
The Intriguing Role of Iron-Sulfur Clusters in the CIAPIN1 Protein Family


1
Magnetic Resonance Center CERM, University of Florence, Via Luigi Sacconi 6, 50019 Sesto Fiorentino, FI, Italy
2
Department of Chemistry ‘Ugo Schiff’, University of Florence, Via della Lastruccia 3, 50019 Sesto Fiorentino, FI, Italy
*
Author to whom correspondence should be addressed.
Inorganics 2022, 10(4), 52; https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics10040052
Submission received: 8 March 2022
/
Revised: 1 April 2022
/
Accepted: 6 April 2022
/
Published: 13 April 2022
(This article belongs to the Special Issue Assembly and Reactivity of Iron–Sulfur Clusters)
Abstract
:Iron-sulfur (Fe/S) clusters are protein cofactors that play a crucial role in essential cellular functions. Their ability to rapidly exchange electrons with several redox active acceptors makes them an efficient system for fulfilling diverse cellular needs. They include the formation of a relay for long-range electron transfer in enzymes, the biosynthesis of small molecules required for several metabolic pathways and the sensing of cellular levels of reactive oxygen or nitrogen species to activate appropriate cellular responses. An emerging family of iron-sulfur cluster binding proteins is CIAPIN1, which is characterized by a C-terminal domain of about 100 residues. This domain contains two highly conserved cysteine-rich motifs, which are both involved in Fe/S cluster binding. The CIAPIN1 proteins have been described so far to be involved in electron transfer pathways, providing electrons required for the biosynthesis of important protein cofactors, such as Fe/S clusters and the diferric-tyrosyl radical, as well as in the regulation of cell death. Here, we have first investigated the occurrence of CIAPIN1 proteins in different organisms spanning the entire tree of life. Then, we discussed the function of this family of proteins, focusing specifically on the role that the Fe/S clusters play. Finally, we describe the nature of the Fe/S clusters bound to CIAPIN1 proteins and which are the cellular pathways inserting the Fe/S clusters in the two cysteine-rich motifs.
1. Introduction
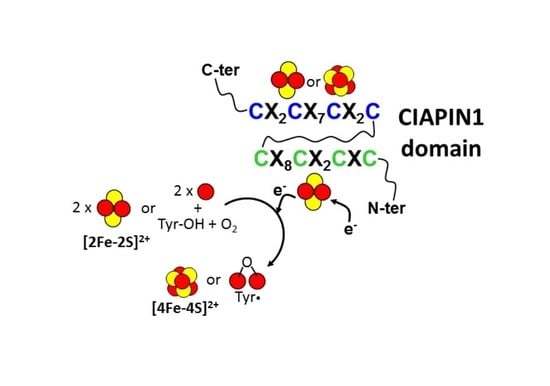
Iron-sulfur (Fe/S) clusters are cofactors composed of iron ions and inorganic sulfur (sulfide anions) that are often ligated to proteins through the sulfur atoms of cysteinyl ligands. The unusual chemical features of the Fe/S cluster enable it to facilitate reduction-oxidation reactions and to carry out numerous complex chemical reactions and sensing activities [1]. This explains their wide use in all three kingdoms of life by biological macromolecules to perform an enormous number of chemical reactions that are essential for life [2]. The most common Fe/S clusters include a rhomboid cluster composed of two iron atoms and two inorganic sulfur atoms ([2Fe-2S]) and a cubane form composed of four iron and four inorganic sulfur atoms ([4Fe-4S]) (Figure 1) [1]. These clusters can be easily obtained in vitro by mixing iron(II)/(III) and sulfide salts in reducing and anaerobic conditions [3]. However, the biosynthesis of these clusters is a complicated, multistep process involving many proteins both in prokaryotes and eukaryotes (recently reviewed in [4,5]). These proteins, which often are Fe/S cluster binding proteins themselves, are indispensable for life, and their absence or dysfunction causes growth defects, diseases or lethality [6,7]. This is a consequence of the fact that these proteins handle the in vivo biosynthesis of all cellular Fe/S clusters as well as their integration into proteins/enzymes via the formation of complex proteinaceous machineries [8]. In prokaryotes, three Fe/S protein biogenesis machineries are known (ISC, SUF, and the more specialized NIF) [9,10,11,12]. The former two systems have been transferred by endosymbiosis from bacteria to mitochondria and plastids of eukaryotes, respectively. In the cytosol, eukaryotes use the cytosolic iron-sulfur protein assembly (CIA) machinery for the biogenesis of cytosolic and nuclear Fe/S proteins [13]. A component of the CIA machinery, that is, the cytosolic CIAPIN1 protein family, is the argument of this review. In the CIA machinery, CIAPIN1 proteins have been shown to play a role in the biosynthesis of cytosolic-nuclear [4Fe-4S] clusters [14,15,16]. The CIAPIN1 protein family is typically characterized by a C-terminal domain (named CIAPIN1) that contains two highly conserved cysteine-rich motifs (CX8CX2CXC and CX2CX7CX2C, where X indicates any amino acid), which are both involved in Fe/S cluster binding. The human protein anamorsin was the first characterized member of the CIAPIN1 protein family and was discovered as an anti-apoptotic protein [17]. The role that the Fe/S clusters bound to the CIAPIN1 proteins play in the CIA machinery have received great interest in recent years, but still many questions remain unresolved. In addition, the Fe/S clusters bound to CIAPIN1 proteins have been shown to play a role in other cellular processes regardless of the CIA machinery, opening a new scenario for the functional role of this family of proteins. Indeed, the CIAPIN1 protein in yeast has been shown to have a role in the biosynthesis of diferric-tyrosyl radical cofactor as well as in the regulation of cell death. This review provides a complete overview of the role that the Fe/S clusters have in the CIAPIN1 protein family, taking into consideration functional, biochemical, structural and bioinorganic aspects.
2. Sequence Analysis of the CIAPIN1 Protein Family
The human anamorsin and the S. cerevisiae Dre2 are the two most studied proteins of the CIAPIN1 protein family and were thus used as starting sequences in our bioinformatic analysis. According to the Pfam database, human anamorsin is formed by two domains, an N-terminal Methyltransf_11 domain and a C-terminal CIAPIN1 domain, and the S. cerevisiae DRE2 protein is formed by an N-terminal DRE2_N domain and a C-terminal CIAPIN1 domain. In both these proteins, the N-terminal domains are structured and globular [18,19], whereas the CIAPIN1 domain is largely disordered, with an alpha-helix in the region connecting the two Fe/S cluster binding motifs [20] that characterize this protein family. Since the latter two motifs are both included in the CIAPIN1 domain, we mapped the sequences of this domain onto the tree domains of life to better understand the distribution of homologs and the origin of the family. It resulted that the CIAPIN1 domain is never found in bacterial and archeal proteins but it is widespread in eukaryotes. In particular, according to the Pfam database, the CIAPIN1 domain is present in 1858 proteins that map to 1510 eukaryotes including six different clades (Table 1). The majority of the organisms in each clade (Table 1) contain only one CIAPIN1 protein with the exception of the Archaeplastida clade whose organisms preferentially contain more than one CIAPIN1 protein. An analysis of the domain architectures of the proteins in each clade (Table 1) shows the presence of three possible domain organizations: (i) a sequence contains just one C-terminal CIAPIN1 domain without any additional domain (Ciapin1_only); (ii) a sequence contains an N-terminal DRE2_N domain and a C-terminal CIAPIN1 domain (Dre2_Ciapin1), as found in the S. cerevisiae Dre2 and (iii) a sequence contains an N-terminal Methyltransf_11 domain and a C-terminal CIAPIN1 domain (Methyltransf11_Ciapin1), as found in the human anamorsin. The Ciapin1_only architecture is present in all the clades whereas all three architectures are only present in Amorphea (Table 1). In particular, Dre2_Ciapin1 architecture is exclusively present in the fungi and specifically in the most evolved subkingdom of fungi, i.e., Dikaria. A similar trend is observed for the Methyltransf11_Ciapin1 architecture, which is indeed present in the more evolved Holozoa (in particular in Eumetazoa, i.e., the animal kingdom [21]) and Embryophyta (the plantae kingdom [22]). In other words, Ciapin1_only architecture is present in all clades and, within each clade, is present from the less evolved to the most evolved families of organisms. On the contrary, Methyltransf11_Ciapin1 is present only in two clades and, within these clades, only among the most evolved organisms. The same holds for Dre2_Ciapin1 architecture that is found only in the “higher fungi” [21,22]. This analysis suggests that the Ciapin1_only is the lowest common ancestor of the CIAPIN1 protein family to which the N-terminal DRE2_N domain in fungi and the N-terminal Methyltransf_11 domain in animals and plantae have been later added in the evolution line. This interpretation might indicate that a new function/pathway for the CIAPIN1 protein family is introduced through the addition of the N-terminal domain in the most evolved organism.
3. Functional Role of the CIAPIN1 Protein Family
The CIAPIN1 protein family has been described to be involved in three cellular processes that are: (i) the maturation of cytosolic-nuclear [4Fe-4S] proteins, (ii) the biosynthesis of diferric-tyrosyl radical cofactor (Fe(III)2-Y•) in the ribonucleotide reductase and (iii) the regulation of cell death (Figure 2).
3.1. The Maturation of Cytosolic and Nuclear [4Fe-4S] Proteins
The role that the CIAPIN1 protein family plays in the biogenesis of cytosolic-nuclear [4Fe-4S] proteins has been mainly characterized for the S. cerevisiae yeast member of the family, Dre2. Dre2 mutant or Dre2-depleted cells were shown to be deficient in cytosolic-nuclear [4Fe-4S] cluster protein activities, as well as in 55Fe incorporation into proteins carrying stable [4Fe-4S] clusters. On the contrary, the maturation of cytosolic [2Fe-2S] proteins, such as NEET proteins [23] and biotin synthase [15], as well as of mitochondrial Fe/S clusters [14,15], is not affected by Dre2 mutant or Dre2-depleted cells. Specifically, the cytosolic-nuclear [4Fe-4S] cluster proteins isopropylmalate isomerase (Leu1), sulfite reductase, RNase L inhibitor RLI1, nuclear glycosylase Ntg2 and IRP1 (the human cytosolic aconitase expressed in yeast cytoplasm) were markedly compromised in both Dre2 mutant and Dre2-depleted cells and 55Fe incorporation into some of these [4Fe-4S] proteins showed a strong decrease. These phenotypes of the Dre2 mutant or Dre2-depleted cells resemble those that are found for other components of the CIA machinery [24,25], which are: Fe/S cluster-containing proteins outside mitochondria were deficient, whereas Fe/S clusters inside mitochondria (aconitase and succinate dehydrogenase as an example) were maintained or in some cases increased. On this basis, it has been concluded that Dre2 is a component of the CIA machinery. Thus, Dre2 needs to be localized in the cytosol to perform its function in the CIA machinery. In agreement with this view, although first reports showed that Dre2 was found in the cytoplasm and partially in the mitochondrial intermembrane space [14,19], it has been recently shown that Dre2 is exclusively a cytosolic protein, with a large fraction of Dre2 bound to the cytosolic side of the outer membrane of mitochondria [26]. It has been also demonstrated that the human Dre2 homolog (called anamorsin) was able to complement the nonviability of a ΔDre2 deletion strain, indicating that the anamorsin is able to substitute Dre2 in the yeast cytosol [14].
The function of Dre2 in the maturation of cystosolic-nuclear [4Fe-4S] protein has been shown to strictly depend on the formation of a complex with the yeast NADPH-dependent diflavin oxidoreductase 1 (Tah18, hereafter) [15], whose physical interaction with Dre2 was identified by immunoprecipitation [27]. According to this model, Tah18 was shown to have the same cellular effects of Dre2 on cytosolic-nuclear [4Fe-4S] proteins upon its depletion, supporting that Tah18 complexed with Dre2 is required for the [4Fe-4S] cluster assembly on cytosolic-nuclear apo proteins [15]. It has also been shown that Tah18 and Dre2 are involved in an early stage of Fe/S protein maturation [15]. The interaction between Tah18 and Dre2 has been deeply investigated and it has been shown that Dre2 C-terminus, but not the N-terminus, interacts in vivo and in vitro with Tah18 [27,28]. It has also been shown that the closest homolog of yeast Tah18 encoded in the human genome, the NADPH-dependent diflavin oxidoreductase 1 (Ndor1, hereafter), can functionally replace Tah18, and that anamorsin and Ndor1 form a stable complex in yeast cells [15], similar to that formed by yeast Dre2 and Tah18. This complex between Ndor1 and anamorsin was also shown to exist in mammalian cells [29]. Taken together, these data indicate that the human Ndor1-anamorsin complex is the functional counterpart of the yeast Tah18-Dre2 complex.
In summary, the available studies define the Tah18-Dre2 complex as an early component of the CIA machinery dedicated to the [4Fe-4S] cluster assembly (Figure 2). The data also showed that this fundamental cellular task of the Tah18-Dre2 complex has been functionally conserved in evolution from yeast to humans and performed in human cells by the Ndor1-anamorsin complex.
3.2. The Diferric-Tyrosyl Radical Cofactor Biosynthesis in Ribonucleotide Reductase
Eukaryotic ribonucleotide reductases (RNRs) require a diferric-tyrosyl radical (Fe(III)2-Y•) cofactor to produce deoxynucleotides essential for DNA replication and repair [30,31,32]. The CIAPIN1 protein family takes part in this cellular process, although so far this involvement has been exclusively demonstrated in yeast. It has been shown indeed that the depletion of Dre2 in S. cerevisiae cells leads to a decrease in both Tyr(•) levels and RNR activity [33,34], strongly suggesting that Dre2 serves in the assembly of the Fe(III)2-Y• cofactor in RNR (Figure 2). In agreement with this model, cells with diminishing Dre2 have significantly reduced the ability to make deoxynucleotides [34].
