Biological Traits of the Pincer Wasp Gonatopus flavifemur (Esaki & Hashimoto) Associated with Different Stages of Its Host, the Brown Planthopper, Nilaparvata lugens (Stål)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Conditions
2.2. Effects of Different Stages of BPHs on Parasitism, Host Feeding, and Offspring Development in Gonatopus flavifemur
2.3. Parasitism and Host-Feeding Functional Response of the Parasitoid Gonatopus flavifemur Associated with Different Stages of BPHs
2.4. Data Analyses
3. Results
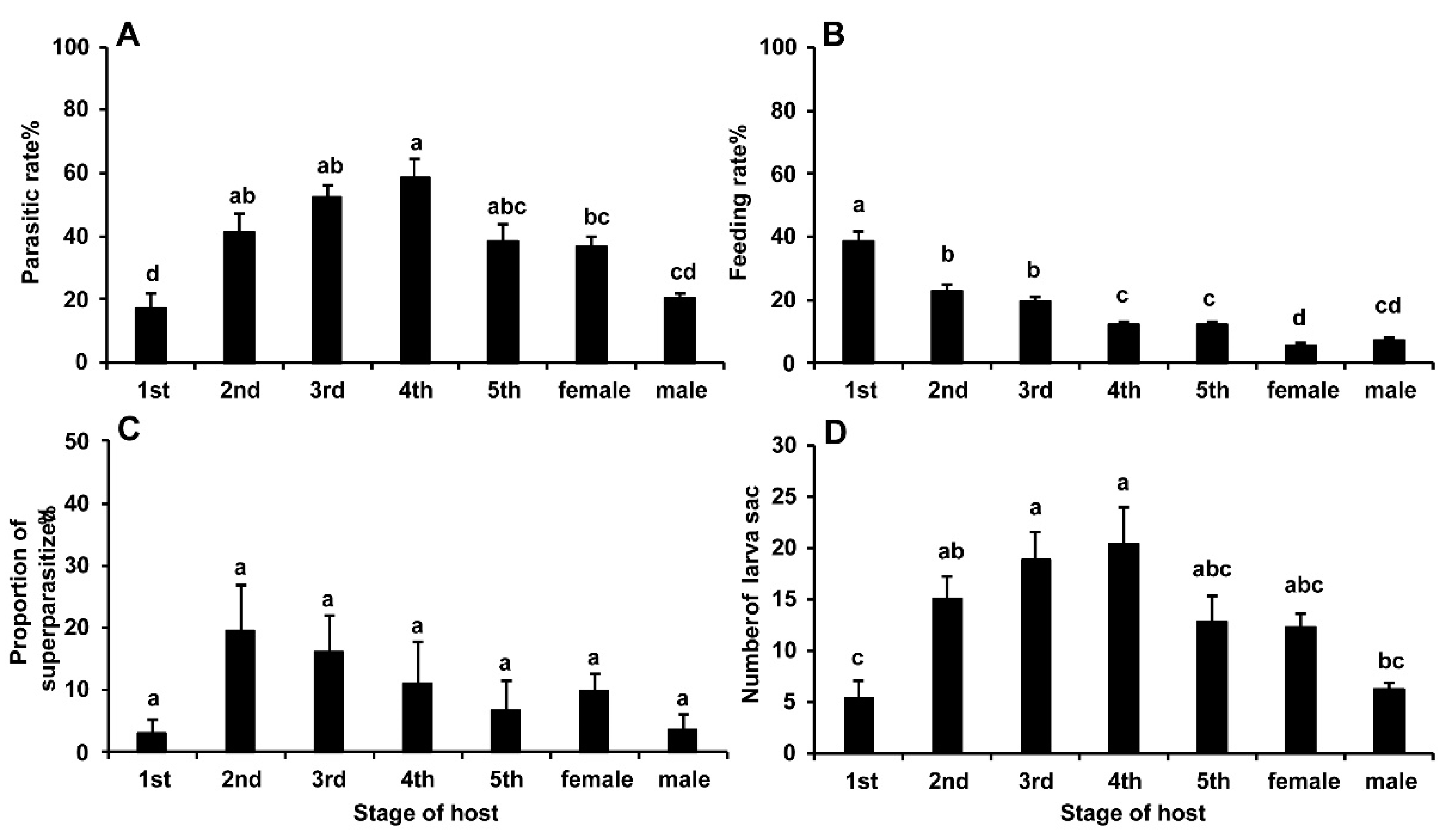
3.1. Effects of Different Stages of BPHs on Parasitism and Host-Feeding Rates in Gonatopus flavifemur
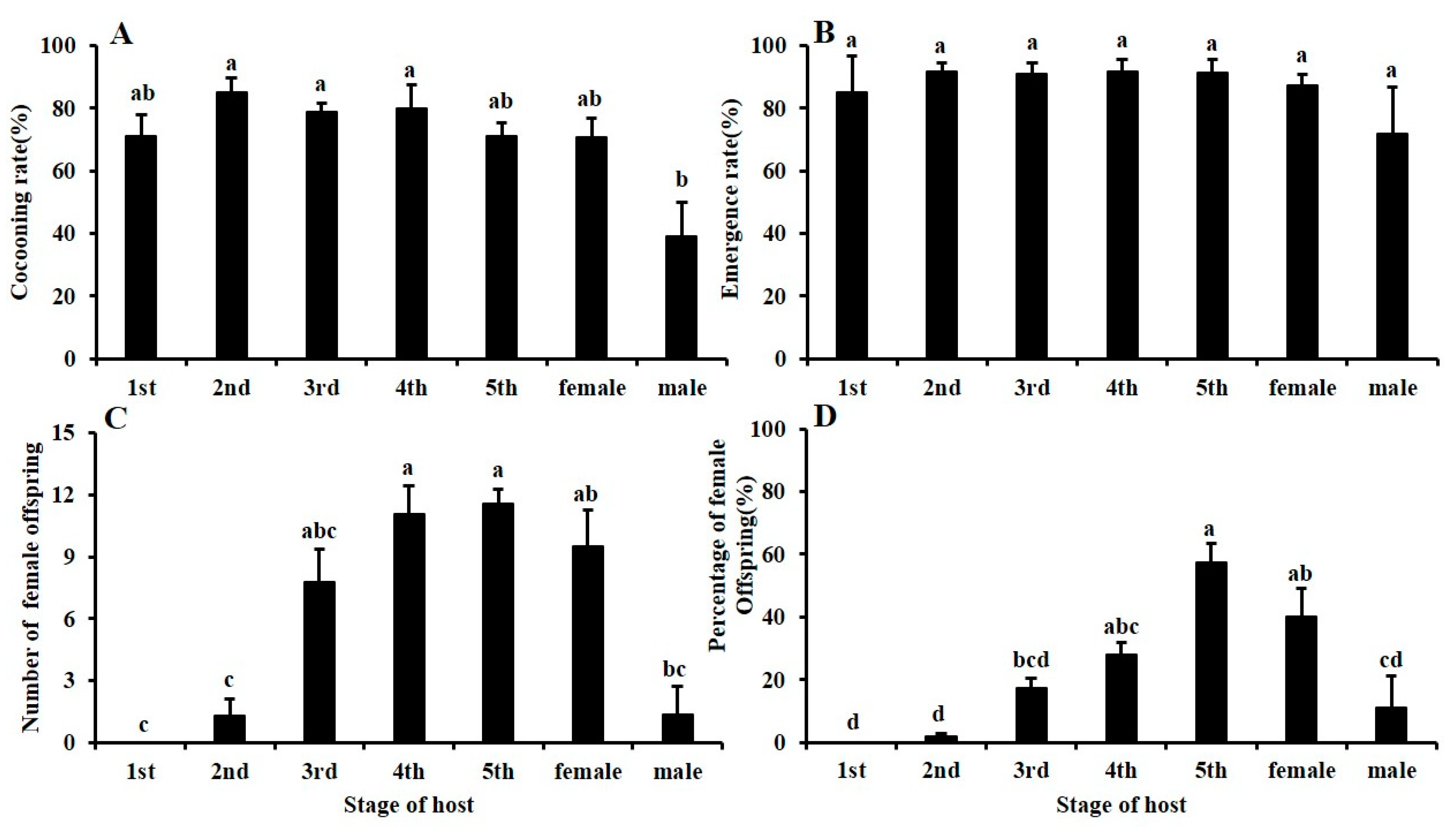
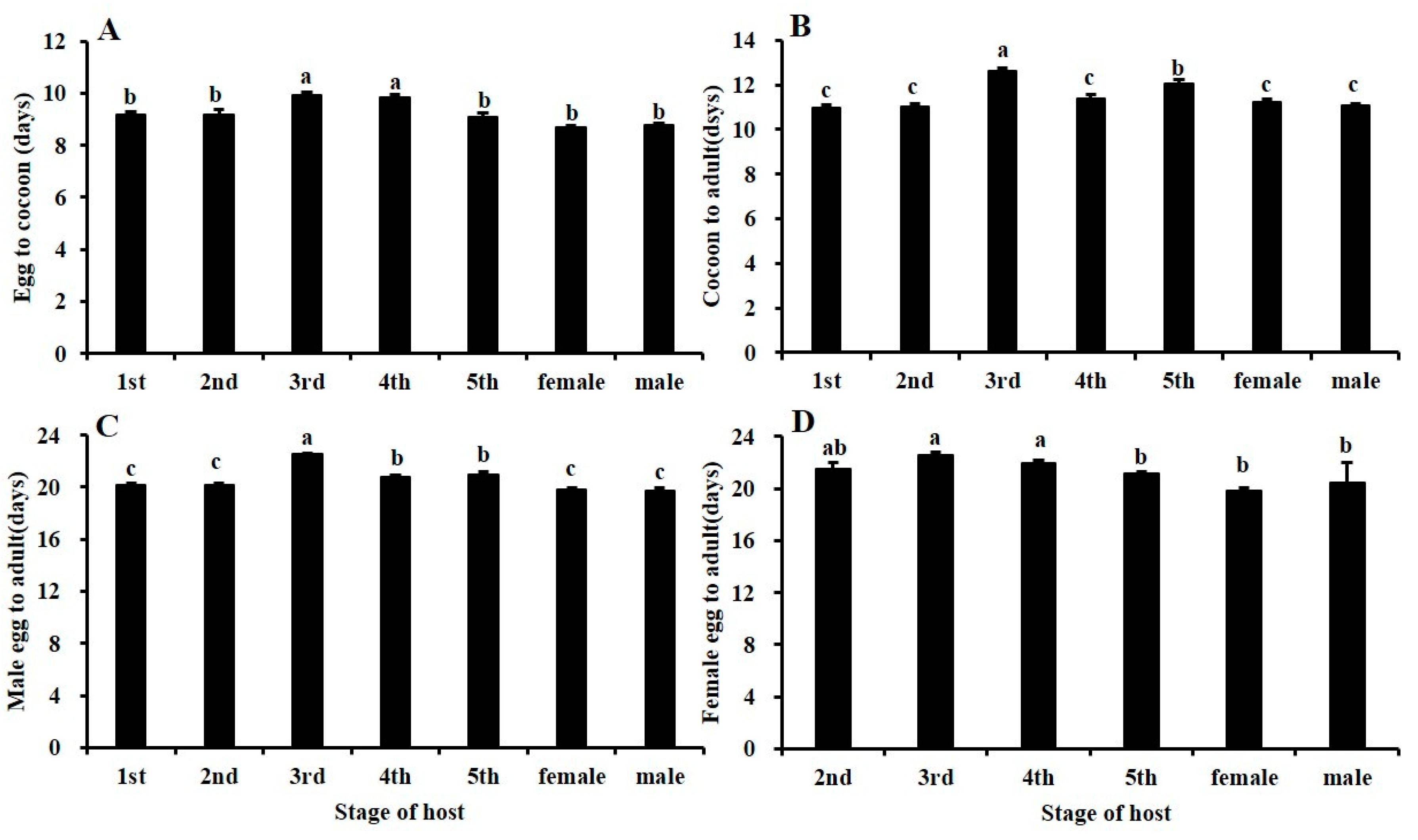
3.2. Effects of Different Stages of BPHs on Gonatopus flavifemur Offspring Development
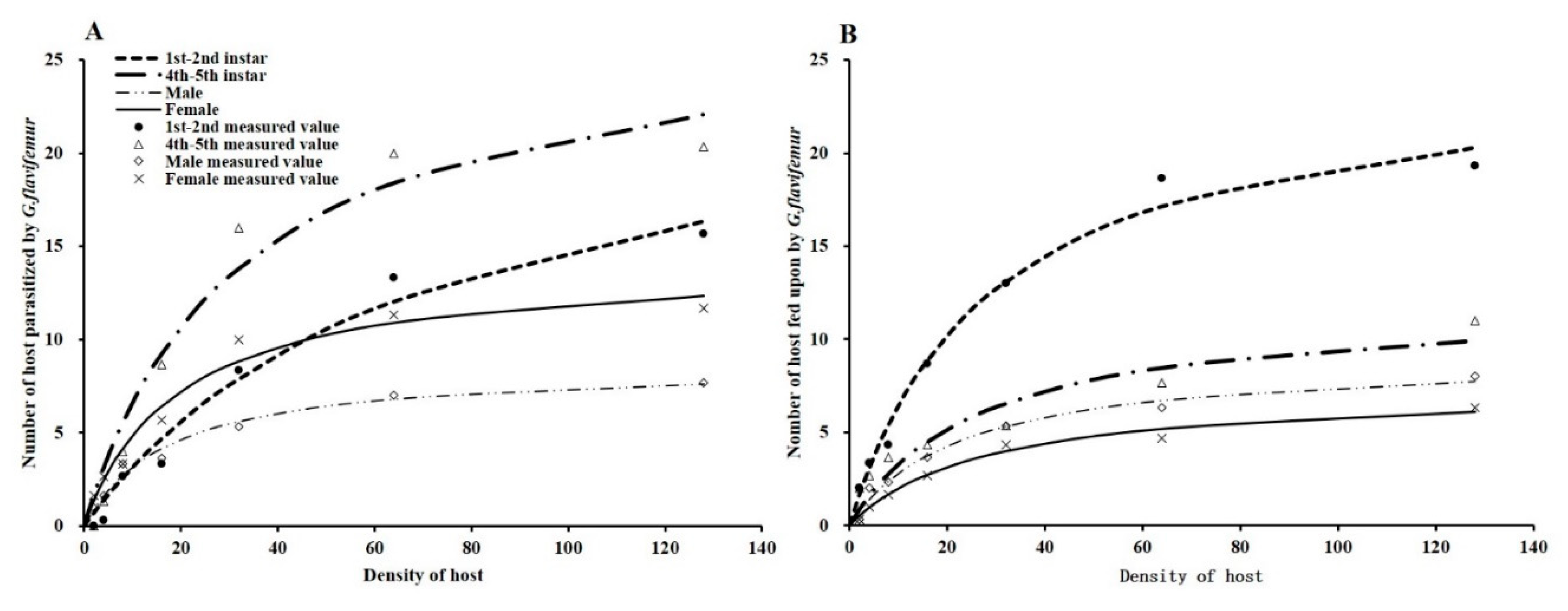
3.3. Parasitism and Host-Feeding Functional Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gurr, G.M.; Liu, J.; Read, D.M.Y.; Catindig, J.L.A.; Cheng, J.A.; Lan, L.P.; Heong, K.L. Parasitoids of Asian rice planthopper (Hemiptera: Delphacidae) pests and prospects for enhancing biological control by ecological engineering. Ann. Appl. Biol. 2011, 158, 149–176. [Google Scholar] [CrossRef]
- He, J.H.; Ma, Y.; Wang, H.Q.; Shi, Y.S.; Qiu, N.Z.; Ren, S.Z.; Chen, X.X.; Chen, Z.F.; Zhen, L.Y.; Pu, T.S.; et al. Directory of Natural Enemies of Rice Insect Pests in China; Science Press: Beijing, China, 1991; pp. 60–62. [Google Scholar]
- Lou, Y.G.; Zhang, G.R.; Zhang, W.Q.; Hu, Y.; Zhang, J. Biological control of rice insect pests in China. Biol. Control 2013, 67, 8–20. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhai, Y.F.; Lin, Q.C.; Sun, Y.X.; Li, Q.; Tao, M.; Zhou, H.X.; Li, L.L.; Yu, Y. Research progress of controlling action of parasitoids on rice planthoppers. J. Environ. Entomol. 2014, 36, 1025–1032. [Google Scholar]
