Evaluation of Drimys winteri (Canelo) Essential Oil as Insecticide against Acanthoscelides obtectus (Coleoptera: Bruchidae) and Aegorhinus superciliosus (Coleoptera: Curculionidae)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Essential Oil Extraction
2.2. Essential Oil Analysis
2.3. Insects
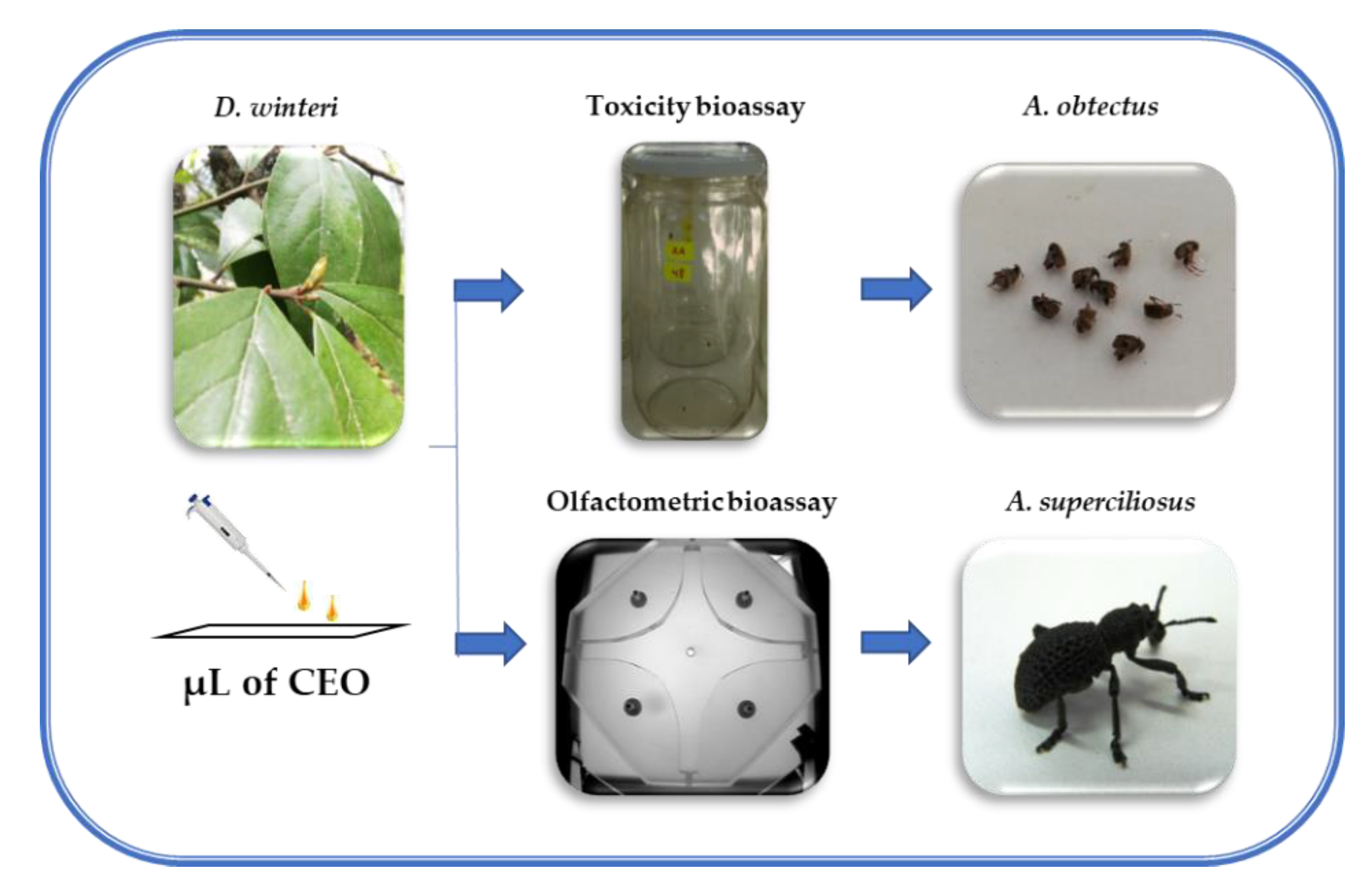
2.4. Toxicological Bioassays for Acanthoscelides obtectus
2.5. Repellency Bioassays for Aegorhinus superciliosus
2.6. Data Analysis
3. Results
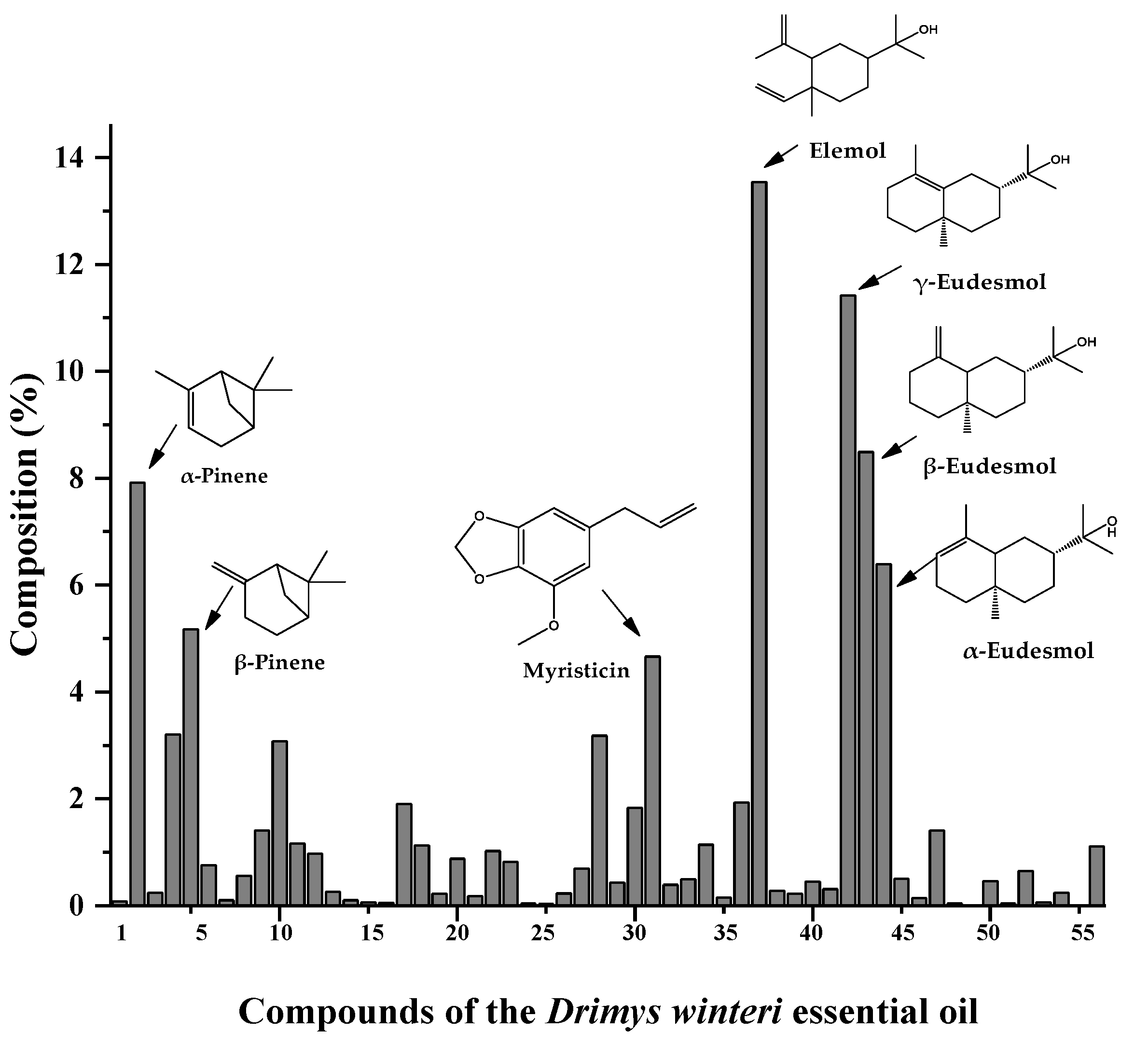
3.1. Yield and Chemical Profile of Drimys winteri Essential Oil
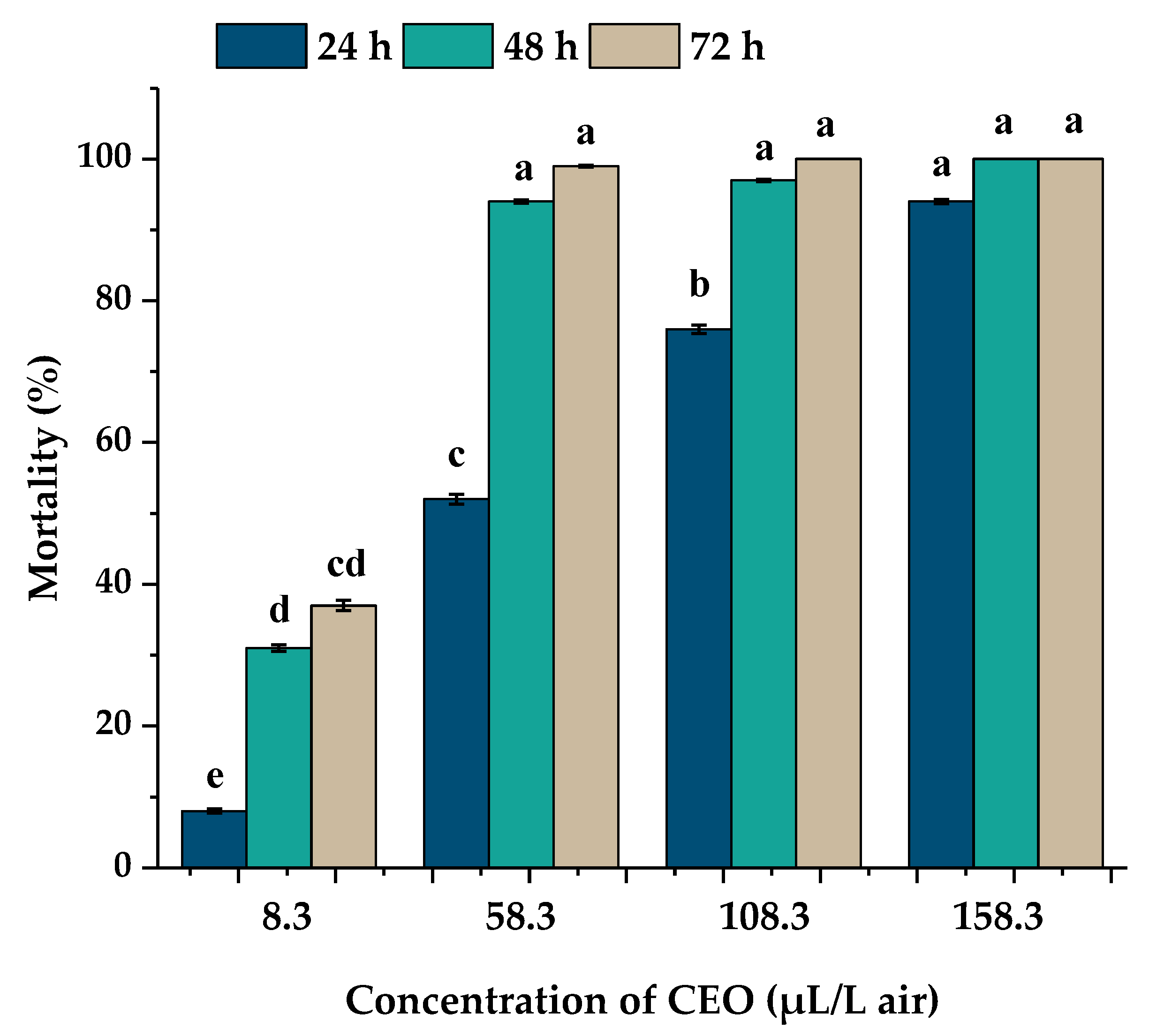
3.2. Mortality and Toxicity Bioassay for A. obtectus
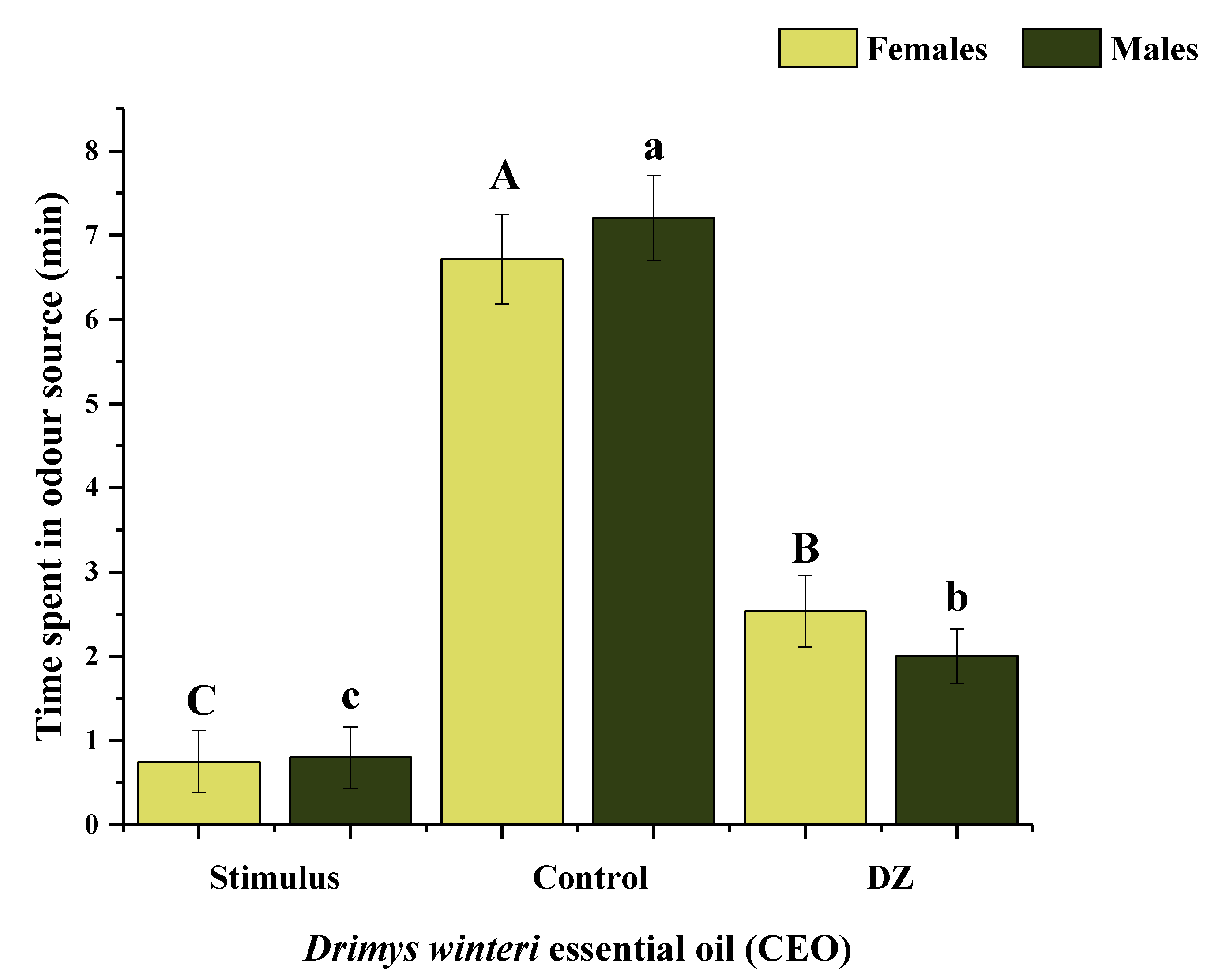
3.3. Repellency Bioassays for A. superciliosus
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Koul, O.; Walia, S.; Dhaliwal, G.S. Essential oils as green pesticides: Potential and constraints. J. Biopest 2008, 4, 63–84. [Google Scholar]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential Oils in Insect Control: Low-Risk Products in a High-Stakes World. Annu. Rev. Ѐntomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Baser, K.H.C.; Buchbauer, G. Handbook of Essential Oils: Science, Technology, and Applications, 2nd ed.; Baser, K.H.C., Buchbauer, G., Eds.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Regnault-Roger, C. The potential of botanical essential oils for insect pest control. Integr. Pest Manag. Rev. 1997, 2, 25–34. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E.E. Repellent activity of essential oils: A review. Bioresour. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Boukouvala, M.C.; Ntalli, N.; Skourti, A.; Karagianni, E.S.; Nika, E.P.; Kontodimas, D.C.; Cappellacci, L.; Petrelli, R.; Cianfaglione, K.; et al. Effectiveness of eight essential oils against two key stored-product beetles, Prostephanus truncatus (Horn) and Trogoderma granarium Everts. Food Chem. Toxicol. 2020, 139, 111255. [Google Scholar] [CrossRef]
- Bedini, S.; Guarino, S.; Echeverria, M.C.; Flamini, G.; Ascrizzi, R.; Loni, A.; Conti, B. Allium sativum, Rosmarinus officinalis, and Salvia officinalis Essential Oils: A Spiced Shield against Blowflies. Insects 2020, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Oviedo, A.; Van Nieuwenhove, G.; Rull, J. Exposure to essential oils and ethanol vapors affect fecundity and survival of two frugivorous fruit fly (Diptera: Tephritidae) pest species. Bull. Ѐntomol. Res. 2020, 1–8. [Google Scholar] [CrossRef]
- Moura, E.D.S.; Faroni, L.R.; Zanuncio, J.C.; Heleno, F.F.; Prates, L.H.F. Insecticidal activity of Vanillosmopsis arborea essential oil and of its major constituent α-bisabolol against Callosobruchus maculatus (Coleoptera: Chrysomelidae). Sci. Rep. 2019, 9, 3723. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, A.M.; Haddi, K.; Ribeiro, B.M.; Corrêia, R.F.T.; Tomé, H.V.V.; Santos-Amaya, O.; Pereira, E.J.G.; Guedes, R.N.C.; Santos, G.R.; Oliveira, E.E.; et al. Essential oil of Siparuna guianensis as an alternative tool for improved lepidopteran control and resistance management practices. Sci. Rep. 2018, 8, 7215. [Google Scholar] [CrossRef] [Green Version]
