Subterranean Acoustic Activity Patterns of Vitacea polistiformis (Lepidoptera: Sesiidae) in Relation to Abiotic and Biotic Factors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vineyard Layout and Grape Varieties
2.2. Procedures and Timing of Pheromone and Temperature Monitoring, Acoustic Recording, and Vine Inspection
2.3. Acoustic Signal Amplification, Sensor Placement, and Inspection of Soil around Sensor
2.4. Digital Signal Processing and Automated Classification of Vibration Impulses
2.5. Estimation of Infestation Likelihood
2.6. Statistical Analyses
3. Results
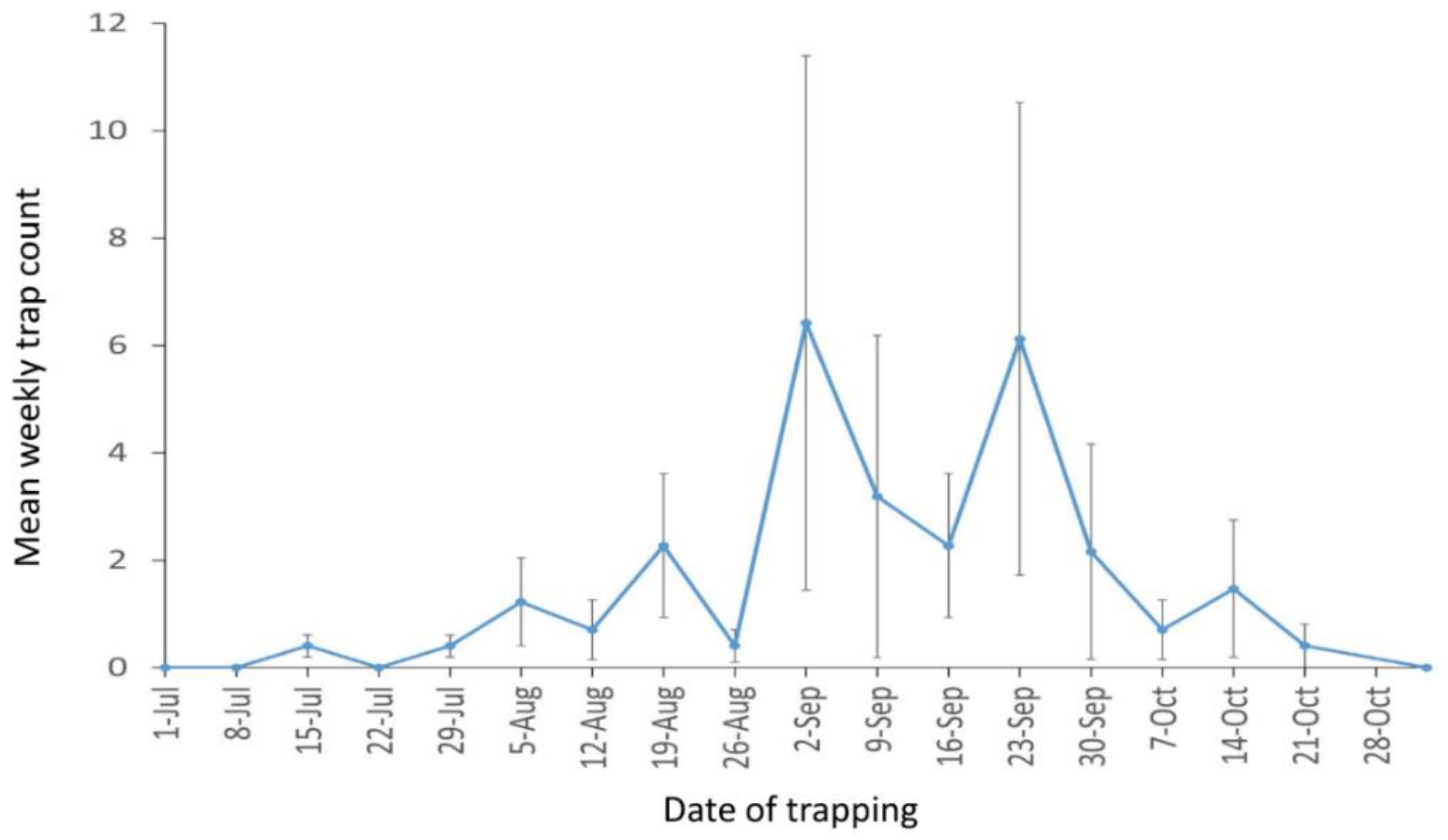
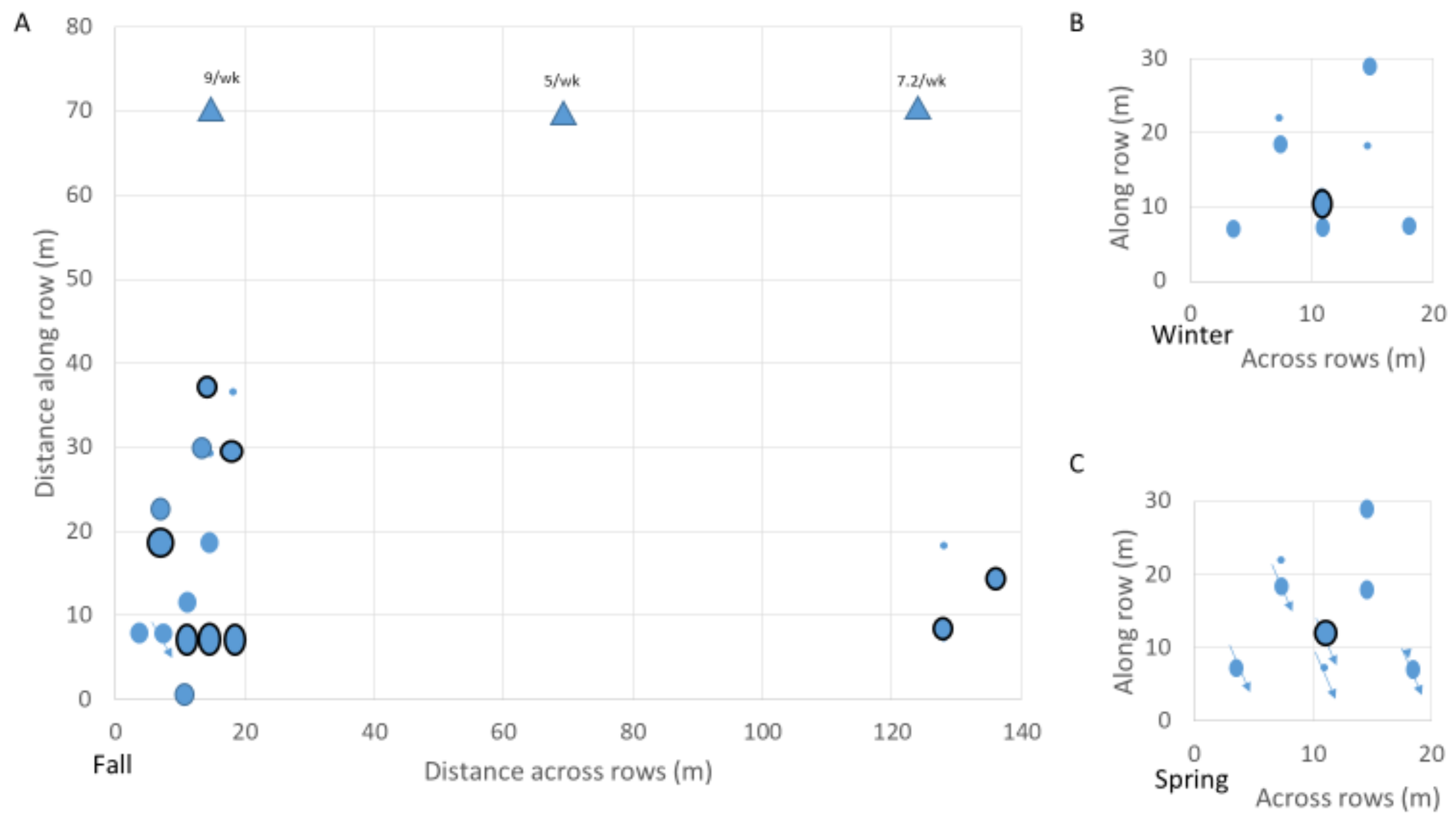
3.1. Pheromone Trap Monitoring and Inspection of Grape Roots and Soil around Sensor Waveguide
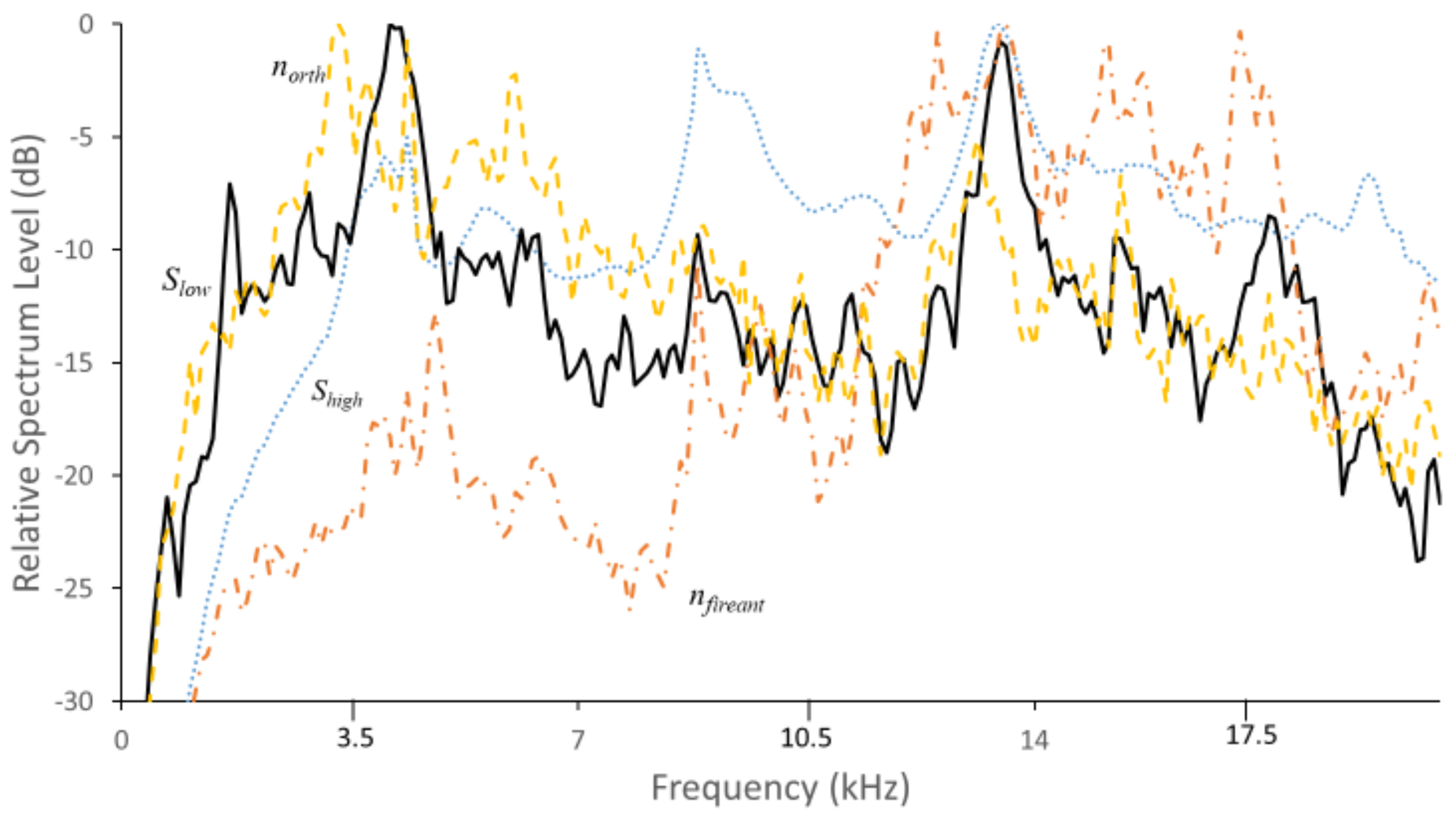
3.2. Mean Spectral Profiles
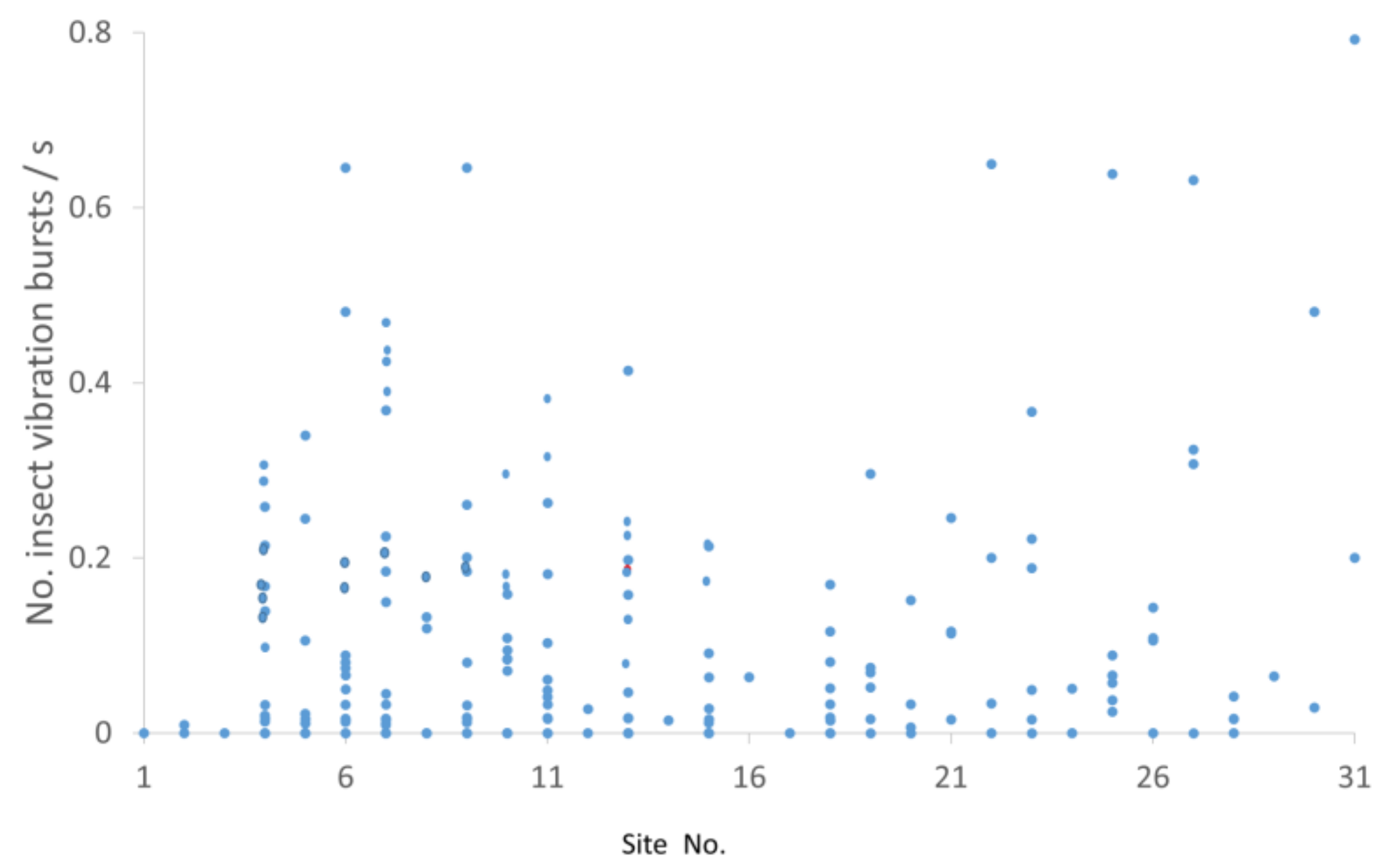
3.3. Insect Vibration Impulse Bursts
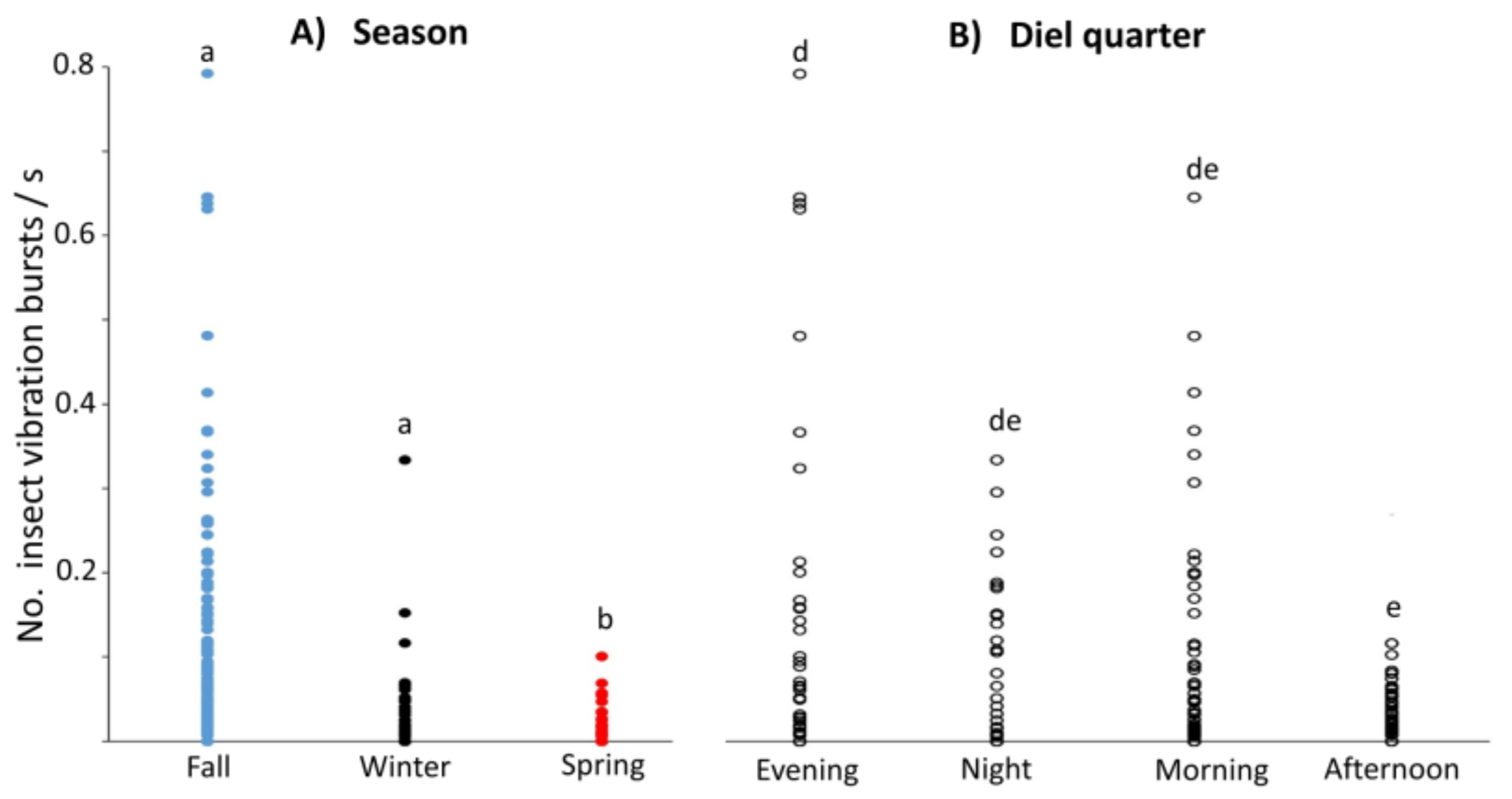
3.4. Mean Vibration Burst Rates at Different Sites across Seasons and Diel Quarters
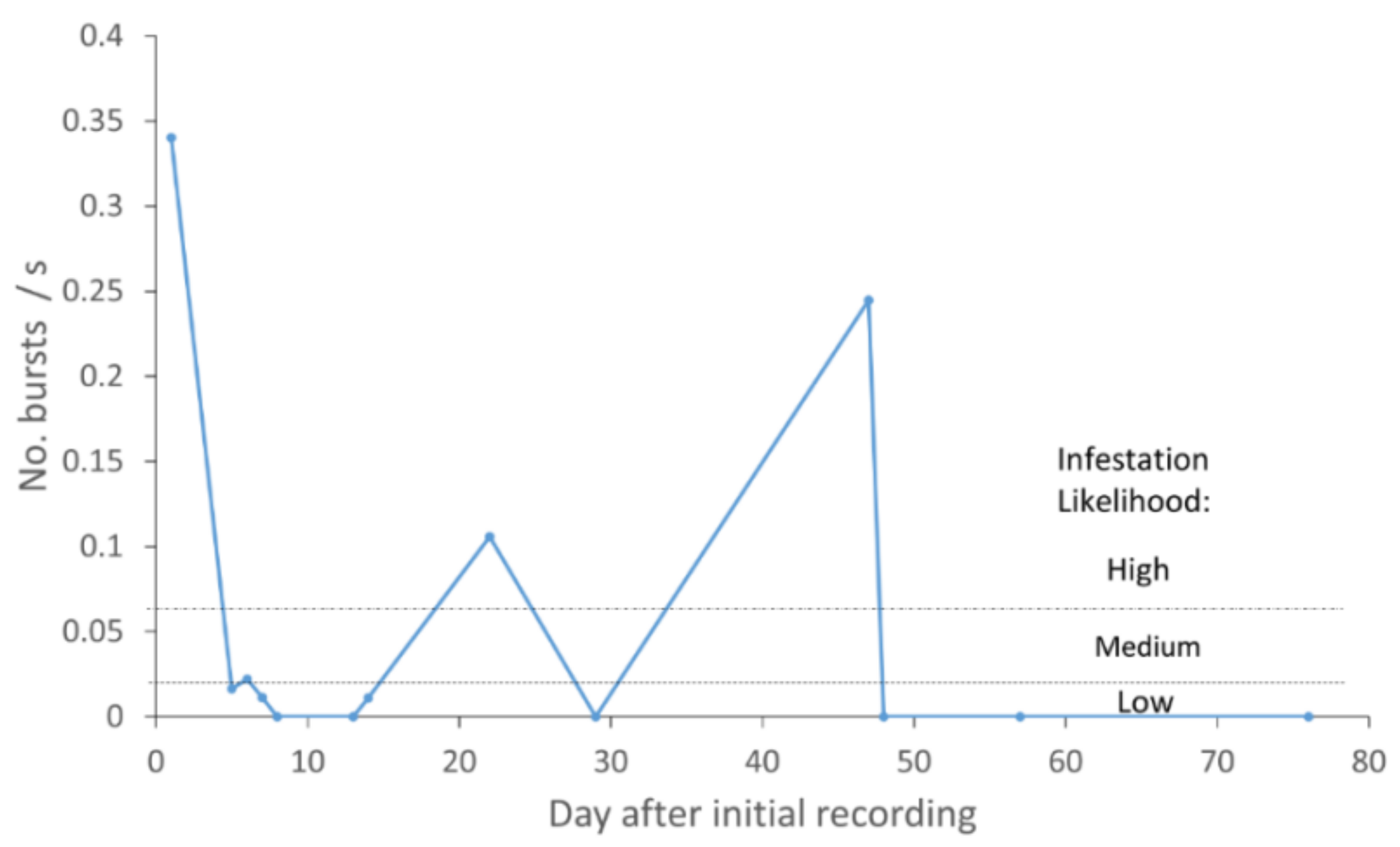
3.5. Spatial and Temporal Assessment of the Likelihood of Infestation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Snow, J.W.; Johnson, D.T.; Meyer, J.R. The seasonal occurrence of grape root borer Vitacea polistiformis (Harris) in the eastern United States. J. Entomol. Sci. 1991, 26, 157–168. [Google Scholar] [CrossRef]
- Bergh, J.C. Grape root borer. In Arthropod Management in Vineyards; Bostanian, N., Vincent, C., Isaacs, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 383–402. [Google Scholar]
- Rijal, J.P.; Bergh, J.C. Grape root borer (Lepidoptera: Sesiidae): An economic pest of commercial vineyards in the eastern United States. J. Integr. Pest Manag. 2017, 8, 30. [Google Scholar] [CrossRef]
- Liburd, O.; Razze, J.M.; Nyoike, T.W. Relative captures of grape root borer, Vitacea polistiformis (Lepidoptera: Sesiidae) in pheromone traps within vineyards and adjacent woodlands. Crop Prot. 2018, 105, 35–40. [Google Scholar] [CrossRef]
