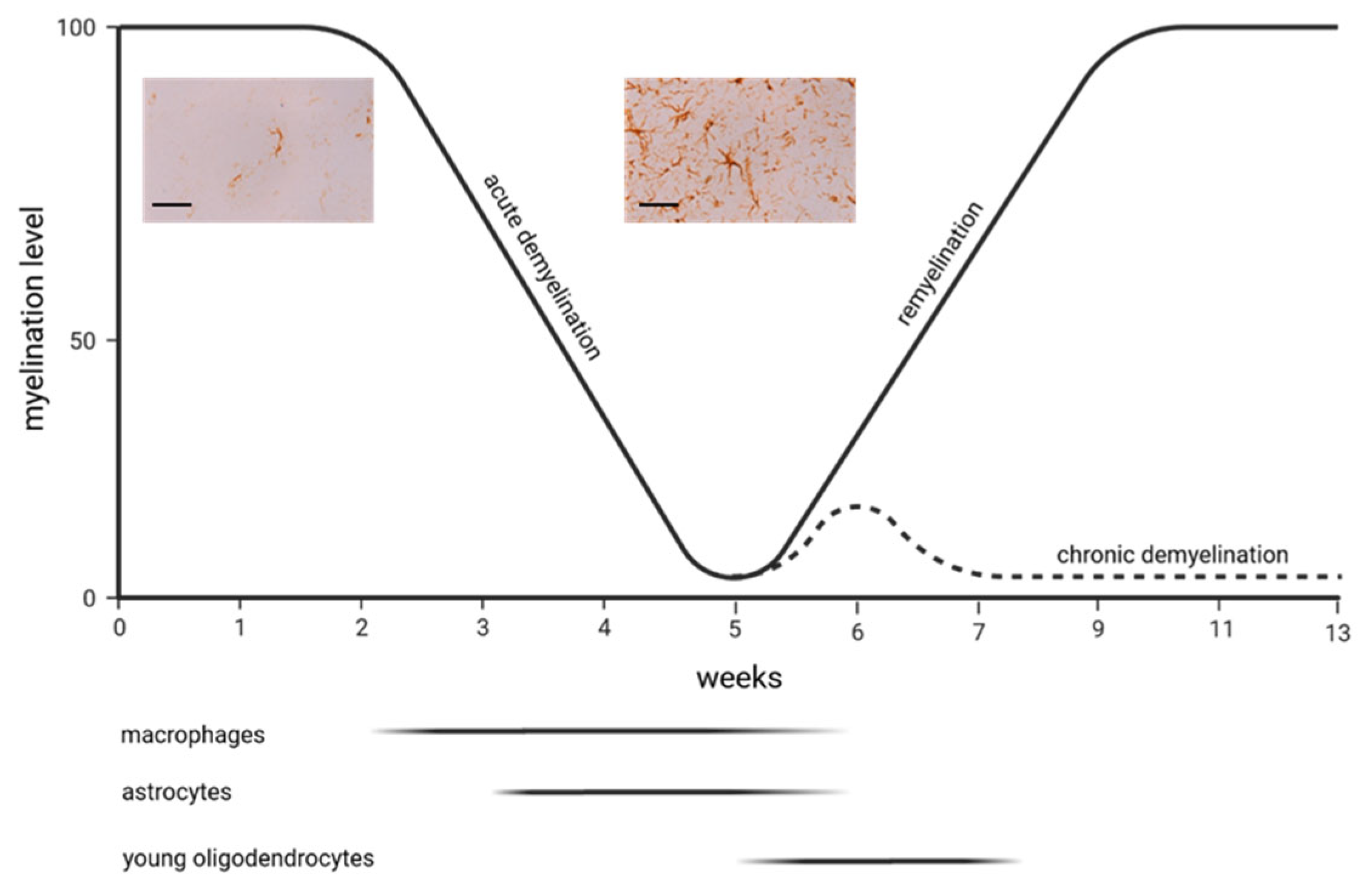
In contrast to the LPC model, demyelination in the cuprizone model occurs more slowly. Feeding young mice (around 8 weeks old) with cuprizone induces early apoptosis in oligodendrocytes, followed by microglia and macrophage activation and, finally, demyelination [31,32,33]. The exact mechanisms of oligodendrocyte death remain unclear, but it is believed to involve mitochondrial disruption due to a cuprizone-induced copper deficit, ferroptosis [34], and endoplasmic reticulum stress responses [35]. Emerging research indicates, however, that the detrimental effects of cuprizone may not be solely attributed to copper chelation or a selective impact on oligodendrocytes. Instead, it is proposed that a reactive cuprizone–copper complex is responsible for the toxic effects of cuprizone and that various cell types within the CNS are affected [36,37,38]. Why cuprizone predominately compromises oligodendrocytes is unknown, but the energy-intensive nature of myelin synthesis might play a role. Although the primary region of interest in many studies is the corpus callosum, demyelination in this model affects multiple white and grey matter brain areas [39]. During the 5–6 weeks of cuprizone exposure, the corpus callosum undergoes “acute demyelination.” When cuprizone intake is ceased at this time point, spontaneous remyelination occurs. If, however, the cuprizone intoxication is extended beyond 12–13 weeks, this leads to “chronic demyelination” with limited endogenous remyelination capacity [40]. All of the aforementioned animal models made, in the past, a significant contribution to the development of new therapeutic approaches in MS therapy. Of note is that some authors apply this model as well in the context of schizophrenia research, where myelin and oligodendrocyte abnormalities are well known [41,42].
In the next section, we will describe key papers investigating the response of astrocytes upon cuprizone-induced demyelination and functional studies that suggest that astrocytes can orchestrate de- and remyelinating pathways in this model. This section does not claim to discuss all publications relevant to astrocytes. Instead, we aim to provide a historical overview of how the function of astrocytes has been better understood over the past decades using the cuprizone model. In contrast, Table 1 lists all relevant works to provide the interested reader with an overview of the available literature.
4. Astrocytes and the Cuprizone Model
Astrocyte activation is a robust early event during cuprizone-induced demyelination [31]. Various laboratories have been able to demonstrate that during the course of cuprizone-induced demyelination, there is a significant upregulation in the expression of various astrocyte marker proteins, such as glial fibrillary acidic protein (GFAP), aldehyde dehydrogenase 1 family member L1 (ALDH1L1), and vimentin, within the demyelinated regions [43,44].
Figure 1 demonstrates findings published by Samuel Ludwin, one of the pioneers working with the cuprizone model from the late 1970s on. In Ludwin’s study, 3H-thymidine was used to label mitotic cells, revealing that alongside macrophages, astrocytes proliferate early during demyelination in the cerebellar peduncle, a frequently studied region in these initial periods of cuprizone research [45]. Despite the limitations of the applied method to label proliferating cells [46], these early studies demonstrated an early activation of astrocytes in the cuprizone model. Of note is that this first notion that prior to astrocyte activation, microglia cells respond to the cuprizone insult, was replicated by Matsushima’s lab in 1998 [47]. Some years later, when anti-GFAP labelling via immunohistochemistry became available, A. Mackenzie from the ARC Institute for Research on Animal Diseases in the UK demonstrated that astrocyte activation is widespread and not restricted to the cerebellar peduncles [39,48,49,50,51,52,53,54,55]. One of the first functional studies was published by Samuel Komoly and colleagues, who, in those days, was working at the National Institute of Neurological Disorders and Stroke, Bethesda/US, and, later, as the director of the neurology department at Pecs/Hungary. In that study, mice were intoxicated with cuprizone for 8 weeks to induce demyelination and, thereafter, provided normal chow to allow endogenous remyelination [56]. Throughout the periods of cuprizone treatment and recovery, brain tissue sections were subjected to hybridization and immunostaining with specific anti-Igf1 and Igf1-receptor probes to determine the precise locations and relative quantities of IGF-I and IGF-I receptor mRNAs and peptides. The authors found that in the white matter of untreated mice, there were no detectable levels of IGF-I or IGF-I receptor mRNAs or peptides. Conversely, in mice treated with cuprizone, astrocytes in regions where myelin was breaking down exhibited significantly elevated levels of both IGF-I mRNA and peptide. As the recovery process commenced, the expression of IGF-I in astrocytes decreased rapidly. At the same time, oligodendrocytes began to express the IGF-I receptor, suggesting that astrocyte-derived IGF1 regulates oligodendrocyte differentiation and remyelination. Although the results of subsequent studies suggested that IGF-1 is probably not a good candidate for treatment in MS [57,58], this was the first work in this model implicating that astrocytes orchestrate de- and remyelination in the diseased brain. In 2000, Brian Popkos’s lab showed that besides microglia accumulation, the extent of astrocyte activation might well be a good indicator of the overall extent of the cuprizone-induced pathology. In that study, the authors investigated the effect of IFN-gamma on myelin injury in the cuprizone model. They used a transgenic mouse line that expresses IFN-gamma under the transcriptional control of the Mbp gene. Careful evaluations revealed that mice overexpressing IFN-gamma preserved the myelin architecture, as demonstrated by LFB-PAS stains and anti-MBP immunohistochemistry. The assessment of glia reactivity showed that both microglia and astrocyte activation were less severe in the IFN-gamma-overexpressing mice, demonstrating that the quantification of glia reactivity is a good indicator of the overall extent of the cuprizone-induced pathology [59]. Our knowledge regarding possible cell–cell interactions in the cuprizone model involving astrocytes became more complex in the following years. In 2011, Matsushima’s lab showed that IL1ß-deficient mice have impaired remyelination capacities, which was paralleled by a lack of IGF-1 synthesis [60]. Because it has been shown in a previous study that astrocytes are the main source of IGF-1 in the cuprizone model, it was suggested that IL1ß might be necessary to induce IGF1 expression in astrocytes, which, in turn, promotes sufficient remyelination.
The cuprizone model also paved the way to imaging the development and progression of neuropathological processes via distinct imaging techniques, such as Positron emission tomography (PET). PET represents a sensitive method for visualizing neuropathological states, such as alterations in neuronal activity, deficiencies of neurotransmitter systems, or the extent of neuroinflammation. During PET, a small amount of a radioactive substance, known as a radiotracer, is introduced into the body. This radiotracer is usually a molecule chemically similar to a naturally occurring compound in the body, such as glucose or water, or it binds to endogenous proteins. As the radiotracer undergoes radioactive decay, it emits positrons, which can be visualized via gamma ray detection. Chen and colleagues were able to show that the mitochondrial translocator protein 18 kDa (TSPO), formerly known as the “peripheral benzodiazepine receptor,” is expressed in the cuprizone model by microglia and astrocytes [61]. Later studies, relying on this initial observation, used specific TSPO-radioligands, such as (18F)-GE180 [62] and others, to visualize activated microglia and astrocytes in living animals [43,63,64,65,66]. Zinnhardt and colleagues demonstrated that the expression of TSPO in either microglia or astrocytes is time-dependent in this model, with a predominant expression in microglia during demyelination and a shift towards astrocytic expression during remyelination [66]. Of note is that different TSPO radioligands, including (18F)-GE180, indicated equally good performance in MS patients [67,68]. Furthermore, the results of a recent study suggest that species-related differences should be considered when interpreting the results of such metabolic imaging techniques [69].
The importance of astrocytes for orchestrating de- and remyelinating events in the CNS was demonstrated in seminal work published by Stangels’s lab from Hannover/Germany. In that study, the authors used a mouse model in which thymidine kinase from the herpes simplex virus was targeted to astrocytes using the mouse GFAP promoter [70]. Ganciclovir functions as a nucleoside analogue, and it undergoes phosphorylation through the action of herpes simplex virus thymidine kinase within the GFAP-positive cell population. This phosphorylated form of ganciclovir competes with naturally occurring nucleotide triphosphates, thereby interfering with the process of DNA synthesis. Consequently, this disruption of DNA synthesis initiates apoptotic cell death, particularly in actively proliferating cells. Astrocyte ablation did not affect cuprizone-induced oligodendrocyte loss. However, without astrocytes, microglia recruitment and myelin debris clearance were hindered, leading to delayed remyelination.