- Rajashekar, Y.; Bakthavatsalam, N.; Shivanandappa, T. Botanicals as Grain Protectants. Psyche A J. Ѐntomol. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Hoffmann, A.; Fraga, C.; Lastra, C.; Veghazi, E. Plantas Medicinales de Uso Común en Chile; Fundación Claudio Gay: Santiago, Chile, 1992; p. 78. [Google Scholar]
- Salas-Eljatib, C.; Corvalán, P.; Pino, N.; Donoso, P.J.; Soto, D.P. Mixed-effects height-diameter models for Drimys winteri in the south (41–43° S) of Chile. Bosque 2019, 40, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Larjavaara, M.; Muller-Landau, H.C. Measuring tree height: A quantitative comparison of two common field methods in a moist tropical forest. Methods Ecol. Evol. 2013, 4, 793–801. [Google Scholar] [CrossRef]
- Corvalán, P.; Araya, L.; Blanco, S.; Cox, F. El Canelo: Una Alternativa de Desarrollo Para La X Region; Fondo de Investigación Agraria y Universidad de Chile: Santiago, Chile, 1987; p. 145. [Google Scholar]
- Rodriguez, S. Antecedentes tecnológicos de Canelo (Drimys winteri Forst.). Bosque 1998, 19, 91–99. [Google Scholar] [CrossRef]
- Muñoz, O.; Montes, M.; Wilkomirsky, T. Plantas Medicinales de Uso en Chile, 1st ed.; Editorial Universitaria: Santiago, Chile, 2001. [Google Scholar]
- Wilhelm, E. Foique, Foye. Botánica indígena de Chile, 1st ed.; Editorial Andrés Bello: Santiago, Chile, 1992; pp. 78–79. [Google Scholar]
- Rodríguez, R.A.; Quezada, M. Nueva combinación en Drimys J.R. et G. Forster (Winteraceae) de Chile. Gayana Bot. 1991, 48, 111–114. [Google Scholar]
- Muñoz-Concha, D.; Vogel, H.; Razmili, I. Variación de compuestos químicos en hojas de poblaciones de Drimys spp. (Magnoliophyta: Winteraceae) en Chile. Rev. Chil. de Hist. Nat. 2004, 77, 43–50. [Google Scholar] [CrossRef]
- Sayah, M.E.; Cechinel, V.; Yunes, R.; Pinheiro, T.; Calixto, J. Action of polygodial, a sesquiterpene isolated from Drimys winteri, in the guinea-pig ileum and trachea ‘in vivo’. Eur. J. Pharmacol. 1998, 344, 215–221. [Google Scholar] [CrossRef]
- Montenegro, I.; Del Corral, S.; Napal, G.N.D.; Carpinella, M.; Mellado, M.; Villegas, A.M.; Villena, J.; Palacios, S.; Fritis, M.C. Antifeedant effect of polygodial and drimenol derivatives against Spodoptera frugiperda and Epilachna paenulataand quantitative structure-activity analysis. Pest Manag. Sci. 2018, 74, 1623–1629. [Google Scholar] [CrossRef]
- Paz, C.; Burgos, V.; Iturra, A.; Rebolledo, R.; Ortiz, L.; Baggio, R.; Becerra, J.; Cespedes-Acuña, C.L. Assessment of insecticidal responses of extracts and compounds of Drimys winteri, Lobelia tupa, Viola portalesia and Vestia foetida against the granary weevil Sitophilus granarius. Ind. Crop. Prod. 2018, 122, 232–238. [Google Scholar] [CrossRef]
- Zapata, N.; Budia, F.; Viñuela, E.; Medina, P. Antifeedant and growth inhibitory effects of extracts and drimanes of Drimys winteri stem bark against Spodoptera littoralis (Lep., Noctuidae). Ind. Crop. Prod. 2009, 30, 119–125. [Google Scholar] [CrossRef]
- Zapata, N.; Smagghe, G. Repellency and toxicity of essential oils from the leaves and bark of Laurelia sempervirens and Drimys winteri against Tribolium castaneum. Ind. Crop. Prod. 2010, 32, 405–410. [Google Scholar] [CrossRef]
