
Disinhibition Is an Essential Network Motif Coordinated by GABA Levels and GABA B Receptors

1
Academia de Fisiología, Escuela Superior de Medicina, Instituto Politécnico Nacional, Plan de San Luis y Díaz Mirón, Colonia Casco de Santo Tomás, Ciudad de México 11340, Mexico
2
Sección de Estudios Posgrado e Investigación de la Escuela Superior de Medicina, Instituto Politécnico Nacional, Plan de San Luis y Díaz Mirón, Colonia Casco de Santo Tomás, Ciudad de Mexico 11340, Mexico
Int. J. Mol. Sci. 2024, 25(2), 1340; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25021340
Submission received: 12 December 2023
/
Revised: 16 January 2024
/
Accepted: 19 January 2024
/
Published: 22 January 2024
(This article belongs to the Collection Feature Papers in Molecular Neurobiology)
{kind=link}
{kind=link}
Abstract
:Network dynamics are crucial for action and sensation. Changes in synaptic physiology lead to the reorganization of local microcircuits. Consequently, the functional state of the network impacts the output signal depending on the firing patterns of its units. Networks exhibit steady states in which neurons show various activities, producing many networks with diverse properties. Transitions between network states determine the output signal generated and its functional results. The temporal dynamics of excitation/inhibition allow a shift between states in an operational network. Therefore, a process capable of modulating the dynamics of excitation/inhibition may be functionally important. This process is known as disinhibition. In this review, we describe the effect of GABA levels and GABAB receptors on tonic inhibition, which causes changes (due to disinhibition) in network dynamics, leading to synchronous functional oscillations.
1. Introduction
The neural network is the substrate of brain function. The functional dynamics of neural networks modify the synaptic physiology, leading to the reorganization of these circuits [1]. These modifications modulate the firing of local units that determine the network’s functional state [2]. In the network’s functional state, neurons respond to both convergent and divergent inputs (from the network itself, recurrent local networks, or even distal networks), which can be excitatory or inhibitory. These dynamics effectively influence the output signals by modulating selective connections, leading to constant changes in the network pathways [3]. Thus, the balance between excitation and inhibition (E/I) is a fundamental parameter that influences the network’s output signal [4]. An imbalance in this parameter impacts the progression of various diseases [5,6].
Networks have a stable state in which they are not entirely inactive. In this state, the neuronal activity (including synaptic strength, intrinsic properties, and E/I balance) produces a broad spectrum of networks with various properties (size, dynamics, and linkage frameworks [7]). In an operational network, the temporal evolution between E/I is proportional among individual neurons and across the global network. This feature allows rapid transitions between states that translate into functional output signals [3]. Therefore, a process able to modulate the network state by controlling the temporal dynamics between E/I is functionally essential. This process is known as disinhibition. Defined as a selective and transient hindrance to inhibition that leads to excitation [8], disinhibition is an evolutionarily conserved process involved in various motor [9,10] and cognitive functional processes, brain regions, neuronal types, and projection neuron compartments [8,11]. A crucial physiological property is its duration, which can range from milliseconds to days [8,12].
Another essential signal modulation mechanism is gain control, which involves regulating the amplitude caused by the E/I balance [13]. In this framework, inhibition in the perisomatic region controls the gain of projection neuron responses, which modulates the arrangement of synaptic inputs [8]. In the network, gain control induces normalization of the average neuron firing rate according to the inputs [8,14,15]; therefore, inhibition adjusts the gain. Consequently, a regenerative depolarization that begins in the dendrites is the process by which distal excitatory inputs lead to neural activation; this process has been described in relation to disinhibition [16].
Neural network oscillations are involved in multiple physiological processes and are accepted as tools for communication among brain regions. Thus, oscillations lead to the organization and coordination of information due to the precise interactions among the activities of different neurons over time. The emergence of oscillations and their frequency ranges depend on both the intrinsic neuronal properties and the network properties [2,17].
Gamma-aminobutyric acid (GABA) is a canonical inhibitory neurotransmitter. GABA performs its function through three types of receptors: GABAA, GABAB, and GABAC. The versatility of GABA is demonstrated not only in the diversity of receptors but also in how it establishes inhibition, with phasic and tonic forms. In the phasic form, the postsynaptic GABAA receptor is activated by an increase in GABA in the synaptic cleft after release. In the tonic form, GABA overflows from the synaptic cleft and activates extrasynaptic GABA A receptors (located in the presynaptic terminal and synapses with adjacent neurons), allowing temporally and spatially slow transmission. The extrasynaptic receptors show a high affinity to GABA and mainly constitute the δ subunits assembled at α4 or α6 [18,19,20,21]. The importance of extrasynaptic GABAA receptors (GABAA Rs) to the tonic current is widely accepted [20] and evidence of the involvement of GABAB receptors (GABAB Rs) as modulators of this current has been presented [22].
According to the above information, in the present review, we describe results that suggest that both GABA levels and GABAB Rs influence disinhibition, which modulates the gain mode of the output signal and thus changes the dynamics of the network (observed as changes in oscillations), thus allowing a functional state.
2. Involvement of GABA B Receptors
GABAB Rs are ubiquitous metabotropic receptors in the brain. These receptors couple with the Gi protein. Consequently, the Gαi/Gαo subunits inhibit adenylyl cyclase (AC), which reduces cAMP levels and thus decreases protein kinase A (PKA) activity [21]. The Gβγ subunits inhibit the Ca2+ channels and activate the GIRK and TREK-type K+ channels [23]. The affinity of the Gβγ subunit to GIRKs increases due to the Na+ that enters during the action potential; this increase in affinity leads to the opening of the channel [24].
GABAB Rs are expressed at both inhibitory and excitatory synapses. They require two different subunits for their function: GABAB1 and GABAB2. GABA binds to the GABAB1 subunit and the GABAB2 subunit causes signaling [25]. Due to gene splicing, GABAB1 has two isoforms, GABAB1a and GABAB1b, which have different N-N-terminal regions. This region determines that the GABA B1a isoform is expressed at the presynaptic terminal in excitatory synapses, thus modulating glutamate release [25,26,27]. However, both isoforms are expressed at the postsynaptic terminal. The GABAB1b subunit in this terminal allows coupling with the K+ channels, reducing this ion’s current [28]. Moreover, this subunit inhibits dendritic Ca2+ spikes by affecting voltage Ca2+ channel (VGCC) sensitivity [28]. This molecular diversity leads to heterogeneous responses that dynamically modulate synaptic transmission and, therefore, the network.
2.1. Presynaptic Modulation
The critical presynaptic function of GABAB Rs is inhibiting the release of neurotransmitters by restricting the entry of Ca2+ into the terminal, followed by the inhibition of VGCC by the Gβγ subunit. However, this is not the only mechanism of GABAB Rs. GABAB Rs also modulate the release mechanism at various levels. For example, they modulate the SNAP-25 protein by decreasing cAMP levels, which reduces vesicular priming. During this process, the Gβγ subunit modulates neurotransmitter release through direct interaction with the SNAP-25 protein, and this interaction is modulated by synaptotagmin-1 [29]. Furthermore, neurexins have been reported to enable GABAB Rs to function at this terminal [30]. Therefore, the activation of presynaptic GABAB Rs by reducing GABA release leads to the formation of a negative feedback loop, which is typical of autoreceptor-mediated synaptic gain control [31].
