Comparative Cutaneous Water Loss and Desiccation Tolerance of Four Solenopsis spp. (Hymenoptera: Formicidae) in the Southeastern United States



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species and Handling
2.2. Total Body Water Content
2.3. Cuticular Permeability
2.4. Rates of Mass Loss, Water Loss, and Mortality
2.5. Statistical Analysis
3. Results
3.1. Body Mass and Water Content
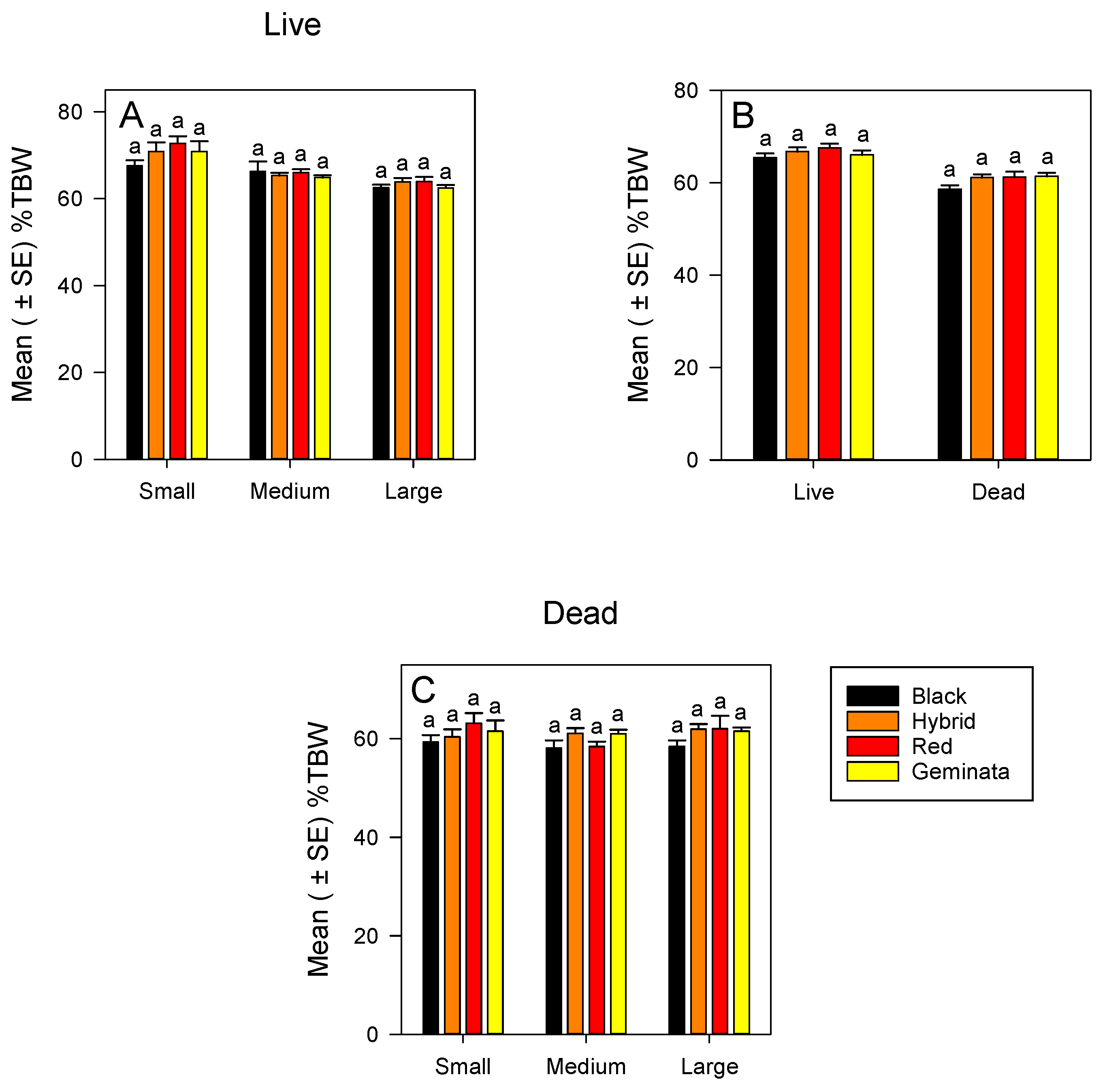
3.1.1. %TBW of Live Ants
3.1.2. %TBW of Dead Ants
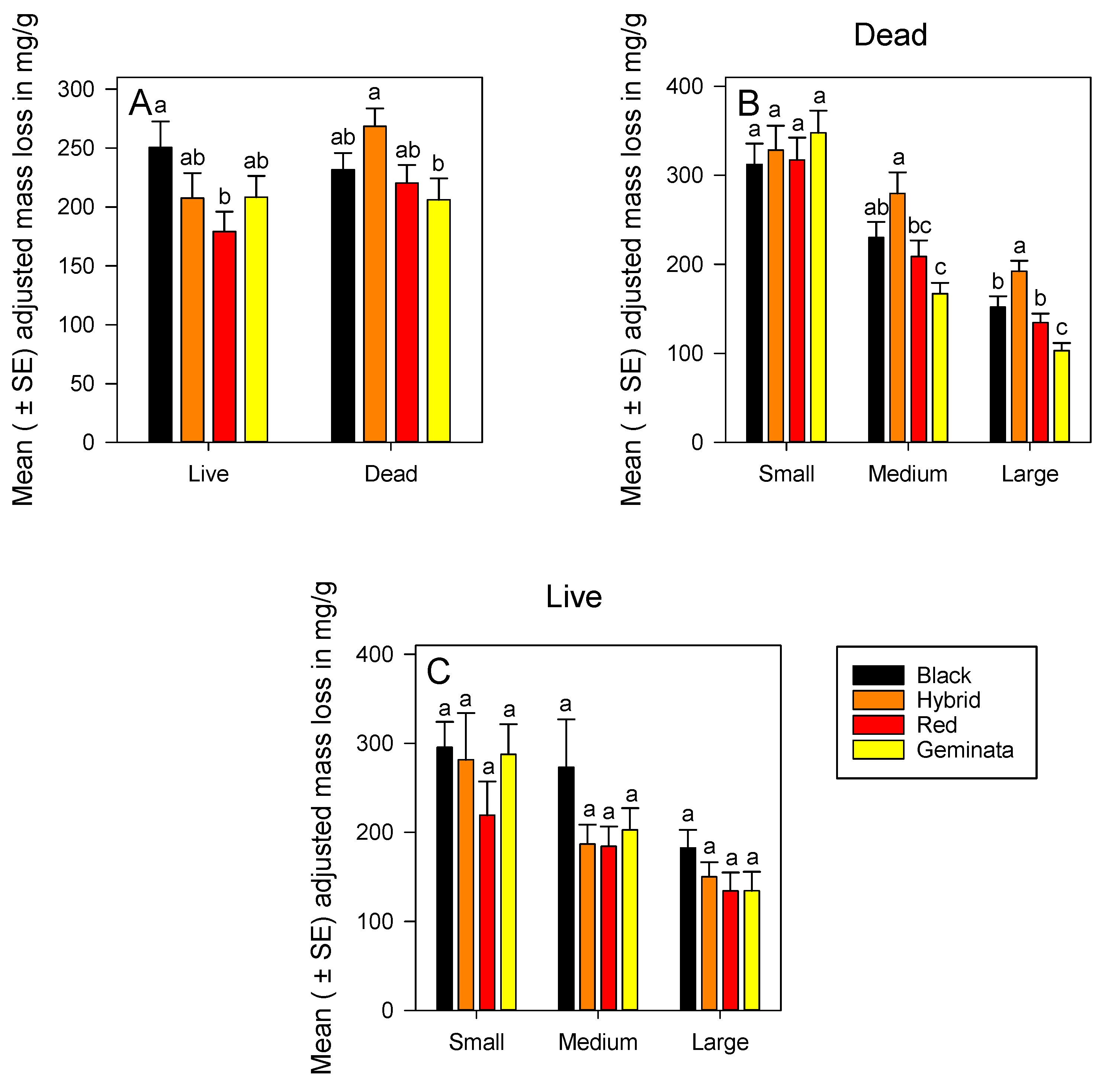
3.1.3. Adjusted Mass Loss of Live Ants
3.1.4. Adjusted Mass Loss of Dead Ants
3.1.5. Rates of % Initial Mass Loss and %TBW Lost
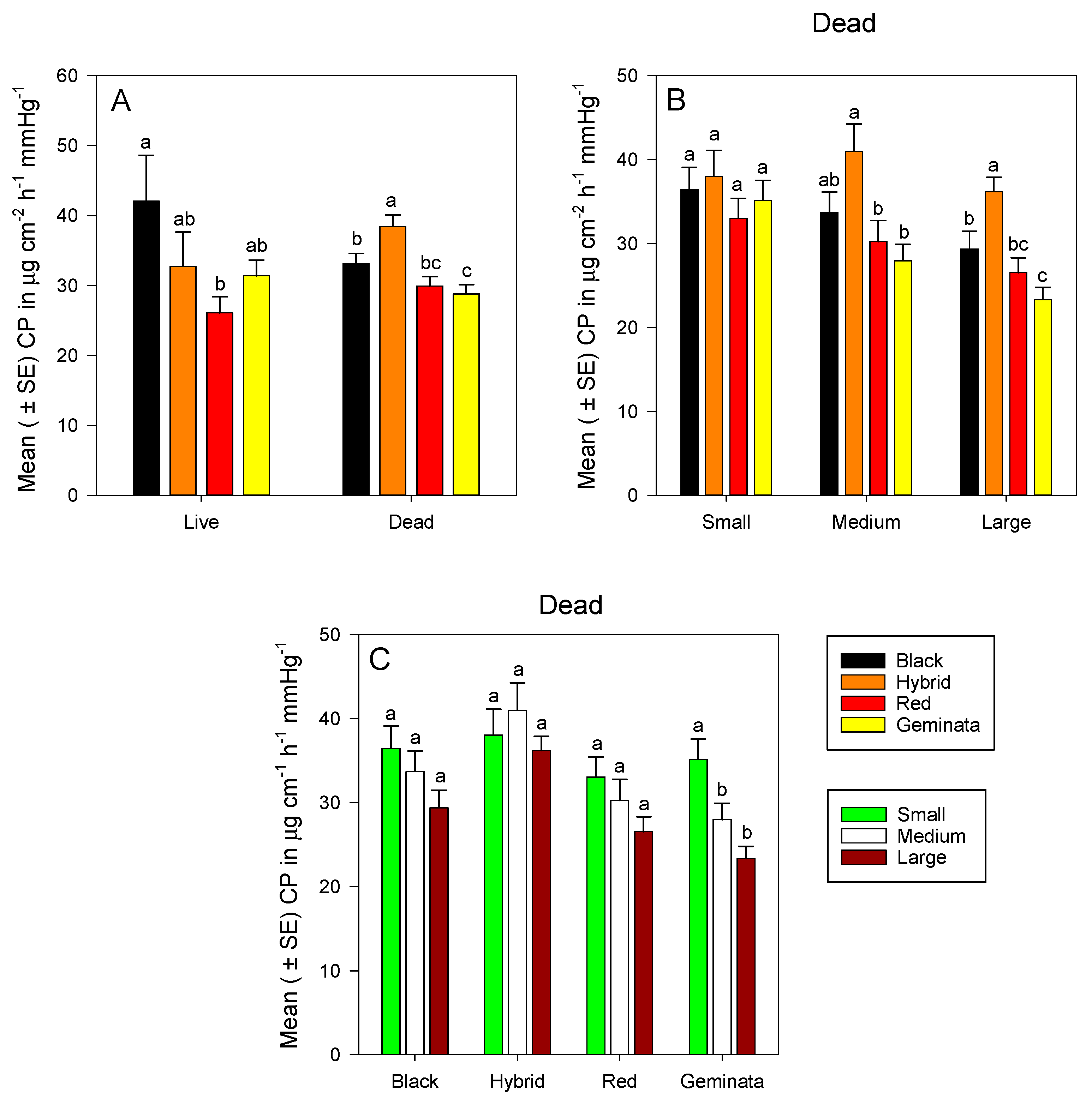
3.2. Cuticular Permeability
3.3. Desiccation Tolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rogers, D.J.; Robinson, T.P. Tsetse Distribution. In The Trypanosomiases; Maudlin, I., Holmes, P.H., Miles, M.A., Eds.; CABI Publishing: Wallingford, England, 2004; pp. 139–180. [Google Scholar]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulyy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danks, H.V. Dehydration in dormant insects. J. Insect Physiol. 2000, 46, 837–852. [Google Scholar] [CrossRef]
- Chown, S.L.; Nicolson, S.W. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Kleynhans, E.; Terblanche, J.S. Complex interactions between temperature and relative humidity on water balance of adult tsetse (Glossinidae: Diptera): Implications for climate change. Front. Physiol. 2011, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Morrison, L.W.; Korzukhin, M.D.; Porter, S.D. Predicted range expansion of the invasive fire ant, Solenopsis invicta, in the eastern United States based on the VEMAP global warming scenario. Divers. Distrib. 2005, 11, 199–204. [Google Scholar] [CrossRef]
- Weldon, C.W.; Boardman, L.; Marlin, D.; Terblanche, J.S. Physiological mechanisms of dehydration tolerance contribute to the invasion potential of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) relative to its less widely distributed congeners. Front. Zool. 2016, 13, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P. The discovery, scope, and puzzle of desiccation tolerance in plants. Plant Ecol. 2000, 151, 5–17. [Google Scholar] [CrossRef]
- Alpert, P.; Oliver, M.J. Drying without dying. In Desiccation and Survival in Plants: Drying without Dying; Black, M., Prichard, H.W., Eds.; CAB International: Wallingford, UK, 2002; pp. 3–43. [Google Scholar]
