
ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice

1
Department of Basic Veterinary Science, Laboratory of Physiology, Joint Graduate School of Veterinary Sciences, Gifu University, 1-1 Yanagido, Gifu 501-1193, Japan
2
Division of Animal Medical Science, Center for One Medicine Innovative Translational Research (COMIT), Gifu University Institute for Advanced Study, 1-1 Yanagido, Gifu 501-1193, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(4), 1985; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25041985
Submission received: 19 December 2023
/
Revised: 31 January 2024
/
Accepted: 5 February 2024
/
Published: 6 February 2024
(This article belongs to the Collection Feature Papers in Molecular Neurobiology)
Abstract
:The tunica muscularis of mammalian esophagi is composed of striated muscle and smooth muscle. Contraction of the esophageal striated muscle portion is mainly controlled by cholinergic neurons. On the other hand, smooth muscle contraction and relaxation are controlled not only by cholinergic components but also by non-cholinergic components in the esophagus. Adenosine triphosphate (ATP) is known to regulate smooth muscle contraction and relaxation in the gastrointestinal tract via purinergic receptors. However, the precise mechanism of purinergic regulation in the esophagus is still unclear. Therefore, the aim of the present study was to clarify the effects of ATP on the mechanical responses of the esophageal muscle in mice. An isolated segment of the mouse esophagus was placed in a Magnus’s tube and longitudinal mechanical responses were recorded. Exogenous application of ATP induced contractile responses in the esophageal preparations. Tetrodotoxin, a blocker of voltage-dependent sodium channels in neurons and striated muscle, did not affect the ATP-induced contraction. The ATP-evoked contraction was blocked by pretreatment with suramin, a purinergic receptor antagonist. RT-PCR revealed the expression of mRNA of purinergic receptor genes in the mouse esophageal tissue. The findings suggest that purinergic signaling might regulate the motor activity of mouse esophageal smooth muscle.
1. Introduction
Motor functions of the gastrointestinal tract are regulated by various transmitters including purines such as adenosine, adenosine diphosphate (ADP) and adenosine triphosphate (ATP) [1,2,3,4,5]. There are two types of purine receptors: one type are P1 receptors for adenosine and the other type are P2 receptors for ATP and ADP [1,2,6,7]. P2 receptors include P2X receptors and P2Y receptors. P2X receptors are ligand-gated ion channels and P2Y receptors are G protein-coupled receptors (GPCRs) [1,2,6,7]. At present, seven P2X (P2X1-7) and eight P2Y (P2Y1, 2, 4, 6, and 11–14) receptor subtypes have been identified [8]. However, there are also some reports that rodents such as rats and mice do not have P2Y11 receptors [9]. P2 receptors are expressed in the gastrointestinal tract, including the esophagus [2,8,10,11,12,13]. Previous studies have demonstrated that purines cause excitatory and inhibitory responses in the smooth muscle cells of the gastrointestinal tract [2,8,11,14,15].
The tunica muscularis of mammalian esophagi contains not only smooth muscle fibers but also striated muscle fibers [16,17]. For example, the upper portion is composed of striated muscle fibers, the lower portion is composed of smooth muscle fibers, and the middle portion is a mixed composition of these fibers in human and cat esophagi [17]. On the other hand, in dogs, ruminants, and rodents, including mice, rats and hamsters, the muscle layer of the esophagus consists largely of striated muscle fibers [17]. The motility of the esophagus is controlled by extrinsic cholinergic vagal neurons [18,19] and by intrinsic myenteric neurons releasing various neurotransmitters such as acetylcholine, tachykinins, nitric oxide, and galanin [17,19,20,21,22]. In the case of striated muscles, cholinergic receptors are nicotinic receptors, while in the case of smooth muscles, they are muscarinic receptors. In addition, the esophagus has longitudinal smooth muscle fibers in the muscularis mucosae even in mammals that have a striated muscle esophagus [23,24]. The mouse esophagus also contains a smooth muscle layer in the muscularis mucosae. The esophageal muscularis mucosa is used for the study of smooth muscle contractility in the esophagus [23,24,25,26,27].
Purinergic receptors have been detected histologically in the esophagus [11,12]. Although there have been functional studies on the purinergic regulation of esophageal smooth muscle [27,28], the precise mechanism is still unclear. Therefore, the aim of the present study was to determine the characteristics of mechanical responses induced by ATP in the mouse esophagus.
