1. Introduction
Mosquito-borne diseases continue to impact the health of populations of many of the world’s tropical and subtropical regions. In the absence of approved vaccines, therapeutics, or prophylaxis, mosquito control remains the primary tool for combating many of these diseases, including dengue, chikungunya, and Zika. These viruses are often transmitted by
Aedes aegypti, and the establishment of
A. aegypti in an area increases the risk of autochthonous disease transmission [
1].
As
A. aegypti continues to spread, so does the risk of vector-borne disease transmission. In North America,
A. aegypti primarily occurs in the southern United States, from Arizona eastward and below 33° N latitudes [
2]. In 2017, the continental U.S. reported 5102 symptomatic Zika virus disease cases. Of those, 224 are attributed to local mosquito-borne transmission. The U.S. territories reported an additional 36,079 cases, including 35,937 attributed to local mosquito-borne transmission [
3]. As
A. aegypti spreads, the need for continued exploration of potential control strategies grows.
Aedes aegypti often oviposit in artificial water containers associated with human activity—for example, flower pots, buckets, gutters, etc. Finding and treating these containers, often referred to as “cryptic breeding sites,” can present a challenge [
4,
5,
6,
7,
8]. Successfully reducing adult
A. aegypti populations, however, requires adequate coverage of cryptic breeding sites [
9,
10,
11,
12,
13]. Auto-dissemination strategies target cryptic breeding sites by relying on mosquitoes themselves to deliver lethal doses of insecticide [
14,
15,
16,
17]. Auto-dissemination methods often employ insect growth regulators such as methoprene or pyriproxyfen (PPF), an active ingredient that is neither lethal nor repellant to adult mosquitoes [
16]. Instead, methoprene or PPF acts as a potent inhibitor of immature mosquito development. PPF is an ideal candidate for autodissemination because it remains lethal at concentrations (LC50) of >0.012 parts per billion [
15]. Approximately 1/1000 th the dry weight of a mosquito adult would adequately treat a 200 mL breeding site [
18]. Residual activity of PPF can persist for up to four months [
14]. Moreover,
Aedes mosquitoes have demonstrated no resistance to PPF to date [
19]. PPF, a juvenile hormone analogue, is not harmful to vertebrates. The WHO has, in fact, defined safe levels for direct treatment of drinking water [
20].
The present study evaluates a technique called Auto-Dissemination Augmented by Males (ADAM). The ADAM approach involves the release of laboratory-reared males, which have been dusted with PPF into an infested area. A prior field study using ADAM has demonstrated
Aedes albopictus males to be effective in transferring lethal PPF concentrations both to breeding sites and to conspecific female mosquitoes [
17]. Using ADAM, the present study similarly examines the effectiveness of
A. aegypti males at disseminating PPF to larval breeding sites. Specifically, hypotheses include that an appropriate dosage of PPF applied to adult
A. aegypti males would not result in high levels of acute mortality, male quality would not be impacted from shipment of PPF treated males from a remote production facility to field release locations, that released PPF-dusted males would deliver PPF into introduced containers, and that the adult
A. aegypti population would be significantly reduced following the release of PPF-dusted males, relative to the
A. aegypti population at a similar, untreated area.
This study included both caged- and open-release trials. In cage studies, the effect of PPF dusting on A. aegypti male survivorship and the ability of PPF dusted A. aegypti males to deliver lethal doses to artificial oviposition containers were assessed. In the open-release trials, PPF-treated males were introduced into A. aegypti infested sites in Clovis, CA; Key Largo, FL; and Los Angeles, CA. These open releases utilized male A. aegypti, which were produced and PPF-dusted at a rearing facility in Kentucky and then shipped to release locations. Measurements assessed shipment survivorship as well as A. aegypti male’s ability to successfully transfer PPF to artificial oviposition sites. The results demonstrate that PPF-dusted adult A. aegypti males (1) can be shipped cross-country with little effect on survivorship and (2) can deliver lethal doses of PPF to artificial oviposition sites in a field setting.
2. Materials and Methods
Aedes aegypti mosquitoes used in caged experiments, bioassays, and field releases in Los Angeles and Florida were the WACO strain [
21].
Aedes aegypti mosquitoes used for Clovis shipments and release were from a colony derived from Clovis, California [
22]. Larvae were fed with a 60 g/L liver powder (ICN Biomedicals, Irvine, CA, USA) slurry. Adults were held in 24.5 cm × 24.5 cm × 24.5 cm BugDorm-4S2222 insect rearing cages (MegaView Science Co., Taichung, Taiwan) and provided a constant supply of 10% sucrose. Females were provided with bovine blood for egg production. All experiments used adult young male mosquitoes that were less than 2 d post emergence at the time of release. Prior to the experiments described here, multiple dose-response treatments were performed to determine a dusting rate that did not appear to affect adult
A. aegypti male survival [
17]. For all experiments, the PPF treatment consisted of a 30%:70% mixture of Esteem 35WP (Valent Biosciences, Libertyville, IL, USA) and DayGlo fluorescent powder (DayGlo Color Corp., Cleveland, OH, USA), respectively. Mosquitoes were dusted using a hand bellow powder duster (Harris Manufacturing Co. LLC, Cartersville, NC, USA) in cardboard mailing tubes (63.5 mm diameter 20.3 cm long), capped on both ends with No-See-Um netting (Equinox, Williamsport, PA, USA). Maps of field sites were generated using OpenStreetMap
® (
https://www.openstreetmap.org).
2.1. Laboratory Survivorship and Mesocosm Cage Studies
The study first examined for effects of PPF treatment on adult survivorship. Males were treated with PPF as described in the previous section. Two replicates of 30 A. aegypti males were dusted with PPF and released into a cage (30 cm × 30 cm × 30 cm). For comparison, two additional replicates of 30 males were left undusted, and survivorship was then monitored in all four cages.