- Li, Y.; Zhang, Q.; Liu, Q.; Meissle, M.; Yang, Y.; Wang, Y.; Hua, H.; Chen, X.; Peng, Y.; Romeis, J. Bt rice in China—Focusing the nontarget risk assessment. Plant Biotechnol. J. 2017, 15, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.C.; Romeis, J.; Liu, K.; Zhang, F.C.; Zheng, X.S.; Xu, H.X.; Chen, G.H.; He, X.C.; Lu, Z.X. Assessing the effects of Cry1C rice and Cry2A rice to Pseudogonatopus flavifemur, a parasitoid of rice planthoppers. Sci. Rep. 2017, 7, 7838. [Google Scholar] [CrossRef] [Green Version]
- Chua, T.H.; Dyck, V.A.; Pena, N.B. Functional response and searching efficiency in Pseudogonatopus flavifemur Esaki and Hash. (Hymenoptera: Dryinidae), a parasite of rice planthoppers. Res. Popul. Ecol. 1984, 26, 74–83. [Google Scholar] [CrossRef]
- Huang, X.F. Natural enemy of brown planthopper: A preliminary observation of Gonatopus flavifemur. Chin. J. Appl. Entomol. 1982, 05, 12–15. [Google Scholar]
- Guglielmino, A. Dryinidae (Hymenoptera Chrysidoidea): An interesting group among the natural enemies of the Auchenorrhyncha (Hemiptera). Denisia 04 2002, 176, 549–556. [Google Scholar]
- Espinosa, M.S.; Van Nieuwenhove, G.A.; Virla, E.G. Host makes the difference: The effect of two planthopper host species on host-feeding and parasitism activities of the pincer wasp Gonatopus bonaerensis. Biocontrol 2019, 64, 655–664. [Google Scholar] [CrossRef]
- Yang, S.L.; Huang, J.X.; Jin, M.X. Study on Planthoppers, rice leafhopper predators-Dryinidae. Nat. Enemies Insects 1982, 4, 1–12. [Google Scholar]