It has also been demonstrated that Tah18, the protein partner of Dre2, also plays a critical role in the formation of the Fe(III)2-Y• cofactor in RNR. Specifically, it was found that the kinetics and levels of in vivo reconstitution of the RNR cofactor are severely impaired in two Tah18 mutants [34]. These data support a model where the conserved protein complex composed of the Fe/S-requiring Dre2 and the diflavin-requiring Tah18, which are required for the [4Fe-4S] cluster assembly of proteins found in the cytosol and nucleus, is also required for the RNR cofactor assembly (Figure 2). Deficiency in this complex has also been shown to lead to the activation of both the DNA-damage checkpoint and the iron regulon, linking iron homeostasis to the maintenance of genome stability [34]. Finally, it has been found that Dre2 is required for RNR cofactor formation but appears to be dispensable for iron loading [35].
In summary, the Tah18-Dre2 complex is required to assemble the Fe(III)2-Y• cofactor of RNR (Figure 2), very likely exploiting a molecular function that is similar for both assemblies of the Fe(III)2-Y• cofactor in RNRs and of [4Fe-4S] clusters of cytosolic and nuclear proteins.
3.3. The Dre2-Tah18 Dependent Regulation of Cell Death
The Tah18-Dre2 complex has been shown to play a regulatory role in yeast cells death in response to high doses of hydrogen peroxide [27]. Indeed, although Dre2 has been identified as a molecular partner of Tah18 in the absence of oxidative stress, after cell exposure to lethal doses of H2O2, the Tah18-Dre2 complex dissociates and Tah18, but not Dre2, relocalizes to the mitochondria and controls mitochondria integrity and cell death (Figure 2). On the contrary, the promotion of Dre2-Tah18 interaction reduces H2O2-induced cell death and Tah18 localization to mitochondria. It has also been shown that only the C-terminus of Dre2 is essential for cell survival while the N-terminus of Dre2, which does not interact with Tah18, is dispensable for survival [28]. From these studies, it was concluded that Tah18-Dre2 interaction in vivo is essential for yeast viability and the existence of a mitochondria-dependent cell death program in yeast involving Tah18 was proposed. In this regard, it has been shown that Tah18 is involved in NO synthesis in yeast, that NO synthase (NOS)-like activity requiring Tah18 induces cell death upon treatment with H2O2 and that Tah18-dependent NO production and cell death are suppressed by enhancement of the interaction between Tah18 and its molecular partner Dre2 [36]. On this basis, it has been proposed that the Tah18-Dre2 complex regulates cell death as a molecular switch via the control of NO production in response to environmental oxidative conditions (Figure 2). In particular, it is possible that, only once Tah18 is translocated to the mitochondria, it activates the cell death pathway dependent on NOS-like activity (Figure 2). A cell death regulation mechanism similar to that observed for the Dre2-Tah18 complex in yeast might be conserved in human cells. Indeed, anamorsin is able to restore yeast viability in the place of Dre2 [14,27] and physically interacts with Tah18 [27]. This view is reinforced by the fact that anamorsin has been identified as an anti-apoptotic factor in human cells [17], thus indicating that anamorsin and Dre2 share this common function.
In summary, the available experimental data support the existence of an oxidative stress-induced cell death pathway in yeast, and possibly also in humans, which is activated in yeast by the Tah18-dependent production of NO, a molecule promoting apoptosis-like cell death (Figure 2) [37]. This cell death pathway is controlled by the Dre2-Tah18 complex (and possibly by anamorsin-Ndor1 complex in human cells) and involves mitochondria (Figure 2).
4. Molecular Function of the Iron-Sulfur Cluster Bound to the CX8CX2CXC Motif in the CIAPIN1 Protein Family
We described in the previous section that all the three cellular processes involving members of the CIAPIN1 protein family depend on the formation of a heterocomplex containing a diflavin reductase. The latter protein tightly binds FAD and FMN cofactors in two domains and catalyzes the transfer of the reducing equivalents from the two-electron donor NADPH to a variety of one-electron acceptors [38], such as an Fe/S cluster. Thus, this suggests that the heterocomplex is able to create an electron transfer pathway working like a reducing system to provide electrons to several potential targets. According to this model, it has been shown in yeast that the diflavin reductase Tah18 functionally interacts in vivo with a Fe/S form of Dre2 [15], thus supporting that the redox center physiologically accepts the electrons from a flavin moiety of Tah18 is a Fe/S center of Dre2. In vitro studies based on UV-visible and EPR spectroscopy showed indeed that Tah18 can transfer electrons from NADPH to an Fe/S cluster of Dre2 [15]. In particular, it has been observed that the reduction of Dre2 by NADPH-Tah18 was observed for only one of the two Fe/S clusters that Dre2 is able to bind into the two distinct binding motifs, that is the one bound to the CX8CX2CXC motif of the CIAPIN1 domain (redox-active Fe/S cluster, hereafter). The same conclusions have been drawn for the human Ndor1-anamorsin complex. Indeed, 55Fe coprecipitation data are consistent with the flavoprotein human Ndor1 to bind to an Fe/S form of anamorsin [15], as well as only the Fe/S cluster bound to the CX8CX2CXC motif of the CIAPIN1 domain of anamorsin is reduced by FMN cofactor of Ndor1 [39].
4.1. CIAPIN1-Dependent Electron Transfer in the Assembly of [4Fe-4S] Clusters
The electrons provided by the Tah18-Dre2 complex have been shown to be essential for assembling [4Fe-4S] clusters [15], which are then inserted into cytosolic-nuclear Fe/S cluster binding proteins through a complex protein machinery [40]. Considering that the formation of the [4Fe-4S]2+ cluster requires a reductive coupling of two [2Fe-2S]2+ clusters [41,42,43,44,45,46], it can be proposed that the complex formed by the Fe/S form of Dre2 (or anamorsin) and the diflavin reductase Tah18 (or Ndor1) provides the two electrons required to couple two [2Fe-2S]2+ clusters into a [4Fe-4S]2+ cluster. However, the mechanism of this step of the CIA machinery is still largely unknown and needs further investigation. In particular, whether the other Fe/S cluster bound to the CX2CX7CX2C motif of the CIAPIN1 domain plays a role in this electron transfer process is still unknown. No information is indeed available on the molecular function of the latter Fe/S cluster, whose nuclearity, i.e., [2Fe-2S] vs. [4Fe-4S], has also been the subject of an intense debate in recent years (see later).
The molecular basis for the complex formation and the electron transfer process have been investigated in detail in the case of Ndor1-anamorsin [20]. It has been found that the two proteins form a stable complex where one electron is transferred from the hydroquinone state of the FMN moiety (FMNH2, hereafter), bound to the FMN domain of Ndor1, to the oxidized state of the redox-active Fe/S cluster of anamorsin (Figure 3). Reduced FMNH2 is obtained through sequential electron transfer events that include the NADPH binding to the FAD domain of the diflavin reductase, the reduction of FAD moiety by NADPH, and finally the transfer of the electrons from reduced FAD to FMN (Figure 3). The stable complex is achieved thanks to a specific protein–protein recognition between a completely unstructured region of anamorsin, which is part of the linker separating the N-terminal domain from the C-terminal domain, and an α-helical face of the FMN-binding domain of Ndor1 (Figure 3). This molecular recognition is governed by hydrophobic and complementary charged interacting residues. On the contrary, the N-terminal domain of anamorsin is not involved in the recognition process (Figure 3). This result is consistent with the absence of any interaction between the N-terminal domain of Dre2 and Tah18, as monitored both in vitro and in vivo [28]. The formation of the stable complex is independent from the presence of the Fe/S centers and from the redox state of the FMN moiety, indicating that the two protein partners do interact permanently and no dissociation occurs along the electron transfer process [20]. Thus, the complex formation is not affected by the redox centers and is stabilized by residues far from the two redox centers. The same behavior was reported for the in vitro interaction of the yeast homologs, where apo-Dre2 showed a stable interaction with Tah18 [28]. The two redox centers (i.e. FMN and the redox active Fe/S cluster), on the other hand, transiently interact via electrostatic interactions established between a negatively charged region surrounding the FMN moiety and a positively charged region surrounding the redox-active Fe/S cluster. All these observations lead to a protein–protein recognition that is based on these molecular aspects: (a) the two proteins form a stable complex thanks to specific interactions involving regions far from the two FMN and Fe/S redox-active cofactors; (b) the areas surrounding the two FMN and Fe/S redox-active centers transiently and weakly interact with each other; and (c) the unstructured region comprising residues 185–223 of anamorsin is essential to stably form the complex and might contribute to positioning the C-terminal CIAPIN1 domain of anamorsin containing the redox-active Fe/S cluster in those orientations that allow the reception of the electron(s) from the hydroquinone state of the FMN redox center. Taking also into consideration that, in diflavin reductases, the electron transfer pathway from NADPH to FMN through the mediation of FAD is coupled with large conformational changes [47,48], we can propose the following molecular model of the electron transfer pathway occurring in the anamorsin-Ndor1 complex (Figure 3): (i) anamorsin is stably bound to both closed and open conformations of Ndor1 due to a specific recognition between an unstructured region of anamorsin and a region of the FMN-binding domain that is solvent exposed in both open and closed conformations of Ndor1; (ii) upon NADPH binding by Ndor1, electrons can be efficiently transferred within the closed conformation of Ndor1 to produce FMNH2; (iii) NADP formation and interflavin electron transfer within Ndor1 significantly populate the open conformation, which allows the formation of the transient interaction between the FMN and the redox-active Fe/S cluster regions; (iv) the latter interaction allows efficient transfer of one electron from FMNH2 to the redox-active Fe/S cluster; and (v) the received electron is finally transferred to an acceptor, such as [2Fe-2S]2+ clusters to obtain reduced [2Fe-2S]+ clusters, which can eventually, with two of them, generate a [4Fe-4S]2+ cluster.
In summary, biochemical, structural and molecular cell observations indicate that the essential function that the Dre2/Tah18 or anamorsin/Ndor1 complex plays in the CIA machinery is determined by the generation of the electron transfer pathway that terminates in the reduction of the redox-active Fe/S cluster of Dre2/anamorsin, which can thus drive the formation of [4Fe-4S] clusters for cytosolic-nuclear proteins.
4.2. CIAPIN1-Dependent Electron Transfer in the Assembly of the Fe(III)2-Y• Cofactor
As discussed before, in S. cerevisiae, the Dre2-Tah18 complex also plays a critical role in the formation of the Fe(III)2-Y• cofactor in RNR. Since Dre2 and the β subunit of RNR can exist in the same protein complex in vivo [34,49], this is consistent with a model where the redox-active Fe/S cluster of Dre2, once reduced by Tah18, is involved in delivering to the β subunit of RNR the electron that is required for the Fe(III)2-Y• cofactor assembly (Figure 3). Moreover, Dre2 interacts in vivo with another component [49] involved in the assembly of the Fe(III)2-Y• cofactor, which is the cytosolic monothiol glutaredoxin-3 (Grx3, hereafter) [33,35]. The latter, once it forms a heterodimer with the highly homologous protein Grx4 by sharing with it a [2Fe-2S]2+ cluster at the subunit interface [50], was proposed to play an important role in the delivery of iron to Fe/S, heme, and di-iron requiring proteins, including RNR [33,51]. On this basis, it was first postulated that Dre2-Tah18 complex might provide the reducing equivalents to allow Fe2+ ion transfer from the reduced [2Fe(III,II)-2S]+ cluster bound at the Grx3/4 dimer interface to apo-ββ′ heterodimer of RNR [34]. However, it was recently shown that, although Dre2 is essential for the assembly of the active Fe(III)2-Y• cofactor of RNR, it is dispensable for the first iron-loading step in RNR cofactor assembly [35]. In summary, we believe that in vitro studies are strongly required to shed light on whether the iron ion of the [2Fe-2S] cluster bound to Grx3/4 heterodimers is transferred to RNR as well as which is the species receiving, from the Dre2-Tah18 complex, the electron that is required for the assembly of the Fe(III)2-Y• cofactor of RNR.
4.3. A Possible Role of CIAPIN1-Dependent Electron Transfer in the Yeast Viability
This electron flow driven by the Dre2-Tah18 complex has also been proposed to be essential for the yeast viability [27]. It is possible indeed that the disruption of the Dre2-Tah18 complex in vivo in the presence of high levels of hydrogen peroxide not only allows Tah18 to relocalize to the mitochondria promoting cell death, but also provokes the interruption of the electron flow between the two proteins, thus abolishing its essential function in the CIA machinery. In summary, this suggests that Dre2-Tah18 complex activity might be regulated by moderate intracellular redox potential changes.
4.4. A Possible New Functional Role of the Reducing System Ndor1-Anamorsin
Recent studies have shown that the Ndor1-anamorsin complex can interact in vitro with the mitochondrial [2Fe-2S] protein mitoNEET forming a transient, low-populated protein–protein complex, which is able to convert the oxidized [2Fe-2S]2+ clusters of mitoNEET into reduced [2Fe-2S]+ clusters [52]. In particular, it was shown that the redox-active Fe/S cluster of anamorsin, once reduced by FMNH2 bound to the FMN-binding domain of Ndor1, is able to work as an electron donor towards mitoNEET, specifically reducing its [2Fe-2S] clusters (Figure 3).
mitoNEET is a small protein localized on the cytosolic face of the outer mitochondrial membrane, being anchored through a single transmembrane helix [53,54]. In this environment, mitoNEET forms dimers with one [2Fe-2S] cluster per monomer. Various studies have shown that mitoNEET displays high affinity for the reduced [2Fe-2S]+ state of the cluster, while upon oxidation the cluster is more labile [55,56,57]. In-cell EPR studies showed that the [2Fe-2S] clusters of mitoNEET are in the reduced [2Fe-2S]+ state when expressed in Escherichia coli cells under normal growth conditions, and on this basis it was assumed to be reduced in unstressed mammalian cells [58]. This fully agrees with the high midpoint potential of the mitoNEET clusters, which is approximatively 0 mV at pH 7.0 [59,60]. Recently, it has been proposed that mitoNEET plays a specific role in the cytosolic Fe/S cluster repair of cytosolic iron regulatory protein 1 (IRP1) in oxidatively stressed cells [23]. This pathway reactivates the cytosolic apo-IRP1 into Fe/S loaded, active aconitase after H2O2-induced oxidative and nitrosative stress, independently of the CIA pathway, which, on the contrary, matures aconitase under normal cellular conditions [13,61]. It was proposed that oxidative stress conditions induce oxidation of the mitoNEET [2Fe-2S] cluster, without its disruption, triggering the [2Fe-2S]2+ transfer to IRP1 and transforming it into active aconitase [23,62]. Hence, mitoNEET may directly control its [2Fe-2S] cluster transfer activity via a redox switch of its [2Fe-2S] cluster, in such a way that indirectly regulates the IRP1-dependent iron homeostasis [63]. IRP1, indeed, is a central component in controlling intracellular iron homeostasis [64]. In this model, an electron source, which is still undefined, is required in order to allow the formation of the [4Fe-4S]2+ cluster on IRP1 via a reductive coupling of the two [2Fe-2S]2+ clusters donated by mitoNEET.