- Pearson, G.A.; Meyer, J.R. Female grape root borer (Lepidoptera: Sesiidae) mating success under synthetic sesiid sex pheromone treatment. J. Entomol. Sci. 1996, 31, 323–330. [Google Scholar] [CrossRef]
- Dutcher, J.D.; All, J.N. Damage impact of larval feeding of the grape root borer in a commercial Concord grape vineyard. J. Econ. Entomol. 1979, 72, 159–161. [Google Scholar] [CrossRef]
- Olien, W.C.; Smith, B.J.; Hegwood, C.P., Jr. Grape root borer: A review of the life cycle strategies for integrated control. HortScience 1993, 28, 1154–1156. [Google Scholar] [CrossRef]
- Weihman, S.W.; Liburd, O.E. Seasonal distribution and evaluation of two trap types for monitoring grape root borer Vitacea polistiformis (Lepidoptera: Sesiidae) in Florida vineyards. Fla. Entomol. 2006, 89, 245–250. [Google Scholar] [CrossRef]
- Sanders, W.R.; Liburd, O.E.; Mankin, R.W.; Meyer, W.L.; Stelinski, L.L. Applications and mechanisms of wax-based semiochemical dispenser technology for disruption of grape root borer mating. J. Econ. Entomol. 2011, 104, 939–946. [Google Scholar] [CrossRef]
- Webb, S.E.; Sprenkel, R.K.; Sharp, J.L. Seasonal flight activity of grape root borer (Lepidoptera: Sesiidae) in Florida. J. Econ. Entomol. 1992, 85, 2161–2169. [Google Scholar] [CrossRef]
- Rijal, J.P.; Zhang, A.; Bergh, J.C. Behavioral response of grape root borer (Lepidoptera: Sesiidae) neonates to grape root volatiles. Environ. Entomol. 2013, 42, 1338–1347. [Google Scholar] [CrossRef]
- Weihman, S. Monitoring and Control Tactics for Grape Root Borer Vitacea polistiformis Harris (Lepidoptera: Sesiidae) in Florida Vineyards. Master’s thesis, University of Florida, Gainesville, FL, USA, 2005. Available online: http://etd.fcla.edu/UF/UFE0009182/weihman_s.pdf (accessed on 22 August2019).
- Clark, G.N.; Enns, W.R. Life history studies of the grape root borer (Lepidoptera: Sesiidae) in Missouri. J. Kans. Entomol. Soc. 1964, 37, 57–63. [Google Scholar]