- Chen, Y.X.; Yang, K.S. Preliminary observe of biological characteristics of Haplogonatopus japonicus. Chin. J. Appl. Entomol. 1987, 4, 237–239. [Google Scholar]
- He, J.H.; Xu, Z.F. Dyrinidae, Hymenoptera, Insecta, Fauna of China; Science Press: Beijing, China, 2002; Volume 29, pp. 360–364. [Google Scholar]
- Esaki, T.; Hashimoto, S. Report on Leafhoppers Injurious to the Rice Plant and Their Natural Enemies (3); Entomological Laboratory, Department of Agriculture, Kyushu Imperial University: Fukuoka, Japan, 1932; pp. 1–42. [Google Scholar]
- Xu, Z.; Olmi, M.; He, J. Dryinidae of the Oriental region (Hymenoptera: Chrysidoidea). Zootaxa 2013, 3614, 1–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmi, M.; Xu, Z. Dryinidae of the Eastern Palaearctic region (Hymenoptera: Chrysidoidea). Zootaxa 2015, 3996, 1–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, T.H.; Dyck, V.A. Assessment of Psudogonatopus flavifemur E. & H. (Dryinidae: Hymenoptera) as a biocontrol agent against the rice brown planthopper. In Proceedings of the International Conference on Plant Protection in the Tropics, Kuala Lumpur, Malaysia, 1–4 March 1982; pp. 253–265. [Google Scholar]
- Heong, K.L.; Hardy, B. Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Int. Rice Res. Inst.: Los Bañon, Philippines, 2009; pp. 157–243. [Google Scholar]
- Wu, S.F.; Zeng, B.; Zheng, C.; Mu, X.C.; Zhang, Y.; Hu, J.; Zhang, S.; Gao, C.F.; Shen, J.L. The evolution of insecticide resistance in the brown planthopper (Nilaparvata lugens Stal) of China in the period 2012–2016. Sci. Rep. 2018, 8, 4586. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-Induced Planthopper Population Resurgence in Rice Cropping Systems. Annu. Rev. Entomol. 2020, 65, 409–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.N.; Wu, J.C.; Ma, F. Brown Planthopper: Occurrence and Control; China Agricultural Press: Beijing, China, 2003; pp. 1–37. [Google Scholar]