- Rebolledo, R.; Abarzúa, J.; Zavala, A.; Quiroz, A.; Alvear, M.; Aguilera, A. The effects of the essential oil and hydrolate of canelo (Drimys winteri) on adults of Aegorhinus superciliosus in the laboratory. Ciencia e Investigación Agraria 2012, 39, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Paul, U.V.; Lossini, J.S.; Edwards, P.J.; Hilbeck, A. Effectiveness of products from four locally grown plants for the management of Acanthoscelides obtectus (Say) and Zabrotes subfasciatus (Boheman) (both Coleoptera: Bruchidae) in stored beans under laboratory and farm conditions in Northern Tanzania. J. Stored Prod. Res. 2009, 45, 97–107. [Google Scholar] [CrossRef]
- Thakur, D.R. Taxonomy, distribution and pest status of indian biotypes of Acanthoscelides obtectus (Coleoptera: Chrysomelidae: Bruchinae) A new record. Pak. J. Zool. 2012, 44, 189–195. [Google Scholar]
- Regnault-Roger, C.; Hamraoui, A. Inhibition of reproduction of Acanthoscelides obtectus Say (Coleoptera), a kidney bean (Phaseolus vulgaris) bruchid, by aromatic essential oils. Crop. Prot. 1994, 13, 624–628. [Google Scholar] [CrossRef]
- Schmale, I.; Wäckers, F.L.; Cardona, C.; Dorn, S. Combining parasitoids and plant resistance for the control of the bruchid Acanthoscelides obtectus in stored beans. J. Stored Prod. Res. 2003, 39, 401–411. [Google Scholar] [CrossRef]
- Silva, F.; Monteiro, A.; Del Sarto, R.; Marra, B.; Dias, S.; Figueira, E.; Oliveira, G.; Rocha, T.; Souza, D.; Da Silva, M.; et al. Proregion of Acanthoscelides obtectus cysteine proteinase: A novel peptide with enhanced selectivity toward endogenous enzymes. Peptides 2007, 28, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Baier, A.H.; Webster, B.D. Control of Acanthoscelides obtectus Say (Coleoptera: Bruchidae) in Phaseolus vulgaris L. seed stored on small farms-I. Evaluation of damage. J. Stored Prod. Res. 1992, 28, 289–293. [Google Scholar] [CrossRef]
- Mallqui, K.S.V.; Oliveira, E.E.; Guedes, R.N.C. Competition between the bean weevils Acanthoscelides obtectus and Zabrotes subfasciatus in common beans. J. Stored Prod. Res. 2013, 55, 32–35. [Google Scholar] [CrossRef]
- Parsons, D.M.J.; Credland, P.F. Determinants of oviposition in Acanthoscelides obtectus: A nonconformist bruchid. Physiol. Ѐntomol. 2003, 28, 221–231. [Google Scholar] [CrossRef]
- Mbogo, K.; Davis, J.; Myers, J. Transfer of the arcelin-phytohaemagglutinin-α amylase inhibitor seed protein locus from tepary bean (Phaseolus acutifolius A. Gray) to common bean (P. vulgaris L.). Biotechnology 2009, 8, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Galindo, J.C.; De La Fuente, M.; Ordas, B.; Domínguez, L.E.G.; Malvar, R.A. Resistance categories to Acanthoscelides obtectus (Coleoptera: Bruchidae) in tepary bean (Phaseolus acutifolius), new sources of resistance for dry bean (Phaseolus vulgaris ) breeding. Crop. Prot. 2017, 98, 255–266. [Google Scholar] [CrossRef]
- Aguilera, A.; Guerrero, J.; Rebolledo, R. Plagas y Enfermedades del Avellano Europeo en La Araucanía, Chile; Ediciones Universidad de La Frontera: Temuco, Chile, 2011; p. 126. [Google Scholar]