A second study examined for the transfer of PPF directly from males into oviposition containers. To exclude the possibility of indirect PPF transfer to breeding sites, i.e., females delivering PPF into the water after having become PPF contaminated by males, no females were introduced into the mesocosm cage for this experiment. Two hundred and fifty dusted A. aegypti males were released into a mesocosm cage (L × W × H; 4 m × 3.5 m × 2.2 m) containing five oviposition containers (16 oz. Plastic Container, 128HRD16 COMBO240, webstaurant.com) containing 100 mL water and lined with seed germination paper (Anchor Paper Company, St. Paul, Minnesota). As a control, 250 untreated males were released into a second, identical mesocosm cage. Both mesocosm cages were housed in a climate-controlled greenhouse (25.5 ± 2.9 °C; RH 74.2 ± 4.1%). Five days after mosquito introduction, all mosquitoes were removed from each of the mesocosm cages. At the same time, the oviposition containers were also removed, and 25 mL samples of water from each were examined using the Kentucky (KY) bioassay method: For the KY bioassay, three samples were placed in 60 mL plastic cups (Dart, Mason, Michigan) with four L3 larvae and three drops of a 60 g/L liver powder slurry. Bioassays measured the number of emerging adults and dead immatures in each cup.
A third experiment assessed the indirect transfer of PPF from males to females. In this experiment, 250 dusted males were placed into the mesocosm cage along with 100 undusted females. As a control, 250 undusted males were released into an identical mesocosm cage along with 100 undusted females. After four days, the females were individually aspirated using different aspirator tips to avoid contamination from both cages and individualized into separate containers to minimize any subsequent transfer of PPF after removal from cages. To assess for PPF on the undusted females, each female was individually examined using the above-described KY lab bioassay method.
2.2. Mosquito Shipment
Open release trials utilized successful shipments of
A. aegypti males that were produced and dusted in Kentucky. The males were transported via commercial courier in cardboard mailing tubes. As a source of water and food during shipment, cotton balls soaked with 10% sucrose solution were placed at both ends of each tube (
Figure 1). One thousand adult male
A. aegypti mosquitoes were anesthetized with chloroform (Fisher Scientific), counted, and placed in each tube. For each shipment, tubes were positioned in a 17.5 cm × 22.5 cm × 30 cm Styrofoam cooler, along with a water-moistened paper towel and a Hobo sensor to monitor conditions (Onset Computer Corporation, Bourne, Massachusetts;