- Sahragard, A.; Jervis, M.A.; Kidd, N.A.C. Influence of host availability on rates of oviposition and host-feeding, and on longevity inDicondylus indianusOlmi (Hym., Dryinidae), a parasitoid of the Rice Brown Planthopper, Nilaparvata lugensStål (Hem., Delphacidae). J. Appl. Entomol. 1991, 112, 153–162. [Google Scholar] [CrossRef]
- He, J.C.; Sun, Y.Q.; Ling, J.J.; Li, B.; Fu, Q. Parasitic and Prey Selectivity of the parasitoid Gonatopus flavifemur (Esaki et Hashimoto) to Two Species of Rice Planthoppers. In Proceedings of the 2015 Annual meeting of the Chinese Society for plant protection, Jilin, China, 9–12 September 2015; p. 632. [Google Scholar]
- Ding, J.H.; Hu, C.L.; Fu, Q.; He, J.C.; Xie, M.C. A Colour Atlas of Commonly Encountered Delphacids in China Rice Regions; Zhejiang Science and Technology Press: Hangzhou, China, 2012; pp. 1–108. [Google Scholar]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect. Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef]
- Holling, C.S. Some Characteristics of Simple Types of Predation and Parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Atlıhan, R.; Kaydan, M.B.; Yarımbatman, A.; Okut, H. Functional response of the coccinellid predator Adalia fasciatopunctata revelierei to walnut aphid (Callaphis juglandis). Phytoparasitica 2010, 38, 23–29. [Google Scholar] [CrossRef]
- Strand, M.R. Developmental traits and life-history evolution in parasitoids. In Parasitoid Population Biology; Hochberg, M.E., Ives, A.R., Eds.; Princeton University Press: New Jersey, USA, 2000; pp. 139–162. [Google Scholar]