The Ndor1-anamorsin complex, by reducing the mitoNEET cluster, might act as a regulator of the termination of the cluster transfer pathway from mitoNEET to IRP1. Once oxidative stress is not present anymore, the reducing properties of the Ndor1-anamorsin complex is reactivated in the cytoplasm and can convert mitoNEET back into its reduced [2Fe-2S]+ form. In this way, the Ndor1-anamorsin complex can stop the mitoNEET cluster transfer pathway to repair IRP1, being indeed the repair function of mitoNEET no longer needed in unstressed cells. This potential new functional role of the Ndor1-anamorsin complex, however, is still not verified at the cellular level and thus needs validation.
5. Spectroscopic Investigations to Unravel the Nature of the Iron-Sulfur Clusters Bound to CIAPIN1 Proteins
The nature of the Fe/S clusters bound to CIAPIN1 proteins has been the subject of an extensive debate in the last few years. The first evidence showing that the CIAPIN1 protein family is an Fe/S cluster binding protein was reported when yeast Dre2 was purified from E. coli cells as recombinant protein [14]. It was observed that the purified protein contains iron and sulfur with a roughly 1:1 stoichiometry per one protein molecule, when the cell culture was supplemented with 125 μM of ferric-nitrilotriacetate [14,65]. This observation indicated an incomplete Fe/S cluster loading with respect to the expected binding of two Fe/S clusters bound in the two conserved Cys-rich motifs of Dre2. UV-visible acquired on the purified protein identified the presence of an oxidized [2Fe-2S]2+ cluster, which can be reduced by dithionite to form the reduced [2Fe-2S]+ cluster, as shown by EPR spectra at temperatures higher than 30 K [14,65,66]. Once registered below 20 K, EPR spectra of the purified Dre2 protein reduced by dithionite also showed signals characteristic of a [4Fe-4S]+ cluster [14,65,66]. In addition, EPR spectra at 6 K showed some spin–spin interaction between the two Fe/S clusters possibly due to their short distance [65]. The zero-field Mössbauer spectrum of as-purified wild-type Dre2 confirmed the presence of two species one accounting for an oxidized [4Fe-4S]2+ cluster and the other for a [2Fe-2S]2+ cluster [66], in line with the EPR studies. Upon reconstitution of Dre2, the content of iron and sulfide per protein was shown to increase up to ~6 and the UV-visible and EPR spectra were interpreted as the sum of one [2Fe-2S] cluster and one [4Fe-4S] cluster [15]. Plant and Trypanosoma brucei homologues of yeast Dre2 have also been purified in the presence of a [2Fe-2S] cluster containing a small subpopulation of a [4Fe-4S] cluster [16,67].
Two recent studies deeply investigated, through UV-visible, EPR and Mössbauer spectroscopies, the Fe/S cluster-coordinating residues bound to yeast Dre2 exploiting several Cys-to-Ala mutants [65,66]. Specifically, one or two Cys residues were removed in each of the two conserved Cys-rich motifs of Dre2, named hereafter motif I mutants for the mutated cysteines of the redox-active Fe/S cluster site and motif II mutants for the mutated cysteines of the other Fe/S cluster site at the C-terminus. In these two studies, anaerobic conditions were strictly maintained, performing cell lysis and a purification procedure in a glove-box with all degassed buffers [65,66]. The resulting data were interpreted by proposing that motif I mutants of Dre2 show spectra typical of [4Fe-4S]2+ proteins, indicating that a [4Fe-4S]2+ cluster is bound at motif II, while purified motif II mutants of Dre2 show spectra typical of a [2Fe-2S]2+ cluster, indicating that a [2Fe-2S]2+ cluster is bound at motif I (Figure 4). Finally, these studies showed that the exposure of anaerobically purified wild-type or motif I mutant Dre2 proteins to air suggested the presence of an oxygen-labile [4Fe-4S]2+ cluster bound to motif II of Dre2. Specifically, UV-visible and EPR data indicated the [4Fe-4S] cluster bound to motif II undergoes some cluster breakdown to a [2Fe-2S]2+ form. A relevant aspect, which was not taken into consideration in the interpretation of the data presented in both studies, concerns the fact that single or double Cys-to-Ala mutants of one motif cannot exclude that the remaining two or three cysteines of this mutated motif might contribute to the binding of the Fe/S cluster in the other not-mutated motif. This scheme of Fe/S cluster binding cannot be neglected for these mutants as it might occur considering that the C-terminal segment containing the two motifs is largely unstructured and flexible and the two Fe/S cluster binding sites are close to each other [65]. Quadruple mutants where all the four cysteines of each motif were mutated would prevent this potential effect in such a way to more properly describe Fe/S cluster coordination properties on both Dre2 motifs.
A further remark concerns the different results found on the [2Fe-2S] vs. [4Fe-4S] cluster content in the two studies described above [65,66]. Indeed, in one study it was shown that the content of the [4Fe-4S] cluster in purified wild-type Dre2 was about 10% that of the [2Fe-2S] cluster and the [2Fe-2S] cluster was always predominant [65]. On the contrary, in the other study [66], zero-field Mössbauer spectrum of an isolated wild-type Dre2 indicated a stoichiometry of 0.25 [2Fe-2S]2+ clusters per [4Fe-4S]2+ cluster calculated for Dre2, and it was reported that this was a consequence of the fact that the cluster coordinated by motif I is a labile [2Fe-2S]2+/1+ type. This discrepancy between the two studies might be due to the high oxygen sensitivity of both [2Fe-2S] and [4Fe-4S] clusters in Dre2, which provides different results depending on the protein isolation protocol that has been applied in the two studies. However, it is still unclear why, in one study, the oxygen sensitivity is observed to largely affect the [2Fe-2S] cluster while, in the other study, it affects the [4Fe-4S] cluster.
An in vivo approach on yeast Dre2 was also exploited to investigate the Fe/S cluster-coordinating residues, showing that exchanges of seven of the eight cysteine residues to alanine lead to a loss of cell viability without affecting protein expression or stability, indicating the essentiality of these residues for yeast Dre2 function [65,66]. This was accompanied by a dramatic decrease of iron incorporation into the cytosolic [4Fe-4S] protein isopropylmalate dehydratase (Leu1) and low enzymatic activity, directly demonstrating the essential function of the seven residues for [4Fe-4S] cluster insertion into Leu1 [66]. Moreover, an almost complete loss of iron binding to any of these seven Dre2 mutants in vivo was observed [66]. This indicates the essential function of the seven cysteines for Fe/S cluster coordination in vivo as well as a mutual dependence of Fe/S cluster binding at the two motifs I and II, i.e., a lack of Fe/S cluster binding to one motif negatively impacts on the stable Fe/S cluster association to the other. Only the first cysteine of motif I, although its mutation elicited a substantial decrease in the in vivo Fe/S cluster binding, is not essential for Dre2 function as a CIA component [65,66]. These results convincingly showed that both Fe/S clusters of Dre2 perform indispensable functions in cytosolic Fe/S protein assembly and that the lethal phenotypes resulting from the mutations are caused by a loss of function of both Fe/S clusters.
The Fe/S cluster binding properties of human anamorsin were deeply investigated by our laboratory. Preliminary UV-visible, EPR and NMR data on anaerobically purified anamorsin, prepared as described in [19], showed that the protein coordinates only one [2Fe-2S]2+ cluster per protein molecule when the cell culture was supplemented with 125 μM of FeCl3, with no evidence of a [4Fe-4S] bound cluster, at a variance of Dre2 [19]. A more detailed spectroscopic EPR and Mössbauer study based on quadruple mutants of each motif showed that: (i) each motif in human anamorsin is able to bind a [2Fe-2S] cluster independently through its four cysteine residues (Figure 4); (ii) the two reduced [2Fe-2S]+ clusters exhibit a unique electronic structure with considerable anisotropy in their coordination environment, different from that observed in reduced, plant-type and vertebrate-type [2Fe-2S] ferredoxin centers; (iii) the reduced cluster bound to motif II reveals an unprecedented valence localization-to-delocalization transition as a function of temperature [39]. The unique electronic properties of both [2Fe-2S] centers can be interpreted by considering that both cysteine-rich motifs are located in a highly unstructured and flexible protein region, whose local conformational heterogeneity can induce anisotropy in metal coordination. In the EPR study by Zhang et al. [65] it was, however, reported that human anamorsin contains a [2Fe-2S] cluster and a [4Fe-4S] cluster, as observed in yeast Dre2. Zhang et al. interpreted the difference between our previous study and its study because of the high oxygen sensitivity of the [4Fe-4S] cluster in anamorsin, which resulted in the formation of a [2Fe-2S] cluster at both sites as observed by us. Very recently, this issue has been solved by us via an in cellulo spectroscopic approach [68] that prevented the possible problem of the oxygen Fe/S cluster sensitivity occurring in vitro during the protein purification procedure. We showed by in cellulo Mössbauer and in cellulo EPR spectroscopies that human anamorsin coordinates only [2Fe-2S] clusters, one per each motif [68]. In addition, EPR spectra, acquired on both reduced E. coli cells expressing WT- and Cys-mutated anamorsin as well as anaerobically purified, reduced WT- and Cys-mutated anamorsin, prepared performing a purification procedure in the glove-box with all degassed buffers [39,68], showed that the [2Fe-2S] cluster bound to motif II of anamorsin displays enhanced electron spin relaxation rates, likely originating from local protein conformational heterogeneity, thus confirming our previous study. The electron spin relaxation properties of the reduced [2Fe-2S]+ cluster bound to motif II of anamorsin are, indeed, peculiar with respect to what is usually observed in ferredoxin-type reduced [2Fe-2S]+ clusters, resembling in part those of fast relaxing reduced [4Fe-4S]+ clusters. Indeed, in ferredoxin-type reduced [2Fe-2S]+ clusters, the slow spin relaxation rates make the EPR signal hardly detectable below 10 K [69], contrary to what was observed for the reduced [2Fe-2S]+ cluster bound to motif II of anamorsin, whose EPR signal was still well detectable at 5 K. These peculiar relaxation properties of the reduced [2Fe-2S]+ cluster bound to motif II were explained considering that the reduced cluster bound to this motif revealed a valence localization-to-delocalization transition as a function of temperature [39]. It has been previously demonstrated that the electron delocalization within mixed-valence FeIIFeIII pairs favors the parallel alignment of the local spins of both the high-spin FeII (SFeII = 2) and high-spin FeIII (SFeIII =5/2) ions, leading to a S = 9/2 total spin ground state [70]. The detection of the EPR signal of a S = 1/2 spin suggests that the partial electron delocalization observed in the reduced [2Fe-2S]+ cluster bound to motif II is not strong enough to make the maximal total spin S = 9/2 value as the ground state. However, it could allow the lowering in energy of the excited S > 1/2 spin states, thus leading to a faster electron spin relaxation rate for the ground S = 1/2 state compared with those typically observed in ferredoxin-type reduced [2Fe-2S]+ clusters, thus more closely mimicking an electron spin relaxation rate value typical of fast relaxing [4Fe-4S]+ clusters. The EPR spectra of wild-type anamorsin reported by Zhang et al. reproduced the same ones reported in our work, and thus our interpretation is consistent with both in vivo and in vitro EPR data obtained by us and Zhang et al., definitively showing that anamorsin exclusively binds [2Fe-2S] clusters at both motifs.
In summary, the deep spectroscopic characterization of Fe/S cluster nature and binding mode on yeast Dre2 and human anamorsin showed two divergent results. While both systems agree that motif I exclusively binds a [2Fe-2S] cluster (Figure 4), which is the redox active site receiving electrons from Tah18/Ndor1, they diverge on the cluster nature bound to motif II. Dre2 was found indeed to bind a [4Fe-4S] cluster, while anamorsin binds a [2Fe-2S] cluster (Figure 4) with the peculiarity of showing enhanced electron spin relaxation rates with respect to ferredoxin-type reduced [2Fe-2S]+ clusters. Detailed structural models of these FeS cluster environments will be fundamental in the future to clarify the divergent results on Fe/S cluster binding of Dre2 vs. anamorsin as well as to provide information on the structural rearrangements occurring during the electron transfer process.
6. Iron-Sulfur Cluster Insertion into CIAPIN1 Proteins
In vivo data on yeast cells provided the first clues on the proteins required to insert Fe/S clusters into Dre2 [15]. It was found that the cysteine desulfurase Nfs1 present in the mitochondrial matrix is essential for Fe/S cluster assembly on Dre2, while no significant effects were observed upon the depletion of five CIA components required to assemble cytosolic [4Fe-4S] clusters, i.e., Cfd1, Nbp35, Nar1, Cia1 and Tah18. In particular, the slight increase of iron associated with Dre2 upon depletion of Tah18 shows that the Fe/S clusters on Dre2 can assemble without the input of electrons from NADPH-Tah18. The latter electron transfer chain was shown to be essential to assembling target [4Fe-4S] proteins. However, although Dre2 can be considered a target Fe/S protein since it stably binds its Fe/S clusters and because the latter were shown to not be transferred to Fe/S acceptor proteins, it unexpectedly does not follow this request of electrons to assemble the [4Fe-4S] cluster bound to motif II. Overall, the in vivo findings, indicating that no CIA [4Fe-4S] assembly components are required for Fe/S assembly on Dre2, would seem to contradict the in vitro observations that are in support of a [4Fe-4S] cluster bound to motif II of Dre2 [66]. Instead, this in vivo observation might suggest that another, still unknown, cytosolic pathway should be active to assemble the [4Fe-4S] cluster in Dre2.