GABAB Rs exert gain modulation at the network level through their presynaptic control. In olfactory inputs, they control differential presynaptic gain [32]. In the presynaptic neurons of the inferior colliculus, GABAB Rs control the excitability gain, and blockading them increases adaptation to the stimulus [33]. In the auditory pathway, they normalize the activity of the neuronal population by controlling the gain [34]. Presynaptic GABAB Rs change the E/I ratio of the neurons in the prefrontal cortex, thereby modulating network excitation in a model of autism [35].
2.2. Postsynaptic Modulation
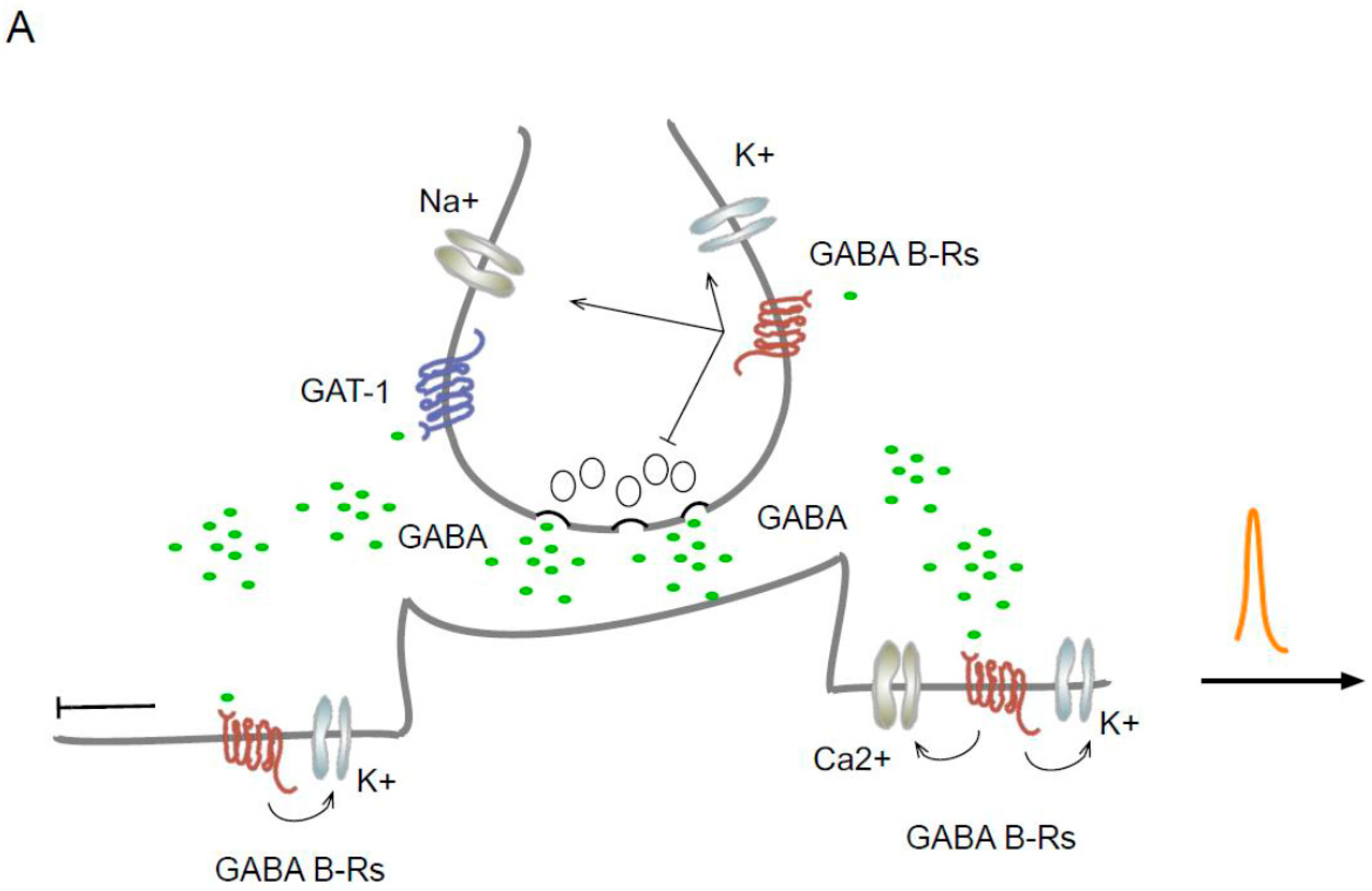
The predominant effect at the postsynaptic level is the inhibition mediated by the activation of the GIRK channels by the Gβγ subunit [36]. Once active, the Gβγ subunit forms a complex with the RGS protein, which binds to the GIRKs and accelerates their kinetics [37]. At the same time, the RGS protein increases the GTPase activity of the Gα subunit, causing rapid desensitization of the K+ current [38], which reduces neuronal excitability and inhibits action potential backpropagation. Furthermore, GABAB Rs activate a two-pore domain K+ channel (TREK2, also called KCNK10 [39,40]). In the dendrites, the joint activation of GIRK, TREK2, and VGCC and the inhibition of action potential backpropagation [41] reduce the generation of Ca2+ spikes (Figure 1A).
2.3. Modulation of Network Dynamics
GABAB Rs modulate the dynamics in the network by decreasing the output current. Once activated, GABAB Rs inhibit the N-type calcium and BK-type potassium channels. Consequently, the neurons increase their degree of depolarization, incrementing their excitation level [23]. It has been suggested that inhibiting the N-type channels leads to operational advantages by expanding the transmission dynamics without influencing neurotransmitter release [23]. In the lateral geniculate nucleus, the activation of GABAB Rs leads to strong hyperpolarization, followed by rebound firing as mediated by the T-type Ca2+ channels, which improves the signal detectability while altering sensory discrimination [42]. GABAB R activation in the network is related to burst firing and constant rhythmic activity. In this way, GABAB Rs may temporally modulate slow network activity, the strength of fast activity, and the relative firing during network oscillations [43].
3. GABA Levels
Recent evidence suggests that the synaptic levels of GABA are essential to synaptic transmission and network physiology. Quantitative reports suggest that the synaptic GABA levels are in the mM range, while the extrasynaptic levels are in the μM range. However, in the nuclei of specific neurons, these levels are higher at rest and during physiological processes [36]. Consequently, it is currently accepted that wiring-based (synaptic) and volume-based (non-synaptic) transmissions both occur in the brain [44,45]. This context supports the proposition that the molecular versatility of GABA has functional implications for the network physiology due to its effect on different neuronal compartments. In this way, GABA levels modulate the output signals and considerably influence the network dynamics.
Accepted functional roles of the GABA levels in both synaptic transmission and the network dynamics include the generation of tonic current and the activation of extrasynaptic GABAA and GABAB Rs (including subtypes of both receptors sensitive to low GABA concentrations [46]). This current has been found in multiple synaptic compartments, including the presynaptic terminals (inhibitory and excitatory), the perisynaptic regions of the excitatory synapses, and the middle of unmyelinated axons and dendrites [45].
