Predation Pressure in Tea (Camellia sinensis) Plantations in Southeastern China Measured by the Sentinel Prey Method


Abstract
:1. Introduction
2. Materials and Methods
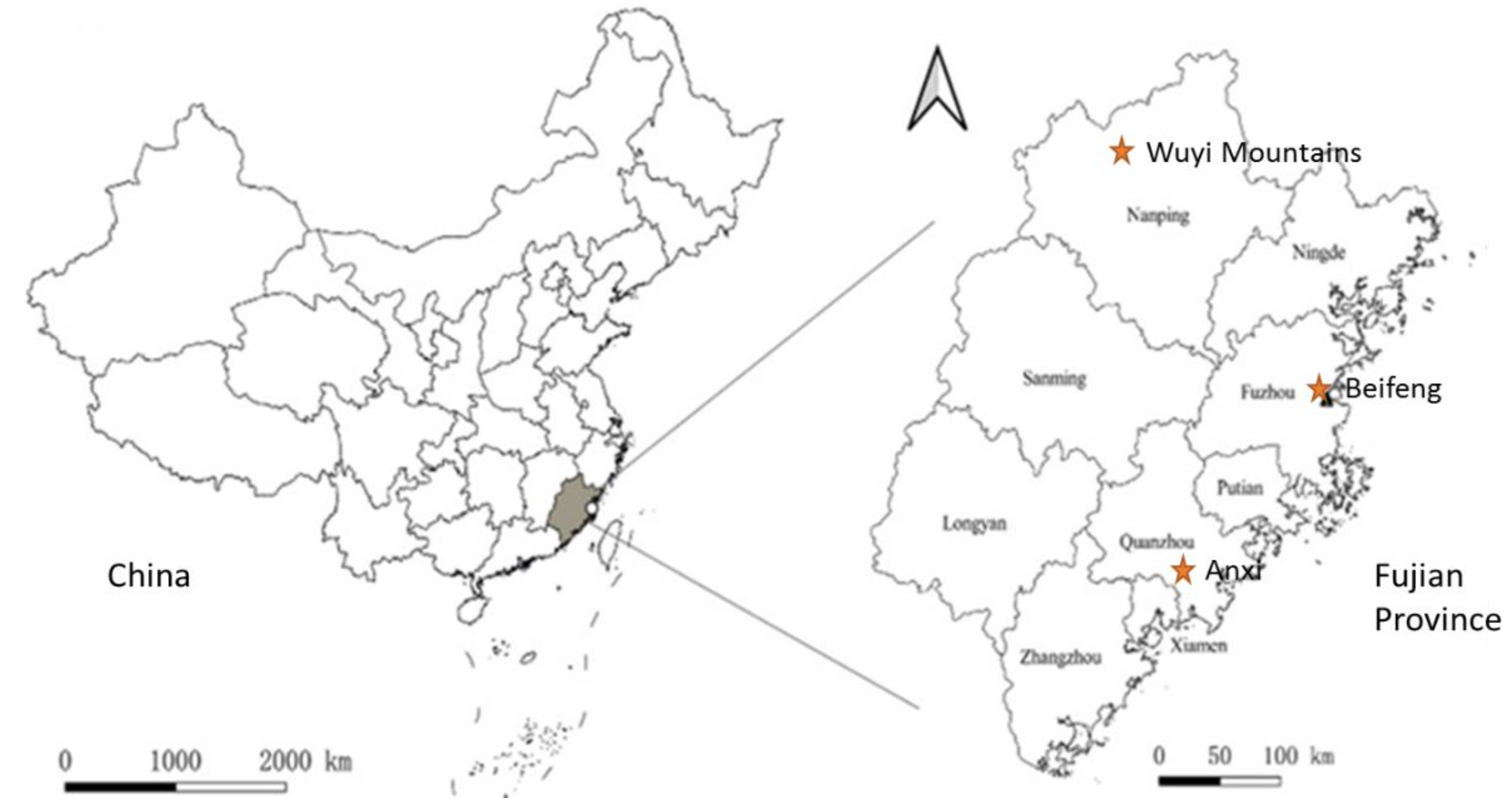
2.1. Study Sites
2.2. Artificial Caterpillars
2.3. Data Evaluation
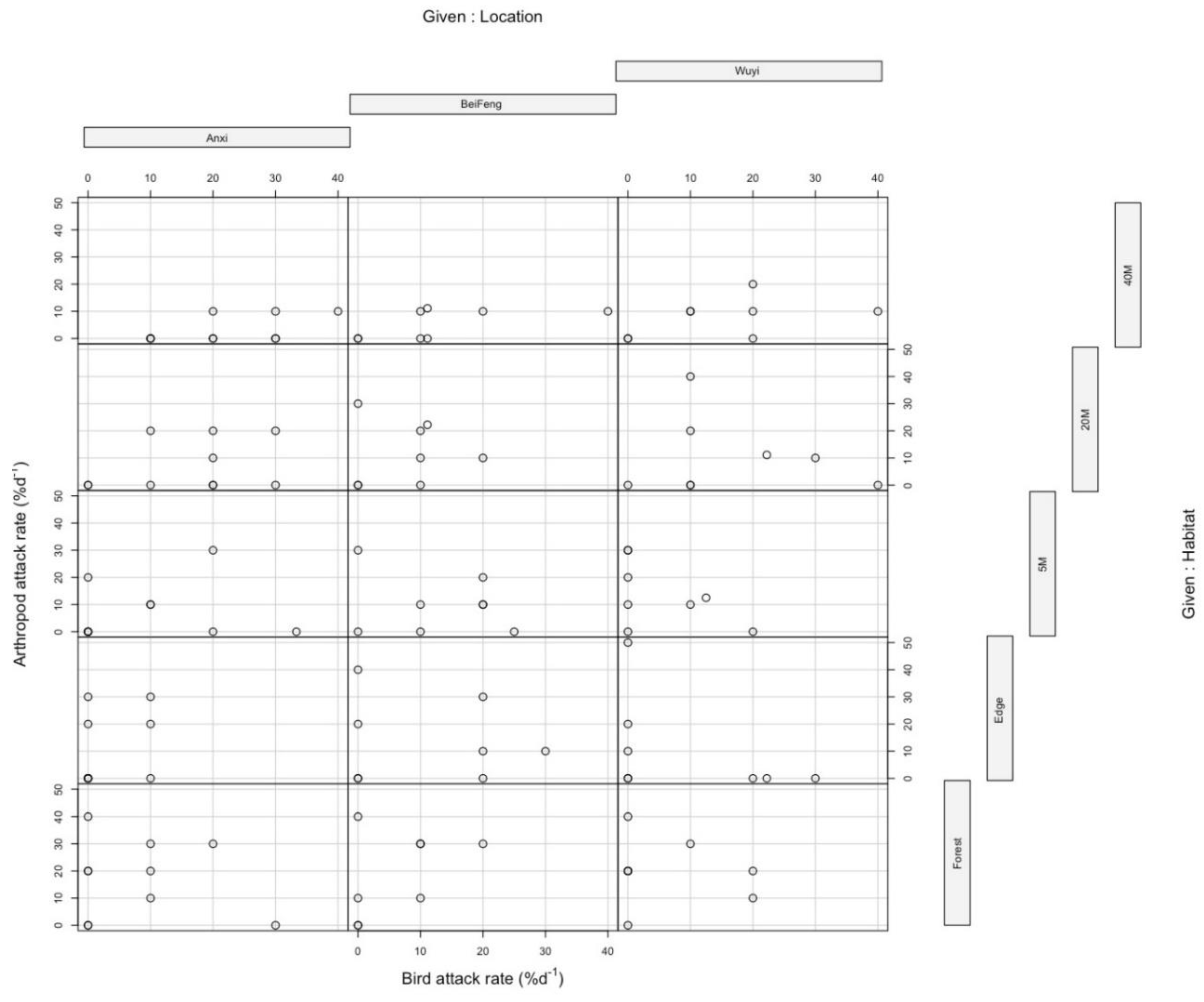
3. Results
3.1. Overall Predation Pressure
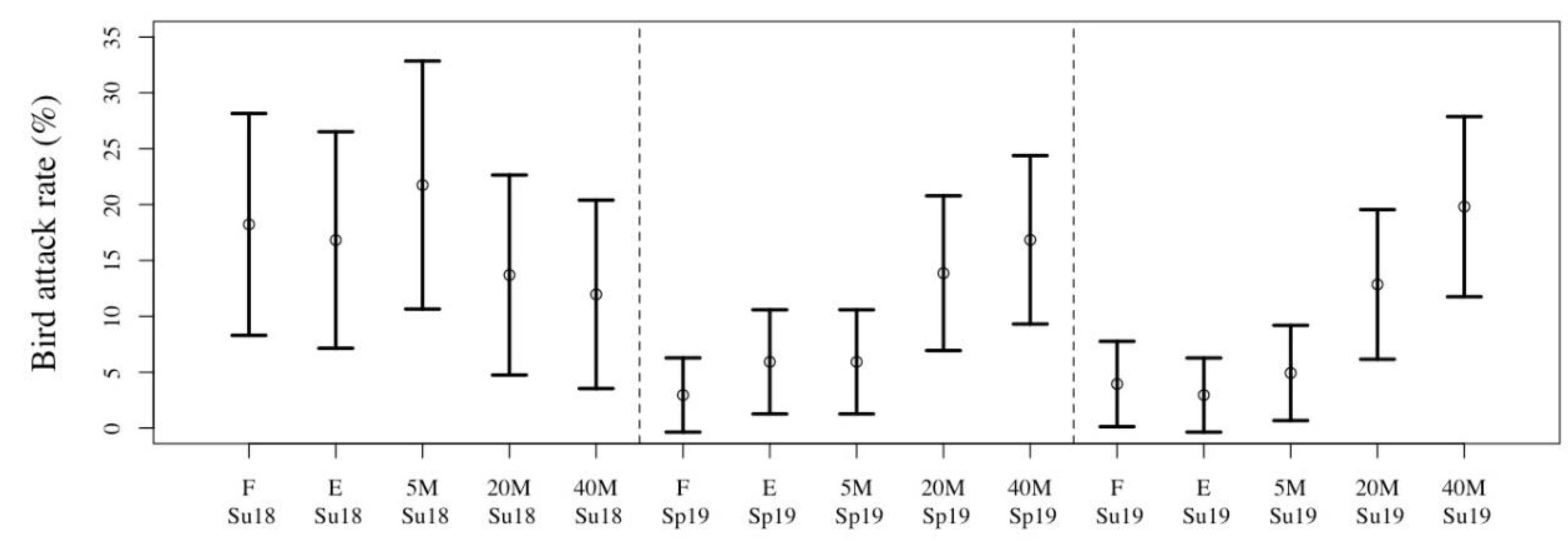
3.2. Seasonal Variation in Predation Pressure
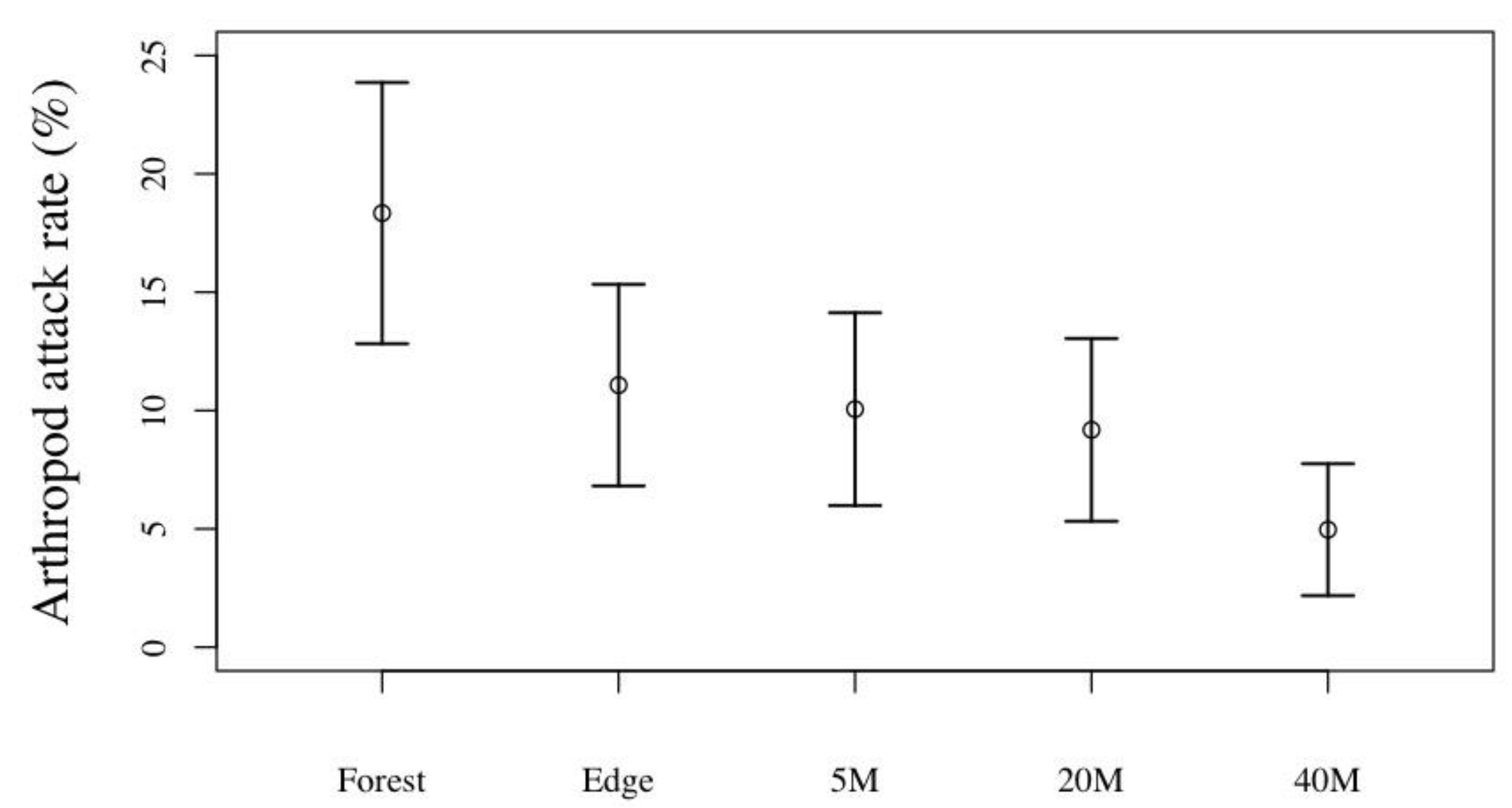
3.3. Predation Pressure in Forest Fragment vs. Tea Plantation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daily, G. Nature’s Services; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Díaz, S.; Pascual, U.; Stenseke, M.; Martín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.A.; Baste, I.A.; Brauman, K.A.; et al. Assessing nature’s contributions to people. Science 2018, 359, 