- Dutcher, J.D.; All, J.N. Survivorship of the grape root borer in commercial grape vineyards with contrasting cultural practices. J. Econ. Entomol. 1978, 71, 751–754. [Google Scholar] [CrossRef]
- Dutcher, J.D.; All, J.N. Models of the distribution of subterranean stages of Vitacea polistiformis in Concord grape vineyards. Environ. Entomol. 1978, 7, 461–465. [Google Scholar] [CrossRef]
- Sarai, D.S. Seasonal history and effect of soil moisture on mortality of newly hatched larvae of the grape root borer in southern Missouri. J. Econ. Entomol. 1972, 65, 182–184. [Google Scholar] [CrossRef]
- All, J.N.; Saunders, M.C.; Dutcher, J.D.; Javid, A.M. Susceptibility of grape root borer larvae, Vitacea polistiformis (Lepidoptera: Sesiidae) to Neoaplectana carpocapsae (Nematoda: Rhabditida): Potential of host kairomones for enhancement of nematode activity in grape vineyards. Misc. Publ. ESA 1981, 12, 9–14. [Google Scholar]
- Williams, R.N.; Fickle, D.S.; Grewal, P.S.; Meyer, J.R. Assessing the potential of entomopathogenic nematodes to control the grape root borer Vitacea polistiformis (Lepidoptera: Sesiidae) through laboratory and greenhouse bioassays. Biol. Sci. Tech. 2012, 12, 35–42. [Google Scholar] [CrossRef]
- Said, R.; Hix, R.L.; Reitz, S. Biological control of grape root borer (Lepidoptera: Sesiidae) with commercially available entomopathogenic nematodes in Florida muscadine and ‘Cynthiana’ grapes. J. Entomol. Sci. 2015, 50, 150–156. [Google Scholar] [CrossRef]
- Hix, R.L. Grape root borer control with nematodes: Vitis rotundifolia (Flugge) muscadines. Arth. Manag. Tests 2008, 33, C8. [Google Scholar]
- Hix, R.L. Grape root borer control with nematodes Vitis aestivalis Michx. ‘Cynthiana’. Arth. Manag. Tests 2009, 34, C17. [Google Scholar]
- All, J.N.; Dutcher, J.D.; Saunders, M.C. Control program for the grape root borer in grape vineyards of the eastern United States. Down Earth 1987, 43, 10–12. [Google Scholar]