2. Results
2.1. Molecular Identification of P2 Receptors in the Mouse Esophagus
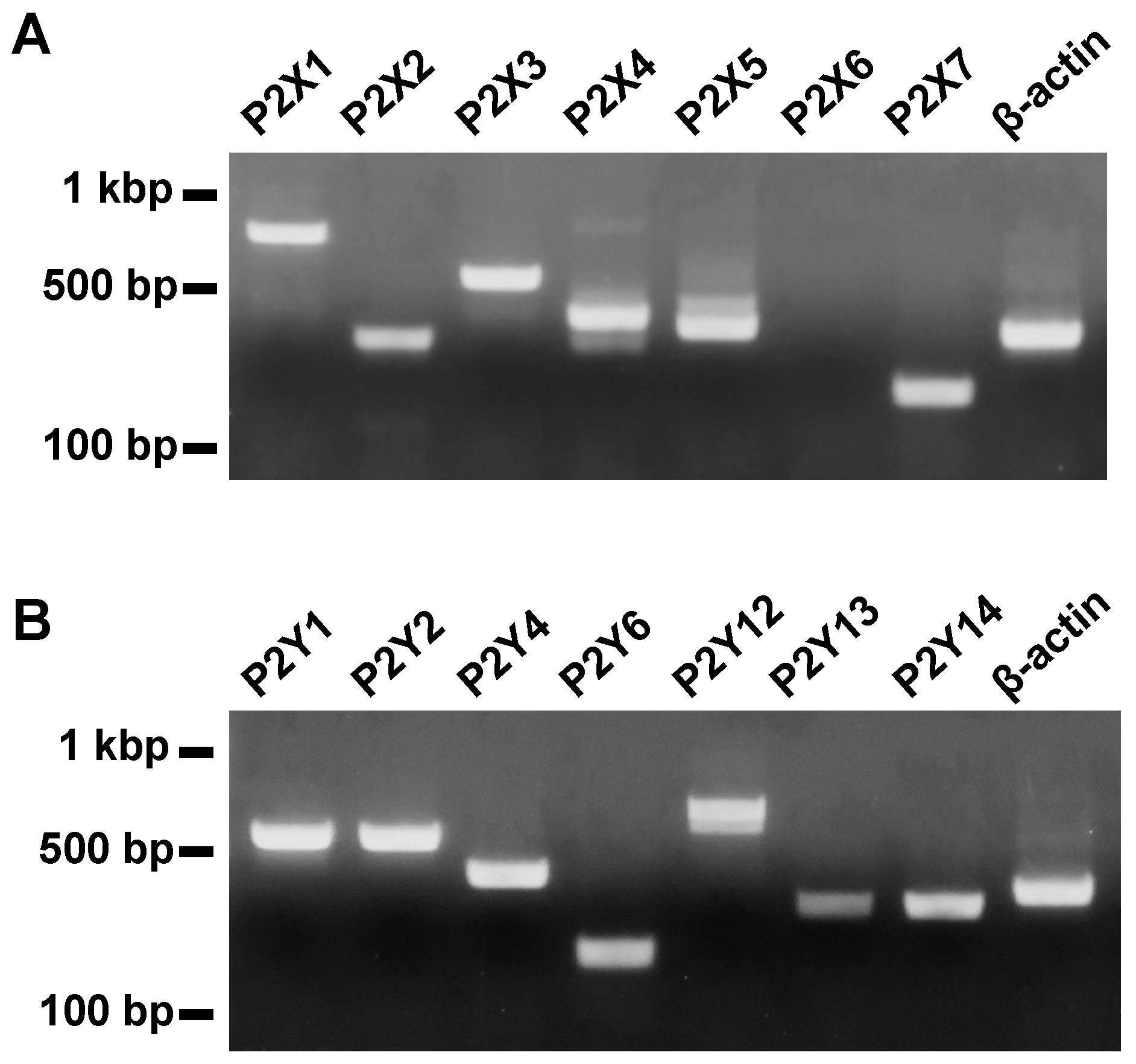
We examined the expression of subtypes of P2 receptors in mouse esophageal tissues by using reverse transcription polymerase chain reaction (RT-PCR). Amplified products of mRNA of P2X1, 2, 3, 4, 5, and 7 receptors and P2Y1, 2, 4, 6, 12, 13, and 14 receptors were observed in appropriate sizes (Figure 1).
2.2. Effects of ATP on the Mechanical Activity of Mouse Esophageal Segments
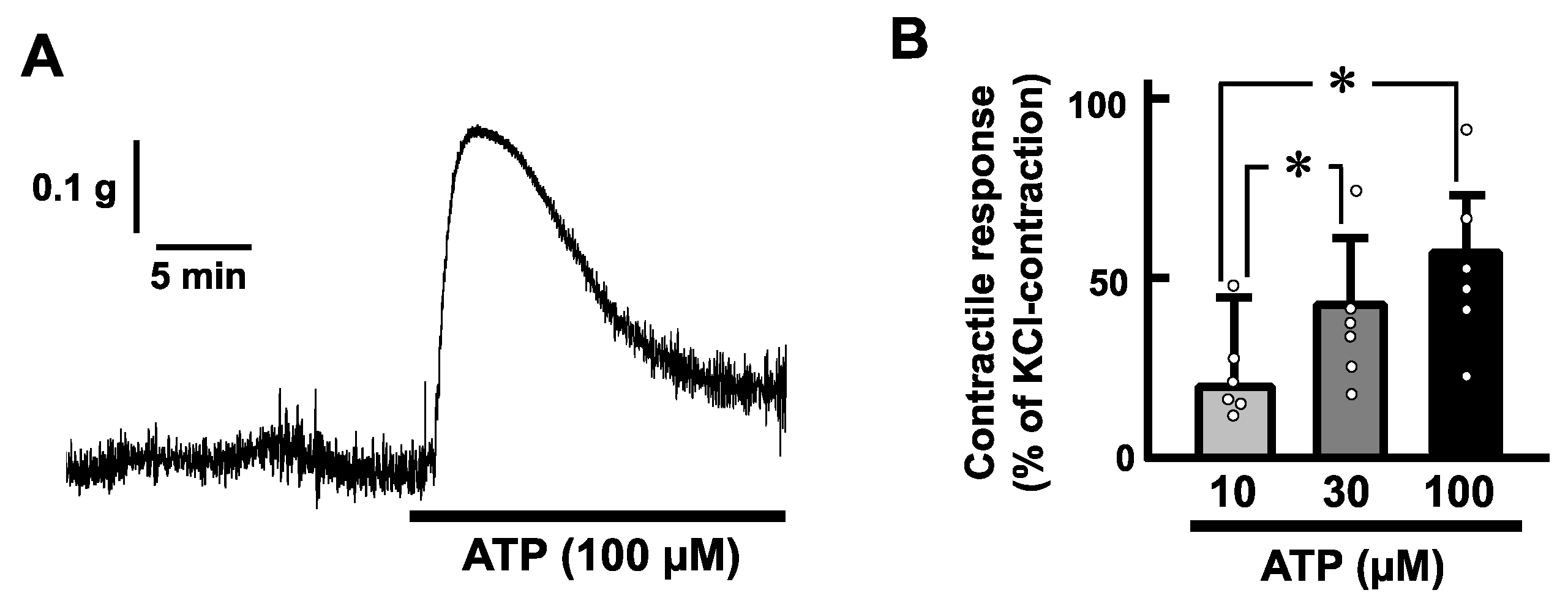
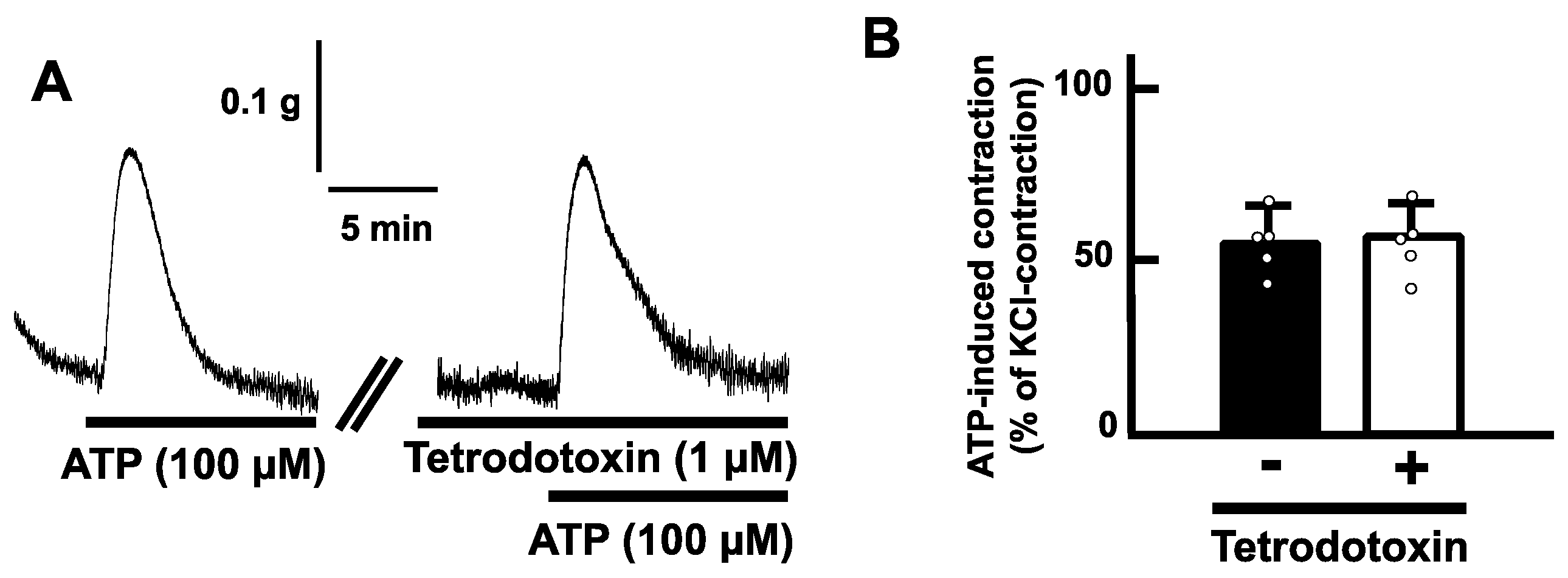
Exogenous application of ATP (100 µM) induced a transient contraction in a longitudinal direction of the mouse esophageal segments (Figure 2A). The contractile responses increased in a concentration-dependent manner (Figure 2B). On the other hand, tetrodotoxin (1 µM) did not affect the ATP-induced contractions of the mouse esophagus (Figure 3).
2.3. Effects of Purinoceptor Antagonists on ATP-Evoked Contractions in the Mouse Esophagus
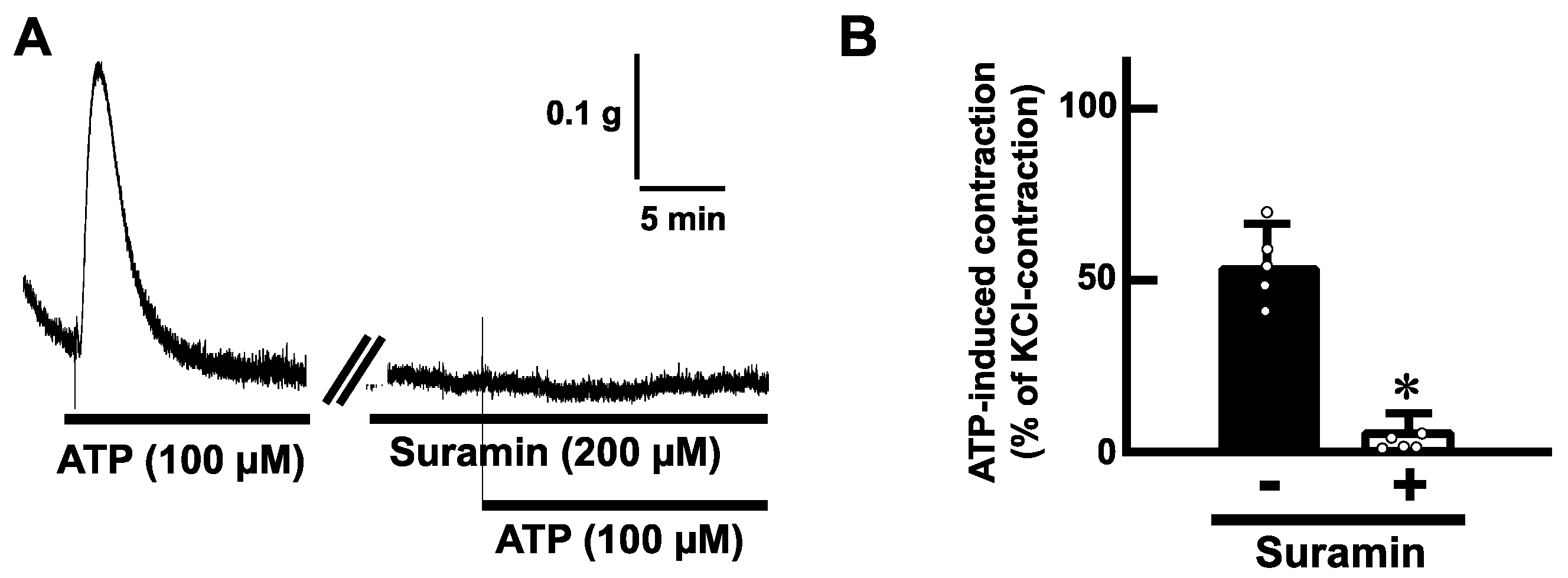
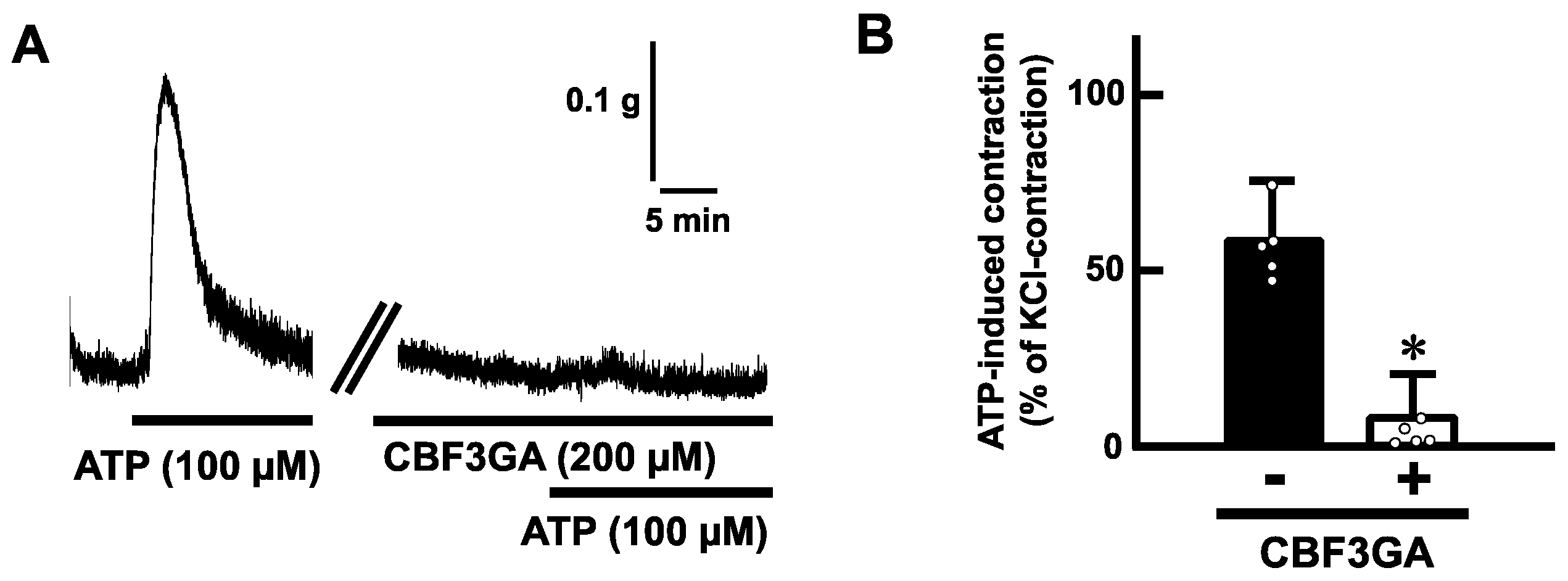
To determine whether ATP-evoked contractions are mediated via purinoceptors, we examined the effects of antagonists for purinoceptors. Pretreatment with suramin (200 µM), a non-selective P2 receptor antagonist, blocked the ATP (100 µM)-induced contractions in mouse esophageal segments (Figure 4). Next, we tested a selective antagonist for the P2Y receptor. Pretreatment with cibacron blue F3GA (CBF3GA) (200 µM), a P2Y receptor antagonist, inhibited the ATP-evoked contractions (Figure 5).