Figure 1). The sensors tracked temperature, light, and humidity during transit for all shipments, with one exception, where a temperature sensor malfunctioned. Coolers were shipped overnight, and in no shipment did the mosquitoes remain in the package for more than 24 hours [
23]. In total, nine shipments were made, with 5–6 tubes each. The shipment process allowed for measurement of mosquito survivorship during cross-country delivery by the commercial courier. After the mosquitoes were released from tubes in California, the number of dead males remaining in the tubes was counted and recorded.
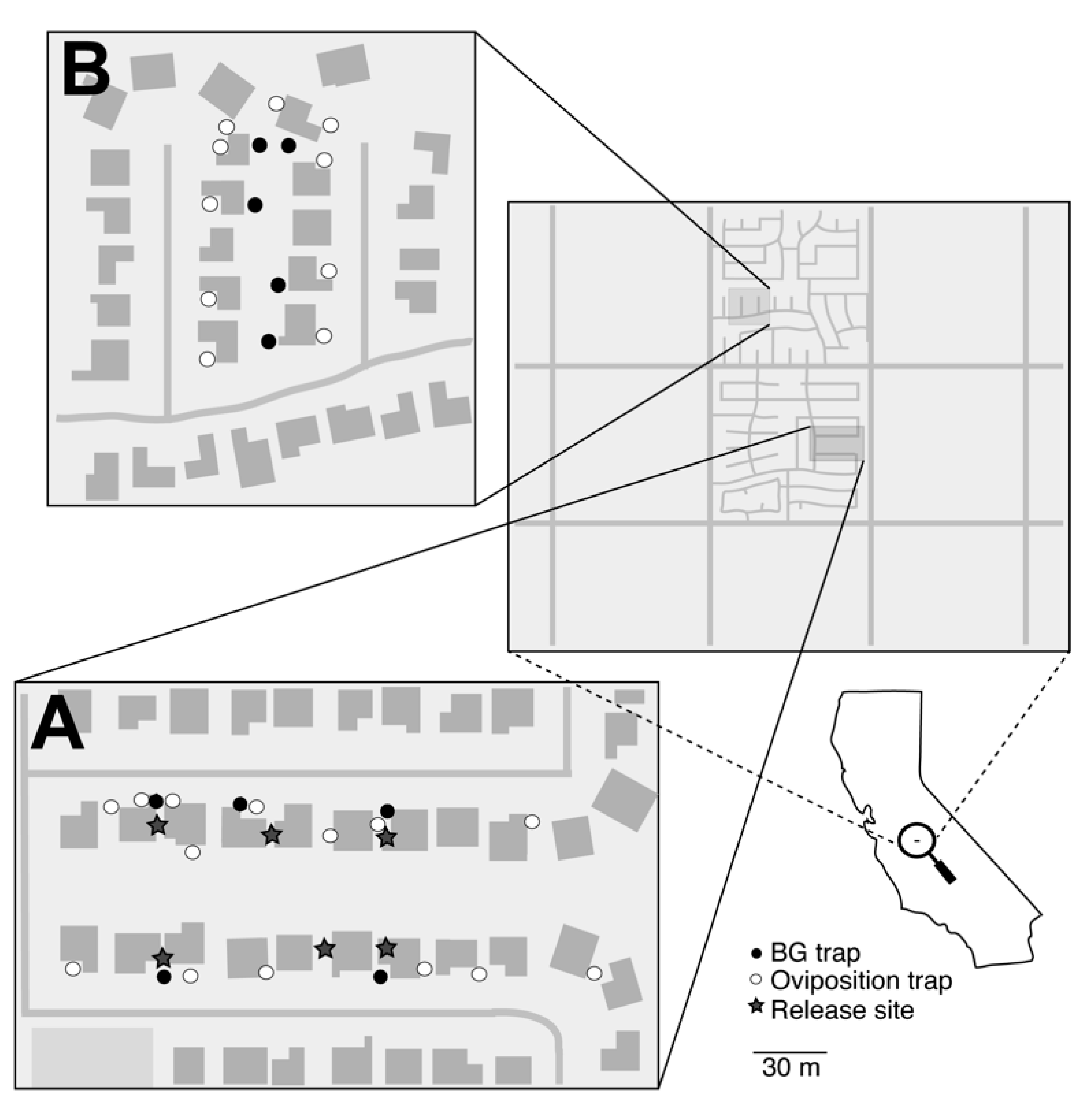
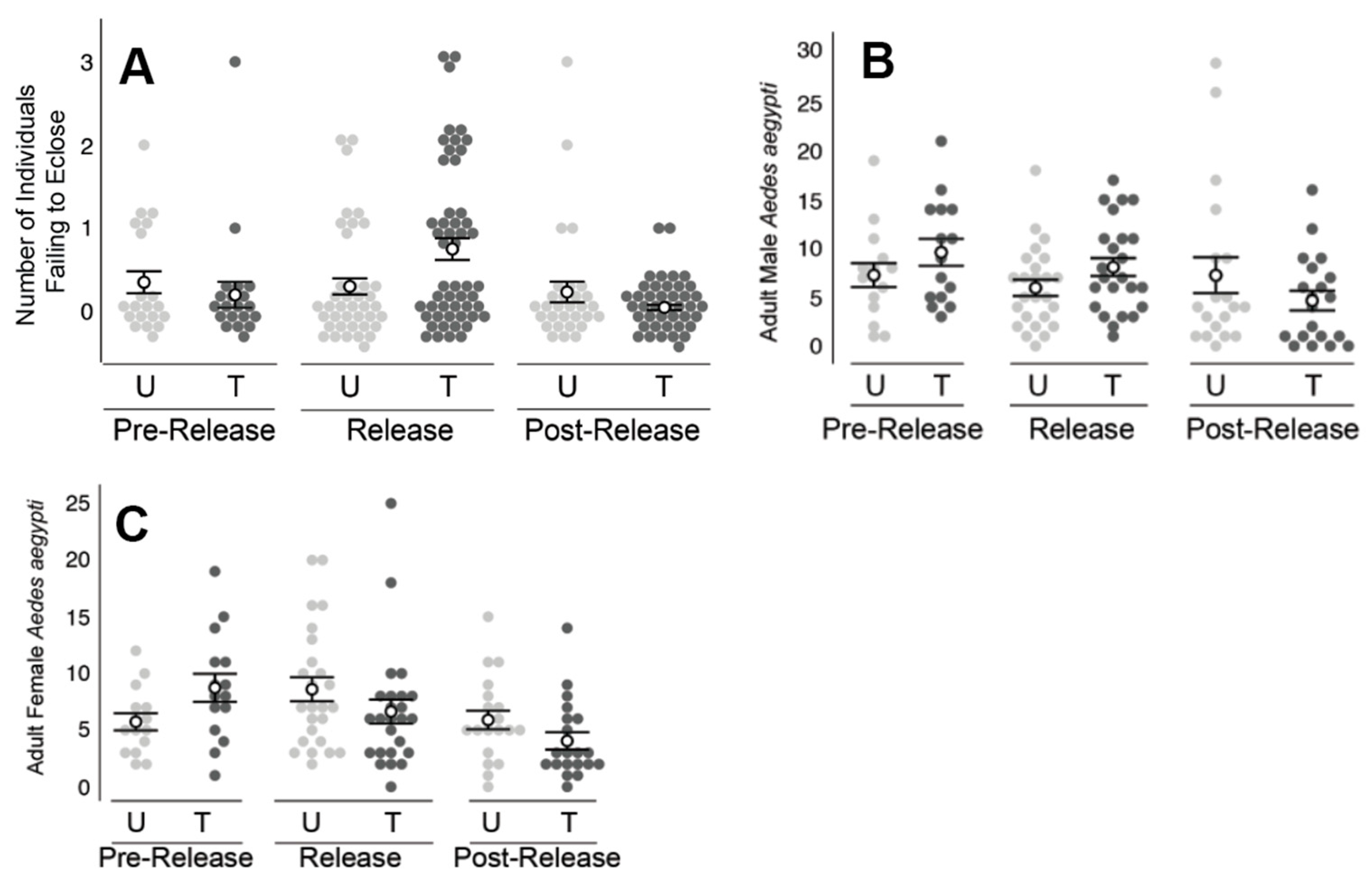
2.3. Fresno/Clovis, CA Field Releases
Clovis, CA has an arid climate with little rainfall (~311 mm annually) throughout the year. The
A. aegypti populations targeted in Clovis were recently established and still expanding at the time of the trial [
24]. The treated and untreated areas in Clovis, CA together encompassed approximately five acres of urban residential neighborhood (
Figure 2). Releases at the treated area were approved by the California Department of Pesticide Regulation [
25]. To raise local awareness about the project, the Consolidated Mosquito Control District produced an educational online video (
http://mosquitobuzz.net/AedesaegyptiADAM.htm) and distributed informational flyers prior to the trial. In addition, several local media outlets produced articles and short news segments intended to inform the public.
Prior to the release, 14 and 10 artificial oviposition sites (ovisites) were placed at the treated and untreated areas, respectively. Each ovisite consisted of a 24-ounce glass mason jar (Jarden Home Brands) painted with black enamel paint (Rust-Oleum Inc.) and held in place with a metal tent peg (Essentials Tool Inc.). Five Biogents sentinel (BG) traps (Biogents AG, Regensburg, Germany) were baited with CO
2 (dry ice, 1–1.5 kg) and a BG lure (Biogents AG, Regensburg, Germany) and placed within each of the treated and untreated areas (
Figure 2). Ovisites were placed in either the front or back yard of residents’ homes, out of direct sunlight and away from yard irrigation. Cups (16 oz. Black Stadium Cups, Promotion Choice) were lined with germination paper (400PT Non-Toxic Paper Toweling; Seedburo) and placed in small ceramic pots to prevent them from tipping over. Finally, to avoid disturbances from animals, large gauge mesh netting (DeerBlock, Easy Gardener Inc., Batesburg, SC, USA) was fitted over the top of each.