6.1. The Possible Role of Monothiol Glutaredoxins Grx3/4 and Fra2 in Fe/S Cluster Insertion into Dre2
The only component of the CIA machinery whose cell depletion caused a Fe/S cluster assembly defect in Dre2 was the Grx4 protein [51]. Grx4 and its homologous protein partner Grx3 contain an amino-terminal thioredoxin-like domain followed by one glutaredoxin (Grx) domain (Figure 5A), and both proteins were found to interact with Dre2 [33,51]. Grx3/4 proteins belong to the monothiol CGFS glutaredoxin family, which typically coordinates a [2Fe-2S] cluster [71,72]. Grx3/4 interacts with itself and with Fra2 in vivo and in vitro, forming heterodimers bridged by a [2Fe-2S] cluster [50,51,73] (Figure 5A). The ligands for the [2Fe-2S] cluster in Grx3/4 heterodimer come from two cysteines of the CGFS motif of Grx3 and Grx4 and two glutathione (GSH) molecules [72,74], whereas in the Fra2-Grx3/4 heterodimer, ligands are provided by the same cysteine from Grx3 or Grx4, a histidine from Fra2, and a GSH molecule [75,76,77]. On the basis of recent findings showing that an Fe/S intermediate is exported from mitochondria to the cytosol to be utilized by the CIA machinery [78], it might be argued that the [2Fe-2S] cluster, once assembled in mitochondria by an Nfs1-containing complex [79], is then transferred to the Grx3/4 and Grx3/4-Fra2 heterodimers that might thus transfer the cluster to motif I of Dre2 (Figure 5A). In agreement with this model, the in vivo function of the Fra2-Grx3/4 heterodimers in yeast was that of delivering its [2Fe-2S] cluster to Aft1/2, two paralogous iron-responsive transcription factors (activators of ferrous transport) regulating iron homeostasis in Saccharomyces cerevisiae [80]. Thus, it might be possible that the same heterodimers can deliver a [2Fe-2S] cluster to Dre2, thus resulting, in this model, the CIAPIN1 domain of Dre2 occupied by a [2Fe-2S] cluster at the motif I only (Figure 5A). However, this Grx3/4-Fra2-dependent model of [2Fe-2S] cluster insertion into Dre2 is still hypothetical and requires experimental support, as well as how the [4Fe-4S] cluster is formed and inserted into motif II of Dre2 is also still unknown (Figure 5A).
6.2. The Role of Monothiol Glutaredoxin GLRX3 and BOLA2 in [2Fe-2S] Cluster Insertion into Anamorsin
The function of the human homologue of yeast Grx3/4, that is, glutaredoxin-3 (GLRX3, also commonly named PICOT), as a [2Fe-2S] cluster donor of anamorsin, was demonstrated by both in vitro and in vivo studies. Firstly, a yeast two-hybrid assay and affinity-capture MS screens showed that GLRX3 binds anamorsin in cells [81]. GLRX3 consists of three domains: one N-terminal thioredoxin (Trx) domain with no Trx-related enzymatic role and two Grx domains (GrxA and GrxB) each able to bind a glutathione-coordinated [2Fe-2S] cluster via protein dimerization [82] (Figure 5B). Protein–protein interaction and cluster transfer in vitro studies provided the first evidence that GLRX3 displays a functional role in cytosolic [2Fe-2S] cluster trafficking by transferring two [2Fe-2S] clusters to its protein partner anamorsin [83]. The transfer mechanism was observed to be dependent on the formation of a protein–protein complex between the N-terminal domains of GLRX3 and anamorsin (Figure 5B). Their interaction was indeed the fundamental requisite to observe [2Fe-2S] cluster transfer from GLRX3 to both motifs I and II of the CIAPIN1 domain of anamorsin (Figure 5B). This mechanism guarantees that two [2Fe-2S] clusters are concomitantly transferred in a single molecular event to anamorsin. NMR-driven biomolecular docking data also defined how the N-domains of the two proteins specifically recognize each other and identified a role of the linker of anamorsin in stabilizing the complex formation [83]. Considering that the linker is also the region of anamorsin tightly interacting with Ndor1 to form a specific and stable protein complex [20], we can suggest that, upon the interaction of the GLRX3-anamorsin complex with Ndor1, the linker might weaken its interaction with GLRX3 while promoting the interaction with Ndor1. In such a way, the stabilizing effect of the linker on the GLRX3–anamorsin interaction would be lost, and, as an outcome, the complex between GLRX3 and anamorsin might switch to the final target complex formed by anamorsin and Ndor1. The linker interaction might be therefore responsible for modulating the formation and the release of the protein–protein complexes required to obtain the mature redox-competent state of anamorsin that receives electrons from Ndor1.
The function of GLRX3 in transferring [2Fe-2S] clusters to anamorsin can nicely explain the in vivo data showing that human GLRX3 silencing causes a decrease of enzymatic activities of cytosolic [4Fe-4S] proteins [84]. This in vivo effect can be interpreted indeed because of an impairment of the GLRX3-dependent anamorsin maturation process, which therefore makes the CIA machinery unable to function in the maturation of cytosolic [4Fe-4S] proteins. This view also fits with the yeast system, which similarly showed a decrease of Fe/S cluster insertion into cytosolic [4Fe-4S] protein targets upon Grx4 depletion [51], as well as a defect in the di-iron cluster assembly of the cytosolic ribonucleotide reductase [33,35,51], which, similarly to the CIA machinery, depends on the Dre2-Tah18 electron transfer chain [34].
In addition to anamorsin, GLRX3 was found to have another protein partner that is BOLA2 [85], a cytosolic protein belonging to a conserved small protein family ubiquitously found in eukaryotes and prokaryotes [75,86]. While in yeast, cytosolic BolA (called Fra2) associates with cytosolic Grx3/4 and affects iron-dependent regulation of transcription factors through the transfer of [2Fe-2S] clusters [80], human BOLA2 must have a different function in the cytosol since these transcription factors are not present in mammalian cells. In vitro studies showed that GLRX3 and BOLA2 form a heterotrimeric complex that consists of one GLRX3 and two BOLA2 molecules that stably coordinate two bridging [2Fe-2S] clusters [86,87] (Figure 5B). Each cluster is shared by BOLA2 and a Grx domain of GLRX3 (Figure 5B), and it is coordinated by the conserved cysteine of the Grx domain, by a glutathione bound to the Grx domain and by a conserved histidine residue in BOLA2 [86]. A fourth ligand for the cluster has not been clearly identified. Protein–protein interaction studies also showed that each GrxA and GrxB domain of GLRX3 interact with a BOLA2 molecule, while the Trx-like domain of GLRX3 is not involved in such interaction in both apo and holo forms of the heterotrimeric complex [87]. The GLRX3-[2Fe-2S]2-BOLA22 complex has been shown to work as a [2Fe-2S] chaperone. In vitro studies showed, indeed, that the latter complex can transfer its [2Fe-2S] clusters to both Fe/S cluster sites of anamorsin [87], following the same cluster transfer mechanism previously described for [2Fe-2S]2 GLRX32 [83] (Figure 5B). In vivo studies also indicate that the GLRX3-BOLA2 complex in mammalian cells functions as a [2Fe-2S] cluster donor to anamorsin being this function dependent on a direct GLRX3-anamorsin interaction, that BOLA2 is only transiently associated with anamorsin and that this transient association is absolutely dependent on the presence of GLRX3 [29].
The fact that both N-terminal domains of GLRX3 and anamorsin are also present in the yeast homologues Grx3/4 and Dre2 supports the model that the latter can perform an analogous function in the yeast system, i.e., they might drive protein–protein recognition between the two proteins in order to specifically deliver the [2Fe-2S] cluster to the redox-active [2Fe-2S] cluster binding site of Dre2 (Figure 5A). A structural comparison of these domains supports this view since the structural elements that have been shown to specifically recognize each other in the human GLRX3-anamorsin interaction [83] are fully conserved in the N-terminal domains of yeast Grx3/4 and Dre2. Indeed, α-helices 2 and 4 of the Trx-like domain of GLRX3 are conserved in the Trx-like domain of Grx4 [88] (PDB ID 2WZ9). Likewise, the fold of both N-terminal domains of Dre2 [18] and anamorsin [19] belongs to the same family of SAM methyl-transferase enzymes. These enzymes usually contain a central β-sheet and six α-helices (αZ and αA-αE), three helices on each side of the β-sheet [89]. The helices C, D and E that are located on the same side of the β-sheet are those involved in protein recognition with the N-terminal domain of GLRX3-Grx3/4 [83] and they are fully conserved in Dre2 [18]. Thus, these structurally common features support a common functional role that these N-terminal domains play in the [2Fe-2S] cluster insertion into both Dre2 and anamorsin.
How the [2Fe-2S] clusters are formed on GLRX3 is a matter of debate in the literature (see reviews [90,91] for details, Figure 5B). Recently, a model on the origin of iron ions used to assemble the [2Fe-2S] cluster on GLRX3 has been proposed. It was argued that the protein Poly(rC)-binding protein 1 (PCBP1), which is a multifunctional protein that coordinates both RNA and iron [92,93], forms an intermediate iron chaperone complex with BOLA2 [94]. Indeed, experimental evidence supports that PCBP1 complexes with BOLA2 via a bridging Fe(II) ligand, and that this complex is then needed for the assembly of the [2Fe-2S] on GLRX3-BOLA2 [95]. However, no spectroscopic evidence showing the formation of the iron chaperone complex, as well as showing its functional role as iron donor towards the apo GLRX3-BOLA2 complex, are still available, thus making this model still largely speculative.
6.3. Cellular Scenario of [2Fe-2S] Cluster Insertion into Anamorsin
In a cellular scenario, two are the possible complexes responsible for the [2Fe-2S] cluster insertion into anamorsin, i.e., [2Fe-2S]2 GLRX32 and GLRX3-[2Fe-2S]2-BOLA22 (Figure 5A). The arising question is whether both are active at the cellular level. In vivo studies in HEK293 cells showed that GLRX3 homodimers were a transient, labile species that did not require Fe/S clusters for formation; furthermore, GLRX3 homodimers with bridging [2Fe-2S] clusters did not accumulate to a detectable degree [29]. Thus, these results suggest that GLRX3 homodimers are not the physiological [2Fe-2S] cluster donor to anamorsin. In agreement with this possible model, in vitro studies showed that GLRX3 homodimers with bound [2Fe-2S] clusters undergo spontaneous rearrangement in the presence of BOLA2 to form GLRX3-[2Fe-2S]2-BOLA22 complexes, suggesting that clusters preferentially form on the heterotrimeric complex [87]. However, it seems a reasonable model where both complexes can be active and thus cellular conditions might select which is the active one. Indeed, both complexes have the N-terminal domain of GLRX3 that is the trigging factor specifically driving [2Fe-2S] cluster transfer from GLRX3 to anamorsin. In addition, the relative cellular levels of GLRX3 and BOLA2, which can be regulated by cellular conditions (i.e., aerobic vs. anaerobic cellular growth, oxidative stress, etc.), might select which of the two complexes mature anamorsin. This model is consistent with the observation that Fe/S protein biogenesis pathways are sensitive toward oxygen and/or oxidative stress for members of both bacterial and eukaryotic Fe/S cluster assembly machineries [96,97,98,99,100]. Consistent with cellular conditions playing a role in anamorsin cluster insertion, bacterial BolA proteins have been observed to be specifically required under aerobic and oxidative stress conditions. For instance, BolA protein is upregulated in E. coli under oxidative stress conditions [101], and in bacterial operons, BolA tends to co-occur not only with a monothiol glutaredoxin, but also with proteins involved in the defense against oxidative stress [102,103]. In eukaryotic genomes, the presence of BOLAs strongly correlates with an aerobic metabolism [104], and some yeast CIA members were found to promote the maturation of cytosolic Fe/S proteins under oxidative stress cellular conditions, these factors being bypassed to some extent in anaerobic conditions [105]. Last but not least, BOLA2 binding to GLRX3 stabilizes [2Fe-2S]2+ clusters against oxidative degradation [86,106], similarly to what was observed in the yeast [2Fe-2S]2+ Fra2-Grx3/4 complexes [50].
An alternative maturation pathway of anamorsin independent of GLRX3 has also been proposed to be operative in human cells. The fact that the iron content found in anamorsin did not completely disappear in cells lacking GLRX3 [29] might suggest that this iron bound to anamorsin could derive from an alternative [2Fe-2S] cluster donor in the cell. In vitro data showing that two human proteins belonging to the NEET protein family, i.e., NAF-1 and mNT, can interact and transfer their [2Fe-2S] clusters to both motifs I and II of anamorsin with second order rate constants similar to those of other known human [2Fe-2S] transfer protein, support that this is the alternative pathway operating in the cell [107]. This model, however, still lacks an in vivo validation. Indeed, a decreased iron content found in anamorsin upon NAF-1 and mNT depletion in GLRX3-deficient cells is still not verified nor are any proteomic data showing an in vivo partnership of these two proteins still available.