In the last few years, single cell sequencing and/or single nuclei sequencing techniques were introduced, and they now allow the scientific community to investigate translatome changes of various cell types in the cuprizone model, including astrocytes. Hou and colleagues performed single-nucleus RNA sequencing from three distinct experimental categories: (1) a demyelination group, which underwent a 5-week cuprizone treatment, (2) a remyelination group, exposed to a 5-week cuprizone treatment followed by a 2-week regular chow diet, and (3) the control group, which received regular chow throughout the entire experimental period. As expected, the relative frequency of mature oligodendrocytes decreased during demyelination and recovered during remyelination [71]. Surprisingly, the frequency of astrocyte nuclei slightly dipped during demyelination but significantly recovered during remyelination. On the first view, this contrasts with the histological finding that severe astrocytosis can be observed after acute cuprizone-induced demyelination. Of note is that in this study, nuclei isolated from the corpus callosum and cortex were analyzed. Under control conditions, most cortical astrocytes express very low, almost undetectable GFAP levels, with a robust increase during demyelination. As such, the robust increase in GFAP+ cell numbers during cuprizone-induced demyelination does not really reflect the accumulation of new astrocytes. Indeed, it has been shown that pro-apoptotic pathways are activated in astrocytes after acute cuprizone-induced demyelination [35], which would explain the observed dip in the frequency of astrocyte nuclei. On the other hand, astrocyte proliferation has been demonstrated in the cuprizone model by several labs [45,72,73], so there remains a certain uncertainty about how to interpret these data. Nevertheless, although astrocyte abundance was minimally affected by cuprizone intoxication, marked transcriptional changes were evident during de- and remyelination. The most important findings of this single-nucleus RNA sequencing study can be summarized as follows. Firstly, homeostatic astrocytes are diverse at a steady state, with subpopulations expressing genes related to the extracellular matrix or neurogenesis. Nine subclusters were identified. Secondly, during demyelination, each subpopulation uniquely upregulated stress responses and mTOR pathway genes. However, certain pathways, such as the IFN-R pathway, were upregulated only in specific subpopulations. Thirdly, in the remyelination phase, pro-inflammatory clusters decreased, while some homeostatic astrocyte clusters were replaced by others. Of note is that the study also provided evidence that only minor expressional changes occur in neurons, verifying our own results that at least after acute cuprizone-induced demyelination, the degeneration of entire neurons, despite the high frequency of acute axonal injury [74], is not a characteristic feature in this model [75].
Schröder et al. applied the ribosomal tagging (RiboTag) approach to obtain insight into astrocyte function in the cuprizone model [76]. The RiboTag approach is a molecular technique used to study gene expression and protein translation within specific cell populations in complex tissues. First, transgenic animals are generated—usually mice—in which a specific ribosomal protein (often Rpl22) is genetically engineered to be tagged with a sequence (in this case, hemagglutinin A or HA), which allows for efficient isolation and purification of ribosome-bound mRNA and associated translating ribosomes. This tagged ribosomal protein is expressed under the control of a cell-type-specific promoter (in that study, under the GFAP promotor), so it is only present in a specific subset of cells within the animal. The transgenic animals are then bred or crossed with other genetically modified mice that express Cre recombinase under the control of a cell-type-specific promoter. This results in the deletion of a “floxed-stop” cassette that was preventing the expression of the tagged ribosomal protein. As a result, the tagged ribosomal protein is now expressed in the desired cell type. Tissues or cells of interest can now be sampled from these transgenic animals and subsequently analyzed.
Schröder and colleagues performed HA-tagged ribosome isolation from four distinct experimental categories: a demyelination group, which underwent a 5-week cuprizone treatment; two remyelination groups, which were exposed to 5-week cuprizone treatment followed by a 0.5- and 2-week regular chow diet; and a control group, which received regular chow throughout the entire experimental period. Upon isolation of the corpus callosum, HA-tagged ribosomes were isolated from the tissue lysate, and RNA was isolated from the HA-tagged ribosomes. Of note is that under conditions of acute demyelination (id est, 5-week cuprizone intoxication), basically all HA+ cells also expressed GFAP, which means that under such conditions, RiboTag mice are suitable for revealing astrocyte-specific transcriptomic signatures. Differential expression analysis of samples from 5-week-cuprizone-fed mice versus controls revealed 1453 differentially expressed genes, of which 1041 were up- and 412 down-regulated. Within the group of differentially upregulated genes, there were numerous chemokines (for example, Ccl6, Cxcl5, and Cxcl10), genes encoding for markers of cell activation (Cd86), and interleukins (Il3ra). Of note is that the results of recent studies suggest that CXCL10 orchestrates the activation of microglia in the cuprizone model [73,77]. In line with the single-nucleus RNA sequencing results from Hou and colleagues, in depth analysis of the RiboTag experiments suggests that astrocytes actively contribute to inflammation during demyelination but adopt a regenerative phenotype during remyelination. Another important finding from Schröder and colleagues was the existence of distinct expression signatures of HA-tagged cells during early and late remyelination, suggesting that astrocytes shape their adoptive phenotype according to the dynamic changes of oligodendrocytes during maturation and remyelination [76]. As a limitation of that study, under control conditions, approximately half of the HA+ cells were positive for APC/CC1 (a frequently used maker of mature oligodendrocytes) but did not express GFAP. The authors suggest that these HA+APC+GFAP- cells are mature oligodendrocytes that did express GFAP during their development [78,79].
5. Conclusions
For a long time, astrocytes lived in the shadows in neuroscience. After an initial focus on neurons, research turned its attention to the myelin producers of the central nervous system, the oligodendrocytes, and the cells of innate immunity, the microglia. However, astrocytes are gradually stepping out of the shadows. It is becoming increasingly clear that astrocytes do not represent a uniform cell population but are highly heterogeneous, with various essential functions. Without astrocytes, normal development and maintenance of neuronal function would not be possible. They are not just a filler substance holding the nervous system together, but rather, like ministers in a government, working diligently behind the scenes to enable higher cognitive functions. Furthermore, astrocytes play an important role in the development and progression of a number of neurological diseases, and we are just at the beginning of understanding the relevance and function of astrocytes in such diseases. This is, maybe, best illustrated with neuromyelitis optica, a neurological disorder that was, for a long time, considered to be a subvariant of MS. Today, we know that an autoimmune attack against the water channel protein aquaporin 4 is a key event during neuromyelitis optica, and this protein is expressed by astrocytes [80,81].
What did we learn by using the cuprizone model? In fact, various studies using this model were able to show that astrocytes are functionally involved in the process of oligodendrocyte damage and demyelination. Proteins expressed by astrocytes and functionally relevant for the cuprizone-induced pathological changes are, among others, the Transient Receptor Potential Ankyrin 1 (TRPA1), a nonselective cation channel with relatively high Ca2+ permeability [82], Lipocalin 2 (LCN2), also known as oncogene 24p3 or neutrophil gelatinase-associated lipocalin (NGAL), an adipocytokine implicated in various immunological functions [83], lymphotoxin-alpha, a cytotoxic protein [84], and the IκB kinase 2 pathway, which induces nuclear factor kappa B activation [85]. Regarding a possible function of astrocyte-expressed factors in the context of remyelination, among others, galectin 3 [86], the CXCR4 ligand, CXCL12 [87] BDNF [88], and the Cav1.2 voltage-gated Ca2+ channel [88] have been identified.
Whether astrocytes are beneficial or detrimental in the context of MS is discussed controversially, and this has been elaborated in several excellent review articles in the past [89,90]. On the one hand, available data suggest that astrocytes help protect demyelinated tissue from additional harm and mitigate the negative impact of neuroinflammatory activities on these tissues through the release of anti-inflammatory cytokines, like TGF-β and interleukins 10 and 27. These actions include blocking inflammatory cells from entering demyelination zones and fostering an environment conducive to remyelination. On the contrary, some studies challenge this view by attributing to astrocytes a role in exacerbating demyelination due to their release of chemokines that draw inflammatory microglial cells to the sites of injury, thereby hindering the remyelination process. Results in the cuprizone model are equally inconclusive. Two studies addressed this question by ablating astrocytes and studying the consequences for myelin repair. Skripuletz and colleagues used Ganciclovir treatment in GFAP–thymidine kinase transgenic mice to ablate astrocytes [73]. While astrocyte ablation did not influence the cuprizone-induced loss of oligodendrocytes, the removal of damaged myelin sheaths by microglia appears to be impaired, thus inhibiting the regeneration of oligodendrocytes and myelin. While Skripuletz and colleagues ablated astrocytes during active demyelination, Madadi and colleagues ablated astrocytes with La-aminoadipate (L-AAA) after chronic cuprizone-induced demyelination [91]. Under such experimental conditions, ablated animals showed better myelin and functional recovery, suggesting that astrocytes impair myelin repair in the more chronic lesion. The results of both studies are difficult to compare. On the one hand, different experimental approaches were used to ablate astrocytes (L-AAA versus thymidine kinase). On the other hand, the ablation of astrocytes was carried out at two entirely different points in time. In the first study, astrocytes were removed from a highly inflammatory environment characterized by a large amount of myelin debris, significant activation of microglia, and infiltration by oligodendrocyte precursor cells. In the second study, in contrast, astrocytes were removed from a less inflammatory environment containing no or only limited myelin debris, less pronounced microglia activation, and sparsely oligodendrocyte precursor cells. Based on our current knowledge, it appears that astrocytes initially play a supportive role in myelin regeneration, which, however, can turn negative as the lesion becomes chronic. Future studies will show whether such a simplified statement is indeed true.

{kind=link}
Table 1.
IHC (Immunohistochemistry); ISH (in situ hybridization); IGF-1 (Insulin-like growth factor 1); IFN-γ (Interferon-gamma); MIP-1alpha (Macrophage Inflammatory Protein-1 Alpha); IL-1β (Interleukin-1β); MHC (Major Histocompatibility Complex); PDGF-A (Platelet-Derived Growth Factor A); OPC (Oligodendrocyte Progenitor Cells); Notch-1 (Neurogenic locus notch homolog protein 1); ADAM12 (ADAM Metallopeptidase Domain 12); CXCL12 (C-X-C motif chemokine 12); Cx47 (Connexin 47); APP (Amyloid precursor protein); TSPO (Translocator protein); COX-1 (Cyclooxygenase-1); FABP7 (Fatty Acid Binding Protein 7); MMP-3 (Metalloproteinase-3); RXRβ (Retinoid X Receptor β); Tumor necrosis factor receptor 1 (TNFR1); GFAP (Glial fibrillary acidic protein); IL6 (Interleukine 6); mGluR (metabotropic glutamate receptor); BDNF (brain derived neurotrophic factor); OSMR (Oncostatin M receptor); NRF2 (Nuclear factor erythroid-2-related factor 2); S1P (Sphingosine-1-Phosphate); NF-κB (nuclear factor k-light-chain-enhancer of activated B cells); GLI1 (GLI Family Zinc Finger 1); SOX10 (SRY-box transcription factor 10); DDIT3 (DNA Damage Inducible Transcript 3); HO-1 (Hämoxygenase-1); PI3K (Phosphoinositide 3-kinase); CTNF (Ciliary neurotrophic factor); PAR-1 (Protease-activated receptor-1); GDNF (Glial cell line-derived neurotrophic factor); P2X7 (purinoceptor 7); 8-OHdG (8-hydroxy-2’ –deoxyguanosine); SIRT-1 (NAD-dependent protein deacetylase sirtuin-1); AQP4 (Aquaporin-4); NgR1 (Nogo-66 receptor 1); SPARC (Secreted protein acidic and rich in cysteine); EBI-2 (EBV-induced gene 2); TrkB (Tropomyosin receptor kinase B); CTR1 (High affinity copper uptake protein 1); ATP7A (ATPase copper transporting alpha); FTH1 (Ferritin Heavy Polypeptide 1); GLAST1 (GLutamate ASpartate Transporter 1); LCN2 (Lipocalin-2); ALDH1L1 (aldehyde dehydrogenase 1 family, member L1); TIMP1 (TIMP metallopeptidase inhibitor 1); TRAP1 (TNF Receptor Associated Protein 1); PTPRZ (Protein Tyrosine Phosphatase Receptor Type Z1); HNK-1 (Human natural killer-1).