270–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, R.S.; Mooney, H.A.; Agard, J.; Capistrano, D.; DeFries, R.S.; Diaz, S.; Dietz, T.; Duraiappah, A.K.; Oteng-Yeboah, A.; Pereira, H.M.; et al. Science for managing ecosystem services: Beyond the Millennium Ecosystem Assessment. Proc. Nat. Acad. Sci. USA 2009, 106, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimpel, G.E.; Mills, N.J. Biological Control. Ecology and Applications; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A functional overview of conservation biological control. Crop. Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M.-S. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef]
- Benn, J.A. Tea in China: A Religious and Cultural History; University of Hawai‘i Press: Honolulu, HI, USA, 2015; p. 288. [Google Scholar]
- Xiao, Z.; Huang, X.; Zang, Z.; Yang, H. Spatio-temporal variation and the driving forces of tea production in China over the last 30 years. J. Geogr. Sci. 2018, 28, 275–290. [Google Scholar] [CrossRef] [Green Version]
- Gurusubramanian, G.; Rahman, A.; Sarmah, M.; Ray, S.; Bora, S. Pesticide usage pattern in tea ecosystem, their retrospects and alternative measures. J. Environ. Biol. 2008, 29, 813–826. [Google Scholar]
- Hazarika, L.K.; Bhuyan, M.; Hazarika, N.B. Insect pest of tea and their management. Annu. Rev. Entomol. 2009, 54, 268–284. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, P.; You, M.S.; Pozsgai, G.; Ma, X.; Zhu, H.; Yang, G. Cover crops enhance natural enemies while help suppressing pests in a tea plantation. Ann. Entomol. Soc. Am. 2019, 112, 348–355. [Google Scholar]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. Roy. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Soh, M.C.K.; Sodhi, N.S.; Lim, S.L.H. High sensitivity of montane bird communities to habitat disturbance in Peninsular Malaysia. Biol. Conserv. 2006, 129, 149–166. [Google Scholar] [CrossRef]
- Meyer, S.T.; Koch, C.; Weisser, W.W. Towards a standardized Rapid Ecosystem Function Assessment (REFA). Trends Ecol. Evol. 2015, 30, 390–397. [Google Scholar] [CrossRef]