- Vanegas, F.; Bratanov, D.; Powell, K.; Weiss, J.; Gonzalez, F. A novel methodology for improving plant pest surveillance in vineyards and crops using UAV-based hyperspectral and spatial data. Sensors 2018, 18, 260. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, J.D.; All, J.N. Predictive models for the summer activity of Vitacea polistiformis in Concord grape vineyards. Environ. Entomol. 1978, 7, 456–460. [Google Scholar] [CrossRef]
- Webb, S.; Mortensen, J.A. Evaluation of bunch grape rootstocks and muscadine varieties for resistance to grape root borer. Proc. Fla. St. Hort. Soc. 1990, 103, 310–313. [Google Scholar]
- Sanders, W.R.; Mankin, R.W.; Liburd, O.E.; Stelinski, L.L. Acoustic detection of arthropod infestation of grape roots: Scouting for grape root borer (Lepidoptera: Sesiidae). Fla. Entomol. 2011, 94, 296–302. [Google Scholar] [CrossRef]
- Brandhorst-Hubbard, J.L.; Flanders, K.L.; Mankin, R.W.; Guertal, E.A.; Crocker, R.L. Mapping of soil insect infestations sampled by excavation and acoustic methods. J. Econ. Entomol. 2001, 94, 1452–1458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Crocker, R.L.; Mankin, R.W.; Flanders, K.L.; Brandhorst-Hubbard, J.L. Acoustic identification and measurement of activity patterns of white grubs in soil. J. Econ. Entomol. 2003, 96, 1704–1710. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Hubbard, J.L.; Flanders, K.L. Acoustic indicators for mapping infestation probabilities of soil invertebrates. J. Econ. Entomol. 2007, 100, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Samson, P.R.; Chandler, K.J. Acoustic detection of Melolonthine larvae in Australian sugarcane. J. Econ. Entomol. 2009, 102, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Hagstrum, D.W.; Smith, M.T.; Roda, A.L.; Kairo, M.T.K. Perspective and promise: A century of insect acoustic detection and monitoring. Am. Entomol. 2011, 57, 30–43. [Google Scholar] [CrossRef]