- Tampe, J.; Pacheco, B.; Bardehle, L.; Fuentes, E.; Salas, L.; Quiroz, A. Attractant Effect of Eucalyptus globulus (Labill) and Foeniculum vulgare (Mill.) Essential Oils on Aegorhinus superciliosus (Guérin) (Coleoptera: Curculionidae). J. Soil Sci. Plant Nutr. 2020, 20, 775–783. [Google Scholar] [CrossRef]
- Aguilera, A. Control selectivo de plagas en frutales de la zona sur. In Seminario de Protección Vegetal; Aguilera, A., Andrade, O., Díaz, J., Espinoza, N., Galdames, R., Norambuena, H., Eds.; INIA Carillanca: Temuco, Chile, 1995; pp. 141–180. [Google Scholar]
- Porca, M.; Oltean, I.; Dobrin, I. Chemical control of bean weevil, Acanthoscelides obtectus say in storage condition. J. Cent. Eur. Agric. 2003, 4, 210–216. [Google Scholar]
- Sousa, A.; Faroni, L.R.; Pimentel, M.; Guedes, R.N.C. Developmental and population growth rates of phosphine-resistant and -susceptible populations of stored-product insect pests. J. Stored Prod. Res. 2009, 45, 241–246. [Google Scholar] [CrossRef]
- Liess, M.; Foit, K.; Becker, A.; Hassold, E.; Dolciotti, I.; Kattwinkel, M.; Duquesne, S. Culmination of Low-Dose Pesticide Effects. Environ. Sci. Technol. 2013, 47, 8862–8868. [Google Scholar] [CrossRef]
- Mearns, J.; Dunn, J.; Lees-Haley, P.R. Psychological effects of organophosphate pesticides: A review and call for research by psychologists. J. Clin. Psychol. 1994, 50, 286–294. [Google Scholar] [CrossRef]
- Devine, G.J.; Furlong, M.J. Insecticide use: Contexts and ecological consequences. Agric. Hum. Values 2007, 24, 281–306. [Google Scholar] [CrossRef]
- Espinoza, J.; Urzua, A.; Tampe, J.; Parra, L.; Quiroz, A. Repellent Activity of the Essential Oil from the Heartwood of Pilgerodendron uviferum (D. Don) Florin against Aegorhinus superciliosus (Coleoptera: Curculionidae). Molecules 2016, 21, 533. [Google Scholar] [CrossRef] [Green Version]
- Tampe, J.; Parra, L.; Huaiquil, K.; Quiroz, A. Chemical composition of the essential oil of Ruta chalepensis (Linnaeus) from Chile and its potential repellent effect against Aegorhinus superciliosus (Guérin) (Coleoptera: Curculionidae). J. Soil Sci. Plant Nutr. 2016, 16, 11–24. [Google Scholar]
- Tampe, J.; Parra, L.; Huaiquil, K.; Mutis, A.; Quiroz, A. Repellent Effect and Metabolite Volatile Profile of the Essential Oil of Achillea millefolium Against Aegorhinus nodipennis (Hope) (Coleoptera: Curculionidae). Neotropical Ѐntomol. 2015, 44, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Stream, C., Ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Ayvaz, A.; Sağdiç, O.; Karaborklu, S.; Ozturk, I. Insecticidal Activity of the Essential Oils from Different Plants Against Three Stored-Product Insects. J. Insect Sci. 2010, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Parra, L.; Mutis, A.; Ceballos, R.; Lizama, M.; Pardo, F.; Perich, F.; Quiroz, A. Volatiles released from Vaccinium corymbosum were attractive to Aegorhinus superciliosus (Coleoptera: Curculionidae) in an olfactometric bioassay. Environ. Ѐntomol. 2009, 38, 781–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia, T.; Perich, F.; Pardo, F.; Palma, G.; Quiroz, A. Identification of volatiles from differently aged red clover (Trifolium pratense) root extracts and behavioural responses of clover root borer (Hylastinus obscurus) (Marsham) (Coleoptera: Scolytidae) to them. Biochem. Syst. Ecol. 2007, 35, 61–67. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: London, UK, 1971; pp. 68–78. [Google Scholar]