It is evident that GABA (by controlling the generation of membrane potential with different forms in various regions) modulates the temporal integration of synaptic inputs and, consequently, the activity patterns of the neuronal populations that make up the circuits, with the tonic current playing a fundamental role. Thus, the stimulating current is modulated by different processes, including the sustained activation of individual cells, the coordination of presynaptic events of different interneurons, an increase in the current density at the release site, and the reuptake mechanisms, which depend on the transporters.
GABA transporters (GATs) are widely expressed in multiple nuclei, compartments, and neuronal glia, with a higher expression in axons than dendrites [47]. The function of GATs depends on several factors. First, the amplitude of the receptor activation is transiently modulated by the GABA concentration. Second, GATs control the receptor kinetics during the recruitment of neighboring synapses. Third, GATs are involved in spatial transmission; they convert spatially confined inhibitory signals into waves without spatial restrictions, which can activate type A or B receptors at both the presynaptic and postsynaptic terminals [46,48]. Thus, GATs functionally participate in the modulation of tonic currents. They reduce the conductance of neuronal inputs, temporarily modulating the incoming excitatory outputs. Therefore, tonic GABA currents control the input–output relationship (neuronal stimulation versus response) through shunt inhibition [49] along the dendrite or axon.
The contribution of GABA Rs to tonic current has been described. Studies on neurons in the locus ceruleus suggest that GABAB Rs generate tonic currents through ERK1-dependent activation [50]. In the basal amygdala, an increase in tonic inhibition related to presynaptic regulation was reported [51]. Interestingly, increases in the tonic current due to the postsynaptic activation of GABAB Rs have been reported in thalamocortical neuron granule cells in both the cerebellum and the dentate gyrus. In the dentate gyrus, the increase in tonic current was related to an increase in membrane traffic and the expression of high-affinity extrasynaptic GABAA Rs induced by GABAB R activation (Figure 1A) [51].
Hefty evidence in the last decades shows the physiological role of glial cells in synaptic transmission, especially astrocytes [52,53]. Astrocytes transform glutamatergic excitation into GABAergic inhibition [54] and contribute to establishing negative feedback [55,56]. During synaptic transmission, astrocytes absorb glutamate, which releases GABA. The GABA released establishes tonic inhibition depending on the network activity and modulates the E/I balance [57].
4. Disinhibition
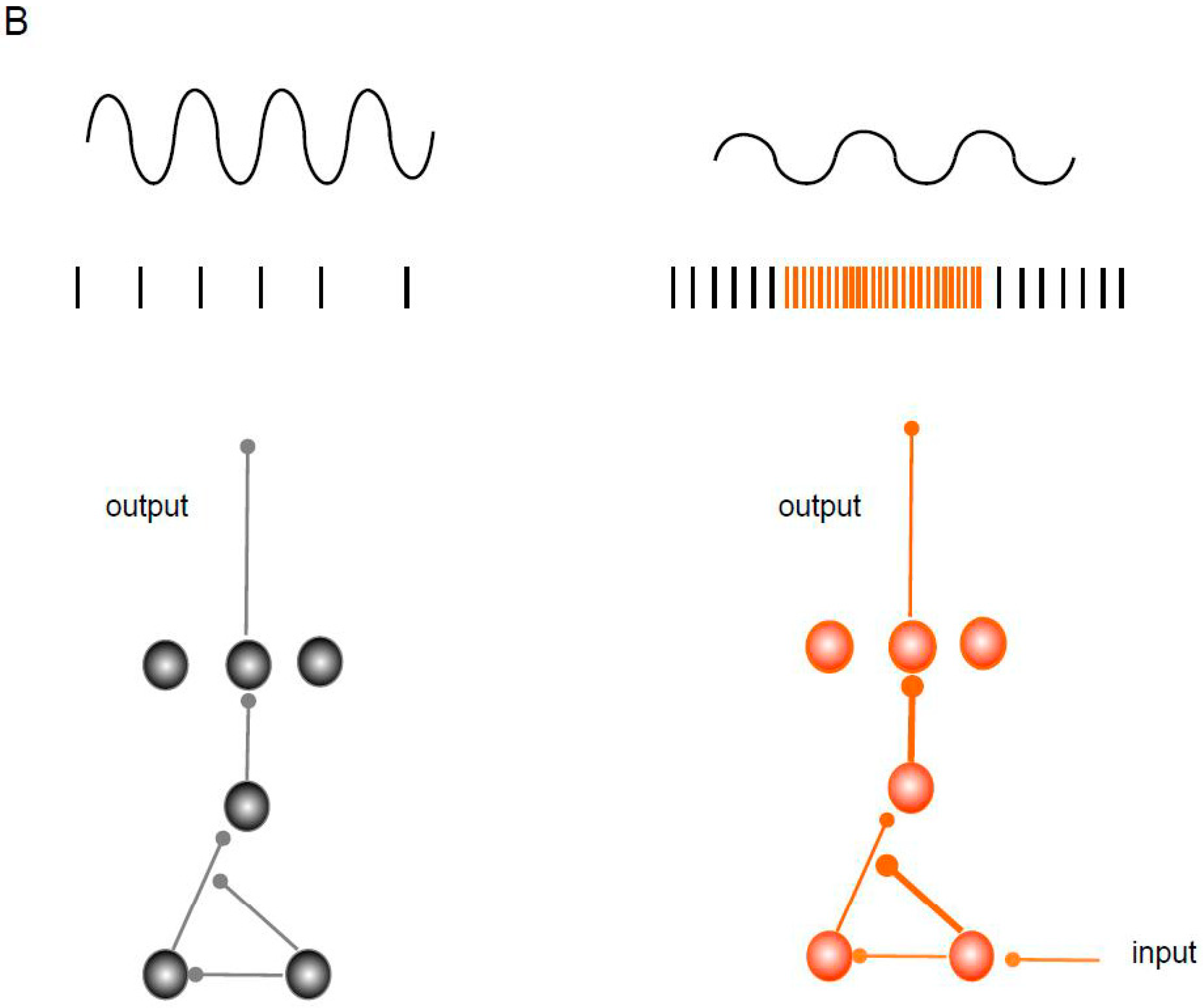
Functionally, a microcircuit may have a “disinhibiting” motif, including serial connections between two inhibitory interneurons and a principal excitatory neuron (Figure 1B) [58]. These circuits are involved in the response to social fear. In this process, the increase in the activity of somatostatin (SST)-positive interneurons (INs) inhibits parvalbumin-positive (PV+) INs, which causes disinhibition of the principal cells of the dorsomedial prefrontal cortex [59]. Similarly, an exaggerated response to fear is mediated by a disinhibitory circuit involving the dorsal raphe nucleus, pericentral reticulotegmental nucleus, and central reticulotegmental nucleus [60].