- Jalinas, J.; Güerri-Agulló, B.; Mankin, R.W.; López-Follana, R.; Lopez-Llorca, L.V. Acoustic assessment of Beauveria bassiana (Hypocreales: Clavicipitaceae) effects on Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) larval activity and mortality. J. Econ. Entomol. 2015, 108, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Jalinas, J.; Güerri-Agulló, B.; Dosunmu, O.G.; Lopez-Llorca, L.V.; Mankin, R.W. Acoustic activity cycles of Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) early instars after Beauveria bassiana (Hypocreales: Clavicipitaceae) treatments. Ann. Entomol. Soc. Am. 2017, 110, 551–557. [Google Scholar] [CrossRef]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Effects of hermetic storage on adult Sitophilus oryzae L. (Coleoptera: Curculionidae) acoustic activity patterns. J. Econ. Entomol. 2017, 110, 2707–2715. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Oxygen consumption and acoustic activity of adult Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae: Bruchinae) during hermetic storage. Insects MDPI 2018, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Effects of hypoxia on acoustic activity of two stored product pests, adult emergence, and grain quality. J. Econ. Entomol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, A.; Fink, F.; Gogala, M.; Traue, D. Plants as transmission channels for insect vibrational songs. Behav. Ecol. Sociobiol. 1982, 11, 269–281. [Google Scholar] [CrossRef]
- Mankin, R.W.; Stanaland, D.; Haseeb, M.; Rohde, B.; Menocal, O.; Carrillo, D. Assessment of plant structural characteristics, health, and ecology using bioacoustic tools. Proc. Mtgs. Acoust. 2018, 33, 010003. [Google Scholar] [CrossRef]
- Hambric, S.A. Structural acoustics tutorial-Part 1: Vibrations in structures. Acoust. Today 2006, 2, 21–33. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Baurle, T.L.; Comas, L.H.; Nielsen, D.; Neilsen, G.H.; Lakso, A.N.; Smart, D.R. Seasonal patterns of root growth in relation to shoot phenology in grape and apple. Acta Hort. 2006, 721, 21–26. [Google Scholar] [CrossRef]