Groups of approximately 5000 PPF dusted males were released within the treatment area twice per week for a total of five weeks. Males were released by hand from the tubes near vegetation at multiple locations within the field site (
Figure 2). Typically, males were released immediately following their arrival by overnight transit to the release locations. Since the commercial shipment timing, male releases were typically performed in the early afternoon (i.e., noon–5pm). Due to repeated releases and the potential for transfer of dust to collected wild-type mosquitoes, including the potential for dust transfer occurring in trap nets, our experimental design did not include monitoring for ‘recaptured males,’ i.e., we did not perform a mark release recapture (MRR) type experiment due to potential artifacts caused by dust accumulation from repeated releases across multiple weeks. Adult populations were monitored weekly with BG traps that were baited with dry ice and a BG lure and were deployed for a single trap night (~24 hours). Ovisites were sampled weekly by transferring approximately 30 mL of water from each ovisite into a 60 ml plastic cup (Dart, Mason, Michigan). The seed germination paper was also removed each week, and cups were relined with new paper and refilled with tap water. Ovisite water samples were bioassayed in the lab using the California method, which consisted of adding four L3 larvae and 0.01g of ground TetraMin
® (Tetra, Spectrum Brands Pet, LLC, Blacksburg, VA, USA) (tropical flakes fish food) to the 30 mL of water collected from each ovisite. Bioassays recorded the number of emerging adults and dead immatures in each cup.
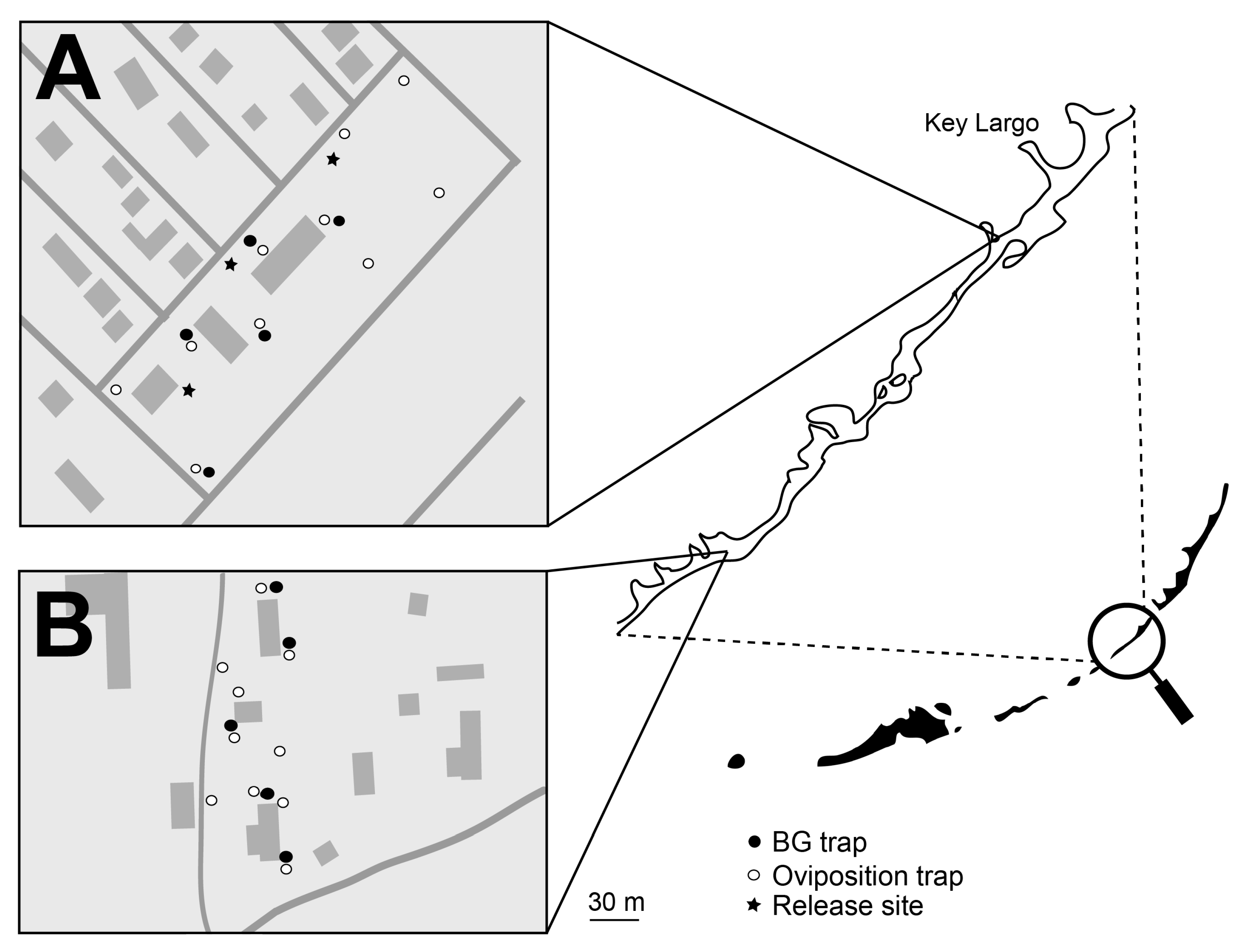
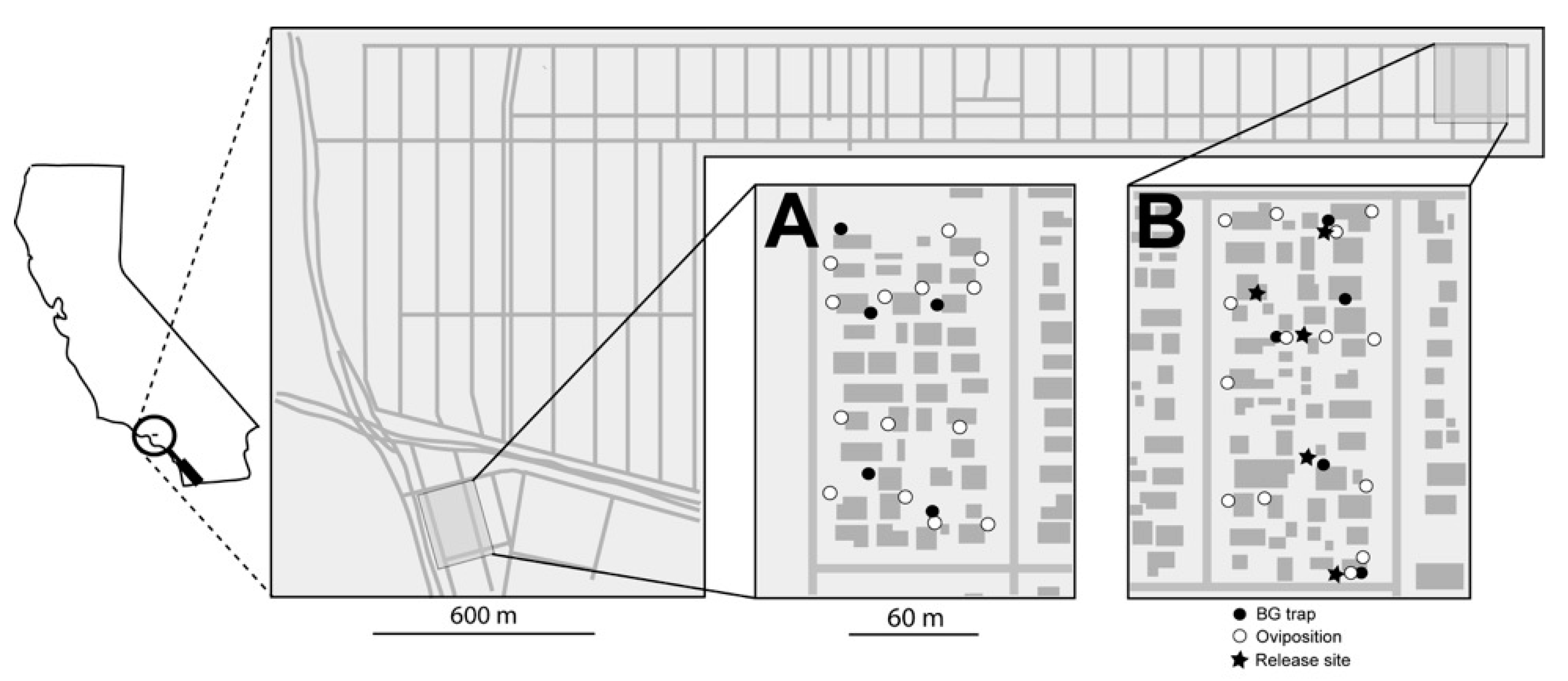
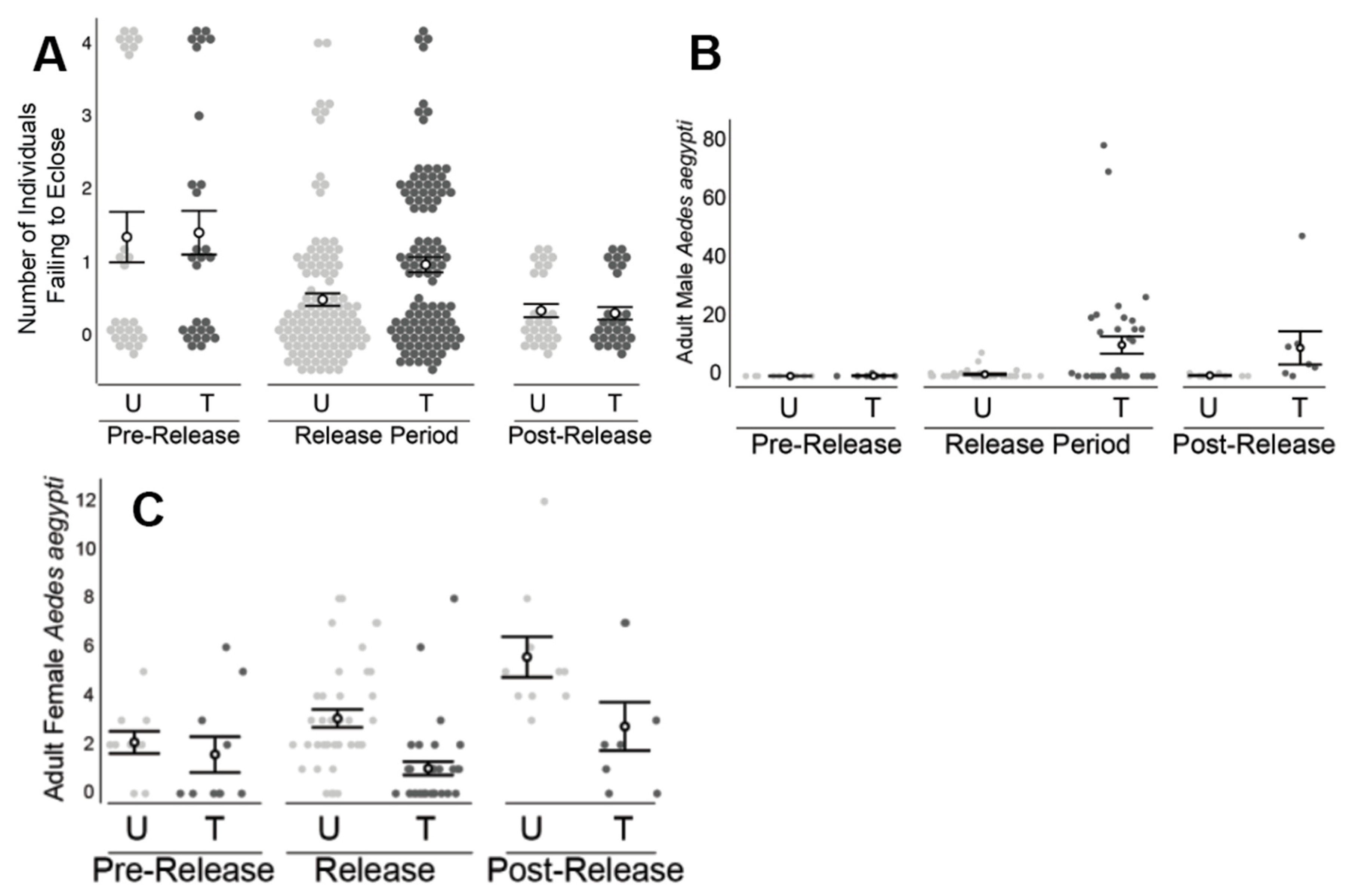
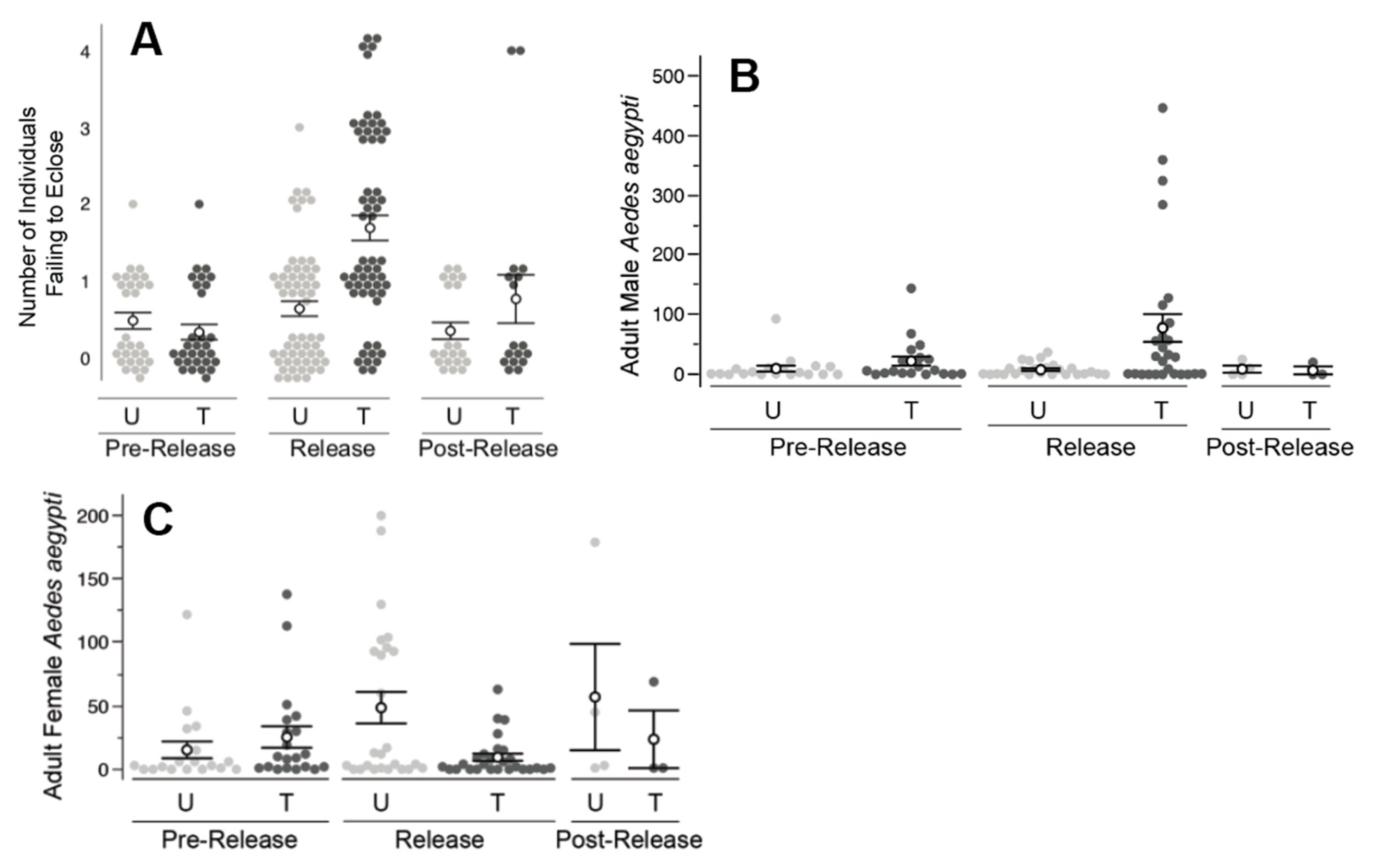
2.4. Florida Keys, FL and Los Angeles, CA Field Releases
The climate of the Florida Keys, FL is tropical with an annual precipitation rate of 1267 mm of rainfall per year.
A. aegypti populations with the Florida Keys are well established and can pose a significant health threat to local residents. Los Angles has a warm and temperate climate with rainy and dry seasons. As with Clovis, CA, the
A. aegypti populations targeted in Los Angeles were recently established and still expanding at the time of the trial [
24].
Treated and untreated areas were defined in the Florida Keys (
Figure 3) and in Los Angeles, CA (
Figure 4). Each area included approximately five and eight acres in Florida and Los Angeles, respectively. Ten ovisites and five BG Sentinel traps were placed at each area. In FL, BG traps were baited with dry ice and a BG lure and were set for ~24 hours, similar to the procedure utilized in Clovis, as described above. In Los Angeles, BG traps were baited with BG lures only (no CO
2). Ovisites were made from 700 mL glass jars (Ball Corporation, Broomfield, CO, USA) that were painted black externally using Flexseal (Flex Seal, Dayton, NJ, USA). These were filled with water, positioned near vegetation and lined with seed germination paper, which was removed and replaced weekly.