An interesting disinhibitory process was described in the lateral entorhinal cortex. In it, optogenetic silencing of VIP INs (positive for vasoactive intestinal peptide) significantly decreased the incidence of the dendritic spikes driven by the lateral entorhinal cortex, suggesting a disinhibitory effect being exerted on the dendritic activity by the INs [61]. A disinhibitory process at the dendritic level was described in the hippocampus associated with the LTP process [62]. The VIP INs in the hippocampus regulate LTP through disinhibition by activating the VPAC1 receptors (G protein-coupled receptors of the VIP/PACAP family). This molecular mechanism affects the expression and phosphorylation of the Kv4.2 K+ channels in the dendrites of the hippocampal pyramidal cells [11].
During attentional selection, selective disinhibition improved the target firing rates with resemblance to multiplicative input gain, another commonly reported effect of attention on neural responses [63]. In the cerebellum, the activation of GABAB Rs in the dentate gyrus (DG) improves the granule cell (GC) activity by reducing the synaptic inhibition imposed by hilar INs. This disinhibitory action facilitates the transfer of signals from the hippocampus. Furthermore, the GABAB(1a,2) and GABAB(1b,2) R subtypes differentially modulate the GC output through dendritic locations and axon terminals. The disinhibition described during spatial and pattern learning in the hippocampus is mediated exclusively by GABAB(1a,2) Rs. A disinhibitory circuit of the distal dendrites of pyramidal cells mediates motor integration in the somatosensory cortex [64]. Furthermore, it was recently reported that both GAT-1 blockade and GABAB agonism disinhibited the neurons in the reticular thalamic nucleus (RTn) [65,66].
Several research results support the functional importance of inhibition in neuronal circuits [67,68]. Therefore, it is accepted that inhibition stabilizes the circuit, thus allowing calculations to be carried out that include network stability, response normalization, and input amplification through a process called inhibitory plasticity [67,68,69]. This process modulates both the generation and firing frequency properties of the excitatory neurons and the input–output function. In the circuit, excitation is propagated by rapid changes in input, the downregulation of inhibitory gains, or temporary changes in the E/I balance. In this way, it establishes coordinated communication between circuits. Thus, disinhibition by increasing the firing frequency allows the transfer of information between circuits. This mechanism has been proposed as the basis of association memory. Through E/I modulation, disinhibition allows the recovery of memories, which are kept in a resting state [68]. Studies that analyze the involvement of the GABA levels, as well as the participation of GABAB Rs, are on the rise.
In addition to various physiological processes, disinhibition has been proposed as a pathophysiological substrate of various diseases. In Parkinson’s disease, disinhibition in the primary motor cortex has been suggested as one of the first manifestations [70] and is correlated with freezing and gait impairment [71]. Obesity is associated with disinhibition of the orbitofrontal cortex. It has been shown that excitability is secondary to the reduction in the tonic inhibition of the pyramidal neurons and the excitability decreases with increasing GABA levels [72].
Tourette’s syndrome (TS) is a hyperkinetic disorder characterized by motor and phonetic tics [73,74]. However, its clinical presentation is commonly accompanied by various abnormal behaviors (motor, sensory, and complex behavioral), such as inappropriate non-obscene behaviors, impulsivity, obsessive-compulsive disorder, and attention deficit hyperactivity disorder [74] The central pathophysiological substrate is disinhibition [10,73,74]. This originates from alterations in the GABAergic signaling and a loss of E/I balance in the cortex–basal ganglia–thalamus–cortex circuit associated with the modulation of motor output, motor learning, and action selection. In TS, alterations in the E/I balance leads to a reduction in the gain in motor cortical excitability. In this context, increasing GABA levels in the somatosensory cortex alter the gain of excitability (by increasing tonic inhibition), which improves the control of motor outputs [73].
The disinhibitory circuit is ubiquitous: its presence is not limited to the cerebral cortex and it communicates with subcortical structures. It has an important implication in regulating muscle tone, which allows motor control [75]. Regarding GABAB, it is suggested that these receptors are extrasynaptic and are tonically activated by the GABA released by astrocytes as well as by the GABAergic interneurons in the spinal motoneurons and descending afferent fibers [76]. A transcendental role of disinhibition has been described in the pathophysiology of pain, especially chronic pain [77,78]. In its development, a loss of inhibition as mediated by parvolabumin-positive interneurons has been suggested, as well as simultaneously the postsynaptic inhibition of the vertical cells. These events allow the pain circuits of lamina I to be activated by innocuous tactile stimuli [77].
5. Oscillation
Various compelling studies have described the modulatory participation of GABAB Rs in slow cortical oscillations. During slow wave oscillations (SWOs), GABAB R blockade modifies three important aspects of the SWO cycle. First, GABAB R blockade increases the number of “up” states; second, GABAB R blockade affects the subsequent duration of the “down” state; and third, GABAB R activation desynchronizes the SWOs [79]. The participation of GABAB Rs according to their synaptic location has also been described. At the presynaptic level, GABAB Rs contribute to spontaneous transitions from the down state, while postsynaptic receptors are essential for the afferent termination of the up state. Thus, GABAB Rs containing the GABA subunit contribute to spontaneous termination of the up state, and GABAB Rs containing the GABAB1b subunit are essential for afferent evoked termination of the upstate [80]. Furthermore, GABAB(1a,2) heteroreceptors selectively located at the thalamocortical relay (TCR)–RTn cell synapses in the thalamus (Th) regulate the oscillation strength, while GABAB(1b,2) Rs control the oscillation frequency [81].
Thalamic oscillations of 3 to 5 Hz are characteristic of absence epilepsy. In a rodent model, selectively blocking the GABA transporters ([GAT] to GAT-1 or GAT-3) was shown to prolong the oscillations; however, blocking both transporters inhibited the oscillations. In this same model, it was reported that extending the activity in a narrow range of GABAB Rs promoted the opening of T-type channels and intensified the oscillations [82].
In addition, tonic inhibition and disinhibition have been suggested to regulate motor activity [83]. An analysis of the cortical oscillations after GAT-1 blockade demonstrated that high GABA levels influence the beta oscillations related to movement [84]. Moreover, it was recently reported that high GABA levels and the participation of GABAB Rs in the external globus pallidus disinhibit the RTn and thus desynchronize the beta oscillations in the motor cortex [65,66]. In the human visual cortex, the GABA levels modulate the gamma oscillations. Increased GABA levels in this region (after GAT-I blockade) lead to decreased gamma oscillations [85]. In the hippocampus, activation of the GABAB Rs selectively suppresses the recruitment of SST Ins during gamma oscillations induced in vitro (Figure 1B) [86].
6. Discussion
The neurophysiological aspects addressed in this paper form an essential part of the physiology of the neuronal dynamics. Thus, the data described firmly support the participation of both the GABA levels and GABAB Rs in neuronal dynamics.
The recent results regarding cortical disinhibition are undeniable [87]. However, the current data regarding cortical disinhibition focus on cortical vision, leaving aside the subcortical structures. Thus, it is widely accepted that the subcortical structures part of the interneuronal network are exclusively inhibitory. In this organization, the data described suggest that convergent afferent synaptic activity can alter the precise temporal arrangement of the network activity through disinhibition. Furthermore, the flow of such information toward a functional neuronal network is highly regulated by synaptic inhibition as mediated by the GABAB Rs [88], contributing to the disinhibition process.