- Appel, A.G.; Miller, M.K.; Mack, T.P. Cutaneous water loss of several stages of the red imported fire ant, Solenopsis invicta (Buren). Comp. Biochem. Physiol 1991, 98, 281–283. [Google Scholar] [CrossRef]
- Beament, J.W.L. The water relations of insect cuticle. Biol. Rev. 1961, 36, 281–320. [Google Scholar] [CrossRef]
- Hadley, N.F. Water Relations of Terrestrial Arthropods; Academic: San Diego, CA, USA, 1994. [Google Scholar]
- Edney, E.B. Water Balance in Land Arthropods; Springer: New York, NY, USA, 1977. [Google Scholar]
- Mazer, C.L.; Appel, A.G. Water loss and desiccation tolerances of longwing butterflies (Lepidoptera: Nymphalidae). Environ. Entomol. 2001, 30, 631–636. [Google Scholar] [CrossRef]
- Manabe, S.; Stouffer, R.J.; Spelman, M.J.; Bryan, K. Transient response of a coupled ocean-atmosphere-land surface model to increasing atmospheric carbon dioxide. J. Clim. 1991, 4, 785–818. [Google Scholar] [CrossRef]
- Fields, P.A.; Graham, J.B.; Rosenblatt, R.H.; Somero, G.N. Effects of expected global climate change on marine faunas. Trends Ecol. Evol. 1993, 8, 361–366. [Google Scholar] [CrossRef]
- Peng, G.; Leslie, L.M.; Shao, Y. Environmental Modeling and Prediction; Springer: Berlin, Germany, 2002. [Google Scholar]
- Viboud, C.; Pakdaman, K.; Boëlle, P.Y.; Wilson, M.L.; Myers, M.F.; Valleron, A.J.; Flahault, A. Association of influenza epidemics with global climate variability. Eur. J. Epidemiol. 2004, 19, 1055–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.W.; Lai, F.Y.L.; Nymadawa, P.; Deng, Y.; Ratnamohan, M.; Petric, M.; Loh, T.P.; Tee, N.W.S.; Dwyer, D.E.; Barr, I.G.; et al. Comparison of the incidence of influenza in relation to climate factors during 2000-2007 in five countries. J. Med. Virol. 2010, 82, 1958–1965. [Google Scholar] [CrossRef]
- Cunze, S.; Heydel, F.; Tackenberg, O. Are plant species able to keep pace with the rapidly changing climate? PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Septiningsih, E.M.; Kohli, A.; Thomson, M.J.; Ye, C.; Redoña, E.; Kumar, A.; Gregorio, G.B.; Wassmann, R.; Ismail, A.M.; et al. Genetic advances in adapting rice to a rapidly changing climate. J. Agron. Crop Sci. 2012, 198, 360–373. [Google Scholar] [CrossRef]
- Winston, P.W. Cuticular water pump in insects. Nature 1967, 214, 383–384. [Google Scholar] [CrossRef]