3. Discussion
In the present study, we investigated the characteristics of mechanical responses induced by ATP in the mouse esophagus. Our major findings are: (1) we detected expression of P2 receptors in the mouse esophagus, (2) exogenous application of ATP evoked contractions of the esophageal smooth muscle, and (3) pretreatment with a selective P2Y receptor antagonist inhibited the ATP-induced contraction. These findings suggest that ATP is involved in excitatory regulation of the longitudinal smooth muscle in the muscularis mucosae of the mouse esophagus via P2Y receptors.
The muscle composition of the tunica muscularis is striated muscle fiber in the mouse esophagus [17]. In addition, the mouse esophagus also contains a smooth muscle layer in the muscularis mucosae. The smooth muscle layer in the muscularis mucosae is longitudinally arranged and thus can express longitudinal mechanical responses exclusively [23,29,30]. Our results showed that application of ATP evoked contraction longitudinally in the esophageal segments. To determine whether ATP acts on smooth muscle or striated muscle, we used tetrodotoxin, which can inhibit contractile activity of striated muscle via blockade of voltage-dependent sodium channels. Pretreatment with tetrodotoxin did not affect the ATP-induced contraction. The concentration of tetrodotoxin used in this study (1 µM) is enough to abolish esophageal striated muscle contractility [23]. Hence, it is reasonable that ATP-induced contraction in the mouse esophagus is a smooth muscle activity in the muscularis mucosae. In addition, since tetrodotoxin can block neuronal activity, the possibility of involvement of neurons in ATP-induced contraction also might be ruled out.
Pretreatment with application of a P2Y receptor antagonist inhibited the ATP-induced contraction in the mouse esophagus. The results indicate that P2Y receptors are involved in ATP-induced contraction of the mouse esophagus. It is known that ATP causes excitatory responses in the smooth muscle cells of other gastrointestinal tracts [2,8,14,15]. The esophagus might have a similar regulation system via P2Y receptors. P2Y receptors are GPCRs [1,2,6,7]. P2Y1, 2, 4, and 6 are coupled preferentially with Gq/11 protein [1], which can activate phospholipase C, generate inositol trisphosphate, increase intracellular calcium, or generate diacylglycerol, and activate protein kinase C and then lead to smooth muscle contractions [29]. ATP acts on several P2Y receptors with high affinity [1]. Further investigation is required to identify the subtype of P2Y receptors involved in the excitatory regulation by ATP in esophageal motility.