Thus, to establish communication between circuits, the phase of the incoming message must be the disinhibitory phase of the oscillation of the receiving circuit [3]. Therefore, in network dynamics, disinhibition could be a mechanism that opens the door to sending information. The current results support the proposition that the exit gate leads to the dendrites, as described in the entorhinal cortex (EC). This region contains a long-range disinhibitory motif that facilitates the integration of inputs from the EC into the hippocampus, increasing the probability of dendritic spiking while inducing the input’s time-dependent plasticity [89].
In network dynamics, the dendrites process information from multiple inputs; simultaneously, they can segregate them into various regions [90]. To generate dendritic spikes, various events must occur, such as intense and repetitive activity, integration with other coincident inputs, and inhibition blockage. Thus, it has been proposed that a disinhibitory circuit participates as a gain modulation mechanism that allows dendritic nonlinearities in a rapid, dynamic, and path-specific manner. Furthermore, it increases the dendritic excitation further when combined with long-range disinhibitory stimuli. During oscillatory synchrony between the EC and the hippocampus, high-amplitude dendritic spikes that do not produce action potentials in the soma are observed [90]. An excitatory process that integrates the GABAA and B receptors was proposed several years ago [91]. It was suggested that in thin dendritic processes, massive activation of the GABAA Rs would lead to depolarizing values. At the same time, the K+ inversion potential would be altered, which would produce inefficiency in the activity of the GABAB Rs. This inactivity would indicate that the equilibrium potential of K+ moves toward the resting potential, increasing the PEPS amplitude without causing hyperpolarization. Consequently, GABAB-type inhibition in large distal dendrites silences the GABAA-mediated inhibition [91].
In microcircuits, information processing between inhibitory internal neurons that modulate inhibitory and excitatory operations is carried out according to lateral inhibition processes. One such process involves the RTn–Th–cortical (TC) circuit. In this circuit, the axonal fields of the RTn neurons and the dendritic domains of the TCs overlap. Based on this morphological characteristic, it has been suggested that disinhibitory feedback loops form due to disinhibition, increasing the contrast between the central and peripheral neurons generated by lateral inhibition [92]. Thus, both processes (lateral inhibition and disinhibition) can establish feedback loops, leading to communication between circuits.
In conclusion, GABA Rs and GABA levels both contribute to the formation of exit gates related to network communication in the cortex and subcortical regions, and these processes modulate motor and cognitive events at various levels.
Funding
This work was supported by funding from Sección de Estudios de Posgrado e Investigación del Instituto Politécnico Nacional to Nelson Villalobos [grant nos. 20211436 and 20231668].
Conflicts of Interest
The author declares no conflicts of interest.
References
- Avena-Koenigsberger, A.; Misic, B.; Sporns, O. Communication dynamics in complex brain networks. Nat. Rev. Neurosci. 2017, 19, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Mejia, M.T.; Villalobos, N. Neurophysiology of Brain Networks Underlies Symptoms of Parkinson’s Disease: A Basis for Diagnosis and Management. Diagnostics 2023, 13, 2394. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.; Ponce-Alvarez, A.; Deco, G.; Aertsen, A.; Kumar, A. Portraits of communication in neuronal networks. Nat. Rev. Neurosci. 2019, 20, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Sun, Q.Q. Circuit-specific and neuronal subcellular-wide E-I balance in cortical pyramidal cells. Sci. Rep. 2018, 8, 3971. [Google Scholar] [CrossRef] [PubMed]
- Sohal, V.S.; Rubenstein, J.L.R. Excitation-inhibition balance as a framework for investigating mechanisms in neuropsychiatric disorders. Mol. Psychiatry 2019, 24, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Eichler, S.A.; Meier, J.C. E-I balance and human diseases—From molecules to networking. Front. Mol. Neurosci. 2008, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Barral, J.; DReyes, A. Synaptic scaling rule preserves excitatory-inhibitory balance and salient neuronal network dynamics. Nat. Neurosci. 2016, 19, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Letzkus, J.J.; Wolff, S.B.; Lüthi, A. Disinhibition, a Circuit Mechanism for Associative Learning and Memory. Neuron 2015, 88, 264–276. [Google Scholar] [CrossRef]
- Chevalier, G.; Deniau, J.M. Disinhibition as a basic process in the expression of striatal functions. Trends Neurosci. 1990, 13, 277–280. [Google Scholar] [CrossRef]
- Ganos, C.; Rothwell, J.; Haggard, P. Voluntary inhibitory motor control over involuntary tic movements. Mov. Disord. 2018, 33, 937–946. [Google Scholar] [CrossRef]
- Artinian, J.; Lacaille, J.C. Disinhibition in learning and memory circuits: New vistas for somatostatin interneurons and long-term synaptic plasticity. Brain Res. Bull. 2018, 141, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Leblois, A.; Bodor, A.L.; Person, A.L.; Perkel, D.J. Millisecond timescale disinhibition mediates fast information transmission through an avian basal ganglia loop. J. Neurosci. 2009, 29, 15420–15433. [Google Scholar] [CrossRef] [PubMed]
- Kremkow, J.; Aertsen, A.; Kumar, A. Gating of signal propagation in spiking neural networks by balanced and correlated excitation and inhibition. J. Neurosci. 2010, 30, 15760–15768. [Google Scholar] [CrossRef] [PubMed]
- Salinas, E.; Thier, P. Gain modulation: A major computational principle of the central nervous system. Neuron 2000, 27, 15–21. [Google Scholar] [CrossRef]
- Louie, K.; Khaw, M.W.; Glimcher, P.W. Normalization is a general neural mechanism for context-dependent decision making. Proc. Natl. Acad. Sci. USA 2013, 110, 6139–6144. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.R.; Murray, J.D.; Wang, X.J. A dendritic disinhibitory circuit mechanism for pathway-specific gating. Nat. Commun. 2016, 7, 12815. [Google Scholar] [CrossRef] [PubMed]