Table 1.
IHC (Immunohistochemistry); ISH (in situ hybridization); IGF-1 (Insulin-like growth factor 1); IFN-γ (Interferon-gamma); MIP-1alpha (Macrophage Inflammatory Protein-1 Alpha); IL-1β (Interleukin-1β); MHC (Major Histocompatibility Complex); PDGF-A (Platelet-Derived Growth Factor A); OPC (Oligodendrocyte Progenitor Cells); Notch-1 (Neurogenic locus notch homolog protein 1); ADAM12 (ADAM Metallopeptidase Domain 12); CXCL12 (C-X-C motif chemokine 12); Cx47 (Connexin 47); APP (Amyloid precursor protein); TSPO (Translocator protein); COX-1 (Cyclooxygenase-1); FABP7 (Fatty Acid Binding Protein 7); MMP-3 (Metalloproteinase-3); RXRβ (Retinoid X Receptor β); Tumor necrosis factor receptor 1 (TNFR1); GFAP (Glial fibrillary acidic protein); IL6 (Interleukine 6); mGluR (metabotropic glutamate receptor); BDNF (brain derived neurotrophic factor); OSMR (Oncostatin M receptor); NRF2 (Nuclear factor erythroid-2-related factor 2); S1P (Sphingosine-1-Phosphate); NF-κB (nuclear factor k-light-chain-enhancer of activated B cells); GLI1 (GLI Family Zinc Finger 1); SOX10 (SRY-box transcription factor 10); DDIT3 (DNA Damage Inducible Transcript 3); HO-1 (Hämoxygenase-1); PI3K (Phosphoinositide 3-kinase); CTNF (Ciliary neurotrophic factor); PAR-1 (Protease-activated receptor-1); GDNF (Glial cell line-derived neurotrophic factor); P2X7 (purinoceptor 7); 8-OHdG (8-hydroxy-2’ –deoxyguanosine); SIRT-1 (NAD-dependent protein deacetylase sirtuin-1); AQP4 (Aquaporin-4); NgR1 (Nogo-66 receptor 1); SPARC (Secreted protein acidic and rich in cysteine); EBI-2 (EBV-induced gene 2); TrkB (Tropomyosin receptor kinase B); CTR1 (High affinity copper uptake protein 1); ATP7A (ATPase copper transporting alpha); FTH1 (Ferritin Heavy Polypeptide 1); GLAST1 (GLutamate ASpartate Transporter 1); LCN2 (Lipocalin-2); ALDH1L1 (aldehyde dehydrogenase 1 family, member L1); TIMP1 (TIMP metallopeptidase inhibitor 1); TRAP1 (TNF Receptor Associated Protein 1); PTPRZ (Protein Tyrosine Phosphatase Receptor Type Z1); HNK-1 (Human natural killer-1).
| Citation | Main Finding(s) |
|---|---|
| [46] | Astrocyte proliferation via 3H-thymidine labelling |
| [48] | Astrocyte activation, demonstrated by IHC and ISH |
| [56] | IGF-1 is expressed by astrocytes and the receptor by oligodendrocytes |
| [92] | Altered astrocytic glutathione-S-transferase isoform expression during demyelination |
| [93] | Altered astrocytic glutathione-S-transferase isoform expression during remyelination |
| [47] | Astrogliosis promptly follows microgliosis during demyelination |
| [59] | Amelioration of cuprizone-induced pathology ameliorates the extent of astrocyte activation; IFN-γ overexpression, driven by the MBP reporter |
| [94] | Amelioration of cuprizone-induced pathology ameliorates the extent of astrocyte activation; MIP-1alpha deficiency |
| [60] | IL1β-deficient mice have lower IGF-1 levels during the remyelination phase |
| [95] | Astrocytes express MHC class I and II |
| [96] | Transgenic mice that overexpress PDGF-A in astrocytes have increased OPC numbers |
| [61] | Peripheral benzodiazepine receptor is expressed by astrocytes and microglia |
| [97] | Osteopontin is expressed by astrocytes and microglia |
| [98] | Notch1 is expressed by various cell types, including astrocytes, within remyelinating lesions |
| [84] | Lymphotoxin-alpha is expressed by astrocytes and exacerbates demyelination |
| [99] | Metallothionein-I and –II are expressed by astrocytes |
| [100] | Different pathologies, including axonal injury and astrocyte activation, are more pronounced in aged versus young mice during demyelination |
| [101] | Complement regulatory protein Crry overexpression in astrocytes protects against demyelination |
| [102] | The acyl-CoA synthetase, lipidosin, is expressed by astrocytes |
| [40] | Platelet-derived growth factor-A overexpression in astrocytes supports remyelination |
| [103] | Metallothionein I/II are expressed by astrocytes |
| [39] | Extent of astrocytosis differs between demyelinated white and grey matter areas |
| [104] | Cortical demyelination, but not astrogliosis per se, is associated with accelerated cortical spreading depression |
| [105] | Astrocyte progenitor cells accumulate in cuprizone lesions |
| [106] | ADAM12 is expressed by astrocytes |
| [107] | Increased numbers of astrocytes in vivo within the subventricular zone during demyelination, and numbers were decreased by intraventricular Noggin infusion |
| [108] | CXCL12 is expressed by astrocytes and microglia |
| [109] | Cx47 is expressed by astrocytes |
| [110] | Glial isoform of APP is expressed by astrocytes |
| [111] | C3a and C5a overexpression exacerbates demyelination and delays remyelination |
| [112] | IV-injected human-embryonic-stem-cell-derived neural precursor cells into mice express GFAP to a limited extent |
| [85] | IκB kinase 2 depletion in astrocytes ameliorates demyelination |
| [113] | Smad1, Smad5, and Smad8, intracellular effectors of the bone morphogenetic protein (BMP) family of proteins, are active in oligodendrocytes and a subset of astrocytes |
| [65] | TSPO is expressed by astrocytes and microglia |
| [114] | COX-1 is expressed by astrocytes and microglia |
| [86,115] | Galectin-1 and -3 are expressed by astrocytes and microglia; galectin-3-deficient mice show impaired remyelination |
| [116] | Serine palmitoyltransferase, the rate-limiting enzyme for ceramide de novo biosynthesis, is expressed by astrocytes |
| [117] | FABP7 is expressed by astrocytes |
| [118] | IGF1 is expressed by astrocytes |
| [119] | MMP3 and MMP9 are expressed by astrocytes |
| [52] | RXRβ is expressed by astrocytes |
| [120] | Carbonic Anhydrase II is expressed by astrocytes |
| [121] | Act1-deletion in astrocytes ameliorates demyelination |
| [122] | p65 is active in astrocytes |
| [87] | TNFR1 and TNFR2 are expressed by astrocytes and microglia; CXCL12 is expressed by astrocytes, which promotes OPC proliferation and differentiation |
| [72] | In contrast to microgliosis, astrocytosis persists during de- and remyelination. Astrocyte reaction is characterized, among others features, by early astrocyte proliferation and increased expression of GFAP, vimentin, and fibronectin. Furthermore, there is an elaboration of a dense network of processes |
| [73] | Astrocyte ablation results in impaired remyelination |
| [123] | IGF1 infusions can decrease astrocyte numbers during remyelination |
| [124] | Glutamate-aspartate transporter is expressed by astrocytes |
| [125] | Receptor protein tyrosine phosphatase β is expressed in astrocytes |
| [126] | IL6 is expressed by astrocytes |
| [88] | mGluR1, mGluR5, and BDNF are expressed by astrocytes; astrocyte-derived BDNF promotes recovery from cuprizone-induced demyelination |
| [77] | CXCL10 is mainly expressed by astrocytes |
| [127] | Oncostatin M receptor is expressed by astrocytes and microglia; OSMR deficiency aggravates demyelination; CNS-targeted OSM treatment ameliorates demyelination |
| [128] | Erk is especially activated in astrocytes and promotes demyelination |
| [129] | Transgenic mice that overexpress IL6 in astrocytes show reduced glia activation, axonal injury, and OPC recruitment |
| [82] | Transient Receptor Potential Ankyrin 1 (TRPA1) is expressed in astrocytes; TRPA1 deficiency significantly attenuated cuprizone-induced demyelination by reducing the apoptosis of mature oligodendrocytes |
| [130] | Activation of the astrocytic Nrf2/ARE system ameliorates demyelination |
| [131] | Transgenic mice that overexpress IL-17A in astrocytes show aggravation of demyelination |
| [132] | Transglutaminase 2 is expressed by astrocytes |
| [133] | Transgenic mice that overexpress IL6 in astrocytes show amelioration in the cuprizone-induced pathologies |
| [134] | S1P receptor 1 is expressed by astrocytes, and its modulation ameliorates demyelination |
| [135] | Astrocytes show NF-κB activation |
| [136] | Gli1 is expressed by astrocytes after chronic, but not acute, demyelination |
| [137] | Cuprizone induces astrocyte atrophy in the rat |
| [138] | Metallothionein I/II and Megalin are expressed by astrocytes |
| [139] | Sox10 converts astrocytes into oligodendrocyte-like cells |
| [140] | Transplanted astrocytes convert into oligodendrocyte-like cells |
| [35] | DDIT3 is expressed by oligodendrocytes and astrocytes |
| [141] | Sox2 converts astrocytes into oligodendrocyte-like cells |
| [91] | Astrocyte ablation augments remyelination after chronic demyelination |
| [63] | TSPO is expressed by microglia and astrocytes |
| [142] | Transferrin can be incorporated by all glial cells among astrocytes |
| [143] | Overexpression of GFAP reduces cuprizone-induced apoptosis, demyelination, and acute axonal damage |
| [144] | Mesenchymal stem cells reduce astogliosis and microgliosis |
| [145] | CD38 is expressed by astrocytes and microglia; CD38 ameliorates demyelination |
| [146] | Astrocytes express NRF2, HO-1, and PI3K; Ginkgolide K augments the expression of these proteins |
| [147] | Astrocytes express SOX2, CNTF, IGF2, and BDNF |
| [148] | Astrocyte-specific deletion of Transient receptor potential ankyrin 1 delays demyelination |
| [149] | PAR1 knock-out mice demonstrate skewing of reactive astrocyte signatures towards a pro-repair phenotype |
| [150] | Astrocytes express BDNF and GDNF; Ginkgolide B augments the expression of these proteins |
| [151] | Deletion of astrocytic Cav1.2 channels leads to reduced astrocytosis, microgliosis, ameliorates inflammation, and promotes remyelination |
| [152] | P2x7 receptors are expressed by astrocytes and microglia; demyelination is ameliorated in P2x7-deficient mice |
| [153] | Induced neural stem cells ameliorate astrocytosis |
| [154] | CD44 is expressed by astrocytes and microglia; Cd44 deficiency does not ameliorate cuprizone-induced pathology |
| [155] | Some astrocytes show oxidative damage to DNA (id est, 8-OHdG+) |
| [156] | SIRT1 is expressed by astrocytes, microglia, and mature oligodendrocytes |
| [157] | Mesenchymal stem cell transplantation ameliorates astrocytosis |
| [158] | AQP4 is expressed in astrocyte endfeet; polarized expression is reduced after demyelination |