PPF-dusted A. aegypti males were released in an area within Los Angeles, CA, across an eight-week period. Male release methodology was analogous to that used in Clovis, CA. Twice per week, approximately 5000 PPF-dusted males were released at the treated area (i.e., total of 10,000 males/week) near vegetation at multiple locations within the field site. Ovisite water samples were collected weekly, and bioassays were performed using a modification on the KY bioassay method. Specifically, the standard method was followed, except samples were placed in 20 mL borosilicate glass scintillation vials (Kimble, Vineland, NJ, USA) with four L3 larvae and three drops of a 60 g/L liver powder slurry. In addition to monitoring during the eight-week release period, monitoring was also conducted during a two-week pre-release period and a two-week post-release period.
Concurrent with the Los Angeles study, PPF-dusted A. aegypti males were released at an area on Key Largo, FL. Male release methodology was analogous to that used in Clovis, CA. The overall design was similar to the Los Angeles study: Bioassays were performed with water samples from the treated and untreated areas, and male and female adult A. aegypti were monitored using BG traps. As with the Clovis study, male recapture assessments using the fluorescent dust were not part of the study design due to the potential transfer of dust between marked and unmarked specimens and artifacts from multiple releases of dusted males within the treated area. The two areas were monitored for a four-week pre-release period, a six-week release period, and a two-week post-release period. In the Key Largo study during the post-release period, approximately half of the traps were disrupted due to weather events.
2.5. Statistical Analyses
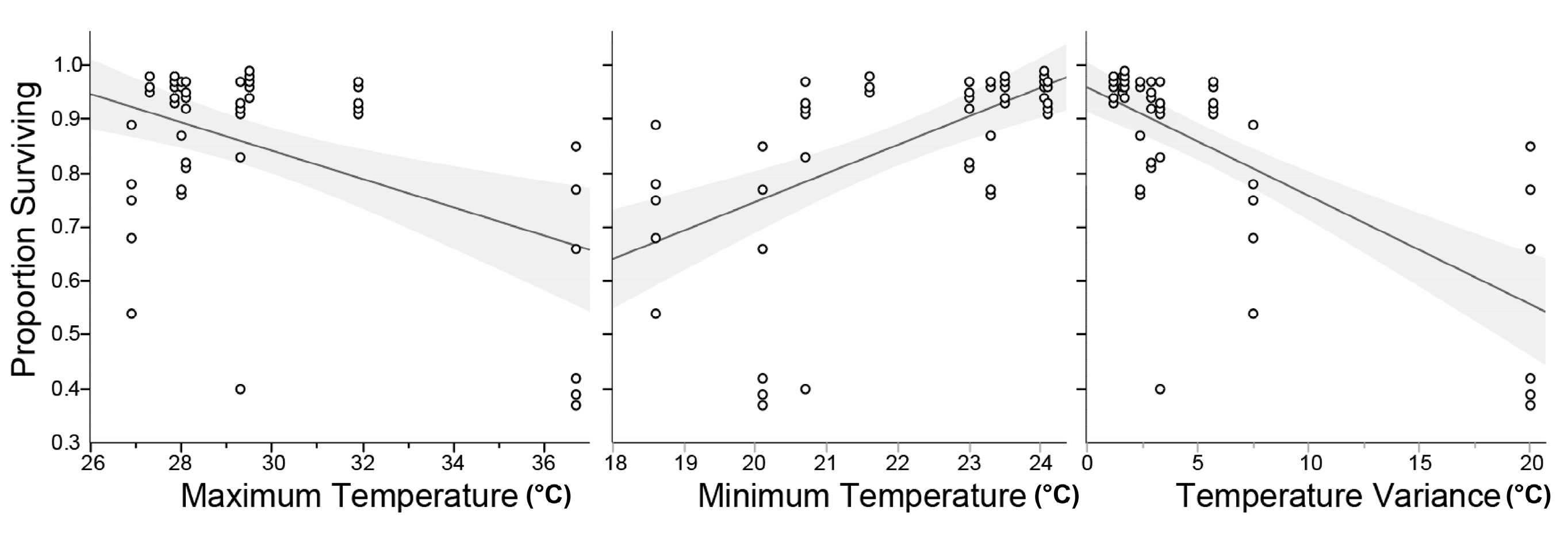
Statistical analyses were performed using JMP 12.1 (SAS Institute, Cary, NC, USA). Log-rank tests were used to compare mortality of treated and untreated males over time. ANOVA was used to examine for differences in dusted male survival, comparing between shipments; proportional survivorship data were arcsine transformed. Subsequently, bivariate correlations were used to examine for effects of temperature on male survival during shipment. Contingency analysis was used to compare the number of surviving adults in bioassays, and the Wilcoxon exact test was used to examine for differences in adult female and male number at field sites.
4. Discussion
While previous work has demonstrated the ability of
A. albopictus males to disseminate PPF [
17], this is the first report of the ADAM method with
A. aegypti populations. The primary goals of the work reported here were: 1) To determine the effects of PPF dusting on male survival, 2) to examine for an ability of ADAM dusted males to disseminate PPF both within artificial environments and in field trials, and 3) to examine for an impact of ADAM male releases on field populations of
A. aegypti females.
Laboratory examination of adult male A. aegypti survival show slightly lower survival of PPF-dusted males, compared to undusted males. However, the effects were not acute, and males within both groups lived for greater than one month. While survival was not directly examined in the field, the results of field trials suggest that PPF-males do survive long enough to disseminate PPF in these conditions. ESTEEM 35WP is a wettable powder and that it includes ingredients (e.g., surfactants) that may be harmful to adult mosquitoes and, therefore, not ideal for the ADAM approach. Further work is needed to develop and test additional PPF formulations that may be less harmful to adult mosquitoes.