- Winston, P.W.; Beament, J.W.L. An active reduction of water level in insect cuticle. J. Exp. Biol. 1969, 50, 541–546. [Google Scholar] [PubMed]
- Berridge, M.J. Osmoregulation in terrestrial arthropods. In Chemical Zoology; Florkin, M., Scheer, B.T., Eds.; Academic Press: London, UK; New York, NY, USA, 1970; Volume 5, pp. 287–319. [Google Scholar]
- Human, K.G.; Weiss, S.; Weiss, A.; Sandler, B.; Gordon, D.M. Effects of abiotic factors on the distribution and activity of the invasive Argentine ant (Hymenoptera: Formicidae). Pop. Ecol. 1998, 27, 822–833. [Google Scholar] [CrossRef]
- Lofgren, C.S.; Banks, W.A.; Glancey, B.M. Biology and control of imported fire ants. Annu. Rev. Entomol. 1975, 20, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.S. Biological Control of Introduced Ant Species, Exotic Ants: Biology, Impact, and Control of Introduced Species; Williams, D.F., Ed.; Westview Press: Boulder, CO, USA, 1994; pp. 293–307. [Google Scholar]
- Vinson, S.B. Invasion of the red imported fire ant (Hymenoptera: Formicidae): Spread biology, and impact. Am. Entomol. 1997, 43, 23–39. [Google Scholar] [CrossRef]
- Cohen, P.; Privman, E. Speciation and hybridization in invasive fire ants. BMC Evol. Biol. 2019, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- Mirenda, J.T.; Vinson, S.B. Division of labour and specification of castes in the red imported fire ant Solenopsis invicta Buren. Anim. Behav. 1981, 29, 410–420. [Google Scholar] [CrossRef]
- Diffie, S.; Vander Meer, R.K.; Bass, M.H. Discovery of hybrid fire ant populations in Georgia and Alabama. J. Entomol. Sci. 1988, 23, 187–191. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Lofgren, C.S.; Alvarez, F.M. Biochemical evidence for hybridization in fire ants. Fla. Entomol. 1985, 68, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Taber, S.W. Fire Ants; Texas A&M University Press: College Station, TX, USA, 2000; 308p. [Google Scholar]
- Buren, W.F.; Allen, G.E.; Whitcomb, W.H.; Lennartz, F.E.; Williams, R.N. Zoogeography of the imported fire ants. J. N. Y. Entomol. Soc. 1974, 82, 113–124. [Google Scholar]
- Tschinkel, W.R. The Fire Ants; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Buren, W.F. Revisionary studies on the taxonomy of the imported fire ants. J. Ga. Entomol. Soc. 1972, 7, 1–27. [Google Scholar]