- Li, Y.X.; Liu, S.S. Hosts Defense to Oviposition Behavior of Parasitoids. Chin. J. Biol. Control 1999, 15, 35–39. [Google Scholar]
- Henry, L.M.; Ma, B.O.; Roitberg, B.D. Size-mediated adaptive foraging: A host-selection strategy for insect parasitoids. Oecologia 2009, 161, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Heinz, K.M.; Parrella, M.P. Attack behavior and host size selection by Diglyphus begini on Liriomyza trifolii in chrysanthemum. Entomol. Exp. Appl. 1989, 53, 147–156. [Google Scholar] [CrossRef]
- Charnov, E.L.; Skinner, S.W. Complementary Approaches to the Understanding of Parasitoid Oviposition Decisions. Environ. Entomol. 1985, 14, 383–391. [Google Scholar] [CrossRef]
- Kidd, N.A.C.; Jervis, M.A. Host-feeding and oviposition strategies of parasitoids in relation to host stage. Res. Popul. Ecol. 1991, 33, 13–28. [Google Scholar] [CrossRef]
- Harvey, J.A.; Strand, M.R. The developmental strategies of endoparasitoid wasps vary with host feeding ecology. Ecology 2002, 83, 2439–2451. [Google Scholar] [CrossRef]
- Liu, Y.H. Host Selection and Development Strategies of Parasitoid Meteoruspulchricornis. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 21 June 2007. [Google Scholar]
- Heimpel, G.E.; Collier, T.R. The Evolution of Host-Feeding Behaviour in Insect Parasitoids. Biol. Rev. 1996, 71, 373–400. [Google Scholar] [CrossRef]
- Rosenheim, J.A.; Rosen, D. Influence of egg load and host size on host-feeding behaviour of the parasitoid Aphytis lingnanensis. Ecol. Entomol. 1992, 17, 263–272. [Google Scholar] [CrossRef]
- Shi, S.S.; Zang, L.S.; Liu, T.X.; Ruan, C.C.; Sun, G. Host-feeding behaviors of parasitoids on hosts and implications for biological control. Acta Entomol. Sin. 2009, 52, 424–433. [Google Scholar]