- Quiroz, A.; Mendez, L.; Mutis, A.; Hormazabal, E.; Ortega, F.; Birkett, M.A.; Parra, L. Antifeedant activity of red clover root isoflavonoids on Hylastinus obscurus. J. Soil Sci. Plant Nutr. 2017, 17, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Verdeguer, M.; García-Rellán, D.; Boira, H.; Pérez, E.; Gandolfo, S.; Blázquez, M.A. Herbicidal Activity of Peumus boldus and Drimys winteri Essential Oils from Chile. Molecules 2011, 16, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, O.; Christen, P.; Cretton, S.; Barrero, A.F.; Lara, A.; Herrador, M.M. Comparison of the essential oils of leaves and stem bark from two different populations of Drimys winteri a Chilean herbal medicine. Nat. Prod. Commun. 2011, 6, 879–882. [Google Scholar] [CrossRef] [Green Version]
- Becerra, J.; Bittner, M.; Hernández, V.; Brintrup, C.; Becerra, J.; Silva, M. Actividad de aceites esenciales de Canelo, Queule, Bailahuén y Culén frente a hongos fitopatógenos. BLACPMA 2010, 9, 212–215. [Google Scholar]
- Monsálvez, M.; Zapata, N.; Vargas, M.; Berti, M.; Bittner, M.; Hernandez, V. Antifungal effects of n-hexane extract and essential oil of Drimys winteri bark against Take-All disease. Ind. Crop. Prod. 2010, 31, 239–244. [Google Scholar] [CrossRef]
- Zapata, N.; Lognay, G.; Smagghe, G. Bioactivity of essential oils from leaves and bark of Laurelia sempervirens and Drimys winteri against Acyrthosiphon pisum. Pest Manag. Sci. 2010, 66, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Barrero, A.F.; Herrador, M.M.; Arteaga, P.; Lara, A.; Cortes, M. Chemical Composition of the Essential Oil from Drimys winteri Forst. Wood. J. Essent. Oil Res. 2000, 12, 685–688. [Google Scholar] [CrossRef]
- Perry, N.B.; Anderson, R.E.; Brennan, N.J.; Douglas, M.H.; Heaney, A.J.; McGrimpsey, J.A.; Smallfield, B.M. Essential oil from Dalmation sage (Salvia officinalis L.), variations among individuals, plant parts, seasons and sites. J. Agric. Food Chem. 1999, 47, 2048–2054. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.; Machial, C.M. Chapter 2 Pesticides based on plant essential oils: From traditional practice to commercialization. In Advances in Phytomedicine; Elsevier BV: Amsterdam, The Netherlands, 2006; Volume 3, pp. 29–44. [Google Scholar]
- Knaak, N.; Fiúza, L.M. Potencial dos óleos essenciais de plantas no controle de insetos e micro-organismos. Neotrop. Biol. Conserv. 2010, 5, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Papachristos, D.P.; Karamanoli, K.I.; Stamopoulos, D.C.; Menkissoglu-Spiroudi, U. The relationship between the chemical composition of three essential oils and their insecticidal activity against Acanthoscelides obtectus (Say). Pest Manag. Sci. 2004, 60, 514–520. [Google Scholar] [CrossRef]
- Chu, S.S.; Jiang, G.H.; Liu, Z.L. Insecticidal compounds from the essential oil of Chinese medicinal herb Atractylodes chinensis. Pest Manag. Sci. 2011, 67, 1253–1257. [Google Scholar] [CrossRef]