- Korzukhin, M.D.; Porter, S.D.; Thompson, L.C.; Wiley, S. Modeling temperature-dependent range limits for the fire ant, Solenopsis invicta (Hymenoptera: Formicidae) in the United States. Environ. Entomol. 2001, 30, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Morrison, L.W.; Porter, S.D.; Daniels, E.; Korzukhin, M.D. Potential global range expansion of the invasive fire ant, Solenopsis invicta. Biol. Invasions 2004, 6, 183–191. [Google Scholar] [CrossRef]
- Callcott, A.M.A.; Collins, H.L. Invasion and range expansion of imported fire ants (Hymenoptera: Formicidae) in North America from 1918-1995. Fla. Entomol. 1996, 79, 240–251. [Google Scholar] [CrossRef]
- Davis, L.R.; Vander Meer, R.K.; Porter, S.D. Red imported fire ants expand their range across the West Indies. Fla. Entomol. 2001, 84, 735–736. [Google Scholar] [CrossRef]
- Mescher, M.C.; Ross, K.G.; Shoemaker, D.D.; Keller, L.; Krieger, M.J.B. Distribution of the two social forms of the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the native South American range. Ecol. Popul. Biol. 2003, 96, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Ascunce, M.; Yang, C.C.; Oakey, J.; Calcaterra, L.; Wu, W.J.; Shih, C.J.; Goudet, J.; Ross, K.; Shoemaker, D. Global invasion history of the fire ant Solenopsis invicta. Science 2011, 331, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rashid, T.; Feng, G. A comparative study between Solenopsis invicta and Solenopsis richteri on tolerance to heat and desiccation stresses. PLoS ONE 2014, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schilman, P.E.; Lighton, J.B.; Holway, D.A. Water balance in the Argentine ant (Linepithema humile) compared with five common native ant species from Southern California. Physiol. Entomol. 2007, 32, 1–7. [Google Scholar] [CrossRef]
- Li, J.; Heinz, K.M. Genetic variation in desiccation resistance and adaptability of the red imported fire ant (Hymenoptera: Formicidae) to arid regions. Ann. Entomol. Soc. Am. 1998, 91, 726–729. [Google Scholar] [CrossRef]
- Xu, M.; Lu, Z.; Lu, Y.; Balusu, R.R.; Ajayi, O.S.; Fadamiro, H.Y.; Appel, A.G.; Chen, L. Cuticular hydrocarbon chemistry, an important factor shaping the current distribution pattern of the imported fire ants in the USA. J. Insect Physiol. 2018, 110, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Braulick, L.S.; Cokendolpher, J.C.; Morrison, W.P. Effect of acute exposure to relative humidity and temperature on four species of fire ants (Solenopsis: Formicidae: Hymenoptera). Tex. J. Sci. 1988, 40, 331–340. [Google Scholar]