- Singer, W. Neuronal oscillations: Unavoidable and useful? Eur. J. Neurosci. 2018, 48, 2389–2398. [Google Scholar] [CrossRef]
- Farrant, M.; Nusser, Z. Variations on an inhibitory theme: Phasic and tonic activation of GABA(A) receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef]
- Farrant, M.; Kaila, K. The cellular, molecular and ionic basis of GABA(A) receptor signalling. Prog. Brain Res. 2007, 160, 59–87. [Google Scholar] [CrossRef]
- Kasaragod, V.B.; Mortensen, M.; Hardwick, S.W.; Wahid, A.A.; Dorovykh, V.; Chirgadze, D.Y.; Smart, T.G.; Miller, P.S. Mechanisms of inhibition and activation of extrasynaptic αβ GABAA receptors. Nature 2022, 602, 529–533. [Google Scholar] [CrossRef]
- Evenseth, L.S.M.; Gabrielsen, M.; Sylte, I. The GABAB Receptor-Structure, Ligand Binding and Drug Development. Molecules 2020, 25, 3093. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Tao, W.; Yang, L.; Spain, W.J.; Ransom, C.B. GABA-B receptors enhance GABA-A receptor currents by modulation of membrane trafficking in dentate gyrus granule cells. Neurosci. Lett. 2022, 773, 136481. [Google Scholar] [CrossRef]
- Garaycochea, J.; Slaughter, M.M. GABAB receptors enhance excitatory responses in isolated rat retinal ganglion cells. J. Physiol. 2016, 594, 5543–5554. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Touhara, K.K.; Weir, K.; Bean, B.P.; MacKinnon, R. Cooperative regulation by G proteins and Na+ of neuronal GIRK2 K+ channels. eLife 2016, 5, e15751. [Google Scholar] [CrossRef]
- Chalifoux, J.R.; Carter, A.G. GABAB receptor modulation of synaptic function. Curr. Opin. Neurobiol. 2011, 21, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Frangaj, A.; Fan, Q.R. Structural biology of GABAB receptor. Neuropharmacology 2018, 136 Pt A, 68–79. [Google Scholar] [CrossRef]
- Shaye, H.; Stauch, B.; Gati, C.; Cherezov, V. Molecular mechanisms of metabotropic GABAB receptor function. Sci. Adv. 2021, 7, eabg3362. [Google Scholar] [CrossRef]
- Luján, R.; Aguado, C.; Ciruela, F.; Cózar, J.; Kleindienst, D.; de la Ossa, L.; Bettler, B.; Wickman, K.; Watanabe, M.; Shigemoto, R.; et al. Differential association of GABAB receptors with their effector ion channels in Purkinje cells. Brain Struct. Funct. 2018, 223, 1565–1587. [Google Scholar] [CrossRef]
- Alten, B.; Guzikowski, N.J.; Zurawski, Z.; Hamm, H.E.; Kavalali, E.T. Presynaptic mechanisms underlying GABAB-receptor-mediated inhibition of spontaneous neurotransmitter release. Cell Rep. 2022, 38, 110255. [Google Scholar] [CrossRef]
- Luo, F.; Sclip, A.; Merrill, S.; Südhof, T.C. Neurexins regulate presynaptic GABAB receptors at central synapses. Nat. Commun. 2021, 12, 2380. [Google Scholar] [CrossRef]
- Roth, F.C.; Draguhn, A. GABA metabolism and transport: Effects on synaptic efficacy. Neural Plast. 2012, 2012, 805830. [Google Scholar] [CrossRef] [PubMed]
- Root, C.M.; Masuyama, K.; Green, D.S.; Enell, L.E.; Nässel, D.R.; Lee, C.H.; Wang, J.W. A presynaptic gain control mechanism fine-tunes olfactory behavior. Neuron 2008, 59, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Ayala, Y.A.; Malmierca, M.S. The effect of inhibition on stimulus-specific adaptation in the inferior colliculus. Brain Struct. Funct. 2018, 223, 1391–1407. [Google Scholar] [CrossRef] [PubMed]
- Stange, A.; Myoga, M.H.; Lingner, A.; Ford, M.C.; Alexandrova, O.; Felmy, F.; Pecka, M.; Siveke, I.; Grothe, B. Adaptation in sound localization: From GABA(B) receptor-mediated synaptic modulation to perception. Nat. Neurosci. 2013, 16, 1840–1847. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, D.; Luhmann, H.J.; Kirischuk, S. Presynaptic GABAB receptor-mediated network excitation in the medial prefrontal cortex of Tsc2+/− mice. Pflug. Arch. Eur. J. Physiol. 2021, 473, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.C.; Chu, Y.L.; Tsai, M.L.; Wong, S.B.; Min, M.Y.; Chen, R.F.; Yang, H.W. GABAB receptor-mediated tonic inhibition of locus coeruleus neurons plays a role in deep anesthesia induced by isoflurane. Neuroreport 2020, 31, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Marron Fernandez de Velasco, E.; Wickman, K. Neuronal G protein-gated K+ channels. Am. J. Physiol. Cell Physiol. 2022, 323, C439–C460. [Google Scholar] [CrossRef]
- Turecek, R.; Schwenk, J.; Fritzius, T.; Ivankova, K.; Zolles, G.; Adelfinger, L.; Jacquier, V.; Besseyrias, V.; Gassmann, M.; Schulte, U.; et al. Auxiliary GABAB receptor subunits uncouple G protein βγ subunits from effector channels to induce desensitization. Neuron 2014, 82, 1032–1044. [Google Scholar] [CrossRef]
- Deng, P.Y.; Xiao, Z.; Yang, C.; Rojanathammanee, L.; Grisanti, L.; Watt, J.; Geiger, J.D.; Liu, R.; Porter, J.E.; Lei, S. GABA(B) receptor activation inhibits neuronal excitability and spatial learning in the entorhinal cortex by activating TREK-2 K+ channels. Neuron 2009, 63, 230–243. [Google Scholar] [CrossRef]
- Chang, Y.T.; Ling, J.; Gu, J.G. Effects of GABAB receptor activation on excitability of IB4-positive maxillary trigeminal ganglion neurons: Possible involvement of TREK2 activation. Mol. Pain 2021, 17, 17448069211042963. [Google Scholar] [CrossRef]
- Leung, L.S.; Peloquin, P. GABA(B) receptors inhibit backpropagating dendritic spikes in hippocampal CA1 pyramidal cells in vivo. Hippocampus 2006, 16, 388–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Jin, Y.; Sun, H.; Ma, C.; Zhang, J.; Wang, M.; Chen, L. Characterization of Rebound Depolarization in Neurons of the Rat Medial Geniculate Body In Vitro. Neurosci. Bull. 2016, 32, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Kohl, M.M.; Paulsen, O. The roles of GABAB receptors in cortical network activity. Adv. Pharmacol. 2010, 58, 205–229. [Google Scholar] [CrossRef] [PubMed]
- Oláh, S.; Füle, M.; Komlósi, G.; Varga, C.; Báldi, R.; Barzó, P.; Tamás, G. Regulation of cortical microcircuits by unitary GABA-mediated volume transmission. Nature 2009, 461, 1278–1281. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.; Kwak, H.; Cheong, E.; Lee, C.J. GABA tone regulation and its cognitive functions in the brain. Nat. Rev. Neuroscience 2023, 24, 523–539. [Google Scholar] [CrossRef]
- Khatri, S.N.; Wu, W.C.; Yang, Y.; Pugh, J.R. Mechanisms of GABAB receptor enhancement of extrasynaptic GABAA receptor currents in cerebellar granule cells. Sci. Rep. 2019, 9, 16683. [Google Scholar] [CrossRef]