- Paluch, G.E.; Zhu, J.; Bartholomay, L.; Coats, J.R. Amyris and Siam-wood essential oils: Insect activity of sesquiterpenes. In Pesticides in Household, Structural and Residential Pest Management; University of Nebraska: Lincoln, NE, USA, 2009; pp. 5–18. [Google Scholar]
- Paluch, G.; Grodnitzky, J.; Bartholomay, L.; Coats, J. Quantitative Structure−Activity Relationship of Botanical Sesquiterpenes: Spatial and Contact Repellency to the Yellow Fever Mosquito, Aedes aegypti. J. Agric. Food Chem. 2009, 57, 7618–7625. [Google Scholar] [CrossRef] [Green Version]
- Baccari, W.; Znati, M.; Zardi-Bergaoui, A.; Chaieb, I.; Flamini, G.; Ascrizzi, R.; Ben Jannet, H. Composition and insecticide potential against Tribolium castaneum of the fractionated essential oil from the flowers of the Tunisian endemic plant Ferula tunetana Pomel ex Batt. Ind. Crop. Prod. 2020, 143, 111888. [Google Scholar] [CrossRef]
- De Souza, M.T.; De Souza, M.T.; Bernardi, O.; Krinski, D.; De Melo, D.J.; Oliveira, D.D.C.; Rakes, M.; Zarbin, P.H.G.; Maia, B.H.L.D.N.S.; Zawadneak, M.A.C. Chemical composition of essential oils of selected species of Piper and their insecticidal activity against Drosophila suzukii and Trichopria anastrephae. Environ. Sci. Pollut. Res. 2020, 27, 13056–13065. [Google Scholar] [CrossRef]
- Tanoh, E.; Boué, G.; Nea, F.; Genva, M.; Wognin, E.; LeDoux, A.; Martin, H.; Tonzibo, Z.; Frederich, M.; Fauconnier, M.-L. Seasonal Effect on the Chemical Composition, Insecticidal Properties and Other Biological Activities of Zanthoxylum leprieurii Guill. & Perr. Essential oils. Foods 2020, 9, 550. [Google Scholar] [CrossRef]
- Ketoh, G.K.; Koumaglo, H.K.; Glitho, I.A. Inhibition of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) development with essential oil extracted from Cymbopogon schoenanthus L. Spreng. (Poaceae), and the wasp Dinarmus basalis (Rondani) (Hymenoptera: Pteromalidae). J. Stored Prod. Res. 2005, 41, 363–371. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Rakotosaona, R.; Randrianarivo, E.; Nicoletti, M.; Maggi, F. Chemical composition and insecticidal activity of the essential oil from Helichrysum faradifani endemic to Madagascar. Nat. Prod. Res. 2017, 32, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Marston, A.; Hostettmann, K.; Msonthi, J.D. solation of antifungal and larvicidal constituents of Diplolophium buchanani by centrifugal partition chromatography. J. Nat. Prod. 1995, 58, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Gupta, M.; Prajapati, V.; Tripathi, A.; Kumar, S. Insecticidal Activity of Myristicin from Piper mullesua. Pharm. Boil. 2001, 39, 226–229. [Google Scholar] [CrossRef]
- Rahman, N.A.A.; Fazilah, A.; Effarizah, M.E. Toxicity of Nutmeg (Myristicin): A Review. Int. J. Adv. Sci. Eng. Inf. Technol. 2015, 5, 212. [Google Scholar] [CrossRef] [Green Version]
- Belzile, A.-S.; Majerus, S.L.; Podeszfinski, C.; Guillet, G.; Durst, T.; Arnason, J. Dillapiol Derivatives as Synergists: Structure–Activity Relationship Analysis. Pestic. Biochem. Physiol. 2000, 66, 33–40. [Google Scholar] [CrossRef]
- Kamdem, D.; Gage, D. Chemical Composition of Essential Oil from the Root Bark of Sassafras albidum. Planta Med. 1995, 61, 574–575. [Google Scholar] [CrossRef]