7. Conclusions and Perspectives
In this review, we discussed the role that the Fe/S clusters bound to the C-terminal CIAPIN1 domain play in the three functional processes described so far for the CIAPIN1 protein family. We found that the three functions are strictly dependent on a complex formation between a CIAPIN1 protein and a diflavin reductase. We described that this complex is the molecular architecture built to provide electrons to the [2Fe-2S] cluster bound to the CX8CX2CXC motif, which can thus work as an electron reservoir for assembling cytosolic [4Fe-4S] clusters and the diferric-tyrosyl radical cofactor of RNR. The destruction of the Fe/S clusters in the complex by oxidative conditions might also be involved in the regulation of cell death in yeast and potentially also in humans. While in yeast a model of the apoptotic pathway has been proposed, the pathway activating the apoptotic cellular response in humans is still elusive. While the function of the cluster bound to the CX8CX2CXC motif has been defined as a redox center, the functional role of the Fe/S cluster bound to the CX2CX7CX2C motif is still unknown. The fact that the nature of the Fe/S cluster bound to the latter motif varies depending on the CIAPIN1 proteins that have been investigated also adds complexity to defining what the Fe/S cluster bound to the CX2CX7CX2C motif functionally does. Finally, the cellular pathway(s) inserting the two Fe/S clusters into the two Cys-rich motifs is still not defined in yeasts, while in humans a model has been described on the basis of consistent in vivo and in vitro data. Looking ahead, we believe that the investigation of the cellular components responsible for the insertion of the Fe/S clusters into CIAPIN1 proteins from organisms other than humans and yeast would provide significant advances to shed light on the functional role of the two clusters in the cellular processes involving the CIAPIN1 protein family. Studies at the molecular level of these pathways, which are still quite few, will also help to unravel the molecular factors required to assemble the Fe/S clusters in the CIAPIN1 protein family.
Author Contributions
Conceptualization, S.C.-B.; investigation, C.A.; data curation, C.A.; writing—original draft preparation, S.C.-B.; writing—review and editing, S.C.-B. and C.A.; visualization, S.C.-B. and C.A.; supervision, S.C.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: https://pfam.xfam.org/family/ciapin1#tabview=tab3 (accessed on 7 March 2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beinert, H.; Holm, R.H.; Munck, E. Iron-sulfur clusters: Nature’s modular, multipurpose structures. Science 1997, 277, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Beinert, H. Iron-sulfur proteins: Ancient structures, still full of surprises. J. Biol. Inorg. Chem. 2000, 5, 2–15. [Google Scholar] [CrossRef]
- Holm, R.H.; Lo, W. Structural Conversions of Synthetic and Protein-Bound Iron-Sulfur Clusters. Chem. Rev. 2016, 116, 13685–13713. [Google Scholar] [CrossRef] [PubMed]
- Braymer, J.J.; Freibert, S.A.; Rakwalska-Bange, M.; Lill, R. Mechanistic concepts of iron-sulfur protein biogenesis in Biology. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118863. [Google Scholar] [CrossRef] [PubMed]
- Maio, N.; Rouault, T.A. Outlining the Complex Pathway of Mammalian Fe-S Cluster Biogenesis. Trends Biochem. Sci. 2020, 45, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Rouault, T.A. Biogenesis of iron-sulfur clusters in mammalian cells: New insights and relevance to human disease. Dis. Model. Mech. 2012, 5, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Sheftel, A.; Stehling, O.; Lill, R. Iron-sulfur proteins in health and disease. Trends Endocrinol. Metab. 2010, 21, 302–314. [Google Scholar] [CrossRef]
- Fontecave, M.; Py, B.; Ollagnier de Choudens, S.; Barras, F. From Iron and Cysteine to Iron-Sulfur Clusters: The Biogenesis Protein Machineries. EcoSal Plus 2008, 3. [Google Scholar] [CrossRef]
- Baussier, C.; Fakroun, S.; Aubert, C.; Dubrac, S.; Mandin, P.; Py, B.; Barras, F. Making iron-sulfur cluster: Structure, regulation and evolution of the bacterial ISC system. Adv. Microb. Physiol. 2020, 76, 1–39. [Google Scholar]
- Garcia, P.S.; Gribaldo, S.; Py, B.; Barras, F. The SUF system: An ABC ATPase-dependent protein complex with a role in Fe-S cluster biogenesis. Res. Microbiol. 2019, 170, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Outten, F.W. Recent advances in the Suf Fe-S cluster biogenesis pathway: Beyond the Proteobacteria. Biochim. Biophys. Acta 2015, 1853, 1464–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, B.; Gerez, C.; de Choudens, S.O. Assembly of Fe/S proteins in bacterial systems: Biochemistry of the bacterial ISC system. Biochim. Biophys. Acta 2015, 1853, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Netz, D.J.; Mascarenhas, J.; Stehling, O.; Pierik, A.J.; Lill, R. Maturation of cytosolic and nuclear iron-sulfur proteins. Trends Cell Biol. 2014, 24, 303–312. [Google Scholar] [CrossRef]
- Zhang, Y.; Lyver, E.R.; Nakamaru-Ogiso, E.; Yoon, H.; Amutha, B.; Lee, D.W.; Bi, E.; Ohnishi, T.; Daldal, F.; Pain, D.; et al. Dre2, a conserved eukaryotic Fe/S cluster protein, functions in cytosolic Fe/S protein biogenesis. Mol. Cell. Biol. 2008, 28, 5569–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netz, D.J.; Stumpfig, M.; Dore, C.; Muhlenhoff, U.; Pierik, A.J.; Lill, R. Tah18 transfers electrons to Dre2 in cytosolic iron-sulfur protein biogenesis. Nat. Chem. Biol. 2010, 6, 758–765. [Google Scholar] [CrossRef]
- Basu, S.; Netz, D.J.; Haindrich, A.C.; Herlerth, N.; Lagny, T.J.; Pierik, A.J.; Lill, R.; Lukeš, J. Cytosolic iron-sulphur protein assembly is functionally conserved and essential in procyclic and bloodstream Trypanosoma brucei. Mol. Microbiol. 2014, 93, 897–910. [Google Scholar] [CrossRef]
- Shibayama, H.; Takai, E.; Matsumura, I.; Kouno, M.; Morii, E.; Kitamura, Y.; Takeda, J.; Kanakura, Y. Identification of a cytokine-induced antiapoptotic molecule anamorsin essential for definitive hematopoiesis. J. Exp. Med. 2004, 199, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Soler, N.; Craescu, C.T.; Gallay, J.; Frapart, Y.M.; Mansuy, D.; Raynal, B.; Baldacci, G.; Pastore, A.; Huang, M.E.; Vernis, L. A S-adenosylmethionine methyltransferase-like domain within the essential, Fe-S-containing yeast protein Dre2. FEBS J. 2012, 279, 2108–2119. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Boscaro, F.; Chatzi, A.; Mikolajczyk, M.; Tokatlidis, K.; Winkelmann, J. Anamorsin is a 2Fe2S cluster-containing substrate of the Mia40-dependent mitochondrial protein trapping machinery. Chem. Biol. 2011, 18, 794–804. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Bertini, I.; Calderone, V.; Ciofi-Baffoni, S.; Giachetti, A.; Jaiswal, D.; Mikolajczyk, M.; Piccioli, M.; Winkelmann, J. Molecular view of an electron transfer process essential for iron-sulfur protein biogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 7136–7141. [Google Scholar] [CrossRef] [Green Version]
- Torruella, G.; Derelle, R.; Paps, J.; Lang, B.F.; Roger, A.J.; Shalchian-Tabrizi, K.; Ruiz-Trillo, I. Phylogenetic relationships within the Opisthokonta based on phylogenomic analyses of conserved single-copy protein domains. Mol. Biol. Evol. 2012, 29, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Timme, R.E.; Bachvaroff, T.R.; Delwiche, C.F. Broad phylogenomic sampling and the sister lineage of land plants. PLoS ONE 2012, 7, e29696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferecatu, I.; Goncalves, S.; Golinelli-Cohen, M.P.; Clemancey, M.; Martelli, A.; Riquier, S.; Guittet, E.; Latour, J.M.; Puccio, H.; Drapier, J.C.; et al. The diabetes drug target MitoNEET governs a novel trafficking pathway to rebuild an Fe-S cluster into cytosolic aconitase/iron regulatory protein 1. J. Biol. Chem. 2014, 289, 28070–28086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balk, J.; Pierik, A.J.; Netz, D.J.; Muhlenhoff, U.; Lill, R. The hydrogenase-like Nar1p is essential for maturation of cytosolic and nuclear iron-sulphur proteins. EMBO J. 2004, 23, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehling, O.; Jeoung, J.H.; Freibert, S.A.; Paul, V.D.; Bänfer, S.; Niggemeyer, B.; Rösser, R.; Dobbek, H.; Lill, R. Function and crystal structure of the dimeric P-loop ATPase CFD1 coordinating an exposed [4Fe-4S] cluster for transfer to apoproteins. Proc. Natl. Acad. Sci. USA 2018, 115, E9085–E9094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleh, V.; Riemer, J.; Dancis, A.; Herrmann, J.M. Protein oxidation in the intermembrane space of mitochondria is substrate-specific rather than general. Microb. Cell 2014, 1, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Vernis, L.; Facca, C.; Delagoutte, E.; Soler, N.; Chanet, R.; Guiard, B.; Faye, G.; Baldacci, G. A newly identified essential complex, Dre2-Tah18, controls mitochondria integrity and cell death after oxidative stress in yeast. PLoS ONE 2009, 4, e4376. [Google Scholar] [CrossRef] [Green Version]