| [159] | Astrocytes phagocytose myelin; astrocytes express BDNF, CTNF, Nestin, SOX2, Notch, and ß-catenin; expression profiles are regulated by ethyl pyruvate |
| [160] | Astrocytes express NgR1, SPARC, and Hevin; astrocytic NgR1 sublocalization alters during demyelination; |
| [161] | Astrocytes express C3; TIC knock-outs show reduced C3 expression |
| [162] | Aastrocyte participation in the tripartite synapse during demyelination |
| [163] | mGluR5 is expressed by astrocytes and orchestrates remyelination |
| [164] | EBI2 receptor is expressed in astrocytes and microglia |
| [165] | Astrocytes express TrkB, CTR1, ATP7A, and ATP7B; demyelination is ameliorated in mice lacking astrocytic TrkB expression |
| [166] | Fth deletion in Glast1/EAAT1-positive astrocytes inhibits remyelination |
| [167] | EAAT2 is expressed by astrocytes |
| [168] | Astrocytes express S100B, GFAP, vimentin, LCN2, and ALDH1L1 |
| [83] | LCN2 is expressed by astrocytes; oligodendrocyte loss is more severe in Lcn2-/- animals |
| [169] | Ongoing astrocytosis weeks after completion of remyelination |
| [170] | C3d, S100a10, Stat3, and Timp1 are expressed by astrocytes |
| [171] | C3d and S100a10 are expressed by astrocytes; Bu Shen Yi Sui capsules promote an A2 phenotype |
| [172] | Combined mesenchymal stem cell transplantation and astrocyte ablation support remyelination |
| [173] | Cuprizone intoxication induces astrocyte endfeet sweelings |
| [174] | Astrocytes express TRAP1; β-hydroxybutyrate downregulates the expression of this protein |
| [71] | High-resolution, single-nucleus RNA sequencing (snRNA-seq) analysis of gene expression changes across various brain cells, including astrocytes |
| [175] | Bone Marrow Mesenchymal Stem Cells reduce astrocytosis |
| [176] | Astrocytes express HNK-1-O-Man+ PTPRZ |
| [177] | A1 versus A2 astrocyte expression profiling |
| [178] | The AQP4 inhibitor TGN020 ameliorates astrocyte and microglia activation |
| [76] | Astrocytic transcriptome signature investigated via ribosomal tagging |
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Lukas Wenzel for providing the anti-GFAP stained images.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hutchinson, N.A.; Koles, Z.J.; Smith, R.S. Conduction velocity in myelinated nerve fibres of Xenopus laevis. J. Physiol. 1970, 208, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Patani, R.; Hardingham, G.E.; Liddelow, S.A. Functional roles of reactive astrocytes in neuroinflammation and neurodegeneration. Nat. Rev. Neurol. 2023, 19, 395–409. [Google Scholar] [CrossRef]
- Valori, C.F.; Sulmona, C.; Brambilla, L.; Rossi, D. Astrocytes: Dissecting Their Diverse Roles in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia. Cells 2023, 12, 1450. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Ortega, P.; Soria-Ortiz, M.B.; Rodríguez, V.M.; Vázquez-Martínez, E.O.; Díaz-Muñoz, M.; Reyes-Haro, D. Anorexia disrupts glutamate-glutamine homeostasis associated with astroglia in the prefrontal cortex of young female rats. Behav. Brain Res. 2022, 420, 113715. [Google Scholar] [CrossRef]
- Als, T.D.; Kurki, M.I.; Grove, J.; Voloudakis, G.; Therrien, K.; Tasanko, E.; Nielsen, T.T.; Naamanka, J.; Veerapen, K.; Levey, D.F.; et al. Depression pathophysiology, risk prediction of recurrence and comorbid psychiatric disorders using genome-wide analyses. Nat. Med. 2023, 29, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- Farsi, Z.; Nicolella, A.; Simmons, S.K.; Aryal, S.; Shepard, N.; Brenner, K.; Lin, S.; Herzog, L.; Moran, S.P.; Stalnaker, K.J.; et al. Brain-region-specific changes in neurons and glia and dysregulation of dopamine signaling in Grin2a mutant mice. Neuron 2023, 111, 3378–3396. [Google Scholar] [CrossRef]
- Yarom, Y.; Naparstek, Y.; Lev-Ram, V.; Holoshitz, J.; Ben-Nun, A.; Cohen, I.R. Immunospecific inhibition of nerve conduction by T lymphocytes reactive to basic protein of myelin. Nature 1983, 303, 246–247. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.J.; Ffrench-Constant, C. Remyelination in the CNS: From biology to therapy. Nat. Rev. Neurosci. 2008, 9, 839–855. [Google Scholar] [CrossRef]
- Friede, R.L. The relationship of body size, nerve cell size, axon length, and glial density in the cerebellum. Proc. Natl. Acad. Sci. USA 1963, 49, 187–193. [Google Scholar] [CrossRef]
- Schwartz, A.M.; Lapham, L.W.; van den Noort, S. Cytologic and cytochemical studies of neuroglia. IV. Experimentally induced protoplasmic astrocytosis in the Bergmann glia of cerebellum. Neurology 1966, 16, 1118–1126. [Google Scholar] [CrossRef]
- Rasmussen, K.E. A morphometric study of the Müller cell cytoplasm in the rat retina. J. Ultrastruct. Res. 1972, 39, 413–429. [Google Scholar] [CrossRef]
- Rhodes, R.H. A light microscopic study of the developing human neural retina. Am. J. Anat. 1979, 154, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.J.; Pilgrim, C. A microprobe analysis of Gomori-positive glial cells in the rat arcuate nucleus. Histochemistry 1978, 55, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Colombo, J.A.; Reisin, H.D. Interlaminar astroglia of the cerebral cortex: A marker of the primate brain. Brain Res. 2004, 1006, 126–131. [Google Scholar] [CrossRef]
- Falcone, C.; McBride, E.L.; Hopkins, W.D.; Hof, P.R.; Manger, P.R.; Sherwood, C.C.; Noctor, S.C.; Martínez-Cerdeño, V. Redefining varicose projection astrocytes in primates. Glia 2022, 70, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Yates, D. Astrocytic cueing of neuronal migration. Nat. Rev. Neurosci. 2020, 21, 120. [Google Scholar] [CrossRef]
- Su, J.; Charalambakis, N.E.; Sabbagh, U.; Somaiya, R.D.; Monavarfeshani, A.; Guido, W.; Fox, M.A. Retinal inputs signal astrocytes to recruit interneurons into visual thalamus. Proc. Natl. Acad. Sci. USA 2020, 117, 2671–2682. [Google Scholar] [CrossRef]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef]
- MacVicar, B.A.; Choi, H.B. Astrocytes Provide Metabolic Support for Neuronal Synaptic Function in Response to Extracellular K(). Neurochem. Res. 2017, 42, 2588–2594. [Google Scholar] [CrossRef]
- Bonvento, G.; Bolaños, J.P. Astrocyte-neuron metabolic cooperation shapes brain activity. Cell Metab. 2021, 33, 1546–1564. [Google Scholar] [CrossRef]
- Roumes, H.; Jollé, C.; Blanc, J.; Benkhaled, I.; Chatain, C.P.; Massot, P.; Raffard, G.; Bouchaud, V.; Biran, M.; Pythoud, C.; et al. Lactate transporters in the rat barrel cortex sustain whisker-dependent BOLD fMRI signal and behavioral performance. Proc. Natl. Acad. Sci. USA 2021, 118, e2112466118. [Google Scholar] [CrossRef]
- Kress, B.T.; Iliff, J.J.; Xia, M.; Wang, M.; Wei, H.S.; Zeppenfeld, D.; Xie, L.; Kang, H.; Xu, Q.; Liew, J.A.; et al. Impairment of paravascular clearance pathways in the aging brain. Ann. Neurol. 2014, 76, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Hablitz, L.M.; Plá, V.; Giannetto, M.; Vinitsky, H.S.; Stæger, F.F.; Metcalfe, T.; Nguyen, R.; Benrais, A.; Nedergaard, M. Circadian control of brain glymphatic and lymphatic fluid flow. Nat. Commun. 2020, 11, 4411. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Charabati, M.; Wheeler, M.A.; Weiner, H.L.; Quintana, F.J. Multiple sclerosis: Neuroimmune crosstalk and therapeutic targeting. Cell 2023, 186, 1309–1327. [Google Scholar] [CrossRef] [PubMed]
- Rawji, K.S.; Neumann, B.; Franklin, R.J.M. Glial aging and its impact on central nervous system myelin regeneration. Ann. N. Y. Acad. Sci. 2023, 1519, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Healy, L.M.; Stratton, J.A.; Kuhlmann, T.; Antel, J. The role of glial cells in multiple sclerosis disease progression. Nat. Rev. Neurol. 2022, 18, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.W.; Keough, M.B.; Haylock-Jacobs, S.; Cua, R.; Döring, A.; Sloka, S.; Stirling, D.P.; Rivest, S.; Yong, V.W. Chondroitin sulfate proteoglycans in demyelinated lesions impair remyelination. Ann. Neurol. 2012, 72, 419–432. [Google Scholar] [CrossRef]
- Huang, J.K.; Jarjour, A.A.; Nait Oumesmar, B.; Kerninon, C.; Williams, A.; Krezel, W.; Kagechika, H.; Bauer, J.; Zhao, C.; Baron-Van Evercooren, A.; et al. Retinoid X receptor gamma signaling accelerates CNS remyelination. Nat. Neurosci. 2011, 14, 45–53. [Google Scholar] [CrossRef]
- Ghasemlou, N.; Jeong, S.Y.; Lacroix, S.; David, S. T cells contribute to lysophosphatidylcholine-induced macrophage activation and demyelination in the CNS. Glia 2007, 55, 294–302. [Google Scholar] [CrossRef]
- Buschmann, J.P.; Berger, K.; Awad, H.; Clarner, T.; Beyer, C.; Kipp, M. Inflammatory response and chemokine expression in the white matter corpus callosum and gray matter cortex region during cuprizone-induced demyelination. J. Mol. Neurosci. 2012, 48, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Voss, E.V.; Škuljec, J.; Gudi, V.; Skripuletz, T.; Pul, R.; Trebst, C.; Stangel, M. Characterisation of microglia during de- and remyelination: Can they create a repair promoting environment? Neurobiol. Dis. 2012, 45, 519–528. [Google Scholar] [CrossRef]
- Blakemore, W.F. Observations on oligodendrocyte degeneration, the resolution of status spongiosus and remyelination in cuprizone intoxication in mice. J. Neurocytol. 1972, 1, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Jhelum, P.; Santos-Nogueira, E.; Teo, W.; Haumont, A.; Lenoël, I.; Stys, P.K.; David, S. Ferroptosis Mediates Cuprizone-Induced Loss of Oligodendrocytes and Demyelination. J. Neurosci. 2020, 40, 9327–9341. [Google Scholar] [CrossRef]
- Fischbach, F.; Nedelcu, J.; Leopold, P.; Zhan, J.; Clarner, T.; Nellessen, L.; Beißel, C.; van Heuvel, Y.; Goswami, A.; Weis, J.; et al. Cuprizone-induced graded oligodendrocyte vulnerability is regulated by the transcription factor DNA damage-inducible transcript 3. Glia 2019, 67, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, L.A.; Calatayud, C.A.; Bertone Uña, A.L.; Millet, V.; Pasquini, J.M.; Soto, E.F. The neurotoxic effect of cuprizone on oligodendrocytes depends on the presence of pro-inflammatory cytokines secreted by microglia. Neurochem. Res. 2007, 32, 279–292. [Google Scholar] [CrossRef]