- Rusch, A.; Birkhofer, K.; Bommarco, R.; Smith, H.G.; Ekbom, B. Predator body sizes and habitat preferences predict predation rates in an agroecosystem. Basic Appl. Ecol. 2015, 16, 250–259. [Google Scholar] [CrossRef]
- Lövei, G.L.; Ferrante, M. A review of the sentinel prey method as a way of quantifying invertebrate predation under field conditions. Insect Sci. 2017, 24, 528–542. [Google Scholar] [CrossRef]
- Howe, A.; Lövei, G.L.; Nachmann, G. Dummy caterpillars as a simple method to asses predation rates of invertebrates in tropical acroecosystem. Entomol. Exp. Appl. 2009, 131, 325–329. [Google Scholar] [CrossRef]
- Ries, L.; Fletcher, R.J.; Battin, J.; Sisk, T.D. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef] [Green Version]
- Lövei, G.L.; Magura, T.; Tóthmérész, B.; Ködöböcz, V. The influence of matrix and edges on species richness patterns of ground beetles (Coleoptera, Carabidae) in habitat islands. Global Ecol. Biogeogr. 2006, 15, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Duelli, P.; Obrist, M.K. Regional biodiversity in an agricultural landscape: The contribution of seminatural habitat islands. Basic Appl. Ecol. 2003, 4, 129–138. [Google Scholar] [CrossRef]
- Blitzer, E.J.; Dormann, C.F.; Holzschuh, A.; Klein, A.M.; Rand, T.A.; Tscharntke, T. Spillover of functionally important organisms between managed and natural habitats. Agric. Ecosyst. Env. 2012, 146, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Low, P.A.; Sam, K.; Mcarthur, C.; Posa, M.R.C.; Hochuli, D.F. Determining predator identity from attack marks left in model caterpillars: Guidelines for best practice. Entomol. Exp. Appl. 2014, 152, 120–126. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 5 March 2020).
- R Studio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2018; Available online: http://www.rstudio.com/ (accessed on 5 March 2020).