- Jones, D.B.; Giles, K.L.; Berberet, R.C.; Royer, T.A.; Elliott, N.C.; Payton, M.E. Functional responses of an introduced parasitoid and an indigenous parasitoid on greenbug at four temperatures. Environ. Entomol. 2003, 32, 425–432. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Jin, L.F. Preliminary study on bionomics of Haplogonatopus japonicus. Nat. Enemies Insects 1992, 14, 57–61. [Google Scholar]
- Li, S.; Chen, W.L.; Jin, D.C. Influence of parasitism at different host stages on the developmental performance of both the parasitoid Haplogonatopusapicalis (Hymenoptera: Dyrinidae) and its host Sogatellafurcifera (Hemiptera: Delphacidae). Acta Entomol. Sin. 2015, 58, 1237–1244. [Google Scholar]
- Vinson, S.B.; Iwantsch, G.F. Host Suitability for Insect Parasitoids. Annu. Rev. Entomol. 1980, 25, 397–419. [Google Scholar] [CrossRef]
- Henry, L.M.; Roitberg, B.D.; Gillespie, D.R. Covariance of phenotypically plastic traits induces an adaptive shift in host selection behaviour. Proc. Biol. Sci. 2006, 273, 2893–2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Meng, L.; Li, B.P. Parasitizing, developmental and reproductive performances of Meteoruspidchricornis (Hymenoptera: Braconidae) in relation to host age at oviposition. Acta Entomol. Sin. 2014, 57, 1032–1036. [Google Scholar]
- Sequeira, R.; Mackauer, M. Covariance of adult size and development time in the parasitoid waspAphidius ervi in relation to the size of its host, Acyrthosiphon pisum. Evol. Ecol. 1992, 6, 34–44. [Google Scholar] [CrossRef]
- Omar, M.Y.; Olmi, M. Anteon Yasumatsui (Dryinidae), a New Nymphal and Adult Parasitoid of Nephotettixspp. (Cicadellidae) in Malaysia; National Seminary on Entomology: Kuala Lumpur, Malaysia, 1996; pp. 18–19. [Google Scholar]