- Munroe, P.D.; Thorvilson, H.G.; Phillips, S.A., Jr. Comparison of desiccation rates among three species of fire ants. Southwest. Entomol. 1996, 21, 173–179. [Google Scholar]
- Wendt, C.F.; Verble-Pearson, R. Critical thermal maxima and body size positively correlate in red imported fire ants, Solenopsis invicta. Southwest. Nat. 2016, 61, 79–83. [Google Scholar] [CrossRef]
- Phillips, S.A., Jr.; Jusino-Atresino, R.; Thorvilson, H.G. Desiccation resistance in populations of the red imported fire ant (Hymenoptera: Formicidae). Environ. Entomol. 1996, 25, 460–464. [Google Scholar] [CrossRef]
- Martin, J.B.; Vinson, S.B. The influence of travel distance on sugar loading decisions and water balance in the central place foraging ant Solenopsis invicta. Insectes Soc. 2008, 55, 129–136. [Google Scholar] [CrossRef]
- Vogt, J.T.; Streett, R.M.; Pereira, D.A.; Callcott, A.M.A. Mississippi areawide fire ant suppression program: Unique aspects of working with black and hybrid imported fire ants. J. Agric. Urban Entomol. 2003, 20, 105–111. [Google Scholar]
- Ross, K.G.; Vander Meer, R.K.; Fletcher, D.J.C.; Vargo, E.L. Biochemical phenotypic and genetic studies of two introduced fire ants and their hybrid (Hymenoptera: Formicidae). Evolution 1987, 41, 280–293. [Google Scholar] [CrossRef]
- Chen, L.; Hu, Q.-B.; Fadamiro, H.Y. Reduction of venom alkaloids in Solenopsis richteri × Solenopsis invicta hybrid: An attempt to identify new alkaloidal components. J. Agric. Food. Chem. 2010, 58, 11534–11542. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Balusu, R.R.; Zhang, W.-Q.; Ajayi, O.S.; Lu, Y.-Y.; Zeng, R.-S.; Fadamiro, H.Y.; Chen, L. Intra- and inter-specific variation in alarm pheromone produced by Solenopsis fire ants. Bull. Entomol. Res. 2018, 108, 667–673. [Google Scholar] [CrossRef]
- Wilson, E.O. Division of labor in fire ants based on physical castes. J. Kans. Entomol. Soc. 1978, 51, 615–636. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; The Belknap Press of Harvard University Press: Cambridge, CA, USA, 1990. [Google Scholar]
- Appel, A.G.; Reierson, D.A.; Rust, M.K. Comparative water relations and temperature sensitivity of cockroaches. Comp. Bioch. Physiol. 1983, 74, 357–361. [Google Scholar] [CrossRef]
- Mack, T.P.; Appel, A.G. Water relations of immature and adult lesser cornstalk borers, Elasmopalpus lignosellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1986, 79, 579–582. [Google Scholar] [CrossRef]