- Melone, M.; Ciappelloni, S.; Conti, F. A quantitative analysis of cellular and synaptic localization of GAT-1 and GAT-3 in rat neocortex. Brain Struct. Funct. 2015, 220, 885–897. [Google Scholar] [CrossRef]
- Scimemi, A. Structure, function, and plasticity of GABA transporters. Front. Cell. Neurosci. 2014, 8, 161. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Silver, R.A. Shunting inhibition modulates neuronal gain during synaptic excitation. Neuron 2003, 38, 433–445. [Google Scholar] [CrossRef]
- Wu, R.N.; Kuo, C.C.; Min, M.Y.; Chen, R.F.; Yang, H.W. Extracellular Signal-Regulated Kinases Mediate an Autoregulation of GABAB-Receptor-Activated Whole-Cell Current in Locus Coeruleus Neurons. Sci. Rep. 2020, 10, 7869. [Google Scholar] [CrossRef]
- Meis, S.; Endres, T.; Munsch, T.; Lessmann, V. Presynaptic Regulation of Tonic Inhibition by Neuromodulatory Transmitters in the Basal Amygdala. Mol. Neurobiol. 2018, 55, 8509–8521. [Google Scholar] [CrossRef] [PubMed]
- Durkee, C.A.; Araque, A. Diversity and Specificity of Astrocyte-neuron Communication. Neuroscience 2019, 396, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Arizono, M.; Nägerl, U.V. Deciphering the functional nano-anatomy of the tripartite synapse using stimulated emission depletion microscopy. Glia 2022, 70, 607–618. [Google Scholar] [CrossRef]
- Héja, L.; Nyitrai, G.; Kékesi, O.; Dobolyi, A.; Szabó, P.; Fiáth, R.; Ulbert, I.; Pál-Szenthe, B.; Palkovits, M.; Kardos, J. Astrocytes convert network excitation to tonic inhibition of neurons. BMC Biol. 2012, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Héja, L.; Simon, Á.; Szabó, Z.; Kardos, J. Feedback adaptation of synaptic excitability via Glu:Na+ symport driven astrocytic GABA and Gln release. Neuropharmacology 2019, 161, 107629. [Google Scholar] [CrossRef] [PubMed]
- Szabó, Z.; Péter, M.; Héja, L.; Kardos, J. Dual Role for Astroglial Copper-Assisted Polyamine Metabolism during Intense Network Activity. Biomolecules 2021, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.; Cho, W.H.; Lee, B.H.; Kim, D.W.; Kim, Y.S.; Park, K.; Hwang, M.; Barcelon, E.; Cho, Y.K.; Lee, C.J.; et al. Cortical astrocytes modulate dominance behavior in male mice by regulating synaptic excitatory and inhibitory balance. Nat. Neurosci. 2023, 26, 1541–1554. [Google Scholar] [CrossRef]
- Chen, Q.; Smith, R.G.; Huang, X.; Wei, W. Preserving inhibition with a disinhibitory microcircuit in the retina. eLife 2020, 9, e62618. [Google Scholar] [CrossRef]
- Xu, H.; Liu, L.; Tian, Y.; Wang, J.; Li, J.; Zheng, J.; Zhao, H.; He, M.; Xu, T.L.; Duan, S.; et al. A Disinhibitory Microcircuit Mediates Conditioned Social Fear in the Prefrontal Cortex. Neuron 2019, 102, 668–682.e5. [Google Scholar] [CrossRef]
- Guo, W.; Fan, S.; Xiao, D.; He, C.; Guan, M.; Xiong, W. A midbrain-reticulotegmental circuit underlies exaggerated startle under fear emotions. Mol. Psychiatry 2022, 27, 4881–4892. [Google Scholar] [CrossRef]
- Bilash, O.M.; Chavlis, S.; Johnson, C.D.; Poirazi, P.; Basu, J. Lateral entorhinal cortex inputs modulate hippocampal dendritic excitability by recruiting a local disinhibitory microcircuit. Cell Rep. 2023, 42, 111962. [Google Scholar] [CrossRef] [PubMed]
- Caulino-Rocha, A.; Rodrigues, N.C.; Ribeiro, J.A.; Cunha-Reis, D. Endogenous VIP VPAC1 Receptor Activation Modulates Hippocampal Theta Burst Induced LTP: Transduction Pathways and GABAergic Mechanisms. Biology 2022, 11, 627. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, D.; Knudsen, E.I. Selective disinhibition: A unified neural mechanism for predictive and post hoc attentional selection. Vis. Res. 2015, 116 Pt B, 194–209. [Google Scholar] [CrossRef]
- Lee, S.; Kruglikov, I.; Huang, Z.J.; Fishell, G.; Rudy, B. A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nat. Neurosci. 2013, 16, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, N.; Almazán-Alvarado, S.; Magdaleno-Madrigal, V.M. Elevation of GABA levels in the globus pallidus disinhibits the thalamic reticular nucleus and desynchronized cortical beta oscillations. J. Physiol. Sci. 2022, 72, 17. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, N.; Magdaleno-Madrigal, V.M. Pallidal GABA B receptors: Involvement in cortex beta dynamics and thalamic reticular nucleus activity. J. Physiol. Sci. 2023, 73, 14. [Google Scholar] [CrossRef] [PubMed]
- Hennequin, G.; Agnes, E.J.; Vogels, T.P. Inhibitory Plasticity: Balance, Control, and Codependence. Annu. Rev. Neurosci. 2017, 40, 557–579. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.K.; Miehl, C.; Gjorgjieva, J. Regulation of circuit organization and function through inhibitory synaptic plasticity. Trends Neurosci. 2022, 45, 884–898. [Google Scholar] [CrossRef]
- Hernández-Frausto, M.; Bilash, O.M.; Masurkar, A.V.; Basu, J. Local and long-range GABAergic circuits in hippocampal area CA1 and their link to Alzheimer’s disease. Front. Neural Circuits 2023, 17, 1223891. [Google Scholar] [CrossRef]
- Ammann, C.; Dileone, M.; Pagge, C.; Catanzaro, V.; Mata-Marín, D.; Hernández-Fernández, F.; Monje, M.H.G.; Sánchez-Ferro, Á.; Fernández-Rodríguez, B.; Gasca-Salas, C.; et al. Cortical disinhibition in Parkinson’s disease. Brain 2020, 143, 3408–3421. [Google Scholar] [CrossRef]
- Sun, H.; Gan, C.; Wang, L.; Ji, M.; Cao, X.; Yuan, Y.; Zhang, H.; Shan, A.; Gao, M.; Zhang, K. Cortical Disinhibition Drives Freezing of Gait in Parkinson’s Disease and an Exploratory Repetitive Transcranial Magnetic Stimulation Study. Mov. Disord. 2023, 38, 2072–2083. [Google Scholar] [CrossRef] [PubMed]
- Seabrook, L.T.; Naef, L.; Baimel, C.; Judge, A.K.; Kenney, T.; Ellis, M.; Tayyab, T.; Armstrong, M.; Qiao, M.; Floresco, S.B.; et al. Disinhibition of the orbitofrontal cortex biases decision-making in obesity. Nat. Neurosci. 2023, 26, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Jackson, G.M.; Draper, A.; Dyke, K.; Pépés, S.E.; Jackson, S.R. Inhibition, Disinhibition, and the Control of Action in Tourette Syndrome. Trends Cogn. Sci. 2015, 19, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Kurvits, L.; Martino, D.; Ganos, C. Clinical Features That Evoke the Concept of Disinhibition in Tourette Syndrome. Front. Psychiatry 2020, 11, 21. [Google Scholar] [CrossRef]