In previous studies, the mouse esophagus contained P2X3 receptors in the myenteric neurons [12]. If exogenous application of ATP acts on the P2X3 receptors, P2X3 receptor-positive myenteric neurons might be activated, which might induce responses in the esophagus. In this study, however, ATP-induced contractile responses were not affected by pretreatment with tetrodotoxin, which is a blocker of voltage-dependent sodium channels in the neurons. In addition, ATP-induced contractile responses were inhibited by pretreatment with the P2Y receptor antagonist. The results show that our obtained data are not associated with the biological activity of P2X3 receptors in the myenteric neurons. In future, examination of the roles of P2X receptors in the esophagus should be performed to advance studies on purinergic regulation of esophageal motor functions.
In this study, we did not identify endogenous sources of ATP in the mouse esophagus. Interestingly, Mihara et al. reported that ATP is released from epithelial keratinocytes in the mouse esophagus in response to TRPV4 activation [31,32]. We therefore consider that epithelial cells are candidate sources. However, it should be noted that we cannot exclude neurons, glial cells, and muscle cells as sources of ATP. Indeed, it is known that skeletal muscle releases ATP via pannexin channels during repetitive contraction [33].
We have identified various regulatory factors in esophageal motility including tachykinins, nitric oxide, galanin, serotonin [20,22,23,24], and purines in this study. Generally, it is important to control contraction and/or relaxation in the longitudinal direction separately from those in the circular direction for effective peristalsis of the gastrointestinal tract [3,34]. In esophageal peristaltic activity, the longitudinal motor response plays an important role and assists effective propulsion [35]. Our findings suggest that the purinergic system may contribute to effective propulsion in the esophagus: whereas, it should be noted that our experiments were performed using isolated esophageal segments in the organ bath. To advance investigation of purinergic regulation of esophageal motility, an in vivo experimental system is necessary. This is because esophageal motility is caused by the vagus nerve reflex that is integrated in the medulla oblongata [18,19]. We devised an in vivo system for recording esophageal motor activity [36], and this system can be used for further experiments.
In the human esophagus, the upper portion is composed of striated muscle fibers, the lower portion is composed of smooth muscle fibers, and the middle portion is a mixed composition of these fibers [17]. In mice, on the other hand, the muscle layer of the esophagus consists largely of striated muscle fibers [17]. However, longitudinal smooth muscle fibers in the muscularis mucosae commonly exist in both the human and mouse esophagus [18,19]. So, the findings obtained in this study can be applied to human healthcare.
In conclusion, the present study clarified that ATP induces contractile responses of longitudinal smooth muscle in the muscularis mucosae of the mouse esophagus via P2Y receptors. This purinergic regulation might contribute to esophageal motility.
4. Materials and Methods
4.1. Animals
Male ddY mice (Mus musculus, 8–12 weeks of age, 30–40 g in weight) were purchased from Japan SLC (Shizuoka, Japan). The number of animals used was 23. They were maintained in plastic cages at 24 ± 2 °C with a 12:12 h light–dark cycle (light on at 07:00–19:00) and given free access to laboratory chow and water. The experiments were approved by the Gifu University Animal Care and Use Committee and were conducted in accordance with the committee guidelines on animal care and use (permission numbers: H30-183, 2019-239, 2020-252). The issuance and expiry date of the license are 1 April 2019 and 31 March 2022, respectively.
4.2. Esophageal Tissue Preparations
The mice were anesthetized with isoflurane and were exsanguinated via axillary arteries. We isolated a 1 cm long segment from the middle thoracic part of the esophagus. The segment of the esophagus was immediately immersed in Krebs’s solution (see below) at room temperature, and the intraluminal contents of the excised segment were flushed using a small cannula containing Krebs’s solution.
4.3. Recording of Mechanical Activity in Esophageal Segments
To record contractile responses in the longitudinal direction, the whole segment was mounted in a Magnus’s tube (10 mL in capacity) filled with Krebs’s solution (pH 7.4). One end of the esophageal segment was tied to the Magnus’s tube and the other end was secured with a silk thread to an isometric force transducer (T7-8-240, Orientec, Tokyo, Japan). The Krebs’s solution was continuously bubbled with a 95% O2 + 5% CO2 gas mixture and maintained at 37 °C. Mechanical responses, which were filtered and amplified by an amplifier (NEC, AS1202, Tokyo, Japan), were recorded using a PowerLab system (AD Instruments, Bella Vista, NSW, Australia). An initial resting tension of 1.0 g was added to the esophageal segment, which was subsequently allowed to equilibrate for at least 30 min. The esophageal segments were used for experiments for at least 6 h.
4.4. Solutions and Drugs
During the experiments, tissues were maintained in Krebs’s solution consisting of (mM): NaCl 118.4, KCl 4.7, CaCl2 2.5, MgSO4 1.2, KH2PO4 1.2, NaHCO3 25, and glucose 11.7. ATP was obtained from Tokyo Chemical Industry (Tokyo, Japan). Tetrodotoxin was obtained from FUJIFILM Wako (Osaka, Japan). Suramin and CBF3GA, which were used as antagonists for purinergic receptors [37,38], were obtained from Sigma-Aldrich (St Louis, MO, USA). The drugs were dissolved in distilled water. We confirmed that the highest concentration of vehicles (0.1%) for the drugs alone had no effect on the basal tone and contractile responses at the concentrations used. Final concentrations in the bath solution were described as the concentrations of drugs.
4.5. RNA Isolation and RT-PCR
The expression of P2 receptor gene mRNAs was assessed by RT-PCR. Total cellular RNA was extracted from homogenates of the mouse esophageal mucosa tissue using TRI Reagent (Molecular Research Center, Cincinnati, OH, USA). First-strand cDNA was synthesized from 2 µg of total RNA by using SuperScript III Reverse Transcriptase (Thermo Fisher Scientific, Waltham, MA, USA) and Random primers (Thermo Fisher Scientific). PCR was performed with Platinum Taq DNA Polymerase High Fidelity (Thermo Fisher Scientific). The primer sets are shown in Table 1. All primers were designed according to previous reports [39,40] and obtained from Thermo Fisher Scientific. Amplifications were performed by 35 cycles. The reaction products were electrophoresed on 1.5% agarose gels and stained with ethidium bromide (0.8 µg/mL). The gels were imaged with a UV transilluminator (UVP Laboratory Products, Upland, CA, USA) and photographed.