- Soler, N.; Delagoutte, E.; Miron, S.; Facca, C.; Baille, D.; d’Autreaux, B.; Craescu, G.; Frapart, Y.M.; Mansuy, D.; Baldacci, G.; et al. Interaction between the reductase Tah18 and highly conserved Fe-S containing Dre2 C-terminus is essential for yeast viability. Mol. Microbiol. 2011, 82, 54–67. [Google Scholar] [CrossRef]
- Frey, A.G.; Palenchar, D.J.; Wildemann, J.D.; Philpott, C.C. A Glutaredoxin-BolA Complex Serves as an Iron-Sulfur Cluster Chaperone for the Cytosolic Cluster Assembly Machinery. J. Biol. Chem. 2016, 291, 22344–22356. [Google Scholar] [CrossRef] [Green Version]
- Stubbe, J.; van Der Donk, W.A. Protein Radicals in Enzyme Catalysis. Chem. Rev. 1998, 98, 705–762. [Google Scholar] [CrossRef]
- Nordlund, P.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 2006, 75, 681–706. [Google Scholar] [CrossRef] [PubMed]
- Chabes, A.; Domkin, V.; Larsson, G.; Liu, A.; Graslund, A.; Wijmenga, S.; Thelander, L. Yeast ribonucleotide reductase has a heterodimeric iron-radical-containing subunit. Proc. Natl. Acad. Sci. USA 2000, 97, 2474–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, L.; Wu, X.; An, X.; Stubbe, J.; Huang, M. Investigation of in vivo diferric tyrosyl radical formation in Saccharomyces cerevisiae Rnr2 protein: Requirement of Rnr4 and contribution of Grx3/4 AND Dre2 proteins. J. Biol. Chem. 2011, 286, 41499–41509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, H.; Zhang, C.; An, X.; Liu, L.; Stubbe, J.; Huang, M. Conserved electron donor complex Dre2-Tah18 is required for ribonucleotide reductase metallocofactor assembly and DNA synthesis. Proc. Natl. Acad. Sci. USA 2014, 111, E1695–E1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Stumpfig, M.; Zhang, C.; An, X.; Stubbe, J.; Lill, R.; Huang, M. The diferric-tyrosyl radical cluster of ribonucleotide reductase and cytosolic iron-sulfur clusters have distinct and similar biogenesis requirements. J. Biol. Chem. 2017, 292, 11445–11451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, Y.; Nasuno, R.; Kawahara, N.; Nishimura, A.; Watanabe, D.; Takagi, H. Regulatory mechanism of the flavoprotein Tah18-dependent nitric oxide synthesis and cell death in yeast. Nitric Oxide 2016, 57, 85–91. [Google Scholar] [CrossRef]
- Almeida, B.; Buttner, S.; Ohlmeier, S.; Silva, A.; Mesquita, A.; Sampaio-Marques, B.; Osório, N.S.; Kollau, A.; Mayer, B.; Leão, C.; et al. NO-mediated apoptosis in yeast. J. Cell Sci. 2007, 120, 3279–3288. [Google Scholar] [CrossRef] [Green Version]
- Murataliev, M.B.; Feyereisen, R.; Walker, F.A. Electron transfer by diflavin reductases. Biochim. Biophys. Acta 2004, 1698, 1–26. [Google Scholar] [CrossRef]
- Banci, L.; Ciofi-Baffoni, S.; Mikolajczyk, M.; Winkelmann, J.; Bill, E.; Pandelia, M.E. Human anamorsin binds [2Fe-2S] clusters with unique electronic properties. J. Biol. Inorg. Chem. 2013, 18, 883–893. [Google Scholar] [CrossRef]
- Paul, V.D.; Lill, R. Biogenesis of cytosolic and nuclear iron-sulfur proteins and their role in genome stability. Biochim. Biophys. Acta 2015, 1853, 1528–1539. [Google Scholar] [CrossRef] [Green Version]
- Kunichika, K.; Nakamura, R.; Fujishiro, T.; Takahashi, Y. The Structure of the Dimeric State of IscU Harboring Two Adjacent [2Fe-2S] Clusters Provides Mechanistic Insights into Cluster Conversion to [4Fe-4S]. Biochemistry 2021, 60, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Azam, T.; Przybyla-Toscano, J.; Vignols, F.; Couturier, J.; Rouhier, N.; Johnson, M.K. The Arabidopsis Mitochondrial Glutaredoxin GRXS15 Provides [2Fe-2S] Clusters for ISCA-Mediated [4Fe-4S] Cluster Maturation. Int. J. Mol. Sci. 2020, 21, 9237. [Google Scholar] [CrossRef] [PubMed]
- Weiler, B.D.; Brück, M.C.; Kothe, I.; Bill, E.; Lill, R.; Mühlenhoff, U. Mitochondrial [4Fe-4S] protein assembly involves reductive [2Fe-2S] cluster fusion on ISCA1-ISCA2 by electron flow from ferredoxin FDX2. Proc. Natl. Acad. Sci. USA 2020, 117, 20555–20565. [Google Scholar] [CrossRef] [PubMed]
- Chandramouli, K.; Unciuleac, M.C.; Naik, S.; Dean, D.R.; Huynh, B.H.; Johnson, M.K. Formation and properties of [4Fe-4S] clusters on the IscU scaffold protein. Biochemistry 2007, 46, 6804–6811. [Google Scholar] [CrossRef]
- Brancaccio, D.; Gallo, A.; Mikolajczyk, M.; Zovo, K.; Palumaa, P.; Novellino, E.; Piccioli, M.; Ciofi-Baffoni, S.; Banci, L. Formation of [4Fe-4S] clusters in the mitochondrial iron-sulfur cluster assembly machinery. J. Am. Chem. Soc. 2014, 136, 16240–16250. [Google Scholar] [CrossRef]
- Brancaccio, D.; Gallo, A.; Piccioli, M.; Novellino, E.; Ciofi-Baffoni, S.; Banci, L. [4Fe-4S] Cluster Assembly in Mitochondria and Its Impairment by Copper. J. Am. Chem. Soc. 2017, 139, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Laursen, T.; Jensen, K.; Moller, B.L. Conformational changes of the NADPH-dependent cytochrome P450 reductase in the course of electron transfer to cytochromes P450. Biochim. Biophys. Acta 2011, 1814, 132–138. [Google Scholar] [CrossRef]
- Ellis, J.; Gutierrez, A.; Barsukov, I.L.; Huang, W.C.; Grossmann, J.G.; Roberts, G.C. Domain motion in cytochrome P450 reductase: Conformational equilibria revealed by NMR and small-angle x-ray scattering. J. Biol. Chem. 2009, 284, 36628–36637. [Google Scholar] [CrossRef] [Green Version]
- Tarassov, K.; Messier, V.; Landry, C.R.; Radinovic, S.; Serna Molina, M.M.; Shames, I.; Malitskaya, Y.; Vogel, J.; Bussey, H.; Michnick, S.W. An in vivo map of the yeast protein interactome. Science 2008, 320, 1465–1470. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mapolelo, D.T.; Dingra, N.N.; Naik, S.G.; Lees, N.S.; Hoffman, B.M.; Riggs-Gelasco, P.J.; Huynh, B.H.; Johnson, M.K.; Outten, C.E. The yeast iron regulatory proteins Grx3/4 and Fra2 form heterodimeric complexes containing a [2Fe-2S] cluster with cysteinyl and histidyl ligation. Biochemistry 2009, 48, 9569–9581. [Google Scholar] [CrossRef] [Green Version]
- Muhlenhoff, U.; Molik, S.; Godoy, J.R.; Uzarska, M.A.; Richter, N.; Seubert, A.; Zhang, Y.; Stubbe, J.; Pierrel, F.; Herrero, E.; et al. Cytosolic monothiol glutaredoxins function in intracellular iron sensing and trafficking via their bound iron-sulfur cluster. Cell Metab. 2010, 12, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Camponeschi, F.; Ciofi-Baffoni, S.; Banci, L. Anamorsin/Ndor1 Complex Reduces [2Fe-2S]-MitoNEET via a Transient Protein-Protein Interaction. J. Am. Chem. Soc. 2017, 139, 9479–9482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamir, S.; Paddock, M.L.; Darash-Yahana-Baram, M.; Holt, S.H.; Sohn, Y.S.; Agranat, L.; Michaeli, D.; Stofleth, J.T.; Lipper, C.H.; Morcos, F.; et al. Structure-function analysis of NEET proteins uncovers their role as key regulators of iron and ROS homeostasis in health and disease. Biochim. Biophys. Acta 2015, 1853, 1294–1315. [Google Scholar] [CrossRef] [Green Version]
- Karmi, O.; Marjault, H.B.; Pesce, L.; Carloni, P.; Onuchic, J.N.; Jennings, P.A.; Mittler, R.; Nechushtai, R. The unique fold and lability of the [2Fe-2S] clusters of NEET proteins mediate their key functions in health and disease. J. Biol. Inorg. Chem. 2018, 23, 599–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, D.W.; Elliott, S.J. Conserved hydrogen bonding networks of MitoNEET tune Fe-S cluster binding and structural stability. Biochemistry 2013, 52, 4687–4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuris, J.A.; Harir, Y.; Conlan, A.R.; Shvartsman, M.; Michaeli, D.; Tamir, S.; Paddock, M.L.; Onuchic, J.N.; Mittler, R.; Cabantchik, Z.I.; et al. Facile transfer of [2Fe-2S] clusters from the diabetes drug target mitoNEET to an apo-acceptor protein. Proc. Natl. Acad. Sci. USA 2011, 108, 13047–13052. [Google Scholar] [CrossRef] [Green Version]
- Golinelli-Cohen, M.P.; Lescop, E.; Mons, C.; Goncalves, S.; Clemancey, M.; Santolini, J.; Guittet, E.; Blondin, G.; Latour, J.M.; Bouton, C. Redox Control of the Human Iron-Sulfur Repair Protein MitoNEET Activity via Its Iron-Sulfur Cluster. J. Biol. Chem. 2016, 291, 7583–7593. [Google Scholar] [CrossRef] [Green Version]
- Landry, A.P.; Ding, H. Redox control of human mitochondrial outer membrane protein MitoNEET [2Fe-2S] clusters by biological thiols and hydrogen peroxide. J. Biol. Chem. 2014, 289, 4307–4315. [Google Scholar] [CrossRef] [Green Version]
- Bak, D.W.; Zuris, J.A.; Paddock, M.L.; Jennings, P.A.; Elliott, S.J. Redox characterization of the FeS protein MitoNEET and impact of thiazolidinedione drug binding. Biochemistry 2009, 48, 10193–10195. [Google Scholar] [CrossRef] [Green Version]