Female mosquitoes were intentionally excluded from the design of one mesocosm experiment reported here. This design was intended to examine for the direct dissemination of PPF by males. Male-dissemination in the absence of females would allow prophylactic treatment of potential larval sites before populations of female mosquitoes appear. As described previously [
17,
26,
27,
28], because artificially reared males can be produced at any time, the ADAM method is not necessarily reliant on naturally occurring mosquitoes. As a result, effective dissemination is not necessarily impacted by the rise and decline of the natural population. Unlike the prior
A. albopictus study [
17], the present study did not specifically examine for direct dissemination in the field, because female
A. aegypti were present throughout the release periods. In other words, because males transfer PPF to females, the possibility cannot be excluded that all field dissemination occurred indirectly, due to male PPF transferal to female
A. aegypti. To exclude this possibility, future studies might examine for dissemination early in the season, when female
A. aegypti are rare or absent. This suggestion would be similar to the approach employed with
A. albopictus in a previous report [
17].
Immature mosquitoes exposed to water collected from ovisites near ADAM male releases experienced significantly higher mortality in all three field trials (Clovis, Los Angeles, and Florida Keys). However, the Clovis study was unlike the other two studies in that an obvious impact on the adult population was not observed. The Clovis study also varied from the other two studies in that, despite the repeated release of PPF-dusted males, BG trap collections detected only a slightly increased number of males, relative to the untreated areas. The number of adult
A. aegypti females collected did not differ between the treated and untreated areas during the release period. It is of note, however, that the female number was significantly higher in the untreated area prior to the start of releases. As shown in
Figure 6, between the pre-release and release periods, the number of female
A. aegypti increased in the untreated area and decreased in the treated area, which suggests an ADAM effect on the
A. aegypti population. While additional studies are needed, these results indicate that an important indicator of success is the detection of elevated male recapture rates.
Similar to the Clovis work, field releases of PPF-dusted A. aegypti males in Los Angeles, CA and the Florida Keys also detected significantly higher mortality of immature mosquitoes exposed to water collected from ovisites near the releases of ADAM males, relative to water sampled from the untreated areas. The increased mortality was coincident with ADAM male releases and was not observed prior to the release of ADAM males nor immediately following the cessation of releases. The decrease in larval mortality following the cessation of releases may be due to the dilution of PPF in sample containers (e.g., due to rainfall and artificial watering of lawns). The types of containers sampled (i.e., small glass jars) were prone to drying out or wash out, particularly in the Clovis and Florida Keys field sites, respectively. Ovicups were often noted to be dry in Clovis when checked weekly. The observation of dry ovicups could also help explain the limited impact on the adult population, if dry ovicups were not attractive oviposition sites for A. aegypti.
In other ways, the results observed in Los Angeles and the Florida Keys adult collections differed from those observed in Clovis. Specifically, the male capture rates detected at both Los Angeles and the Florida Keys were much higher in the treated areas, and significant male increases were observed during the release period only. In other words, the treated and untreated pairs were not observed to differ in the number of A. aegypti females prior to the start of releases. Consistent with expectations for a population impact of ADAM males, a lower number of adult A. aegypti females was observed at the treated areas, and this decrease in the number of females was coincident with the start of male releases.
There are potential sources of variation that could have contributed to the observed results between the different field studies including: (1) The Clovis study was the first of the
A. aegypti trials described here, (2) different cooperating personnel operated at each of the field locations, and (3) used differing methods (e.g., BG trapping in Los Angeles did not include the use of CO
2), (4) the targeted populations of
A. aegypti differed between the cities [
29], (5) a different strain of
A. aegypti was used in Clovis relative to the other two studies, and (6) the weather and geography of Clovis differs relative to the other two areas. Future examination could increase the number of males released in Clovis and examine for an elevated male recapture rate and impact on the female
A. aegypti numbers. However, because fieldwork is currently constrained in scale by regulatory approvals, operating at a larger scale is not currently possible. At this small scale, immigration of females could have affected adult collection numbers, complicating interpretation of the experiment. Repeating the ADAM field studies in Clovis, Los Angeles, and the Florida Keys allowed us to examine the ADAM method in different environments, different types and densities of housing, and different climates, e.g., arid Clovis versus tropical Florida Keys. Furthermore, the overall
A. aegypti population densities are higher in the Florida Keys, where
A. aegypti have long been established, relative to Clovis and Los Angeles, where
A. aegypti has only recently established [
24]. Clearly, additional replication would be beneficial in assessing optimal male release rates, and the results reported here support the effectiveness of the ADAM approach for PPF dissemination across a range of habitats. Investigations into field performance of ADAM mosquito strains could be studied in upcoming field assessments. Male performance (i.e., male competitiveness) is widely considered a key factor in a successful sterile insect technique (SIT) programs, and PPF dissemination rates in the field could vary between mosquito strains and local environmental factors at a release location.
Ideally, the ADAM method would employ locally reared males, which would preclude the need for shipping adult A. aegypti males. However, the results presented here show that male A. aegypti can remain useful for PPF dissemination following cross country shipment. While the dusting and shipment affect male longevity, the results demonstrate that a sufficient number of males survive and disseminate PPF. Since the conditions during shipment (temperature extremes and variation) affect male fitness, additional work can investigate improved methods for shipment (e.g., how to better maintain a desired temperature during shipment), if future work cannot be based on local production of ADAM males.
Lastly, the fieldwork reported here took place in areas that were not co-infested with
A. albopictus and
A. aegypti. If possible, future studies might examine areas with both
A. albopictus and
A. aegypti. In these areas, the release of one species could be examined for an intra- and inter-specific effect on immature development. This is possible because both
A. albopictus and
A. aegypti larvae occur in similar habitats and because both are ‘container breeders’ [
29]. Furthermore,
A. albopictus males have been shown to attempt mating with
A. aegypti females, known as ‘satyrization’ [
30,
31,
32,
33], which could result in the interspecific transfer of PPF from released
A. albopictus males to
A. aegypti. If this were demonstrated, then releases of one species could be used to control additional container-breeding mosquito species. The potential for interspecific effects also demonstrates the need to monitor for potential non-target impacts, e.g., non-Culicid insects that co-occur within the same containers.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