4.6. Data Analysis
Data are presented as means ± standard error of the mean (S.E.M.). n indicates the number of separate preparations. The values of contractile responses are maximum amplitudes of contractions induced by application of ATP that are normalized as percentages of KCl (60 mM)-induced contractions in the same preparations. KCl was applied into Krebs’s solution at the latest time point of each experiment. The significance of differences between mean values was determined by one-way analysis of variance followed by the Turkey–Kramer test for the comparison of multiple groups or by the paired t-test for comparison of two groups. A p value less than 0.05 denotes the presence of a statistically significant difference.
Author Contributions
Formal analysis, Y.S. (Yuji Suzuki), Y.S. (Yasutake Shimizu), and T.S.; investigation, Y.S. (Yuji Suzuki) and T.S.; methodology, Y.S. (Yuji Suzuki) and T.S.; supervision, T.S.; writing—original draft, Y.S. (Yuji Suzuki) and T.S.; writing—review and editing, Y.S. (Yasutake Shimizu). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by research grants from the Koshiyama Science & Technology Foundation (Gifu, Japan), Research grants from the OGAWA Science and Technology Foundation (Gifu, Japan), and grants-in-aid for scientific research (KAKENHI) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan [Research Project Numbers: 17K08122, 20K06409 and 23K05553].
Institutional Review Board Statement
The animal study protocol was approved by the Gifu University Animal Care and Use Committee (permission numbers are H30-183, 2019-239, and 2020-252 and date of approval are 1 April 2019, 1 April 2020 and 1 April 2021, respectively).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors are indebted to Ayako Takahashi (Gifu University, Japan) for technical support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Burnstock, G.; Ralevic, V. Purinergic signaling and blood vessels in health and disease. Pharmacol. Rev. 2013, 66, 102–192. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signalling in the gastrointestinal tract and related organs in health and disease. Purinergic Signal. 2014, 10, 3–50. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The Enteric Nervous System; Blackwell Publishing: Malden, MA, USA, 2006; pp. 103–131. [Google Scholar]
- Hansen, M.B. Neurohumoral control of gastrointestinal motility. Physiol. Res. 2003, 52, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M. G protein-coupled receptors in gastrointestinal physiology. IV. Neural regulation of gastrointestinal smooth muscle. Am. J. Physiol. 1998, 275, G1–G7. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Development and perspectives of the purinoceptor concept. J. Auton. Pharmacol. 1996, 16, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. The past, present and future of purine nucleotides as signalling molecules. Neuropharmacology 1997, 36, 1127–1139. [Google Scholar] [CrossRef]
- Jiménez, M.; Clavé, P.; Accarino, A.; Gallego, D. Purinergic neuromuscular transmission in the gastrointestinal tract; functional basis for future clinical and pharmacological studies. Br. J. Pharmacol. 2014, 171, 4360–4375. [Google Scholar] [CrossRef]
- Dreisig, K.; Kornum, B.R. A critical look at the function of the P2Y11 receptor. Purinergic Signal. 2016, 12, 427–437. [Google Scholar] [CrossRef]
- Giaroni, C.; Knight, G.E.; Zanetti, E.; Chiaravalli, A.M.; Lecchini, S.; Frigo, G.; Burnstock, G. Postnatal development of P2 receptors in the murine gastrointestinal tract. Neuropharmacology 2006, 50, 690–704. [Google Scholar] [CrossRef]
- Goyal, R.K.; Sullivan, M.P.; Chaudhury, A. Progress in understanding of inhibitory purinergic neuromuscular transmission in the gut. Neurogastroenterol. Motil. 2013, 25, 203–207. [Google Scholar] [CrossRef]
- Kestler, C.; Neuhuber, W.L.; Raab, M. Distribution of P2X3 receptor immunoreactivity in myenteric ganglia of the mouse esophagus. Histochem. Cell Biol. 2009, 131, 13–27. [Google Scholar] [CrossRef]
- Wang, Z.-J.; Neuhuber, W.L. Intraganglionic laminar endings in the rat esophagus contain purinergic P2X2 and P2X3 receptor immunoreactivity. Anat. Embryol. 2003, 207, 363–371. [Google Scholar] [CrossRef]
- Yuan, W.; Wang, Z.; Li, J.; Li, D.; Liu, D.; Bai, G.; Walsh, M.P.; Gui, Y.; Zheng, X.-L. Uridine adenosine tetraphosphate induces contraction of circular and longitudinal gastric smooth muscle by distinct signaling pathways. IUBMB Life 2013, 65, 623–632. [Google Scholar] [CrossRef]
- Zizzo, M.G.; Mulè, F.; Serio, R. Evidence that ATP or a related purine is an excitatory neurotransmitter in the longitudinal muscle of mouse distal colon. Br. J. Pharmacol. 2007, 151, 152–160. [Google Scholar] [CrossRef]
- Neuhuber, W.L.; Wörl, J. Enteric co-innervation of striated muscle in the esophagus: Still enigmatic? Histochem. Cell Biol. 2016, 146, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Wörl, J.; Neuhuber, W.L. Enteric co-innervation of motor endplates in the esophagus: State of the art ten years after. Histochem. Cell Biol. 2005, 123, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Bieger, D.; Neuhuber, W.L. Neural circuits and mediators regulating swallowing in the brainstem. GI Motil. Online 2006. [Google Scholar]
- Clouse, R.E.; Diamant, N.E. Motor Function of the Esophagus. In Physiology of the Gastrointestinal Tract, 4th ed.; Johnson, L.R., Ed.; Elsevier Academic Press: Burlington, MA, USA, 2006; pp. 913–926. [Google Scholar]
