Oral Ingestion of Bacterially Expressed dsRNA Can Silence Genes and Cause Mortality in a Highly Invasive, Tree-Killing Pest, the Emerald Ash Borer


Abstract
:1. Introduction
2. Materials and Methods
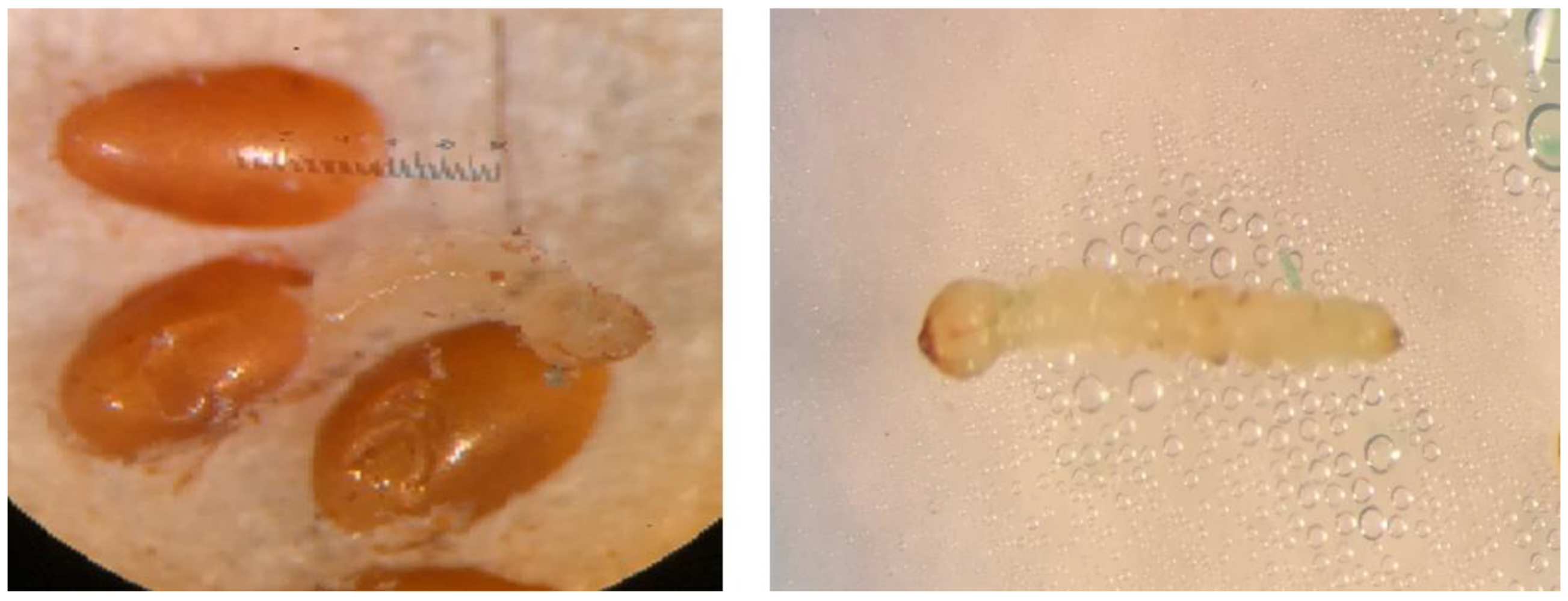