- Edney, E.B.; McFarlane, J.M. The effect of temperature on transpiration in the desert cockroach, Arenivaga investigata and in Periplaneta americana. Physio. Zool. 1974, 47, 1–12. [Google Scholar] [CrossRef]
- Haar, L.; Gallagher, J.S.; Kell, G.S. NBS/NRC Steam Tables; Hemisphere Publishing Corp.: New York, NY, USA, 1984. [Google Scholar]
- SAS Institute Inc. SAS/STAT 13.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Systat Software Inc. SigmaPlot, Version 13.0; Systat Software Inc.: San Jose, CA, USA, 2014. [Google Scholar]
- Robertson, J.L.; Preisler, H.; Russell, R.M. PoloPlus: Probit and Logit Analysis User’s Guide; LeOra Software: Petaluma, CA, USA, 2007. [Google Scholar]
- Elzen, G.W. Oxygen consumption and water loss in the imported fire ant Solenopsis invicta Buren. Comp. Biochem. Physiol. 1986, 84, 13–17. [Google Scholar] [CrossRef]
- Wills, B.D.; Powell, S.; Rivera, M.D.; Suarez, A.V. Correlates and consequences of worker polymorphism in ants. Annu. Rev. Entomol. 2018, 63, 575–598. [Google Scholar] [CrossRef] [PubMed]
- Roeder, K.A.; Prather, R.M.; Paraskevopoulos, A.W.; Roeder, D.V. The economics of optimal foraging by the red imported fire ant. Environ. Entomol. 2020, 49, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Lighton, J.R.B.; Feener, D.H., Jr. Water-loss rate and cuticular permeability in foragers of the desert ant Pogonomyrmex rugosus. Physiol. Zool. 1989, 62, 1232–1256. [Google Scholar] [CrossRef]
- Hood, W.G.; Tschinkel, W.R. Desiccation resistance in arboreal and terrestrial ants. Physiol. Entomol. 1990, 15, 23–35. [Google Scholar] [CrossRef]
- Porter, S.D.; Tschinkel, W.R. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): Effects of Weather and Season. Environ. Entomol. 1987, 16, 802–808. [Google Scholar] [CrossRef]
- Mankowski, M.E. Biology of the Carpenter Ants Camponotus Vicinus (Mayr) and Camponotus Modoc (Wheeler) in Western Oregon; Oregon State University: Corvallis, OR, USA, 2001. [Google Scholar]
- Fox, E.G.P.; Pianaro, A.; Solis, D.R.; Delabie, J.H.C.; Vairo, B.C.; Machado, E.A.; Bueno, O.C. Intraspecific and Intracolonial Variation in the Profile of Venom Alkaloids and Cuticular Hydrocarbons of the Fire Ant Solenopsis saevissima Smith (Hymenoptera: Formicidae). Psyche 2012, 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hung, A.C.F.; Vinson, S.B. Factors affecting the distribution of fire ants in Texas. Southwest. Nat. 1978, 23, 205–214. [Google Scholar] [CrossRef]