- Boudaka, A.; Wörl, J.; Shiina, T.; Shimizu, Y.; Takewaki, T.; Neuhuber, W.L. Galanin modulates vagally induced contractions in the mouse esophagus. Neurogastroenterol. Motil. 2009, 21, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Izumi, N.; Matsuyama, H.; Ko, M.; Shimizu, Y.; Takewaki, T. Role of intrinsic nitrergic neurones on vagally mediated striated muscle contractions in the hamster oesophagus. J. Physiol. 2003, 551, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Shiina, T.; Shimizu, Y.; Boudaka, A.; Wörl, J.; Takewaki, T. Tachykinins are involved in local reflex modulation of vagally mediated striated muscle contractions in the rat esophagus via tachykinin NK1 receptors. Neuroscience 2006, 139, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Shiina, T.; Shima, T.; Hirayama, H.; Kuramoto, H.; Takewaki, T.; Shimizu, Y. Contractile responses induced by physalaemin, an analogue of substance P, in the rat esophagus. Eur. J. Pharmacol. 2010, 628, 202–206. [Google Scholar] [CrossRef]
- Shiina, T.; Naitou, K.; Nakamori, H.; Suzuki, Y.; Horii, K.; Sano, Y.; Shimaoka, H.; Shimizu, Y. Serotonin-induced contractile responses of esophageal smooth muscle in the house musk shrew (Suncus murinus). Neurogastroenterol. Motil. 2016, 28, 1641–1648. [Google Scholar] [CrossRef]
- Bieger, D.; Triggle, C. Pharmacological properties of mechanical responses of the rat oesophageal muscularis mucosae to vagal and field stimulation. Br. J. Pharmacol. 1985, 84, 93–106. [Google Scholar] [PubMed]
- Christensen, J.; Percy, W.H. A pharmacological study of oesophageal muscularis mucosae from the cat, dog and American opossum (Didelphis virginiana). Br. J. Pharmacol. 1984, 83, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Percy, W.H.; Miller, A.J.; Brunz, J.T. Pharmacologic characteristics of rabbit esophageal muscularis mucosae in vitro. Dig. Dis. Sci. 1997, 42, 2537–2546. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.H.; Jung, H.; Cho, E.J.; Jeong, J.H.; Sohn, U.D. The Signaling Mechanism of Contraction Induced by ATP and UTP in Feline Esophageal Smooth Muscle Cells. Mol. Cells 2015, 38, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Storr, M.; Geisler, F.; Neuhuber, W.L.; Schusdziarra, V.; Allescher, H.D. Endomorphin-1 and -2, endogenous ligands for the μ-opioid receptor, inhibit striated and smooth muscle contraction in the rat oesophagus. Neurogastroenterol. Motil. 2000, 12, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Storr, M.; Geisler, F.; Neuhuber, W.L.; Schusdziarra, V.; Allescher, H.D. Characterization of vagal input to the rat esophageal muscle. Auton. Neurosci. Basic Clin. 2001, 91, 1–9. [Google Scholar] [CrossRef]
- Boudaka, A.; Tominaga, M. Physiological and Pathological Significance of Esophageal TRP Channels: Special Focus on TRPV4 in Esophageal Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 4550. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H.; Boudaka, A.; Sugiyama, T.; Moriyama, Y.; Tominaga, M. Transient receptor potential vanilloid 4 (TRPV4)-dependent calcium influx and ATP release in mouse oesophageal keratinocytes. J. Physiol. 2011, 589, 3471–3482. [Google Scholar] [CrossRef]
- Riquelme, M.A.; Cea, L.A.; Vega, J.L.; Boric, M.P.; Monyer, H.; Bennett, M.V.; Frank, M.; Willecke, K.; Sáez, J.C. The ATP required for potentiation of skeletal muscle contraction is released via pannexin hemichannels. Neuropharmacology 2013, 75, 594–603. [Google Scholar] [CrossRef]
- Furness, J.B. The enteric nervous system: Normal functions and enteric neuropathies. Neurogastroenterol. Motil. 2008, 20 (Suppl. S1), 32–38. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.K. Longitudinal muscle of the esophagus: Its role in esophageal health and disease. Curr. Opin. Gastroenterol. 2013, 29, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Horii, K.; Shiina, T.; Naitou, K.; Nakamori, H.; Horii, Y.; Shimaoka, H.; Shimizu, Y. Characterization of peristaltic motility in the striated muscle portion of the esophagus using a novel in vivo method in rats. Neurogastroenterol. Motil. 2019, 31, e13518. [Google Scholar] [CrossRef] [PubMed]
- Uneyama, H.; Uneyama, C.; Ebihara, S.; Akaike, N. Suramin and reactive blue 2 are antagonists for a newly identified purinoceptor on rat megakaryocyte. Br. J. Pharmacol. 1994, 111, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Fallon, M.; Bobbin, R.P. ATP antagonists cibacron blue, basilen blue and suramin alter sound-evoked responses of the cochlea and auditory nerve. Hear. Res. 1994, 78, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Hayato, R.; Ohtubo, Y.; Yoshii, K. Functional expression of ionotropic purinergic receptors on mouse taste bud cells. J. Physiol. 2007, 584, 473–488. [Google Scholar] [CrossRef]
- Hao, M.M.; Bergner, A.J.; Hirst, C.S.; Stamp, L.A.; Casagranda, F.; Bornstein, J.C.; Boesmans, W.; Vanden Berghe, P.; Young, H.M. Spontaneous calcium waves in the developing enteric nervous system. Dev. Biol. 2017, 428, 74–87. [Google Scholar] [CrossRef]
Figure 1.
Expression of subtypes of P2 receptors in the mouse esophageal tissue determined by RT-PCR. Amplified products of mRNA of P2X1, P2X2, P2X3, P2X4, P2X5, P2X6, P2X7, P2Y1, P2Y2, P2Y4, P2Y6, P2Y12, P2Y13, P2Y14, and β-actin were detected in appropriate sizes (n = 3). (A) Shows expression of P2X receptors and β-actin. (B) Shows expression of P2Y receptors and β-actin.
Figure 1.