- Conlan, A.R.; Axelrod, H.L.; Cohen, A.E.; Abresch, E.C.; Zuris, J.; Yee, D.; Nechushtai, R.; Jennings, P.A.; Paddock, M.L. Crystal structure of Miner1: The redox-active 2Fe-2S protein causative in Wolfram Syndrome 2. J. Mol. Biol. 2009, 392, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Stehling, O.; Mascarenhas, J.; Vashisht, A.A.; Sheftel, A.D.; Niggemeyer, B.; Rosser, R.; Pierik, A.J.; Wohlschlegel, J.A.; Lill, R. Human CIA2A-FAM96A and CIA2B-FAM96B integrate iron homeostasis and maturation of different subsets of cytosolic-nuclear iron-sulfur proteins. Cell Metab. 2013, 18, 187–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, G.; Liu, D.; Pan, F.; Zhao, J.; Li, T.; Ma, Y.; Shen, B.; Lyu, J. His-87 ligand in mitoNEET is crucial for the transfer of iron sulfur clusters from mitochondria to cytosolic aconitase. Biochem. Biophy.s Res. Commun. 2016, 470, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Mons, C.; Ferecatu, I.; Riquier, S.; Lescop, E.; Bouton, C.; Golinelli-Cohen, M.P. Combined Biochemical, Biophysical, and Cellular Methods to Study Fe-S Cluster Transfer and Cytosolic Aconitase Repair by MitoNEET. Methods Enzym. 2017, 595, 83–106. [Google Scholar]
- Kühn, L.C. Iron regulatory proteins and their role in controlling iron metabolism. Metallomics 2015, 7, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, C.; Dancis, A.; Nakamaru-Ogiso, E. EPR studies of wild type and mutant Dre2 identify essential [2Fe--2S] and [4Fe--4S] clusters and their cysteine ligands. J. Biochem. 2017, 161, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netz, D.J.; Genau, H.M.; Weiler, B.D.; Bill, E.; Pierik, A.J.; Lill, R. The conserved protein Dre2 uses essential [2Fe-2S] and [4Fe-4S] clusters for its function in cytosolic iron-sulfur protein assembly. Biochem. J. 2016, 473, 2073–2085. [Google Scholar] [CrossRef]
- Bernard, D.G.; Netz, D.J.; Lagny, T.J.; Pierik, A.J.; Balk, J. Requirements of the cytosolic iron-sulfur cluster assembly pathway in Arabidopsis. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matteucci, S.; Camponeschi, F.; Clémancey, M.; Ciofi-Baffoni, S.; Blondin, G.; Banci, L. In Cellulo Mössbauer and EPR Studies Bring New Evidence to the Long-Standing Debate on Iron-Sulfur Cluster Binding in Human Anamorsin. Angew. Chem. Int. Ed. 2021, 60, 14841–14845. [Google Scholar] [CrossRef] [PubMed]
- Hagen, W.R. EPR spectroscopy of complex biological iron-sulfur systems. J. Biol. Inorg. Chem. 2018, 23, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondin, G.; Girerd, J.-J. Interplay of electron exchange and electron transfer in metal polynuclear complexes in proteins or chemical models. Chem. Rev. 1990, 90, 1359–1376. [Google Scholar] [CrossRef]
- Iwema, T.; Picciocchi, A.; Traore, D.A.; Ferrer, J.L.; Chauvat, F.; Jacquamet, L. Structural basis for delivery of the intact [Fe2S2] cluster by monothiol glutaredoxin. Biochemistry 2009, 48, 6041–6043. [Google Scholar] [CrossRef] [PubMed]
- Picciocchi, A.; Saguez, C.; Boussac, A.; Cassier-Chauvat, C.; Chauvat, F. CGFS-type monothiol glutaredoxins from the cyanobacterium Synechocystis PCC6803 and other evolutionary distant model organisms possess a glutathione-ligated [2Fe-2S] cluster. Biochemistry 2007, 46, 15018–15026. [Google Scholar] [CrossRef] [PubMed]
- Kumanovics, A.; Chen, O.S.; Li, L.; Bagley, D.; Adkins, E.M.; Lin, H.; Dingra, N.N.; Outten, C.E.; Keller, G.; Winge, D.; et al. Identification of FRA1 and FRA2 as genes involved in regulating the yeast iron regulon in response to decreased mitochondrial iron-sulfur cluster synthesis. J. Biol. Chem. 2008, 283, 10276–10286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talib, E.A.; Outten, C.E. Iron-sulfur cluster biogenesis, trafficking, and signaling: Roles for CGFS glutaredoxins and BolA proteins. Biochim. Biophys. Acta. Mol. Cell Res. 2020, 1868, 118847. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Outten, C.E. Monothiol CGFS glutaredoxins and BolA-like proteins: [2Fe-2S] binding partners in iron homeostasis. Biochemistry 2012, 51, 4377–4389. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mapolelo, D.T.; Dingra, N.N.; Keller, G.; Riggs-Gelasco, P.J.; Winge, D.R.; Johnson, M.K.; Outten, C.E. Histidine 103 in Fra2 is an iron-sulfur cluster ligand in the [2Fe-2S] Fra2-Grx3 complex and is required for in vivo iron signaling in yeast. J. Biol. Chem. 2011, 286, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.B.; Tang, Y.; Zhang, J.; Dai, Y.N.; Abdalla, M.; Chen, Y.; Zhou, C.Z. Structural and Biochemical Insights into the Multiple Functions of Yeast Grx3. J. Mol. Biol. 2018, 430, 1235–1248. [Google Scholar] [CrossRef]
- Pandey, A.K.; Pain, J.; Dancis, A.; Pain, D. Mitochondria export iron-sulfur and sulfur intermediates to the cytoplasm for iron-sulfur cluster assembly and tRNA thiolation in yeast. J. Biol. Chem. 2019, 294, 9489–9502. [Google Scholar] [CrossRef]
- Freibert, S.A.; Boniecki, M.T.; Stümpfig, C.; Schulz, V.; Krapoth, N.; Winge, D.R.; Mühlenhoff, U.; Stehling, O.; Cygler, M.; Lill, R. N-terminal tyrosine of ISCU2 triggers [2Fe-2S] cluster synthesis by ISCU2 dimerization. Nat. Commun. 2021, 12, 6902. [Google Scholar] [CrossRef]
- Poor, C.B.; Wegner, S.V.; Li, H.; Dlouhy, A.C.; Schuermann, J.P.; Sanishvili, R.; Hinshaw, J.R.; Riggs-Gelasco, P.J.; Outten, C.E.; He, C. Molecular mechanism and structure of the Saccharomyces cerevisiae iron regulator Aft2. Proc. Natl. Acad. Sci. USA 2014, 111, 4043–4048. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Shibayama, H.; Tanaka, H.; Tanimura, A.; Matsumura, I.; Kanakura, Y. PICOT is a molecule which binds to anamorsin. Biochem. Biophys. Res. Commun. 2011, 408, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Haunhorst, P.; Berndt, C.; Eitner, S.; Godoy, J.R.; Lillig, C.H. Characterization of the human monothiol glutaredoxin 3 (PICOT) as iron-sulfur protein. Biochem. Biophys. Res. Commun. 2010, 394, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Ciofi-Baffoni, S.; Gajda, K.; Muzzioli, R.; Peruzzini, R.; Winkelmann, J. N-terminal domains mediate [2Fe-2S] cluster transfer from glutaredoxin-3 to anamorsin. Nat. Chem. Biol. 2015, 11, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Haunhorst, P.; Hanschmann, E.M.; Brautigam, L.; Stehling, O.; Hoffmann, B.; Muhlenhoff, U.; Lill, R.; Berndt, C.; Lillig, C.H. Crucial function of vertebrate glutaredoxin 3 (PICOT) in iron homeostasis and hemoglobin maturation. Mol. Biol. Cell 2013, 24, 1895–1903. [Google Scholar] [CrossRef]
- Luck, K.; Kim, D.K.; Lambourne, L.; Spirohn, K.; Begg, B.E.; Bian, W.; Brignall, R.; Cafarelli, T.; Campos-Laborie, F.J.; Charloteaux, B.; et al. A reference map of the human binary protein interactome. Nature 2020, 580, 402–408. [Google Scholar] [CrossRef]
- Li, H.; Mapolelo, D.T.; Randeniya, S.; Johnson, M.K.; Outten, C.E. Human glutaredoxin 3 forms [2Fe-2S]-bridged complexes with human BolA2. Biochemistry 2012, 51, 1687–1696. [Google Scholar] [CrossRef]
- Banci, L.; Camponeschi, F.; Ciofi-Baffoni, S.; Muzzioli, R. Elucidating the molecular function of human BOLA2 in GRX3-Dependent anamorsin maturation pathway. J. Am. Chem. Soc. 2015, 137, 16133–16134. [Google Scholar] [CrossRef]
- Gibson, L.M.; Dingra, N.N.; Outten, C.E.; Lebioda, L. Structure of the thioredoxin-like domain of yeast glutaredoxin 3. Acta Crystallogr. Sect. D Biol. Crystallogr. 2008, 64, 927–932. [Google Scholar] [CrossRef]
- Martin, J.L.; McMillan, F.M. SAM (dependent) I AM: The S-adenosylmethionine-dependent methyltransferase fold. Curr. Opin. Struct. Biol. 2002, 12, 783–793. [Google Scholar] [CrossRef]
- Philpott, C.C.; Ryu, M.S.; Frey, A.; Patel, S. Cytosolic iron chaperones: Proteins delivering iron cofactors in the cytosol of mammalian cells. J. Biol. Chem. 2017, 292, 12764–12771. [Google Scholar] [CrossRef] [Green Version]