- Pimm, S.L.; Bartell, D.P. Statistical model for predicting range expansion of the red imported fire ant, Solenopsis invicta, in Texas. Environ. Entomol. 1980, 9, 653–658. [Google Scholar] [CrossRef]
- Moody, J.V.; Francke, O.F.; Merickel, F.W. The distribution of fire ants, Solenopsis (Solenopsis) in western Texas. J. Kans. Entomol. Soc. 1981, 54, 469–480. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Colony | Town/City, State | Latitude | Longitude |
|---|---|---|---|---|
| Solenopsis geminata | 1 | Gainesville, FL | 29°34’19.49” N | 82°27’22.89” W |
| 2 | Gainesville, FL | 29°34’24.92” N | 82°27’30.49” W | |
| 3 | Gainesville, FL | 29°34’25.23” N | 82°27’29.59” W | |
| S. invicta | 1 | Auburn, AL | 32°37’31.47” N | 85°30’07.78” W |
| 2 | Auburn, AL | 32°34’36.73” N | 85°29’52.24” W | |
| 3 | Auburn, AL | 32°36’59.58” N | 85°30’27.14” W | |
| S. richteri | 1 | Waverly, TN | 36°05’01.41” N | 87°48’31.32” W |
| 2 | Hohenwald, TN | 35°33’24.66” N | 87°31’46.43” W | |
| 3 | Mount Pleasant, TN | 35°36’38.52” N | 87°15’46.40” W | |
| S. invicta × S. richteri | 1 | Huntsville, AL | 34°32’18.66” N | 86°30’00.27” W |
| 2 | Huntsville, AL | 34°34’45.70” N | 86°33’13.75” W | |
| 3 | Decatur, AL | 34°31’47.41” N | 86°54’02.96” W |
| Species | Size Class | N for Live Ants | Initial Mass (mg) Live Ants | N for Dead Ants | Initial Mass (mg) Dead Ants |
|---|---|---|---|---|---|
| S. richteri | Small | 15 | 0.89 ± 0.03 **A | 15 | 0.72 ± 0.03 cA |
| Medium | 15 | 2.94 ± 0.03 *A | 15 | 1.47 ± 0.09 bB | |
| Large | 15 | 3.19 ± 0.12 *B | 15 | 3.31 ± 0.15 aB | |
| S. invicta × S. richteri | Small | 15 | 1.18 ± 0.42 **A | 15 | 0.70 ± 0.02 cA |
| Medium | 15 | 1.71 ± 0.16 **A | 15 | 1.49 ± 0.11 bB | |
| Large | 14 | 3.08 ± 0.27 *B | 14 | 3.21 ± 0.26 aB | |
| S. invicta | Small | 15 | 0.59 ± 0.02 ***A | 15 | 0.52 ± 0.02 cB |
| Medium | 14 | 1.61 ± 0.13 **A | 15 | 1.41 ± 0.09 bB | |
| Large | 15 | 3.23 ± 0.19 *B | 15 | 3.52 ± 0.14 aB | |
| S. geminata | Small | 15 | 0.63 ± 0.09 ***A | 15 | 0.47 ± 0.02 cB |
| Medium | 15 | 2.07 ± 0.14 **A | 15 | 2.23 ± 0.19 bA | |
| Large | 15 | 5.26 ± 0.35 *A | 15 | 5.63 ± 0.37 aA |
| Species | Size | N | LT50 (h) | (95% CI) | Slope ± SE | χ2 | Df |
|---|---|---|---|---|---|---|---|
| S. richteri | Small | 15 | 2.21 | (1.59–2.71) | 6.14 ± 1.75 | 0.09 | 4 |