Expression of subtypes of P2 receptors in the mouse esophageal tissue determined by RT-PCR. Amplified products of mRNA of P2X1, P2X2, P2X3, P2X4, P2X5, P2X6, P2X7, P2Y1, P2Y2, P2Y4, P2Y6, P2Y12, P2Y13, P2Y14, and β-actin were detected in appropriate sizes (n = 3). (A) Shows expression of P2X receptors and β-actin. (B) Shows expression of P2Y receptors and β-actin.

Figure 2.
ATP-evoked contractions in the mouse esophagus. (A) A representative trace demonstrating the effect of ATP on longitudinal tension of the mouse esophagus is shown. (B) Dose-dependency of the contractile responses evoked by ATP in the mouse esophagus is summarized (n = 6). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± standard error of the mean (S.E.M.). * p < 0.05.
Figure 2.
ATP-evoked contractions in the mouse esophagus. (A) A representative trace demonstrating the effect of ATP on longitudinal tension of the mouse esophagus is shown. (B) Dose-dependency of the contractile responses evoked by ATP in the mouse esophagus is summarized (n = 6). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± standard error of the mean (S.E.M.). * p < 0.05.

Figure 3.
Effects of tetrodotoxin on ATP-evoked contraction in the mouse esophagus. (A) Representative traces of the contraction induced by ATP (100 µM) of the mouse esophagus in the absence or presence of tetrodotoxin (1 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of tetrodotoxin (1 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar graph represents the mean of data ± S.E.M.
Figure 3.
Effects of tetrodotoxin on ATP-evoked contraction in the mouse esophagus. (A) Representative traces of the contraction induced by ATP (100 µM) of the mouse esophagus in the absence or presence of tetrodotoxin (1 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of tetrodotoxin (1 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar graph represents the mean of data ± S.E.M.

Figure 4.
Effects of a non-selective antagonist for P2 receptors on ATP-evoked contraction in the mouse esophagus. (A) Representative tracings demonstrating the contraction induced by ATP (100 µM) in the mouse esophagus under the absence or presence of suramin (200 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of suramin (200 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± S.E.M. * p < 0.05, compared to the control.
Figure 4.
Effects of a non-selective antagonist for P2 receptors on ATP-evoked contraction in the mouse esophagus. (A) Representative tracings demonstrating the contraction induced by ATP (100 µM) in the mouse esophagus under the absence or presence of suramin (200 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of suramin (200 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± S.E.M. * p < 0.05, compared to the control.

Figure 5.
Effects of a selective antagonist for P2Y receptors on ATP-evoked contraction in the mouse esophagus. (A) Representative tracings demonstrating the contraction induced by ATP (100 µM) of the mouse esophagus in the absence or presence of CBF3GA (200 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of CBF3GA (200 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± S.E.M. * p < 0.05, compared to the control.
Figure 5.
Effects of a selective antagonist for P2Y receptors on ATP-evoked contraction in the mouse esophagus. (A) Representative tracings demonstrating the contraction induced by ATP (100 µM) of the mouse esophagus in the absence or presence of CBF3GA (200 µM) are shown. (B) Summarized bar graphs of contraction evoked by ATP (100 µM) of the mouse esophagus in the absence (−) or presence (+) of CBF3GA (200 µM) are shown (n = 5). The values of contractile responses are normalized as percentages of KCl (60 mM)-induced contractions. Each bar represents the mean of data ± S.E.M. * p < 0.05, compared to the control.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of primers for RT-PCR.
| Gene | Sequence (5′-3′) | Predicted Size (bp) |
|---|---|---|
| P2X1 | Forward CATTGTGCAGAGAACCCAGAA Reverse ATGTCCTCCGCATACTTGAAC | 776 |
| P2X2 | Forward ACGTTCATGAACAAAAACAAG Reverse TCAAAGTTGGGCCAAACCTTTGG | 360 |
| P2X3 | Forward AAGAGTGGGCAGTTACAAGGG Reverse GAAAACCCACCCCACAAAGT | 576 |
| P2X4 | Forward GAGAATGACGCTGGTGTGCC Reverse TTGGTGAGTGTGCGTTGCTC | 437 |
| P2X5 | Forward TAGTTAATGGCAAGGCGGGA Reverse AGCTCTGGCTACGTCTTCAC | 409 |
| P2X6 | Forward TACGTACTAACAGACGCA Reverse ATATCAGGGTTCTTTGGG | 254 |
| P2X7 | Forward TGTTTCCTTTGGCTGCTCCT Reverse CGCTCACCAAAGCAAAGCTAAT | 239 |
| P2Y1 | Forward TGGCGTGGTGTACCCTCTCAAGTC Reverse CGGGACAGTCTCCTTCTGAATGTA | 558 |
| P2Y2 | Forward CTCACGCGCACCCTCTACTA Reverse TCGGGTGCACTGCCTTTCTT | 549 |
| P2Y4 | Forward CTTTGGCTTTCCCTTCTTGA Reverse GTCCGCCCACCTGCTGAT | 427 |
| P2Y6 | Forward GCCCTGTGCTGGAGACCTTC Reverse CATGGCCCCAGTGACAAACA | 226 |
| P2Y12 | Forward CAGTGCAAGGGGTGGCATCT Reverse TGGCACACCAAGGTTCTCAG | 618 |
| P2Y13 | Forward GAAGAGAGGCACATGCAACA Reverse TTACTAATGCCAGGCCAACC | 345 |
| P2Y14 | Forward CAGTGCATGGAGCTCAAAAA Reverse GCAGCCGAGAGTAGCAGAGT | 347 |
| β-actin | Forward TGACCCTGAAGTACCCCATTG Reverse TCAGGATCTTCATGAGGTAG | 387 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Suzuki, Y.; Shimizu, Y.; Shiina, T. ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice. Int. J. Mol. Sci. 2024, 25, 1985. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25041985
AMA Style
Suzuki Y, Shimizu Y, Shiina T. ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice. International Journal of Molecular Sciences. 2024; 25(4):1985. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25041985
Chicago/Turabian StyleSuzuki, Yuji, Yasutake Shimizu, and Takahiko Shiina. 2024. "ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice" International Journal of Molecular Sciences 25, no. 4: 1985. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25041985
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.