- Guo, M.F. Study of parasitic behavior of Trichogramma (ІІ) affect of mating behavior on sex ratio of progeny. Nat. Enemies Insects 1992, 14, 51–53. [Google Scholar]
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press: Princeton, NJ, USA, 1982; p. 18. [Google Scholar]
- Chong, J.H.; Oetting, R.D. Functional response and progeny production of the Madeira mealybug parasitoid, Anagyrus sp nov nr. sinope: The effect of host stage preference. Biol. Control 2007, 41, 78–85. [Google Scholar] [CrossRef]
- Yamada, Y.Y.; Miyamoto, K. Payoff from self and conspecific superparasitism in a dryinid parasitoid, Haplogonatopus atratus. Oikos 1998, 81, 209–216. [Google Scholar] [CrossRef]
- Yamada, Y.Y.; Ikawa, K. Superparasitism strategy in a semisolitary parasitoid with imperfect self/non-self recognition, Echthrodelphax fairchildii. Entomol. Exp. Appl. 2005, 114, 143–152. [Google Scholar] [CrossRef]
- Mita, T.; Sanada-Morimura, S.; Matsumura, M.; Matsumoto, Y. Genetic variation of two apterous wasps Haplogonatopus apicalis and H. oratorius (Hymenoptera: Dryinidae) in East Asia. Appl. Entomol. Zool. 2013, 48, 119–124. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Stage When Parasitized | Host Stage Reached (%) at Time of Parasitoid Cocooning | |||||
|---|---|---|---|---|---|---|
| 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | Adult | |
| 1st instar | 0 | 0 | 100.00 | 0 | 0 | 0 |
| 2nd instar | - | 0 | 0 | 94.44 | 5.56 | 0 |
| 3rd instar | - | - | 0 | 0 | 96.58 | 3.42 |
| 4th instar | - | - | - | 0 | 80.58 | 19.42 |
| 5th instar | - | - | - | - | 0 | 100.00 |