- Ganguly, J.; Kulshreshtha, D.; Almotiri, M.; Jog, M. Muscle Tone Physiology and Abnormalities. Toxins 2021, 13, 282. [Google Scholar] [CrossRef]
- Delgado-Ramírez, X.; Alvarado-Cervantes, N.S.; Jiménez-Barrios, N.; Raya-Tafolla, G.; Felix, R.; Martínez-Rojas, V.A.; Delgado-Lezama, R. GABAB Receptors Tonically Inhibit Motoneurons and Neurotransmitter Release from Descending and Primary Afferent Fibers. Life 2023, 13, 1776. [Google Scholar] [CrossRef]
- Hughes, D.I.; Todd, A.J. Central Nervous System Targets: Inhibitory Interneurons in the Spinal Cord. Neurotherapeutics 2020, 17, 874–885. [Google Scholar] [CrossRef]
- Cioffi, C.L. Inhibition of Glycine Re-Uptake: A Potential Approach for Treating Pain by Augmenting Glycine-Mediated Spinal Neurotransmission and Blunting Central Nociceptive Signaling. Biomolecules 2021, 11, 864. [Google Scholar] [CrossRef]
- Perez-Zabalza, M.; Reig, R.; Manrique, J.; Jercog, D.; Winograd, M.; Parga, N.; Sanchez-Vives, M.V. Modulation of cortical slow oscillatory rhythm by GABAB receptors: An in vitro experimental and computational study. J. Physiol. 2020, 598, 3439–3457. [Google Scholar] [CrossRef]
- Craig, M.T.; Mayne, E.W.; Bettler, B.; Paulsen, O.; McBain, C.J. Distinct roles of GABAB1a- and GABAB1b-containing GABAB receptors in spontaneous and evoked termination of persistent cortical activity. J. Physiol. 2013, 591, 835–843. [Google Scholar] [CrossRef]
- Ulrich, D.; Lalanne, T.; Gassmann, M.; Bettler, B. GABAB receptor subtypes differentially regulate thalamic spindle oscillations. Neuropharmacology 2018, 136 Pt A, 106–116. [Google Scholar] [CrossRef]
- Lu, A.C.; Lee, C.K.; Kleiman-Weiner, M.; Truong, B.; Wang, M.; Huguenard, J.R.; Beenhakker, M.P. Nonlinearities between inhibition and T-type calcium channel activity bidirectionally regulate thalamic oscillations. eLife 2020, 9, e59548. [Google Scholar] [CrossRef]
- Benjamin, P.R.; Staras, K.; Kemenes, G. What roles do tonic inhibition and disinhibition play in the control of motor programs? Front. Behav. Neurosci. 2010, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Muthukumaraswamy, S.D.; Myers, J.F.; Wilson, S.J.; Nutt, D.J.; Lingford-Hughes, A.; Singh, K.D.; Hamandi, K. The effects of elevated endogenous GABA levels on movement-related network oscillations. NeuroImage 2013, 66, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Magazzini, L.; Muthukumaraswamy, S.D.; Campbell, A.E.; Hamandi, K.; Lingford-Hughes, A.; Myers, J.F.; Nutt, D.J.; Sumner, P.; Wilson, S.J.; Singh, K.D. Significant reductions in human visual gamma frequency by the gaba reuptake inhibitor tiagabine revealed by robust peak frequency estimation. Hum. Brain Mapp. 2016, 37, 3882–3896. [Google Scholar] [CrossRef] [PubMed]
- Booker, S.A.; Harada, H.; Elgueta, C.; Bank, J.; Bartos, M.; Kulik, A.; Vida, I. Presynaptic GABAB receptors functionally uncouple somatostatin interneurons from the active hippocampal network. eLife 2020, 9, e51156. [Google Scholar] [CrossRef]
- Kullander, K.; Topolnik, L. Cortical disinhibitory circuits: Cell types, connectivity and function. Trends Neurosci. 2021, 44, 643–657. [Google Scholar] [CrossRef]
- Brown, J.T.; Davies, C.H.; Randall, A.D. Synaptic activation of GABA(B) receptors regulates neuronal network activity and entrainment. Eur. J. Neurosci. 2007, 25, 2982–2990. [Google Scholar] [CrossRef]
- Basu, J.; Zaremba, J.D.; Cheung, S.K.; Hitti, F.L.; Zemelman, B.V.; Losonczy, A.; Siegelbaum, S.A. Gating of hippocampal activity, plasticity, and memory by entorhinal cortex long-range inhibition. Science 2016, 351, aaa5694. [Google Scholar] [CrossRef]
- Stuyt, G.; Godenzini, L.; Palmer, L.M. Local and Global Dynamics of Dendritic Activity in the Pyramidal Neuron. Neuroscience 2022, 489, 176–184. [Google Scholar] [CrossRef]
- Koch, C.; Poggio, T.; Torre, V. Nonlinear interactions in a dendritic tree: Localization, timing, and role in information processing. Proc. Natl. Acad. Sci. USA 1983, 80, 2799–2802. [Google Scholar] [CrossRef] [PubMed]
- Pinault, D.; Deschênes, M. Anatomical evidence for a mechanism of lateral inhibition in the rat thalamus. Eur. J. Neurosci. 1998, 10, 3462–3469. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Localization of GAT-1 and GABAB Rs in synaptic regions and a disinhibitory circuit. (A) Schematic representation of the localization and effect of GAT-1 and GABAB Rs. In dendrites, the GABAB receptor activates the GIRK, TREK2, and VGCC-type channels and inhibits the backpropagation of the action potential while reducing the generation of Ca2+ spikes. (B) The scheme represents a disinhibitory circuit as a mechanism for activating specific pathways and its effect on oscillation. The microcircuit includes serial connections between two inhibitory interneurons and a main excitatory neuron and their effect on oscillation. The thick lines represent disinhibition.
Figure 1.
Localization of GAT-1 and GABAB Rs in synaptic regions and a disinhibitory circuit. (A) Schematic representation of the localization and effect of GAT-1 and GABAB Rs. In dendrites, the GABAB receptor activates the GIRK, TREK2, and VGCC-type channels and inhibits the backpropagation of the action potential while reducing the generation of Ca2+ spikes. (B) The scheme represents a disinhibitory circuit as a mechanism for activating specific pathways and its effect on oscillation. The microcircuit includes serial connections between two inhibitory interneurons and a main excitatory neuron and their effect on oscillation. The thick lines represent disinhibition.


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Villalobos, N. Disinhibition Is an Essential Network Motif Coordinated by GABA Levels and GABA B Receptors. Int. J. Mol. Sci. 2024, 25, 1340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25021340
AMA Style
Villalobos N. Disinhibition Is an Essential Network Motif Coordinated by GABA Levels and GABA B Receptors. International Journal of Molecular Sciences. 2024; 25(2):1340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25021340
Chicago/Turabian StyleVillalobos, Nelson. 2024. "Disinhibition Is an Essential Network Motif Coordinated by GABA Levels and GABA B Receptors" International Journal of Molecular Sciences 25, no. 2: 1340. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25021340
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.