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Selected Papers of Hirotugu Akaike; Parzen, E., Tanabe, K., Kitagawa, G., Eds.; Springer: Berlin, Germany, 1998; pp. 199–213. [Google Scholar]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Software 2016, 69, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, M.; Lövei, G.L.; Magagnoli, S.; Minarcikova, L.; Tomescu, E.L.; Burgio, G.; Cagan, L.; Ichim, M.C. Predation pressure in maize across Europe and in Argentina: An intercontinental comparison. Insect Sci. 2019, 26, 545–554. [Google Scholar] [CrossRef]
- Van Bael, S.A.; Sunshine, A.; Philpott, S.M.; Greenberg, R.; Bichier, P.; Barber, N.A.; Mooney, K.A.; Gruner, D.S. Birds as predators in tropical agroforestry systems. Ecology 2008, 89, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Bullock, J.M.; McCracken, M.; Chapman, R.E.; Ball, S.L.; Edwards, M.E.; Nowakowski, M.; Pywell, R.F. Spill-over of pest control and pollination services into arable crops. Agric. Ecosyst. Env. 2016, 231, 15–23. [Google Scholar] [CrossRef]
- Knapp, M.; Seidl, M.; Knappová, J.; Macek, M.; Saska, P. Temporal changes in the spatial distribution of carabid beetles around arable field-woodlot boundaries. Sci. Rep. 2019, 9, 8967. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, M.; Gonzalez, E.; Lövei, G.L. Predators do not spill over from forest fragments to maize fields in a landscape mosaic in central Argentina. Ecol. Evol. 2017, 7, 7699–7707. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L.; Tóthmérész, B. Various edge response of ground beetles in edges under natural versus anthropogenic influence: A meta-analysis using life-history traits. Acta Zool. Acad. Sci. Hung. 2019, 65, 3–20. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L. Environmental filtering is the main assembly rule of ground beetles in the forest and its edge but not in the adjacent grassland. Insect Sci. 2019, 26, 154–163. [Google Scholar] [CrossRef]
- Török, J. Food composition of nestling blackbirds in an oak forest bordering on an orchard. Opuscula Zool. 1981, 17–18, 145–156. [Google Scholar]
- Lövei, G.L.; Sunderland, K.D. The ecology of ground beetles. Annu. Rev. Entomol. 1996, 41, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Mansion-Vaquié, A.; Ferrante, M.; Cook, S.M.; Pell, J.K.; Lövei, G.L. Manipulating field margins to increase predation intensity in fields of winter wheat (Triticum aestivum). J. Appl. Entomol. 2017, 141, 600–611. [Google Scholar] [CrossRef] [Green Version]
- Parmenter, R.R.; MacMahon, J.A. Factors influencing species composition and population sizes in a ground beetle community (Carabidae): Predation by rodents. Oikos 1988, 52, 350–356. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location/Season/ Habitat | Attack Rates (% d−1) by | ||
|---|---|---|---|
| All Predators | Arthropods | Birds | |
| Wuyi | 23.0 ± 14.9 (40) | 12.3 ± 13.1 (40) | 10.4 ± 11.8 (40) |
| Beifeng | 22.0 ± 16.0 (40) | 11.8 ±12.4 (40) | 10.2 ± 10.0 (40) |
| Anxi | 21.5 ± 17.1 (50) | 8.8 ±11.5 (50) | 11.7 ± 11.6 (50) |
| Summer 2018 | 25.6 ± 12.2 (30) | 8.7 ±10.0 (30) | 16.6 ± 9.6 (30) |
| Spring 2019 | 19.0 ±17.3 (50) | 9.8 ±12.0 (50) | 9.2 ± 11.8 (50) |
| Summer 2019 | 23.0 ± 16.3 (50) | 13.2 ±13.6 (50) | 9.0 ± 10.4 (50) |
| Forest | 25.4 ± 16.3 (26) | 18.5 ±13.5 (26) | 6.9 ± 8.8 (26) |
| Forest edge | 18.9 ± 13.6 (26) | 11.2 ±14.8 (26) | 7.4 ± 10.5 (26) |
| Tea plantation, 5m | 19.4 ± 13.6 (26) | 10.1 ±11.0 (26) | 9.3 ± 10.1 (26) |
| Tea plantation, 20m | 24.1 ± 16.2 (26) | 9.4 ± 11.4 (26) | 13.6 ± 11.0 (26) |
| Tea plantation, 40m | 22.8 ± 16.3 (26) | 5.0 ± 5.9 (26) | 17.0 ± 12.2 (26) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imboma, T.S.; Gao, D.-p.; You, M.-s.; You, S.; Lövei, G.L. Predation Pressure in Tea (Camellia sinensis) Plantations in Southeastern China Measured by the Sentinel Prey Method. Insects 2020, 11, 212. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11040212
Imboma TS, Gao D-p, You M-s, You S, Lövei GL. Predation Pressure in Tea (Camellia sinensis) Plantations in Southeastern China Measured by the Sentinel Prey Method. Insects. 2020; 11(4):212. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11040212
Chicago/Turabian StyleImboma, Titus S., De-ping Gao, Min-sheng You, Shijun You, and Gabor L Lövei. 2020. "Predation Pressure in Tea (Camellia sinensis) Plantations in Southeastern China Measured by the Sentinel Prey Method" Insects 11, no. 4: 212. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11040212