- Nassar, A.R.; Kliewer, W.M. Free amino acids in various parts of Vitis vinifera at different stages of development. Proc. Am. Soc. Hortic. Sci. 1966, 89, 281–294. [Google Scholar]
- Mahmud, K.P.; Holzapfel, B.P.; Guisard, Y.; Smith, J.P.; Nielsen, S.; Rogiers, S.Y. Circadian regulation of grapevine root and shoot growth and their modulation by photoperiod and temperature. J. Plant Physiol. 2018, 222, 86–93. [Google Scholar] [CrossRef]
- Mankin, R.W.; Moore, A. Acoustic detection of Oryctes rhinoceros (Coleoptera: Scarabaeidae: Dynastinae) and Nasutitermes luzonicus (Isoptera: Termitidae) in palm trees in urban Guam. J. Econ. Entomol. 2010, 103, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Al-Ayedh, H.Y.; Aldryhim, Y.; Rohde, B. Acoustic detection of Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) and Oryctes elegans (Coleoptera: Scarabaeidae) in Phoenix dactylifera (Arecales: Arecacae) trees and offshoots in Saudi Arabian orchards. J. Econ. Entomol. 2016, 109, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Charif, R.A.; Waack, A.M.; Strickman, L.M. Raven Pro 1.3 User’s Manual; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2008. [Google Scholar]
- Mankin, R.W.; Brandhorst-Hubbard, J.L.; Flanders, K.L.; Zhang, M.; Crocker, R.L.; Lapointe, S.L.; McCoy, C.W.; Fisher, J.R.; Weaver, D.K. Eavesdropping on insects hidden in soil and interior structures of plants. J. Econ. Entomol. 2000, 93, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Mankin, R.W.; Mizrach, A.; Hetzroni, A.; Levsky, S.; Nakache, Y.; Soroker, V. Temporal and spectral features of sounds of wood-boring beetle larvae: Identifiable patterns of activity enable improved discrimination from background noise. Fla. Entomol. 2008, 91, 241–247. [Google Scholar] [CrossRef]
- Dosunmu, O.G.; Herrick, N.J.; Haseeb, M.; Hix, R.L.; Mankin, R.W. Acoustic detectability of Rhynchophorus cruentatus (Coleoptera: Dryophthoridae). Fla. Entomol. 2014, 97, 431–438. [Google Scholar] [CrossRef]