- Maio, N.; Rouault, T.A. Mammalian iron sulfur cluster biogenesis: From assembly to delivery to recipient proteins with a focus on novel targets of the chaperone and co-chaperone proteins. IUBMB Life 2022. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Bencze, K.Z.; Stemmler, T.L.; Philpott, C.C. A cytosolic iron chaperone that delivers iron to ferritin. Science 2008, 320, 1207–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makeyev, A.V.; Liebhaber, S.A. The poly(C)-binding proteins: A multiplicity of functions and a search for mechanisms. Rna 2002, 8, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.J.; Frey, A.G.; Palenchar, D.J.; Achar, S.; Bullough, K.Z.; Vashisht, A.; Wohlschlegel, J.A.; Philpott, C.C. A PCBP1-BolA2 chaperone complex delivers iron for cytosolic [2Fe-2S] cluster assembly. Nat. Chem. Biol. 2019, 15, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.J.; Protchenko, O.; Shakoury-Elizeh, M.; Baratz, E.; Jadhav, S.; Philpott, C.C. The iron chaperone and nucleic acid-binding activities of poly(rC)-binding protein 1 are separable and independently essential. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Py, B.; Gerez, C.; Angelini, S.; Planel, R.; Vinella, D.; Loiseau, L.; Talla, E.; Brochier-Armanet, C.; Garcia, S.R.; Latour, J.M.; et al. Molecular organization, biochemical function, cellular role and evolution of NfuA, an atypical Fe-S carrier. Mol. Microbiol. 2012, 86, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.C.; Unciuleac, M.C.; Dean, D.R. Controlled expression and functional analysis of iron-sulfur cluster biosynthetic components within Azotobacter vinelandii. J. Bacteriol. 2006, 188, 7551–7561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, G.; Lu, J.; Bitoun, J.P.; Huang, H.; Ding, H. IscA/SufA paralogues are required for the [4Fe-4S] cluster assembly in enzymes of multiple physiological pathways in Escherichia coli under aerobic growth conditions. Biochem. J. 2009, 420, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, S.; Gerez, C.; Ollagnier-de, C.S.; Sanakis, Y.; Fontecave, M.; Barras, F.; Py, B. NfuA, a new factor required for maturing Fe/S proteins in Escherichia coli under oxidative stress and iron starvation conditions. J. Biol. Chem. 2008, 283, 14084–14091. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, S.; Naik, S.G.; O’Carroll, I.P.; Huynh, B.H.; Dean, D.R.; Johnson, M.K.; Dos Santos, P.C. A proposed role for the Azotobacter vinelandii NfuA protein as an intermediate iron-sulfur cluster carrier. J. Biol. Chem. 2008, 283, 14092–14099. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Freire, P.; Vicente, M.; Arraiano, C.M. The stationary-phase morphogene bolA from Escherichia coli is induced by stress during early stages of growth. Mol. Microbiol. 1999, 32, 789–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynen, M.A.; Spronk, C.A.; Gabaldón, T.; Snel, B. Combining data from genomes, Y2H and 3D structure indicates that BolA is a reductase interacting with a glutaredoxin. FEBS Lett. 2005, 579, 591–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreini, C.; Rosato, A.; Banci, L. The relationship between environmental dioxygen and iron-sulfur proteins explored at the genome level. PLoS ONE 2017, 12, e0171279. [Google Scholar] [CrossRef] [PubMed]
- Willems, P.; Wanschers, B.F.; Esseling, J.; Szklarczyk, R.; Kudla, U.; Duarte, I.; Forkink, M.; Nooteboom, M.; Swarts, H.; Gloerich, J.; et al. BOLA1 is an aerobic protein that prevents mitochondrial morphology changes induced by glutathione depletion. Antioxid. Redox. Signal. 2013, 18, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.D.; Muhlenhoff, U.; Stumpfig, M.; Seebacher, J.; Kugler, K.G.; Renicke, C.; Taxis, C.; Gavin, A.C.; Pierik, A.J.; Lill, R. The deca-GX3 proteins Yae1-Lto1 function as adaptors recruiting the ABC protein Rli1 for iron-sulfur cluster insertion. Elife 2015, 4, e08231. [Google Scholar] [CrossRef]
- Nuttle, X.; Giannuzzi, G.; Duyzend, M.H.; Schraiber, J.G.; Narvaiza, I.; Camponeschi, F.; Ciofi-Baffoni, S.; Sudmant, P.H.; Penn, O.; Chiatante, G.; et al. Emergence of a Homo sapiens-specific gene family and the evolution of autism risk at chromosome 16p11.2. Nature 2016, 536, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Lipper, C.H.; Paddock, M.L.; Onuchic, J.N.; Mittler, R.; Nechushtai, R.; Jennings, P.A. Cancer-Related NEET Proteins Transfer 2Fe-2S Clusters to Anamorsin, a Protein Required for Cytosolic Iron-Sulfur Cluster Biogenesis. PLoS ONE 2015, 10, e0139699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic structures of rhomboid [2Fe-2S] and cubane [4Fe-4S] clusters.

Figure 2.
The three cellular processes that involve CIAPIN1 proteins (Dre2 and anamorsin) once complexed with the NADPH-dependent diflavin oxidoreductase 1 (Tah18 and Ndor1). Red sphere is Fe(III) ion, green sphere is Fe(II) ion, yellow sphere is sulfide ion and red/yellow sphere is Fe/S cluster.
Figure 2.
The three cellular processes that involve CIAPIN1 proteins (Dre2 and anamorsin) once complexed with the NADPH-dependent diflavin oxidoreductase 1 (Tah18 and Ndor1). Red sphere is Fe(III) ion, green sphere is Fe(II) ion, yellow sphere is sulfide ion and red/yellow sphere is Fe/S cluster.

Figure 3.
The electron transfer pathway responsible for the formation/reduction of Fe/S clusters and for the formation of the diferric-tyrosyl radical. First, the reduction of the iron-sulfur cluster bound to the CX8CX2CXC motif of the CIAPIN1 domain occurs following a multistep pathway. Then, this reduced Fe/S cluster transfers one electron at a time to reductively couple two [2Fe-2S]2+ clusters into a [4Fe-4S]2+ cluster, to drive the formation of the diferric-tyrosyl radical cofactor in yeast ribonucleotide reductase and to reduce [2Fe-2S]2+ clusters of human mitoNEET. Red sphere is Fe(III) ion, green sphere is Fe(II) ion, yellow sphere is sulfide ion and red/yellow sphere is Fe/S cluster.
Figure 3.
The electron transfer pathway responsible for the formation/reduction of Fe/S clusters and for the formation of the diferric-tyrosyl radical. First, the reduction of the iron-sulfur cluster bound to the CX8CX2CXC motif of the CIAPIN1 domain occurs following a multistep pathway. Then, this reduced Fe/S cluster transfers one electron at a time to reductively couple two [2Fe-2S]2+ clusters into a [4Fe-4S]2+ cluster, to drive the formation of the diferric-tyrosyl radical cofactor in yeast ribonucleotide reductase and to reduce [2Fe-2S]2+ clusters of human mitoNEET. Red sphere is Fe(III) ion, green sphere is Fe(II) ion, yellow sphere is sulfide ion and red/yellow sphere is Fe/S cluster.

Figure 4.
Fe/S cluster binding nature and coordination at the two highly conserved cysteine-rich motifs of Dre2 and anamorsin (CX8CX2CXC, motif I, and CX2CX7CX2C, motif II, where X indicates any amino acid) obtained by EPR and Mössbauer spectroscopies. Red/green sphere is Fe(III) ion (for Mössbauer spectra) or Fe(II) ion (for EPR spectra), green sphere is Fe(II) ion, red sphere is Fe(III) ion and yellow sphere is sulfide ion.
Figure 4.
Fe/S cluster binding nature and coordination at the two highly conserved cysteine-rich motifs of Dre2 and anamorsin (CX8CX2CXC, motif I, and CX2CX7CX2C, motif II, where X indicates any amino acid) obtained by EPR and Mössbauer spectroscopies. Red/green sphere is Fe(III) ion (for Mössbauer spectra) or Fe(II) ion (for EPR spectra), green sphere is Fe(II) ion, red sphere is Fe(III) ion and yellow sphere is sulfide ion.

Figure 5.
Model of Fe/S cluster insertion into Dre2 (A) and anamorsin (B). Red sphere is Fe(III) ion, green sphere is Fe(II) ion and yellow sphere is sulfide ion.
Figure 5.
Model of Fe/S cluster insertion into Dre2 (A) and anamorsin (B). Red sphere is Fe(III) ion, green sphere is Fe(II) ion and yellow sphere is sulfide ion.


{kind=link}
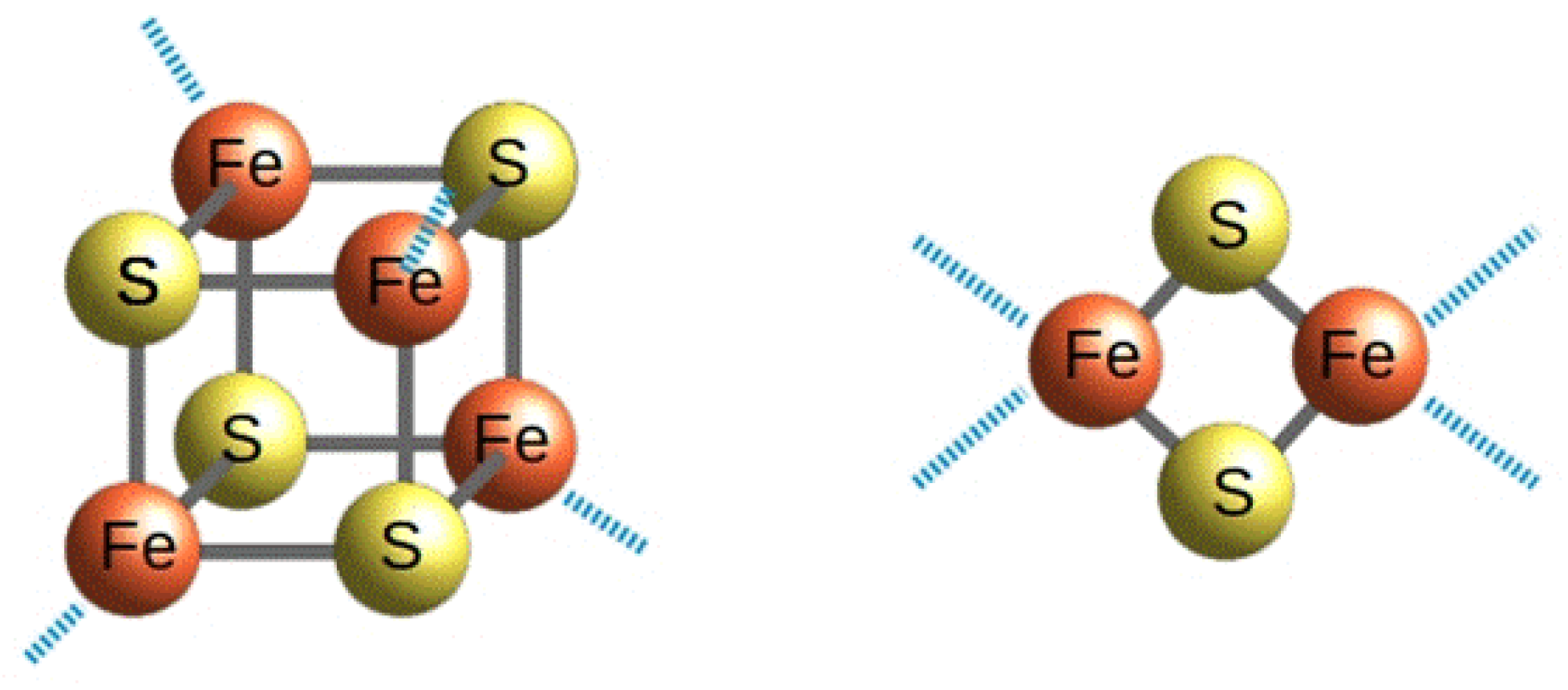{kind=link}
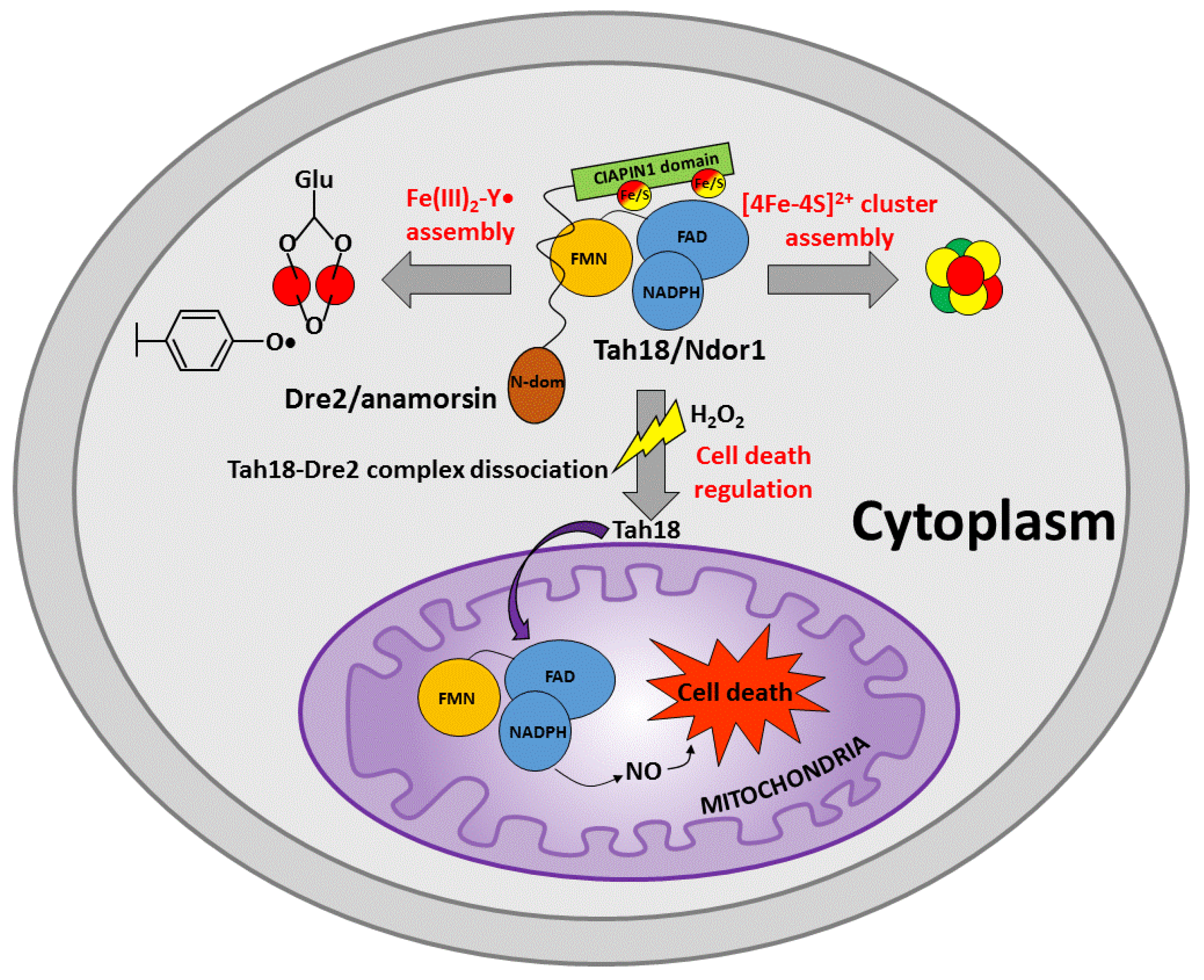{kind=link}
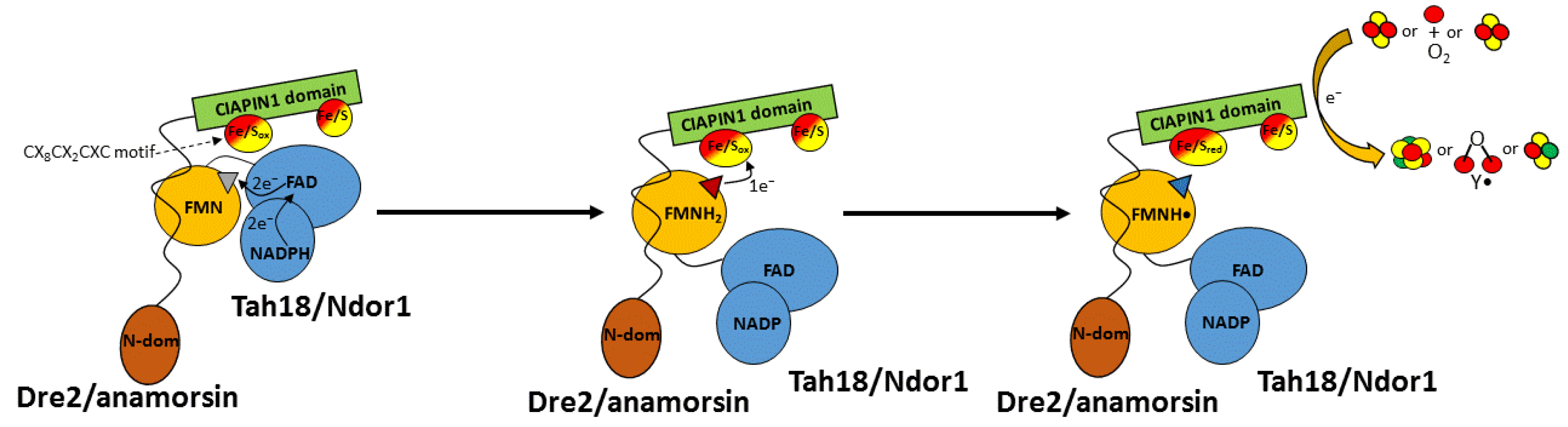{kind=link}
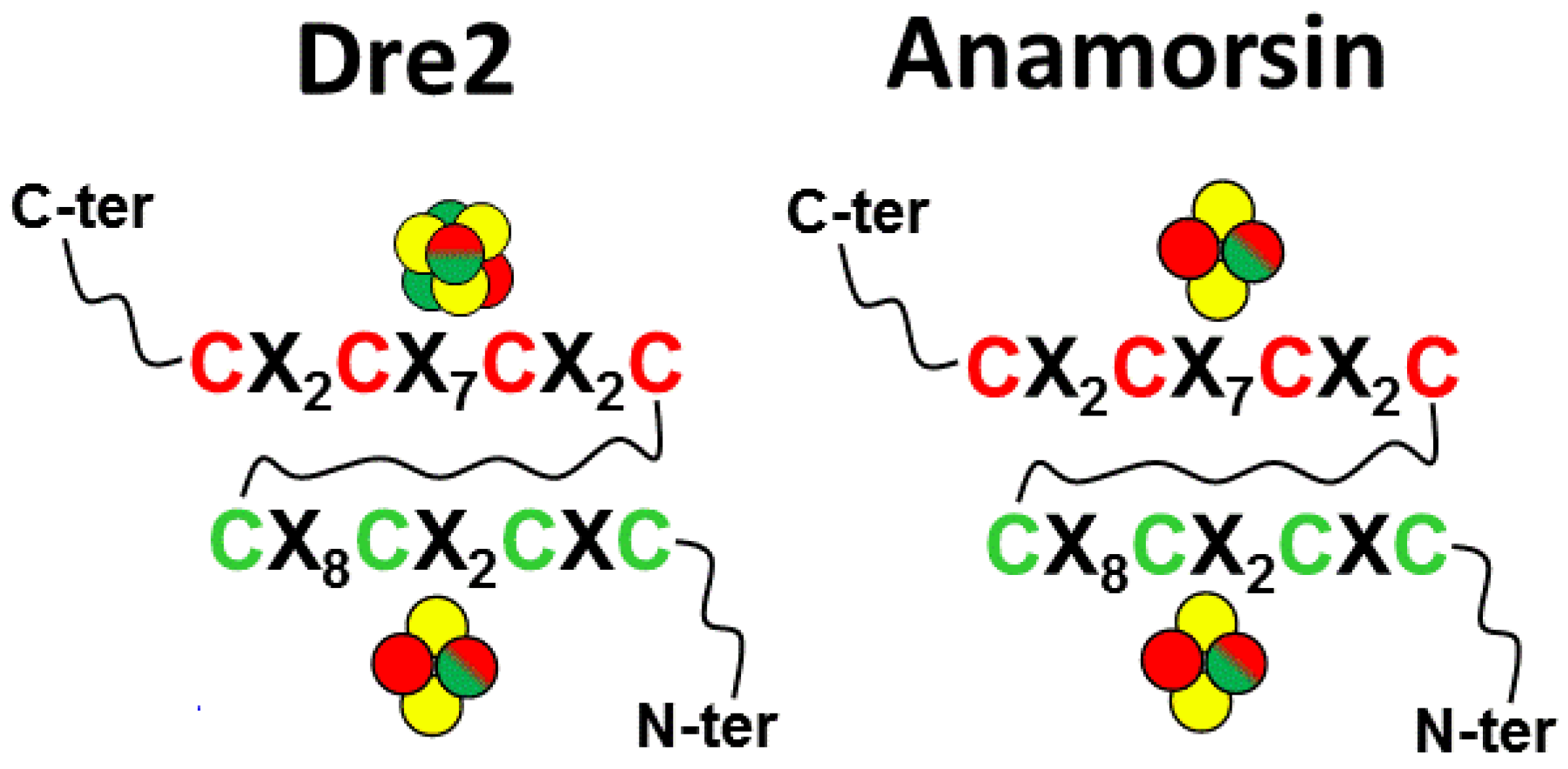{kind=link}
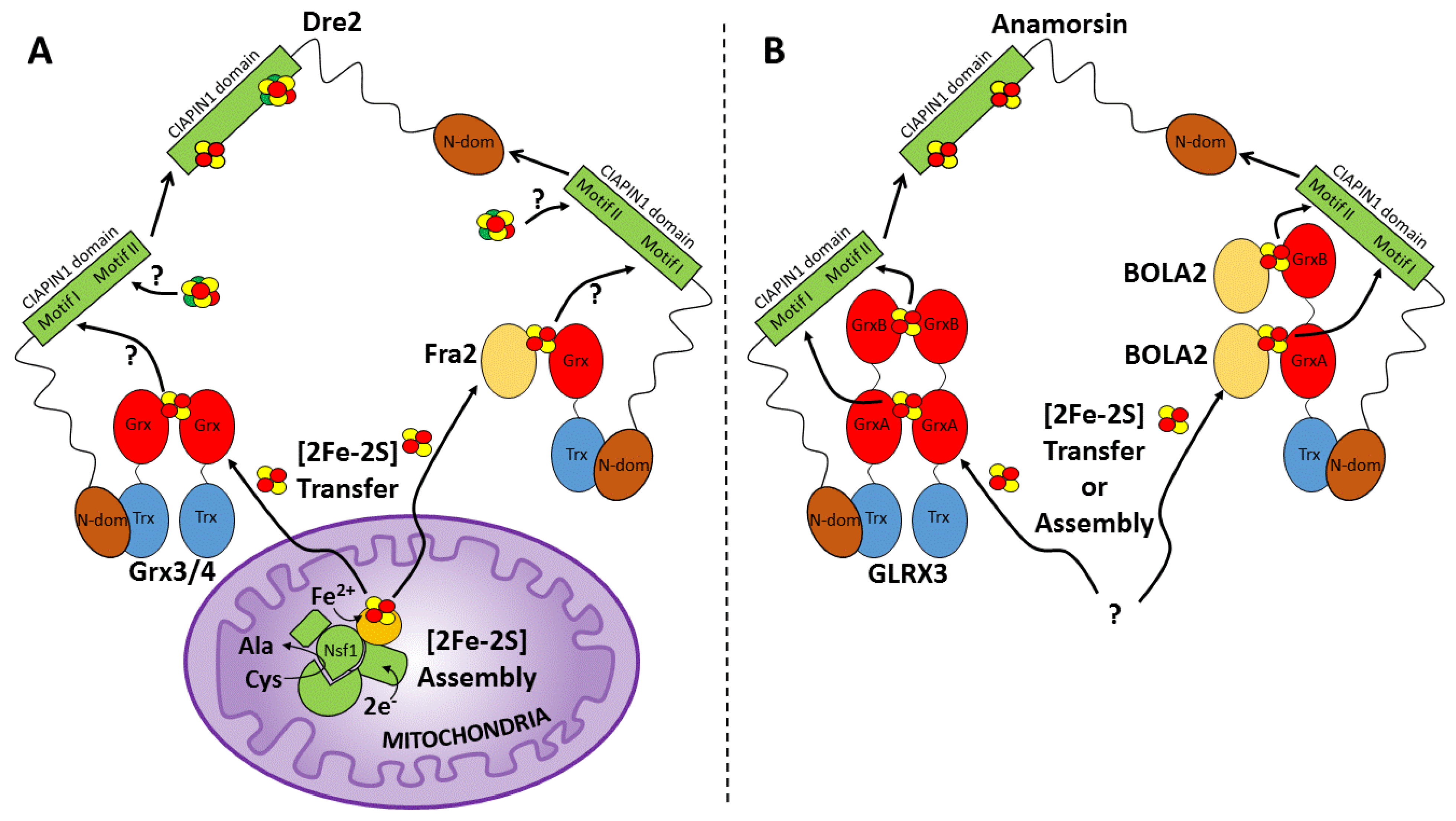{kind=link}
Table 1.
Distribution of CIAPIN1 proteins per clade and domain architectures.
| Clade | # Organisms per Clade | # Organisms with One CIAPIN1 Protein | # Organisms with More Than One CIAPIN1 Protein | Domain Architecture |
|---|---|---|---|---|
| Amorphea | 1214 | 1080 | 134 | Ciapin1_only + Methyltransf11_Ciapin1 + Dre2_Ciapin1 |
| Cryptista | 1 | 0 | 1 | Ciapin1_only |
| Discoba | 18 | 17 | 1 | Ciapin1_only * |
| Haptista | 1 | 1 | 0 | Ciapin1_only |
| TSar | 91 | 82 | 9 | Ciapin1_only |
| Archaeplastida | 185 | 83 | 102 | Ciapin1_only + Methyltransf11_Ciapin1 |
* The A0A6A5BSL2 protein from Naegleria fowleri is an exception because it has a Methyltransf11_Ciapin1 architecture.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ciofi-Baffoni, S.; Andreini, C. The Intriguing Role of Iron-Sulfur Clusters in the CIAPIN1 Protein Family. Inorganics 2022, 10, 52. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics10040052
AMA Style
Ciofi-Baffoni S, Andreini C. The Intriguing Role of Iron-Sulfur Clusters in the CIAPIN1 Protein Family. Inorganics. 2022; 10(4):52. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics10040052
Chicago/Turabian StyleCiofi-Baffoni, Simone, and Claudia Andreini. 2022. "The Intriguing Role of Iron-Sulfur Clusters in the CIAPIN1 Protein Family" Inorganics 10, no. 4: 52. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics10040052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.