- Çetin, H.; Uysal, M.; Sahbaz, A.; Alaoglu, O.; Akgul, A.; Ozcan, M. Fumigant effects of essential medical and aromatic plant oils to bean weevil [Acanthoscelides obtectus Say (Coleoptera: Chrysomelidae)] adults. Selcuk J. Agric. Food Sci. 2014, 1, 6–11. [Google Scholar]
- Gokturk, T.; Kordali, S.; Ak, K.; Kesdek, M.; Bozhuyuk, A.U. Insecticidal effects of some essential oils against Tribolium confusum (du Val.) and Acanthoscelides obtectus (Say), (Coleoptera: Tenebrionidae and Bruchidae) adults. Int. J. Trop. Insect Sci. 2020. [Google Scholar] [CrossRef]
- Jumbo, L.O.V.; Faroni, L.R.; Oliveira, E.E.; Pimentel, M.A.; Silva, G.N. Potential use of clove and cinnamon essential oils to control the bean weevil, Acanthoscelides obtectus Say, in small storage units. Ind. Crop. Prod. 2014, 56, 27–34. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Hamraoui, A. Fumigant toxic activity and reproductive inhibition induced by monoterpenes on Acanthoscelides obtectus (Say) (coleoptera), a bruchid of kidney bean (Phaseolus vulgaris L.). J. Stored Prod. Res. 1995, 31, 291–299. [Google Scholar] [CrossRef]
- Ryan, M.F.; Byrne, O. Plant-insect coevolution and inhibition of acetylcholinesterase. J. Chem. Ecol. 1988, 14, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Ghabbari, M.; Guarino, S.; Caleca, V.; Saiano, F.; Sinacori, M.; Baser, N.; Ben Jemâa, J.M.; Verde, G.L. Behavior-modifying and insecticidal effects of plant extracts on adults of Ceratitis capitata (Wiedemann) (Diptera Tephritidae). J. Pest Sci. 2018, 91, 907–917. [Google Scholar] [CrossRef]
- Mutis, A.; Parra, L.; Manosalva, L.; Palma, R.; Candia, O.; Lizama, M.; Pardo, F.; Perich, F.; Quiroz, A. Electroantennographic and Behavioral Responses of Adults of Raspberry Weevil Aegorhinus superciliosus (Coleoptera: Curculionidae) to Odors Released From Conspecific Females. Environ. Ѐntomol. 2010, 39, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT | RI | Compound | % | Identification |
|---|---|---|---|---|---|
| 1 | 8.17 | 922 | α-thujene | 0.08 | RI, MS |
| 2 | 8.36 | 929 | α-pinene | 7.92 | RI, MS |
| 3 | 8.66 | 940 | camphene | 0.24 | RI, MS |
| 4 | 9.37 | 965 | sabinene | 3.20 | RI, MS |
| 5 | 9.45 | 968 | β-pinene | 5.17 | RI, MS |
| 6 | 9.91 | 983 | β-myrcene | 0.76 | RI, MS |
| 7 | 10.24 | 993 | α-phellandrene | 0.10 | RI, MS |
| 8 | 10.60 | 1005 | α-terpinene | 0.56 | RI, MS |
| 9 | 10.89 | 1016 | eucalyptol | 1.41 | RI, MS |
| 10 | 10.98 | 1019 | limonene | 3.08 | RI, MS |
| 11 | 11.25 | 1029 | β-trans-ocimene | 1.16 | RI, MS |
| 12 | 11.83 | 1049 | γ-terpinene | 0.97 | RI, MS |
| 13 | 12.71 | 1078 | terpinolene | 0.26 | RI, MS |
| 14 | 13.70 | 1111 | 3-octyl acetate | 0.10 | RI, MS |
| 15 | 14.01 | 1122 | allo-ocimene | 0.06 | RI, MS |
| 16 | 14.78 | 1150 | borneol | 0.05 | RI, MS |
| 17 | 15.15 | 1162 | (-)-terpinen-4-ol | 1.90 | RI, MS |
| 18 | 15.51 | 1174 | α-terpineol | 1.13 | RI, MS |
| 19 | 18.10 | 1267 | safrol | 0.22 | RI, MS |
| 20 | 20.16 | 1344 | α-cubebene | 0.88 | RI, MS |
| 21 | 20.82 | 1370 | copaene | 0.18 | RI, MS |
| 22 | 21.18 | 1383 | β-elemene | 1.02 | RI, MS |
| 23 | 21.85 | 1409 | caryophyllene | 0.82 | RI, MS |
| 24 | 22.10 | 1420 | β-cubebene | 0.04 | RI, MS |
| 25 | 22.36 | 1431 | aromadendrene | 0.03 | RI, MS |
| 26 | 22.66 | 1443 | α-caryophyllene | 0.23 | RI, MS |