- Görres, C.M.; Chesmore, D. Active sound production of scarab beetle larvae opens up new possibilities for species-specific pest monitoring in soils. Sci. Rep. 2019, 9, 10115. [Google Scholar] [CrossRef]
- SAS Institute Inc. Base SAS 9.4 Procedures Guide. Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- JMP. User Manual Version 14; SAS Institute Inc.: Cary, NC, USA, 2019. [Google Scholar]
- Rashid, T.; Chen, J.; Vogt, J.T.; McLeod, P.J. Arthropod prey of imported fire ants (Hymenoptera: Formicidae) in Mississippi sweetpotato fields. Insect Sci. 2013, 20, 467–471. [Google Scholar] [CrossRef]
- Rim, K.; Price, S.J.; Wenninger, E.J.; Long, R.; Ramirez, R.A. Biology and management of clover root curculio (Coleoptera: Curculionidae). J. Integr. Pest Manag. 2019, 10, 23. [Google Scholar] [CrossRef]
- Mankin, R.W.; Hagstrum, D.W.; Nansen, C.; Meikle, W.G. Almond moth oviposition patterns in continuous layers of peanuts. J. Stored Prod. Res. 2014, 59, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Rijal, J.P.; Brewster, C.C.; Bergh, J.C. Spatial distribution of grape root borer (Lepidoptera: Sesiidae) infestations in Virginia vineyards and implications for sampling. Environ. Entomol. 2014, 43, 716–728. [Google Scholar] [CrossRef]
- Rijal, J.P.; Brewster, C.C.; Bergh, J.C. Effects of biotic and abiotic factors on grape root borer (Lepidoptera: Sesiidae) infestations in commercial vineyards in Virginia. Environ. Entomol. 2014, 43, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Shade, R.E.; Furgason, E.S.; Murdock, L. Detection of hidden insect infestations by feeding-generated ultrasonic signals. Am. Entomol. 1990, 36, 231–234. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Flinn, P.W. Comparison of acoustical detection of several species of stored-grain beetles (Coleoptera: Curculionidae, Tenebrionidae, Bostrichidae, Cucujidae) over a range of temperatures. J. Econ. Entomol. 1993, 86, 1271–1278. [Google Scholar] [CrossRef]