- Taraboletti, A.; Walker, T.; Avila, R.; Huang, H.; Caporoso, J.; Manandhar, E.; Leeper, T.C.; Modarelli, D.A.; Medicetty, S.; Shriver, L.P. Cuprizone Intoxication Induces Cell Intrinsic Alterations in Oligodendrocyte Metabolism Independent of Copper Chelation. Biochemistry 2017, 56, 1518–1528. [Google Scholar] [CrossRef]
- Morgan, M.L.; Teo, W.; Hernandez, Y.; Brideau, C.; Cummins, K.; Kuipers, H.F.; Stys, P.K. Cuprizone-induced Demyelination in Mouse Brain is not due to Depletion of Copper. ASN Neuro 2022, 14, 17590914221126367. [Google Scholar] [CrossRef]
- Gudi, V.; Moharregh-Khiabani, D.; Skripuletz, T.; Koutsoudaki, P.N.; Kotsiari, A.; Skuljec, J.; Trebst, C.; Stangel, M. Regional differences between grey and white matter in cuprizone induced demyelination. Brain Res. 2009, 1283, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Vana, A.C.; Flint, N.C.; Harwood, N.E.; Le, T.Q.; Fruttiger, M.; Armstrong, R.C. Platelet-derived growth factor promotes repair of chronically demyelinated white matter. J. Neuropathol. Exp. Neurol. 2007, 66, 975–988. [Google Scholar] [CrossRef]
- Fessel, J. Abnormal oligodendrocyte function in schizophrenia explains the long latent interval in some patients. Transl. Psychiatry 2022, 12, 120. [Google Scholar] [CrossRef]
- Gouvêa-Junqueira, D.; Falvella, A.C.B.; Antunes, A.; Seabra, G.; Brandão-Teles, C.; Martins-de-Souza, D.; Crunfli, F. Novel Treatment Strategies Targeting Myelin and Oligodendrocyte Dysfunction in Schizophrenia. Front. Psychiatry 2020, 11, 379. [Google Scholar] [CrossRef]
- Behrangi, N.; Heinig, L.; Frintrop, L.; Santrau, E.; Kurth, J.; Krause, B.; Atanasova, D.; Clarner, T.; Fragoulis, A.; Joksch, M.; et al. Siponimod ameliorates metabolic oligodendrocyte injury via the sphingosine-1 phosphate receptor 5. Proc. Natl. Acad. Sci. USA 2022, 119, e2204509119. [Google Scholar] [CrossRef]
- Gudi, V.; Gingele, S.; Skripuletz, T.; Stangel, M. Glial response during cuprizone-induced de- and remyelination in the CNS: Lessons learned. Front. Cell. Neurosci. 2014, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Ludwin, S.K. An autoradiographic study of cellular proliferation in remyelination of the central nervous system. Am. J. Pathol. 1979, 95, 683–696. [Google Scholar] [PubMed]
- Hu, V.W.; Black, G.E.; Torres-Duarte, A.; Abramson, F.P. 3H-thymidine is a defective tool with which to measure rates of DNA synthesis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 1456–1457. [Google Scholar] [CrossRef]
- Hiremath, M.M.; Saito, Y.; Knapp, G.W.; Ting, J.P.; Suzuki, K.; Matsushima, G.K. Microglial/macrophage accumulation during cuprizone-induced demyelination in C57BL/6 mice. J. Neuroimmunol. 1998, 92, 38–49. [Google Scholar] [CrossRef]
- Mackenzie, A. Immunohistochemical demonstration of glial fibrillary acidic protein in scrapie. J. Comp. Pathol. 1983, 93, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Norkute, A.; Hieble, A.; Braun, A.; Johann, S.; Clarner, T.; Baumgartner, W.; Beyer, C.; Kipp, M. Cuprizone treatment induces demyelination and astrocytosis in the mouse hippocampus. J. Neurosci. Res. 2009, 87, 1343–1355. [Google Scholar] [CrossRef]
- Koutsoudaki, P.N.; Skripuletz, T.; Gudi, V.; Moharregh-Khiabani, D.; Hildebrandt, H.; Trebst, C.; Stangel, M. Demyelination of the hippocampus is prominent in the cuprizone model. Neurosci. Lett. 2009, 451, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Skripuletz, T.; Bussmann, J.H.; Gudi, V.; Koutsoudaki, P.N.; Pul, R.; Moharregh-Khiabani, D.; Lindner, M.; Stangel, M. Cerebellar cortical demyelination in the murine cuprizone model. Brain Pathol. 2010, 20, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Steelman, A.J.; Thompson, J.P.; Li, J. Demyelination and remyelination in anatomically distinct regions of the corpus callosum following cuprizone intoxication. Neurosci. Res. 2012, 72, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Awad, H.; Slowik, A.; Beyer, C.; Kipp, M.; Clarner, T. Regional heterogeneity of cuprizone-induced demyelination: Topographical aspects of the midline of the corpus callosum. J. Mol. Neurosci. 2013, 49, 80–88. [Google Scholar] [CrossRef]
- Wagenknecht, N.; Becker, B.; Scheld, M.; Beyer, C.; Clarner, T.; Hochstrasser, T.; Kipp, M. Thalamus Degeneration and Inflammation in Two Distinct Multiple Sclerosis Animal Models. J. Mol. Neurosci. 2016, 60, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Sen, M.K.; Almuslehi, M.S.M.; Coorssen, J.R.; Mahns, D.A.; Shortland, P.J. Behavioural and histological changes in cuprizone-fed mice. Brain Behav. Immun. 2020, 87, 508–523. [Google Scholar] [CrossRef]
- Komoly, S.; Hudson, L.D.; Webster, H.D.; Bondy, C.A. Insulin-like growth factor I gene expression is induced in astrocytes during experimental demyelination. Proc. Natl. Acad. Sci. USA 1992, 89, 1894–1898. [Google Scholar] [CrossRef] [PubMed]
- Cannella, B.; Pitt, D.; Capello, E.; Raine, C.S. Insulin-like growth factor-1 fails to enhance central nervous system myelin repair during autoimmune demyelination. Am. J. Pathol. 2000, 157, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Lovett-Racke, A.E.; Bittner, P.; Cross, A.H.; Carlino, J.A.; Racke, M.K. Regulation of experimental autoimmune encephalomyelitis with insulin-like growth factor (IGF-1) and IGF-1/IGF-binding protein-3 complex (IGF-1/IGFBP3). J. Clin. Investig. 1998, 101, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Gillig, T.A.; Ye, P.; D’Ercole, A.J.; Matsushima, G.K.; Popko, B. Interferon-gamma protects against cuprizone-induced demyelination. Mol. Cell. Neurosci. 2000, 16, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.L.; Suzuki, K.; Chaplin, D.D.; Matsushima, G.K. Interleukin-1beta promotes repair of the CNS. J. Neurosci. 2001, 21, 7046–7052. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.K.; Baidoo, K.; Verina, T.; Guilarte, T.R. Peripheral benzodiazepine receptor imaging in CNS demyelination: Functional implications of anatomical and cellular localization. Brain 2004, 127 Pt 6, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Wickstrøm, T.; Clarke, A.; Gausemel, I.; Horn, E.; Jørgensen, K.; Khan, I.; Mantzilas, D.; Rajanayagam, T.; in ’t Veld, D.J.; Trigg, W. The development of an automated and GMP compliant FASTlab™ Synthesis of [(18) F]GE-180; a radiotracer for imaging translocator protein (TSPO). J. Label. Compd. Radiopharm. 2014, 57, 42–48. [Google Scholar] [CrossRef]
- Nack, A.; Brendel, M.; Nedelcu, J.; Daerr, M.; Nyamoya, S.; Beyer, C.; Focke, C.; Deussing, M.; Hoornaert, C.; Ponsaerts, P.; et al. Expression of Translocator Protein and [18F]-GE180 Ligand Uptake in Multiple Sclerosis Animal Models. Cells 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.K.; Guilarte, T.R. Imaging the peripheral benzodiazepine receptor response in central nervous system demyelination and remyelination. Toxicol. Sci. 2006, 91, 532–539. [Google Scholar] [CrossRef]
- Mattner, F.; Bandin, D.L.; Staykova, M.; Berghofer, P.; Gregoire, M.C.; Ballantyne, P.; Quinlivan, M.; Fordham, S.; Pham, T.; Willenborg, D.O.; et al. Evaluation of [¹²³I]-CLINDE as a potent SPECT radiotracer to assess the degree of astroglia activation in cuprizone-induced neuroinflammation. Eur. J. Nucl. Med. Mol. Imaging 2011, 38, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Zinnhardt, B.; Belloy, M.; Fricke, I.B.; Orije, J.; Guglielmetti, C.; Hermann, S.; Wagner, S.; Schäfers, M.; Van der Linden, A.; Jacobs, A.H. Molecular Imaging of Immune Cell Dynamics During De- and Remyelination in the Cuprizone Model of Multiple Sclerosis by [(18)F]DPA-714 PET and MRI. Theranostics 2019, 9, 1523–1537. [Google Scholar] [CrossRef]
- Unterrainer, M.; Mahler, C.; Vomacka, L.; Lindner, S.; Havla, J.; Brendel, M.; Böning, G.; Ertl-Wagner, B.; Kümpfel, T.; Milenkovic, V.M.; et al. TSPO PET with [(18)F]GE-180 sensitively detects focal neuroinflammation in patients with relapsing-remitting multiple sclerosis. Eur. J. Nucl. Med. Mol. Imaging 2018, 45, 1423–1431. [Google Scholar] [CrossRef]
- Vomacka, L.; Albert, N.L.; Lindner, S.; Unterrainer, M.; Mahler, C.; Brendel, M.; Ermoschkin, L.; Gosewisch, A.; Brunegraf, A.; Buckley, C.; et al. TSPO imaging using the novel PET ligand [(18)F]GE-180: Quantification approaches in patients with multiple sclerosis. EJNMMI Res. 2017, 7, 89. [Google Scholar] [CrossRef]
- Nutma, E.; Fancy, N.; Weinert, M.; Tsartsalis, S.; Marzin, M.C.; Muirhead, R.C.J.; Falk, I.; Breur, M.; de Bruin, J.; Hollaus, D.; et al. Translocator protein is a marker of activated microglia in rodent models but not human neurodegenerative diseases. Nat. Commun. 2023, 14, 5247. [Google Scholar] [CrossRef] [PubMed]
- Bush, T.G.; Savidge, T.C.; Freeman, T.C.; Cox, H.J.; Campbell, E.A.; Mucke, L.; Johnson, M.H.; Sofroniew, M.V. Fulminant jejuno-ileitis following ablation of enteric glia in adult transgenic mice. Cell 1998, 93, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Zhou, Y.; Cai, Z.; Terekhova, M.; Swain, A.; Andhey, P.S.; Guimaraes, R.M.; Ulezko Antonova, A.; Qiu, T.; Sviben, S.; et al. Transcriptomic atlas and interaction networks of brain cells in mouse CNS demyelination and remyelination. Cell Rep. 2023, 42, 112293. [Google Scholar] [CrossRef] [PubMed]
- Hibbits, N.; Yoshino, J.; Le, T.Q.; Armstrong, R.C. Astrogliosis during acute and chronic cuprizone demyelination and implications for remyelination. ASN Neuro 2012, 4, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Skripuletz, T.; Hackstette, D.; Bauer, K.; Gudi, V.; Pul, R.; Voss, E.; Berger, K.; Kipp, M.; Baumgärtner, W.; Stangel, M. Astrocytes regulate myelin clearance through recruitment of microglia during cuprizone-induced demyelination. Brain 2013, 136 Pt 1, 147–167. [Google Scholar] [CrossRef] [PubMed]
- Höflich, K.M.; Beyer, C.; Clarner, T.; Schmitz, C.; Nyamoya, S.; Kipp, M.; Hochstrasser, T. Acute axonal damage in three different murine models of multiple sclerosis: A comparative approach. Brain Res. 2016, 1650, 125–133. [Google Scholar] [CrossRef]
- Hochstrasser, T.; Rühling, S.; Hecher, K.; Fabisch, K.H.; Chrzanowski, U.; Brendel, M.; Eckenweber, F.; Sacher, C.; Schmitz, C.; Kipp, M. Stereological Investigation of Regional Brain Volumes after Acute and Chronic Cuprizone-Induced Demyelination. Cells 2019, 8, 1024. [Google Scholar] [CrossRef] [PubMed]