| 27 | 23.16 | 1463 | (+)-epi-bicyclosesquiphellandrene | 0.69 | RI, MS |
| 28 | 23.32 | 1470 | germacrene d | 3.18 | RI, MS |
| 29 | 23.54 | 1478 | γ-muurolene | 0.43 | RI, MS |
| 30 | 23.69 | 1484 | γ-elemene | 1.83 | RI, MS |
| 31 | 23.83 | 1490 | myristicin | 4.66 | RI, MS |
| 32 | 24.08 | 1500 | γ-cadinene | 0.39 | RI, MS |
| 33 | 24.17 | 1504 | (-)-calamenene | 0.49 | RI, MS |
| 34 | 24.30 | 1509 | (-)-β-cadinene | 1.14 | RI, MS |
| 35 | 24.52 | 1519 | (+)-δ-cadinene | 0.15 | RI, MS |
| 36 | 24.75 | 1529 | hedycaryol | 1.93 | RI, MS |
| 37 | 24.90 | 1536 | elemol | 13.54 | RI, MS |
| 38 | 25.19 | 1549 | e-nerolidol | 0.28 | RI, MS |
| 39 | 25.44 | 1559 | spathulenol | 0.22 | RI, MS |
| 40 | 25.67 | 1569 | globulol | 0.45 | RI, MS |
| 41 | 25.83 | 1576 | ledol | 0.31 | RI, MS |
| 42 | 26.77 | 1617 | γ-eudesmol | 11.42 | RI, MS |
| 43 | 27.10 | 1632 | β-eudesmol | 8.49 | RI, MS |
| 44 | 27.23 | 1638 | α-eudesmol | 6.39 | RI, MS |
| 45 | 27.45 | 1648 | bulnesol | 0.50 | RI, MS |
| 46 | 28.00 | 1673 | eudesm-7(11)-en-4-ol | 0.14 | RI, MS |
| 47 | 29.42 | 1740 | drimenol | 1.41 | RI, MS |
| 48 | 32.56 | 1893 | sclaren | 0.04 | MS |
| 49 | 32.72 | 1901 | drimenin | 0.01 | MS |
| 50 | 32.99 | 1916 | rimuen | 0.46 | RI, MS |
| 51 | 34.16 | 1977 | ethyl palmitate | 0.04 | RI, MS |
| 52 | 34.94 | 2019 | kaur-16-ene | 0.65 | RI, MS |
| 53 | 37.08 | 2138 | ethyl linoleate | 0.06 | RI, MS |
| 54 | 37.23 | 2147 | ethyl oleate | 0.24 | RI, MS |
| 55 | 37.78 | 2179 | ethyl stearate | 0.01 | RI, MS |
| 56 | 44.51 | 2601 | hexacosane | 1.11 | RI, MS |
| - | - | - | monoterpenes | 28.08 | - |
| - | - | - | sesquiterpenes | 56.60 | - |
| - | - | - | diterpenes | 1.15 | - |
| - | - | - | phenylpropanoids | 4.88 | - |
| - | - | - | others | 1.57 | - |
| Time (h) | LC50 (95% FCI) | LC90 (95% FCI) | χ2 | p | Slope (±SE) |
|---|---|---|---|---|---|
| 24 | 60.1 (49.87–70.49) | 163.05 (139.20–196.09) | 9.04 | <0.001 | −3.85 ± 0.42 |
| 48 | 14.8 (11.24–19.05) | 40.18 (31.14–53.42) | 8.09 | <0.001 | −1.63 ± 0.20 |
| 72 | 11.15 (8.91–13.8) | 30.26 (24.04–39.75) | 7.15 | <0.001 | −1.31 ± 0.18 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tampe, J.; Espinoza, J.; Chacón-Fuentes, M.; Quiroz, A.; Rubilar, M. Evaluation of Drimys winteri (Canelo) Essential Oil as Insecticide against Acanthoscelides obtectus (Coleoptera: Bruchidae) and Aegorhinus superciliosus (Coleoptera: Curculionidae). Insects 2020, 11, 335. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060335
Tampe J, Espinoza J, Chacón-Fuentes M, Quiroz A, Rubilar M. Evaluation of Drimys winteri (Canelo) Essential Oil as Insecticide against Acanthoscelides obtectus (Coleoptera: Bruchidae) and Aegorhinus superciliosus (Coleoptera: Curculionidae). Insects. 2020; 11(6):335. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060335
Chicago/Turabian StyleTampe, Jocelyne, Javier Espinoza, Manuel Chacón-Fuentes, Andrés Quiroz, and Mónica Rubilar. 2020. "Evaluation of Drimys winteri (Canelo) Essential Oil as Insecticide against Acanthoscelides obtectus (Coleoptera: Bruchidae) and Aegorhinus superciliosus (Coleoptera: Curculionidae)" Insects 11, no. 6: 335. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060335