| Medium | 15 | 3.03 | (2.19–3.76) | 4.07 ± 0.84 | 1.44 | 4 | |
| Large | 15 | 3.65 | (2.57–4.62) | 3.20 ± 0.67 | 2.20 | 4 | |
| S. invicta × S. richteri | Small | 15 | 2.08 | (1.36–2.58) | 5.65 ± 1.69 | 0.12 | 4 |
| Medium | 15 | 3.63 | (2.87–4.34) | 5.02 ± 0.96 | 1.77 | 4 | |
| Large | 14 | 5.38 | (4.24–6.67) | 3.50 ± 0.68 | 3.76 | 4 | |
| S. invicta | Small | 15 | 2.49 | (1.44–3.29) | 3.21 ± 0.76 | 0.60 | 4 |
| Medium | 14 | 4.50 | (3.61–5.35) | 4.81 ± 0.91 | 0.93 | 4 | |
| Large | 15 | 8.52 | (6.14–13.81) | 3.38 ± 0.67 | 4.29 | 4 | |
| S. geminata | Small | 15 | 1.45 | (0.27–2.22) | 3.06 ± 1.03 | 1.08 | 4 |
| Medium | 15 | 4.15 | (3.28–4.99) | 4.44 ± 0.85 | 2.57 | 4 | |
| Large | 15 | 7.59 | (6.11–9.79) | 3.35 ± 0.67 | 2.66 | 4 |
| Species | Size | N | %TBW Lost at LT50 | %TBW Lost at Lower CI | %TBW Lost at Upper CI | Mean %TBW Lost at LT50 for All Sizes in a Species a |
|---|---|---|---|---|---|---|
| S. richteri | Small | 15 | 48.05 | 39.49 | 53.53 | 45.42 |
| Medium | 15 | 48.69 | 39.93 | 54.77 | ||
| Large | 15 | 39.53 | 30.56 | 46.33 | ||
| S. invicta × S. richteri | Small | 15 | 42.64 | 27.88 | 48.27 | 43.31 |
| Medium | 15 | 44.57 | 38.12 | 49.76 | ||
| Large | 14 | 42.73 | 35.82 | 49.61 | ||
| S. invicta | Small | 15 | 40.97 | 28.07 | 48.39 | 45.79 |
| Medium | 14 | 46.59 | 40.29 | 51.50 | ||
| Large | 15 | 49.80 | 39.13 | 68.17 | ||
| S. geminata | Small | 15 | 35.74 | 9.35 | 46.05 | 42.47 |
| Medium | 15 | 44.91 | 38.47 | 50.24 | ||
| Large | 15 | 46.76 | 39.90 | 55.63 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajayi, O.S.; Appel, A.G.; Chen, L.; Fadamiro, H.Y. Comparative Cutaneous Water Loss and Desiccation Tolerance of Four Solenopsis spp. (Hymenoptera: Formicidae) in the Southeastern United States. Insects 2020, 11, 418. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070418
Ajayi OS, Appel AG, Chen L, Fadamiro HY. Comparative Cutaneous Water Loss and Desiccation Tolerance of Four Solenopsis spp. (Hymenoptera: Formicidae) in the Southeastern United States. Insects. 2020; 11(7):418. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070418
Chicago/Turabian StyleAjayi, Olufemi S., Arthur G. Appel, Li Chen, and Henry Y. Fadamiro. 2020. "Comparative Cutaneous Water Loss and Desiccation Tolerance of Four Solenopsis spp. (Hymenoptera: Formicidae) in the Southeastern United States" Insects 11, no. 7: 418. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070418