- Schröder, L.J.; Mulenge, F.; Pavlou, A.; Skripuletz, T.; Stangel, M.; Gudi, V.; Kalinke, U. Dynamics of reactive astrocytes fosters tissue regeneration after cuprizone-induced demyelination. Glia 2023, 71, 2573–2590. [Google Scholar] [CrossRef] [PubMed]
- Clarner, T.; Janssen, K.; Nellessen, L.; Stangel, M.; Skripuletz, T.; Krauspe, B.; Hess, F.M.; Denecke, B.; Beutner, C.; Linnartz-Gerlach, B.; et al. CXCL10 triggers early microglial activation in the cuprizone model. J. Immunol. 2015, 194, 3400–3413. [Google Scholar] [CrossRef]
- Casper, K.B.; McCarthy, K.D. GFAP-positive progenitor cells produce neurons and oligodendrocytes throughout the CNS. Mol. Cell. Neurosci. 2006, 31, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Barragan, J.; Bashiruddin, S.; Smith, C.J.; Tyrrell, C.; Parsons, M.J.; Doris, R.; Kucenas, S.; Downes, G.B.; Velez, C.M.; et al. Gfap-positive radial glial cells are an essential progenitor population for later-born neurons and glia in the zebrafish spinal cord. Glia 2016, 64, 1170–1189. [Google Scholar] [CrossRef]
- Lennon, V.A.; Kryzer, T.J.; Pittock, S.J.; Verkman, A.S.; Hinson, S.R. IgG marker of optic-spinal multiple sclerosis binds to the aquaporin-4 water channel. J. Exp. Med. 2005, 202, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Hinson, S.R.; Pittock, S.J.; Lucchinetti, C.F.; Roemer, S.F.; Fryer, J.P.; Kryzer, T.J.; Lennon, V.A. Pathogenic potential of IgG binding to water channel extracellular domain in neuromyelitis optica. Neurology 2007, 69, 2221–2231. [Google Scholar] [CrossRef]
- Sághy, É.; Sipos, É.; Ács, P.; Bölcskei, K.; Pohóczky, K.; Kemény, Á.; Sándor, Z.; Szőke, É.; Sétáló, G., Jr.; Komoly, S.; et al. TRPA1 deficiency is protective in cuprizone-induced demyelination-A new target against oligodendrocyte apoptosis. Glia 2016, 64, 2166–2180. [Google Scholar] [CrossRef]
- Gasterich, N.; Bohn, A.; Sesterhenn, A.; Nebelo, F.; Fein, L.; Kaddatz, H.; Nyamoya, S.; Kant, S.; Kipp, M.; Weiskirchen, R.; et al. Lipocalin 2 attenuates oligodendrocyte loss and immune cell infiltration in mouse models for multiple sclerosis. Glia 2022, 70, 2188–2206. [Google Scholar] [CrossRef]
- Plant, S.R.; Arnett, H.A.; Ting, J.P. Astroglial-derived lymphotoxin-alpha exacerbates inflammation and demyelination, but not remyelination. Glia 2005, 49, 1–14. [Google Scholar] [CrossRef]
- Raasch, J.; Zeller, N.; van Loo, G.; Merkler, D.; Mildner, A.; Erny, D.; Knobeloch, K.P.; Bethea, J.R.; Waisman, A.; Knust, M.; et al. IkappaB kinase 2 determines oligodendrocyte loss by non-cell-autonomous activation of NF-kappaB in the central nervous system. Brain 2011, 134 Pt 4, 1184–1198. [Google Scholar] [CrossRef]
- Pasquini, L.A.; Millet, V.; Hoyos, H.C.; Giannoni, J.P.; Croci, D.O.; Marder, M.; Liu, F.T.; Rabinovich, G.A.; Pasquini, J.M. Galectin-3 drives oligodendrocyte differentiation to control myelin integrity and function. Cell Death Differ. 2011, 18, 1746–1756. [Google Scholar] [CrossRef]
- Patel, J.R.; Williams, J.L.; Muccigrosso, M.M.; Liu, L.; Sun, T.; Rubin, J.B.; Klein, R.S. Astrocyte TNFR2 is required for CXCL12-mediated regulation of oligodendrocyte progenitor proliferation and differentiation within the adult CNS. Acta Neuropathol. 2012, 124, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Fulmer, C.G.; VonDran, M.W.; Stillman, A.A.; Huang, Y.; Hempstead, B.L.; Dreyfus, C.F. Astrocyte-derived BDNF supports myelin protein synthesis after cuprizone-induced demyelination. J. Neurosci. 2014, 34, 8186–8196. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Piaton, G.; Lubetzki, C. Astrocytes--friends or foes in multiple sclerosis? Glia 2007, 55, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Correale, J.; Farez, M.F. The Role of Astrocytes in Multiple Sclerosis Progression. Front. Neurol. 2015, 6, 180. [Google Scholar] [CrossRef]
- Madadi, S.; Pasbakhsh, P.; Tahmasebi, F.; Mortezaee, K.; Khanehzad, M.; Boroujeni, F.B.; Noorzehi, G.; Kashani, I.R. Astrocyte ablation induced by La-aminoadipate (L-AAA) potentiates remyelination in a cuprizone demyelinating mouse model. Metab. Brain Dis. 2019, 34, 593–603. [Google Scholar] [CrossRef]
- Cammer, W.; Zhang, H. Atypical localization of the oligodendrocytic isoform (PI) of glutathione-S-transferase in astrocytes during cuprizone intoxication. J. Neurosci. Res. 1993, 36, 183–190. [Google Scholar] [CrossRef]
- Tansey, F.A.; Zhang, H.; Cammer, W. Rapid upregulation of the Pi isoform of glutathione-S-transferase in mouse brains after withdrawal of the neurotoxicant, cuprizone. Mol. Chem. Neuropathol. 1997, 31, 161–170. [Google Scholar] [CrossRef]
- McMahon, E.J.; Cook, D.N.; Suzuki, K.; Matsushima, G.K. Absence of macrophage-inflammatory protein-1alpha delays central nervous system demyelination in the presence of an intact blood-brain barrier. J. Immunol. 2001, 167, 2964–2971. [Google Scholar] [CrossRef]
- Arnett, H.A.; Wang, Y.; Matsushima, G.K.; Suzuki, K.; Ting, J.P. Functional genomic analysis of remyelination reveals importance of inflammation in oligodendrocyte regeneration. J. Neurosci. 2003, 23, 9824–9832. [Google Scholar] [CrossRef]
- Woodruff, R.H.; Fruttiger, M.; Richardson, W.D.; Franklin, R.J. Platelet-derived growth factor regulates oligodendrocyte progenitor numbers in adult CNS and their response following CNS demyelination. Mol. Cell. Neurosci. 2004, 25, 252–262. [Google Scholar] [CrossRef]
- Selvaraju, R.; Bernasconi, L.; Losberger, C.; Graber, P.; Kadi, L.; Avellana-Adalid, V.; Picard-Riera, N.; Baron Van Evercooren, A.; Cirillo, R.; Kosco-Vilbois, M.; et al. Osteopontin is upregulated during in vivo demyelination and remyelination and enhances myelin formation in vitro. Mol. Cell. Neurosci. 2004, 25, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Stidworthy, M.F.; Genoud, S.; Li, W.W.; Leone, D.P.; Mantei, N.; Suter, U.; Franklin, R.J. Notch1 and Jagged1 are expressed after CNS demyelination, but are not a major rate-determining factor during remyelination. Brain 2004, 127 Pt 9, 1928–1941. [Google Scholar] [CrossRef] [PubMed]
- Zatta, P.; Raso, M.; Zambenedetti, P.; Wittkowski, W.; Messori, L.; Piccioli, F.; Mauri, P.L.; Beltramini, M. Copper and zinc dismetabolism in the mouse brain upon chronic cuprizone treatment. Cell. Mol. Life Sci. CMLS 2005, 62, 1502–1513. [Google Scholar] [CrossRef] [PubMed]
- Irvine, K.A.; Blakemore, W.F. Age increases axon loss associated with primary demyelination in cuprizone-induced demyelination in C57BL/6 mice. J. Neuroimmunol. 2006, 175, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.T.; Martin, C.B.; Ingersoll, S.A.; Barnum, S.R.; Martin, B.K. Astrocyte-specific expression of a soluble form of the murine complement control protein Crry confers demyelination protection in the cuprizone model. Glia 2007, 55, 1405–1415. [Google Scholar] [CrossRef]
- Song, S.Y.; Kato, C.; Adachi, E.; Moriya-Sato, A.; Inagawa-Ogashiwa, M.; Umeda, R.; Hashimoto, N. Expression of an acyl-CoA synthetase, lipidosin, in astrocytes of the murine brain and its up-regulation during remyelination following cuprizone-induced demyelination. J. Neurosci. Res. 2007, 85, 3586–3597. [Google Scholar] [CrossRef]
- Biancotti, J.C.; Kumar, S.; de Vellis, J. Activation of inflammatory response by a combination of growth factors in cuprizone-induced demyelinated brain leads to myelin repair. Neurochem. Res. 2008, 33, 2615–2628. [Google Scholar] [CrossRef]
- Merkler, D.; Klinker, F.; Jürgens, T.; Glaser, R.; Paulus, W.; Brinkmann, B.G.; Sereda, M.W.; Stadelmann-Nessler, C.; Guedes, R.C.; Brück, W.; et al. Propagation of spreading depression inversely correlates with cortical myelin content. Ann. Neurol. 2009, 66, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Doucette, J.R.; Jiao, R.; Nazarali, A.J. Age-related and cuprizone-induced changes in myelin and transcription factor gene expression and in oligodendrocyte cell densities in the rostral corpus callosum of mice. Cell. Mol. Neurobiol. 2010, 30, 607–629. [Google Scholar] [CrossRef] [PubMed]
- Baertling, F.; Kokozidou, M.; Pufe, T.; Clarner, T.; Windoffer, R.; Wruck, C.J.; Brandenburg, L.O.; Beyer, C.; Kipp, M. ADAM12 is expressed by astrocytes during experimental demyelination. Brain Res. 2010, 1326, 1–14. [Google Scholar] [CrossRef]
- Cate, H.S.; Sabo, J.K.; Merlo, D.; Kemper, D.; Aumann, T.D.; Robinson, J.; Merson, T.D.; Emery, B.; Perreau, V.M.; Kilpatrick, T.J. Modulation of bone morphogenic protein signalling alters numbers of astrocytes and oligodendroglia in the subventricular zone during cuprizone-induced demyelination. J. Neurochem. 2010, 115, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.R.; McCandless, E.E.; Dorsey, D.; Klein, R.S. CXCR4 promotes differentiation of oligodendrocyte progenitors and remyelination. Proc. Natl. Acad. Sci. USA 2010, 107, 11062–11067. [Google Scholar] [CrossRef]
- Parenti, R.; Cicirata, F.; Zappalà, A.; Catania, A.; La Delia, F.; Cicirata, V.; Tress, O.; Willecke, K. Dynamic expression of Cx47 in mouse brain development and in the cuprizone model of myelin plasticity. Glia 2010, 58, 1594–1609. [Google Scholar] [CrossRef]
- Clarner, T.; Buschmann, J.P.; Beyer, C.; Kipp, M. Glial amyloid precursor protein expression is restricted to astrocytes in an experimental toxic model of multiple sclerosis. J. Mol. Neurosci. 2011, 43, 268–274. [Google Scholar] [CrossRef]
- Ingersoll, S.A.; Martin, C.B.; Barnum, S.R.; Martin, B.K. CNS-specific expression of C3a and C5a exacerbate demyelination severity in the cuprizone model. Mol. Immunol. 2010, 48, 219–230. [Google Scholar] [CrossRef]
- Crocker, S.J.; Bajpai, R.; Moore, C.S.; Frausto, R.F.; Brown, G.D.; Pagarigan, R.R.; Whitton, J.L.; Terskikh, A.V. Intravenous administration of human embryonic stem cell-derived neural precursor cells attenuates cuprizone-induced central nervous system (CNS) demyelination. Neuropathol. Appl. Neurobiol. 2011, 37, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, S.V.; O’Keeffe, G.W.; Sullivan, A.M. BMP-Smad 1/5/8 signalling in the development of the nervous system. Prog. Neurobiol. 2013, 109, 28–41. [Google Scholar] [CrossRef]