- Mankin, R.W.; Shuman, D.; Weaver, D.K. Thermal treatments to increase acoustic detectability of Sitophilus oryzae (Coleoptera: Curculionidae) larvae. J. Econ. Entomol. 1999, 92, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Coupel-Ledru, A.; Lebon, E.; Christophe, A.; Gallo, A.; Gago, P.; Pantin, F.; Doligez, A.; Simonneau, T. Reduced nighttime transpiration is a relevant breeding target for high water-use in efficiency in grapevine. Proc. Natl. Acad. Sci. USA 2016, 113, 8963–8968. [Google Scholar] [CrossRef] [PubMed]
- Hotta, C.T.; Gardner, M.J.; Hubbard, K.E.; Baek, S.J.; Dalchau, N.; Suhita, D.; Dodd, A.N.; Webb, A.A.R. Modulation of environmental responses of plants by circadian clocks. Plant Cell Environ. 2007, 30, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.R.; Allen, M.F. Diurnal patterns of productivity of arbuscular mycorrhizal fungi revealed with the Soil Ecosystem Observatory. New Phytol. 2013, 200, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Hassell, M.P.; Southwood, T.R.E. Foraging strategies of insects. Annu. Rev. Ecol. Syst. 1978, 9, 75–98. [Google Scholar] [CrossRef]
- Behmer, S.T. Insect herbivore nutrient regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inyang, E.I.; Hix, R.L.; Tsolova, V.; Rohde, B.B.; Dosunmu, O.; Mankin, R.W. Subterranean Acoustic Activity Patterns of Vitacea polistiformis (Lepidoptera: Sesiidae) in Relation to Abiotic and Biotic Factors. Insects 2019, 10, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10090267
Inyang EI, Hix RL, Tsolova V, Rohde BB, Dosunmu O, Mankin RW. Subterranean Acoustic Activity Patterns of Vitacea polistiformis (Lepidoptera: Sesiidae) in Relation to Abiotic and Biotic Factors. Insects. 2019; 10(9):267. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10090267
Chicago/Turabian StyleInyang, Edidiong I., Raymond L. Hix, Violeta Tsolova, Barukh B. Rohde, Omotola Dosunmu, and Richard W. Mankin. 2019. "Subterranean Acoustic Activity Patterns of Vitacea polistiformis (Lepidoptera: Sesiidae) in Relation to Abiotic and Biotic Factors" Insects 10, no. 9: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10090267