| Behavior | Stage of Host | Functional Response Equation | R2 | Attack Rate (a) | Handling Time (Th/day) | Maximum Parasitism/Host Feeding (Namax) |
|---|---|---|---|---|---|---|
| Parasitism | 1st–2nd instar | Na = 0.357N/(1 + 0.014N) | 0.976 | 0.357 | 0.039 | 25.491 |
| 4th–5th instar | Na = 0.864N/(1 + 0.031N) | 0.959 | 0.864 | 0.036 | 27.579 | |
| Male | Na = 0.502N/(1 + 0.057N) | 0.981 | 0.500 | 0.116 | 8.636 | |
| Female | Na = 0.726N/(1 + 0.051N) | 0.970 | 0.726 | 0.070 | 14.229 | |
| Host-feeding | 1st–2nd instar | Na = 0.860N/(1 + 0.035N) | 0.985 | 0.860 | 0.040 | 24.882 |
| 4th–5th instar | Na = 0.449N/(1 + 0.037N) | 0.968 | 0.449 | 0.083 | 12.039 | |
| Male | Na = 0.397N/(1 + 0.044N) | 0.982 | 0.397 | 0.110 | 9.117 | |
| Female | Na = 0.267N/(1 + 0.036N) | 0.983 | 0.267 | 0.135 | 7.431 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; He, Y.; Lai, F.; Chen, X.; Fu, Q. Biological Traits of the Pincer Wasp Gonatopus flavifemur (Esaki & Hashimoto) Associated with Different Stages of Its Host, the Brown Planthopper, Nilaparvata lugens (Stål). Insects 2020, 11, 279. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11050279
He J, He Y, Lai F, Chen X, Fu Q. Biological Traits of the Pincer Wasp Gonatopus flavifemur (Esaki & Hashimoto) Associated with Different Stages of Its Host, the Brown Planthopper, Nilaparvata lugens (Stål). Insects. 2020; 11(5):279. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11050279
Chicago/Turabian StyleHe, Jiachun, Yuting He, Fengxiang Lai, Xiangsheng Chen, and Qiang Fu. 2020. "Biological Traits of the Pincer Wasp Gonatopus flavifemur (Esaki & Hashimoto) Associated with Different Stages of Its Host, the Brown Planthopper, Nilaparvata lugens (Stål)" Insects 11, no. 5: 279. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11050279