- Palumbo, S.; Toscano, C.D.; Parente, L.; Weigert, R.; Bosetti, F. Time-dependent changes in the brain arachidonic acid cascade during cuprizone-induced demyelination and remyelination. Prostaglandins Leukot. Essent. Fat. Acids 2011, 85, 29–35. [Google Scholar] [CrossRef]
- Hoyos, H.C.; Marder, M.; Ulrich, R.; Gudi, V.; Stangel, M.; Rabinovich, G.A.; Pasquini, L.A.; Pasquini, J.M. The Role of Galectin-3: From Oligodendroglial Differentiation and Myelination to Demyelination and Remyelination Processes in a Cuprizone-Induced Demyelination Model. Adv. Exp. Med. Biol. 2016, 949, 311–332. [Google Scholar] [CrossRef]
- Kim, S.; Steelman, A.J.; Zhang, Y.; Kinney, H.C.; Li, J. Aberrant upregulation of astroglial ceramide potentiates oligodendrocyte injury. Brain Pathol. 2012, 22, 41–57. [Google Scholar] [CrossRef]
- Kipp, M.; Gingele, S.; Pott, F.; Clarner, T.; van der Valk, P.; Denecke, B.; Gan, L.; Siffrin, V.; Zipp, F.; Dreher, W.; et al. BLBP-expression in astrocytes during experimental demyelination and in human multiple sclerosis lesions. Brain Behav. Immun. 2011, 25, 1554–1568. [Google Scholar] [CrossRef]
- Gudi, V.; Škuljec, J.; Yildiz, Ö.; Frichert, K.; Skripuletz, T.; Moharregh-Khiabani, D.; Voss, E.; Wissel, K.; Wolter, S.; Stangel, M. Spatial and temporal profiles of growth factor expression during CNS demyelination reveal the dynamics of repair priming. PLoS ONE 2011, 6, e22623. [Google Scholar] [CrossRef] [PubMed]
- Skuljec, J.; Gudi, V.; Ulrich, R.; Frichert, K.; Yildiz, O.; Pul, R.; Voss, E.V.; Wissel, K.; Baumgärtner, W.; Stangel, M. Matrix metalloproteinases and their tissue inhibitors in cuprizone-induced demyelination and remyelination of brain white and gray matter. J. Neuropathol. Exp. Neurol. 2011, 70, 758–769. [Google Scholar] [CrossRef]
- Silvestroff, L.; Bartucci, S.; Pasquini, J.; Franco, P. Cuprizone-induced demyelination in the rat cerebral cortex and thyroid hormone effects on cortical remyelination. Exp. Neurol. 2012, 235, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Liu, L.; Spangler, R.; Spear, C.; Wang, C.; Gulen, M.F.; Veenstra, M.; Ouyang, W.; Ransohoff, R.M.; Li, X. IL-17-induced Act1-mediated signaling is critical for cuprizone-induced demyelination. J. Neurosci. 2012, 32, 8284–8292. [Google Scholar] [CrossRef]
- Brück, W.; Pförtner, R.; Pham, T.; Zhang, J.; Hayardeny, L.; Piryatinsky, V.; Hanisch, U.K.; Regen, T.; van Rossum, D.; Brakelmann, L.; et al. Reduced astrocytic NF-κB activation by laquinimod protects from cuprizone-induced demyelination. Acta Neuropathol. 2012, 124, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Sabo, J.K.; Aumann, T.D.; Kilpatrick, T.J.; Cate, H.S. Investigation of sequential growth factor delivery during cuprizone challenge in mice aimed to enhance oligodendrogliogenesis and myelin repair. PLoS ONE 2013, 8, e63415. [Google Scholar] [CrossRef]
- Azami Tameh, A.; Clarner, T.; Beyer, C.; Atlasi, M.A.; Hassanzadeh, G.; Naderian, H. Regional regulation of glutamate signaling during cuprizone-induced demyelination in the brain. Ann. Anat. 2013, 195, 415–423. [Google Scholar] [CrossRef]
- Kanekiyo, K.; Inamori, K.; Kitazume, S.; Sato, K.; Maeda, J.; Higuchi, M.; Kizuka, Y.; Korekane, H.; Matsuo, I.; Honke, K.; et al. Loss of branched O-mannosyl glycans in astrocytes accelerates remyelination. J. Neurosci. 2013, 33, 10037–10047. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, T.; Tamura, M.; Kondo, M.A.; Sakaue, M.; Okada, K.; Takemoto, K.; Fukunari, A.; Miwa, K.; Ohzeki, H.; Kano, S.; et al. Cuprizone short-term exposure: Astrocytic IL-6 activation and behavioral changes relevant to psychosis. Neurobiol. Dis. 2013, 59, 63–68. [Google Scholar] [CrossRef]
- Janssens, K.; Maheshwari, A.; Van den Haute, C.; Baekelandt, V.; Stinissen, P.; Hendriks, J.J.; Slaets, H.; Hellings, N. Oncostatin M protects against demyelination by inducing a protective microglial phenotype. Glia 2015, 63, 1729–1737. [Google Scholar] [CrossRef]
- Okazaki, R.; Doi, T.; Hayakawa, K.; Morioka, K.; Imamura, O.; Takishima, K.; Hamanoue, M.; Sawada, Y.; Nagao, M.; Tanaka, S.; et al. The crucial role of Erk2 in demyelinating inflammation in the central nervous system. J. Neuroinflammation 2016, 13, 235. [Google Scholar] [CrossRef] [PubMed]
- Petković, F.; Campbell, I.L.; Gonzalez, B.; Castellano, B. Astrocyte-targeted production of interleukin-6 reduces astroglial and microglial activation in the cuprizone demyelination model: Implications for myelin clearance and oligodendrocyte maturation. Glia 2016, 64, 2104–2119. [Google Scholar] [CrossRef] [PubMed]
- Draheim, T.; Liessem, A.; Scheld, M.; Wilms, F.; Weißflog, M.; Denecke, B.; Kensler, T.W.; Zendedel, A.; Beyer, C.; Kipp, M.; et al. Activation of the astrocytic Nrf2/ARE system ameliorates the formation of demyelinating lesions in a multiple sclerosis animal model. Glia 2016, 64, 2219–2230. [Google Scholar] [CrossRef]
- Zimmermann, J.; Emrich, M.; Krauthausen, M.; Saxe, S.; Nitsch, L.; Heneka, M.T.; Campbell, I.L.; Müller, M. IL-17A Promotes Granulocyte Infiltration, Myelin Loss, Microglia Activation, and Behavioral Deficits During Cuprizone-Induced Demyelination. Mol. Neurobiol. 2018, 55, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Espitia Pinzon, N.; Sanz-Morello, B.; Brevé, J.J.; Bol, J.G.; Drukarch, B.; Bauer, J.; Baron, W.; van Dam, A.M. Astrocyte-derived tissue Transglutaminase affects fibronectin deposition, but not aggregation, during cuprizone-induced demyelination. Sci. Rep. 2017, 7, 40995. [Google Scholar] [CrossRef]
- Petković, F.; Campbell, I.L.; Gonzalez, B.; Castellano, B. Reduced cuprizone-induced cerebellar demyelination in mice with astrocyte-targeted production of IL-6 is associated with chronically activated, but less responsive microglia. J. Neuroimmunol. 2017, 310, 97–102. [Google Scholar] [CrossRef]
- Kim, S.; Bielawski, J.; Yang, H.; Kong, Y.; Zhou, B.; Li, J. Functional antagonism of sphingosine-1-phosphate receptor 1 prevents cuprizone-induced demyelination. Glia 2018, 66, 654–669. [Google Scholar] [CrossRef]
- Sanadgol, N.; Golab, F.; Mostafaie, A.; Mehdizadeh, M.; Khalseh, R.; Mahmoudi, M.; Abdollahi, M.; Vakilzadeh, G.; Taghizadeh, G.; Sharifzadeh, M. Low, but not high, dose triptolide controls neuroinflammation and improves behavioral deficits in toxic model of multiple sclerosis by dampening of NF-κB activation and acceleration of intrinsic myelin repair. Toxicol. Appl. Pharmacol. 2018, 342, 86–98. [Google Scholar] [CrossRef]
- Sanchez, M.A.; Sullivan, G.M.; Armstrong, R.C. Genetic detection of Sonic hedgehog (Shh) expression and cellular response in the progression of acute through chronic demyelination and remyelination. Neurobiol. Dis. 2018, 115, 145–156. [Google Scholar] [CrossRef]
- Omotoso, G.O.; Ukwubile, I.I.; Arietarhire, L.; Sulaimon, F.; Gbadamosi, I.T. Kolaviron protects the brain in cuprizone-induced model of experimental multiple sclerosis via enhancement of intrinsic antioxidant mechanisms: Possible therapeutic applications? Pathophysiology 2018, 25, 299–306. [Google Scholar] [CrossRef]
- Jakovac, H.; Grubić Kezele, T.; Radošević-Stašić, B. Expression Profiles of Metallothionein I/II and Megalin in Cuprizone Model of De- and Remyelination. Neuroscience 2018, 388, 69–86. [Google Scholar] [CrossRef]
- Mokhtarzadeh Khanghahi, A.; Satarian, L.; Deng, W.; Baharvand, H.; Javan, M. In vivo conversion of astrocytes into oligodendrocyte lineage cells with transcription factor Sox10; Promise for myelin repair in multiple sclerosis. PLoS ONE 2018, 13, e0203785. [Google Scholar] [CrossRef]
- Zare, L.; Baharvand, H.; Javan, M. In vivo conversion of astrocytes to oligodendrocyte lineage cells using chemicals: Targeting gliosis for myelin repair. Regen. Med. 2018, 13, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Farhangi, S.; Dehghan, S.; Totonchi, M.; Javan, M. In vivo conversion of astrocytes to oligodendrocyte lineage cells in adult mice demyelinated brains by Sox2. Mult. Scler. Relat. Disord. 2019, 28, 263–272. [Google Scholar] [CrossRef]
- Carden, T.R.; Correale, J.; Pasquini, J.M.; Pérez, M.J. Transferrin Enhances Microglial Phagocytic Capacity. Mol. Neurobiol. 2019, 56, 6324–6340. [Google Scholar] [CrossRef]
- Kramann, N.; Menken, L.; Pförtner, R.; Schmid, S.N.; Stadelmann, C.; Wegner, C.; Brück, W. Glial fibrillary acidic protein expression alters astrocytic chemokine release and protects mice from cuprizone-induced demyelination. Glia 2019, 67, 1308–1319. [Google Scholar] [CrossRef]
- Barati, S.; Kashani, I.R.; Tahmasebi, F.; Mehrabi, S.; Joghataei, M.T. Effect of mesenchymal stem cells on glial cells population in cuprizone induced demyelination model. Neuropeptides 2019, 75, 75–84. [Google Scholar] [CrossRef]
- Roboon, J.; Hattori, T.; Ishii, H.; Takarada-Iemata, M.; Le, T.M.; Shiraishi, Y.; Ozaki, N.; Yamamoto, Y.; Sugawara, A.; Okamoto, H.; et al. Deletion of CD38 Suppresses Glial Activation and Neuroinflammation in a Mouse Model of Demyelination. Front. Cell. Neurosci. 2019, 13, 258. [Google Scholar] [CrossRef]
- Li, Q.Y.; Miao, Q.; Sui, R.X.; Cao, L.; Ma, C.G.; Xiao, B.G.; Xiao, W.; Yu, W.B. Ginkgolide K supports remyelination via induction of astrocytic IGF/PI3K/Nrf2 axis. Int. Immunopharmacol. 2019, 75, 105819. [Google Scholar] [CrossRef]
- An, J.; Yin, J.J.; He, Y.; Sui, R.X.; Miao, Q.; Wang, Q.; Yu, J.Z.; Yu, J.W.; Shi, F.D.; Ma, C.G.; et al. Temporal and Spatial Dynamics of Astroglial Reaction and Immune Response in Cuprizone-Induced Demyelination. Neurotox. Res. 2020, 37, 587–601. [Google Scholar] [CrossRef]
- Kriszta, G.; Nemes, B.; Sándor, Z.; Ács, P.; Komoly, S.; Berente, Z.; Bölcskei, K.; Pintér, E. Investigation of Cuprizone-Induced Demyelination in mGFAP-Driven Conditional Transient Receptor Potential Ankyrin 1 (TRPA1) Receptor Knockout Mice. Cells 2019, 9, 81. [Google Scholar] [CrossRef]
- Yoon, H.; Choi, C.I.; Triplet, E.M.; Langley, M.R.; Kleppe, L.S.; Kim, H.N.; Simon, W.L.; Scarisbrick, I.A. Blocking the Thrombin Receptor Promotes Repair of Demyelinated Lesions in the Adult Brain. J. Neurosci. 2020, 40, 1483–1500. [Google Scholar] [CrossRef]
- Yin, J.J.; He, Y.; An, J.; Miao, Q.; Sui, R.X.; Wang, Q.; Yu, J.Z.; Xiao, B.G.; Ma, C.G. Dynamic Balance of Microglia and Astrocytes Involved in the Remyelinating Effect of Ginkgolide B. Front. Cell. Neurosci. 2019, 13, 572. [Google Scholar] [CrossRef]
- Zamora, N.N.; Cheli, V.T.; Santiago González, D.A.; Wan, R.; Paez, P.M. Deletion of Voltage-Gated Calcium Channels in Astrocytes during Demyelination Reduces Brain Inflammation and Promotes Myelin Regeneration in Mice. J. Neurosci. 2020, 40, 3332–3347. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Chico, A.; Manterola, A.; Cipriani, R.; Katona, I.; Matute, C.; Mato, S. P2x7 receptors control demyelination and inflammation in the cuprizone model. Brain Behav. Immun. Health 2020, 4, 100062. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.M.; Knutsen, A.K.; Peruzzotti-Jametti, L.; Korotcov, A.; Bosomtwi, A.; Dardzinski, B.J.; Bernstock, J.D.; Rizzi, S.; Edenhofer, F.; Pluchino, S.; et al. Transplantation of induced neural stem cells (iNSCs) into chronically demyelinated corpus callosum ameliorates motor deficits. Acta Neuropathol. Commun. 2020, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Reinbach, C.; Stadler, M.S.; Pröbstl, N.; Chrzanowski, U.; Schmitz, C.; Kipp, M.; Hochstrasser, T. CD44 expression in the cuprizone model. Brain Res. 2020, 1745, 146950. [Google Scholar] [CrossRef]
- Luo, M.; Deng, M.; Yu, Z.; Zhang, Y.; Xu, S.; Hu, S.; Xu, H. Differential Susceptibility and Vulnerability of Brain Cells in C57BL/6 Mouse to Mitochondrial Dysfunction Induced by Short-Term Cuprizone Exposure. Front. Neuroanat. 2020, 14, 30. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, N.; Zhang, R.; Jin, L.; Petridis, A.K.; Loers, G.; Zheng, X.; Wang, Z.; Siebert, H.C. Cuprizone-Induced Demyelination in Mouse Hippocampus Is Alleviated by Ketogenic Diet. J. Agric. Food Chem. 2020, 68, 11215–11228. [Google Scholar] [CrossRef]
- Tahmasebi, F.; Pasbakhsh, P.; Barati, S.; Madadi, S.; Kashani, I.R. The effect of microglial ablation and mesenchymal stem cell transplantation on a cuprizone-induced demyelination model. J. Cell. Physiol. 2021, 236, 3552–3564. [Google Scholar] [CrossRef]
- Rohr, S.O.; Greiner, T.; Joost, S.; Amor, S.; Valk, P.V.; Schmitz, C.; Kipp, M. Aquaporin-4 Expression during Toxic and Autoimmune Demyelination. Cells 2020, 9, 2187. [Google Scholar] [CrossRef]
- He, Y.; An, J.; Yin, J.J.; Miao, Q.; Sui, R.X.; Han, Q.X.; Ding, Z.B.; Huang, J.J.; Ma, C.G.; Xiao, B.G. Ethyl Pyruvate-Derived Transdifferentiation of Astrocytes to Oligodendrogenesis in Cuprizone-Induced Demyelinating Model. Neurotherapeutics 2021, 18, 488–502. [Google Scholar] [CrossRef]
- Espírito-Santo, S.; Coutinho, V.G.; Dezonne, R.S.; Stipursky, J.; Dos Santos-Rodrigues, A.; Batista, C.; Paes-de-Carvalho, R.; Fuss, B.; Gomes, F.C.A. Astrocytes as a target for Nogo-A and implications for synapse formation in vitro and in a model of acute demyelination. Glia 2021, 69, 1429–1443. [Google Scholar] [CrossRef]
- Marzan, D.E.; Brügger-Verdon, V.; West, B.L.; Liddelow, S.; Samanta, J.; Salzer, J.L. Activated microglia drive demyelination via CSF1R signaling. Glia 2021, 69, 1583–1604. [Google Scholar] [CrossRef]
- Baltan, S.; Jawaid, S.S.; Chomyk, A.M.; Kidd, G.J.; Chen, J.; Battapady, H.D.; Chan, R.; Dutta, R.; Trapp, B.D. Neuronal hibernation following hippocampal demyelination. Acta Neuropathol. Commun. 2021, 9, 34. [Google Scholar] [CrossRef]
- Saitta, K.S.; Lercher, L.D.; Sainato, D.M.; Patel, A.; Huang, Y.; McAuliffe, G.; Dreyfus, C.F. CHPG enhances BDNF and myelination in cuprizone-treated mice through astrocytic metabotropic glutamate receptor 5. Glia 2021, 69, 1950–1965. [Google Scholar] [CrossRef] [PubMed]
- Klejbor, I.; Shimshek, D.R.; Klimaszewska-Łata, J.; Velasco-Estevez, M.; Moryś, J.; Karaszewski, B.; Szutowicz, A.; Rutkowska, A. EBI2 is expressed in glial cells in multiple sclerosis lesions, and its knock-out modulates remyelination in the cuprizone model. Eur. J. Neurosci. 2021, 54, 5173–5188. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.; Triolo, D.; Bassani, C.; Bedogni, F.; Di Dario, M.; Dina, G.; Fredrickx, E.; Fermo, I.; Martinelli, V.; Newcombe, J.; et al. Dysregulated copper transport in multiple sclerosis may cause demyelination via astrocytes. Proc. Natl. Acad. Sci. USA 2021, 118, e2025804118. [Google Scholar] [CrossRef] [PubMed]
- Cheli, V.T.; Santiago González, D.A.; Wan, Q.; Denaroso, G.; Wan, R.; Rosenblum, S.L.; Paez, P.M. H-ferritin expression in astrocytes is necessary for proper oligodendrocyte development and myelination. Glia 2021, 69, 2981–2998. [Google Scholar] [CrossRef]
- An, J.; He, Y.; Yin, J.J.; Ding, Z.B.; Han, Q.X.; Chen, Y.Y.; Wang, Q.; Chai, Z.; Yu, J.Z.; Song, L.J.; et al. Temporal and spatial evolution of various functional neurons during demyelination induced by cuprizone. J. Neurophysiol. 2021, 126, 1756–1771. [Google Scholar] [CrossRef]
- Castillo-Rodriguez, M.L.A.; Gingele, S.; Schröder, L.J.; Möllenkamp, T.; Stangel, M.; Skripuletz, T.; Gudi, V. Astroglial and oligodendroglial markers in the cuprizone animal model for de- and remyelination. Histochem. Cell Biol. 2022, 158, 15–38. [Google Scholar] [CrossRef]
- Kawabe, Y.; Tanaka, T.; Isonishi, A.; Nakahara, K.; Tatsumi, K.; Wanaka, A. Characterization of Glial Populations in the Aging and Remyelinating Mouse Corpus Callosum. Neurochem. Res. 2022, 47, 2826–2838. [Google Scholar] [CrossRef]
- Silva Oliveira Junior, M.; Schira-Heinen, J.; Reiche, L.; Han, S.; de Amorim, V.C.M.; Lewen, I.; Gruchot, J.; Göttle, P.; Akkermann, R.; Azim, K.; et al. Myelin repair is fostered by the corticosteroid medrysone specifically acting on astroglial subpopulations. EBioMedicine 2022, 83, 104204. [Google Scholar] [CrossRef]
- Zha, Z.; Liu, Y.J.; Liu, S.S.; Zhang, N.; Li, J.L.; Qi, F.; Jin, L.Y.; Xue, B.; Yang, T.; Fan, Y.P.; et al. Bu Shen Yi Sui Capsule Promotes Myelin Repair by Modulating the Transformation of A1/A2 Reactive Astrocytes In Vivo and In Vitro. Oxidative Med. Cell. Longev. 2022, 2022, 3800004. [Google Scholar] [CrossRef] [PubMed]
- Madadi, S.; Shiri, E.; Pasbakhsh, P.; Tahmasebi, F.; Kazemzadeh, S.; Zibara, K.; Kashani, I.R. Combination Therapy of Mesenchymal Stem Cell Transplantation and Astrocyte Ablation Improve Remyelination in a Cuprizone-Induced Demyelination Mouse Model. Mol. Neurobiol. 2022, 59, 7278–7292. [Google Scholar] [CrossRef]
- Fallier-Becker, P.; Bonzheim, I.; Pfeiffer, F. Cuprizone feeding induces swollen astrocyte endfeet. Pflug. Arch. 2022, 474, 1275–1283. [Google Scholar] [CrossRef]
- Sun, W.; Wen, M.; Liu, M.; Wang, Q.; Liu, Q.; Li, L.; Siebert, H.C.; Loers, G.; Zhang, R.; Zhang, N. Effect of β-hydroxybutyrate on behavioral alterations, molecular and morphological changes in CNS of multiple sclerosis mouse model. Front. Aging Neurosci. 2022, 14, 1075161. [Google Scholar] [CrossRef]
- Lei, C.; Chen, H.; Chen, K. Effects of Bone Marrow Mesenchymal Stem Cells on Myelin Repair and Emotional Changes of a Cuprizone-Induced Demyelination Model. J. Integr. Neurosci. 2023, 22, 40. [Google Scholar] [CrossRef]
- Takahashi, K.; Kanekiyo, K.; Sakuda, K.; Muto, Y.; Iguchi, M.; Matsuda, N.; Hashimoto, Y.; Kanai, K.; Ogawa, H.; Hirase, H.; et al. Brain-specific glycosylation of protein tyrosine phosphatase receptor type Z (PTPRZ) marks a demyelination-associated astrocyte subtype. J. Neurochem. 2023, 166, 547–559. [Google Scholar] [CrossRef]
- Frankle, L.; Riley, A.; Tomor, R.; Lee, H.; Jarzembak, K.; Benedict, O.; Sternbach, S.; Shelestak, J.; McDonough, J.; Clements, R. Changes to Astrocyte-associated Protein Expression at Different Timepoints of Cuprizone Treatment. BioRxiv 2023. [Google Scholar] [CrossRef]
- Mohamadi, Y.; Borhani-Haghighi, M. TGN020 application against aquaporin 4 improved multiple sclerosis by inhibiting astrocytes, microglia, and NLRP3 inflammasome in a cuprizone mouse model. J. Chem. Neuroanat. 2023, 132, 102306. [Google Scholar] [CrossRef]
Figure 1.
The proliferation of different glia subpopulations during the course of cuprizone-induced demyelination [45]. The curved line in the graph illustrates the myelination levels of the midline of the corpus callosum during acute demyelination (week 5) and the subsequent remyelination phase as the animals are provided normal chow after acute demyelination. The dotted line illustrates the myelination levels of the midline of the corpus callosum if the intoxication with cuprizone is continued until week 13 (chronic demyelination). The straight lines beneath the graph illustrate the periods of glia cell proliferation. The inserts show cortical astrocytes, visualized using anti-GFAP immunohistochemistry, in control (left image) and 5-week-cuprizone-intoxicated mice (right image). The inserted bars are 50 µm.
Figure 1.
The proliferation of different glia subpopulations during the course of cuprizone-induced demyelination [45]. The curved line in the graph illustrates the myelination levels of the midline of the corpus callosum during acute demyelination (week 5) and the subsequent remyelination phase as the animals are provided normal chow after acute demyelination. The dotted line illustrates the myelination levels of the midline of the corpus callosum if the intoxication with cuprizone is continued until week 13 (chronic demyelination). The straight lines beneath the graph illustrate the periods of glia cell proliferation. The inserts show cortical astrocytes, visualized using anti-GFAP immunohistochemistry, in control (left image) and 5-week-cuprizone-intoxicated mice (right image). The inserted bars are 50 µm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kipp, M. Astrocytes: Lessons Learned from the Cuprizone Model. Int. J. Mol. Sci. 2023, 24, 16420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216420
AMA Style
Kipp M. Astrocytes: Lessons Learned from the Cuprizone Model. International Journal of Molecular Sciences. 2023; 24(22):16420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216420
Chicago/Turabian StyleKipp, Markus. 2023. "Astrocytes: Lessons Learned from the Cuprizone Model" International Journal of Molecular Sciences 24, no. 22: 16420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216420
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.