2.1. Insect Rearing
2.2. Target Gene Selection, Total RNA Extraction, cDNA Synthesis, PCR Amplification, and Construction of Recombinant L4440 Vector
2.3. Bacterial Transformation and Expression of dsRNA
2.4. Biological Activity of Recombinant Bacteria Expressing dsRNA
2.5. Molecular Validation of Gene Silencing
3. Results
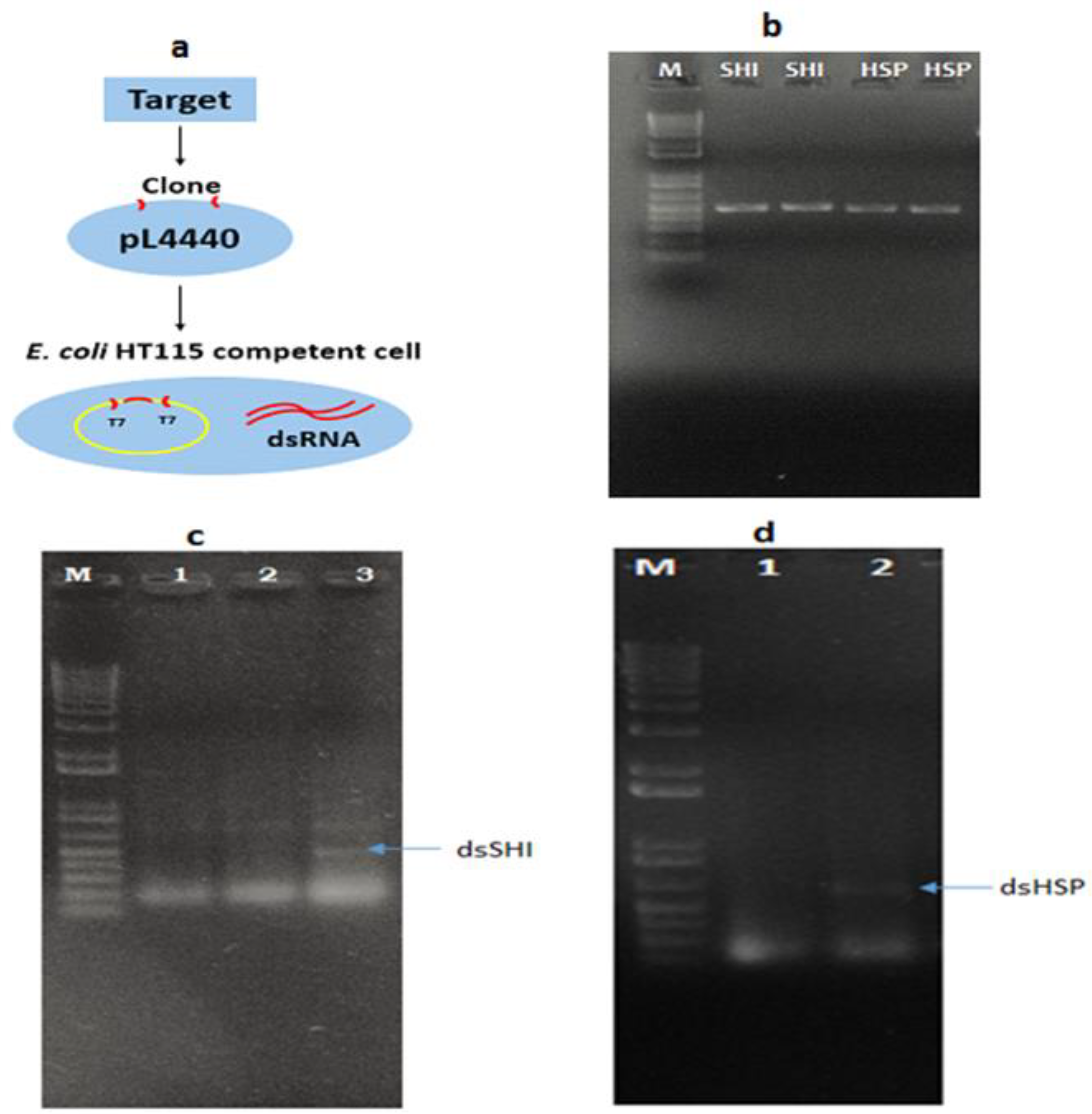
3.1. Bacterial Transformation and Expression of dsRNA
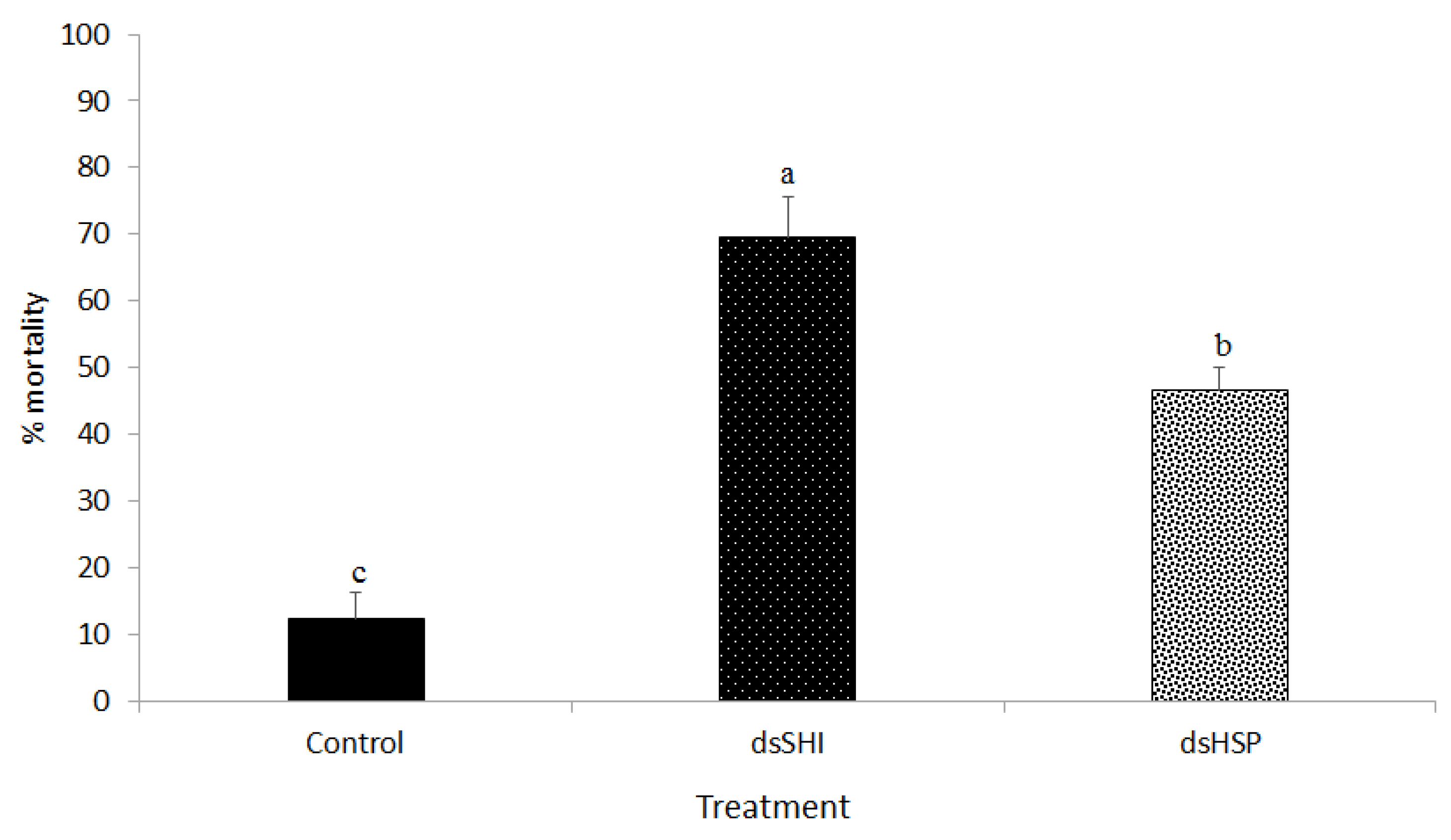
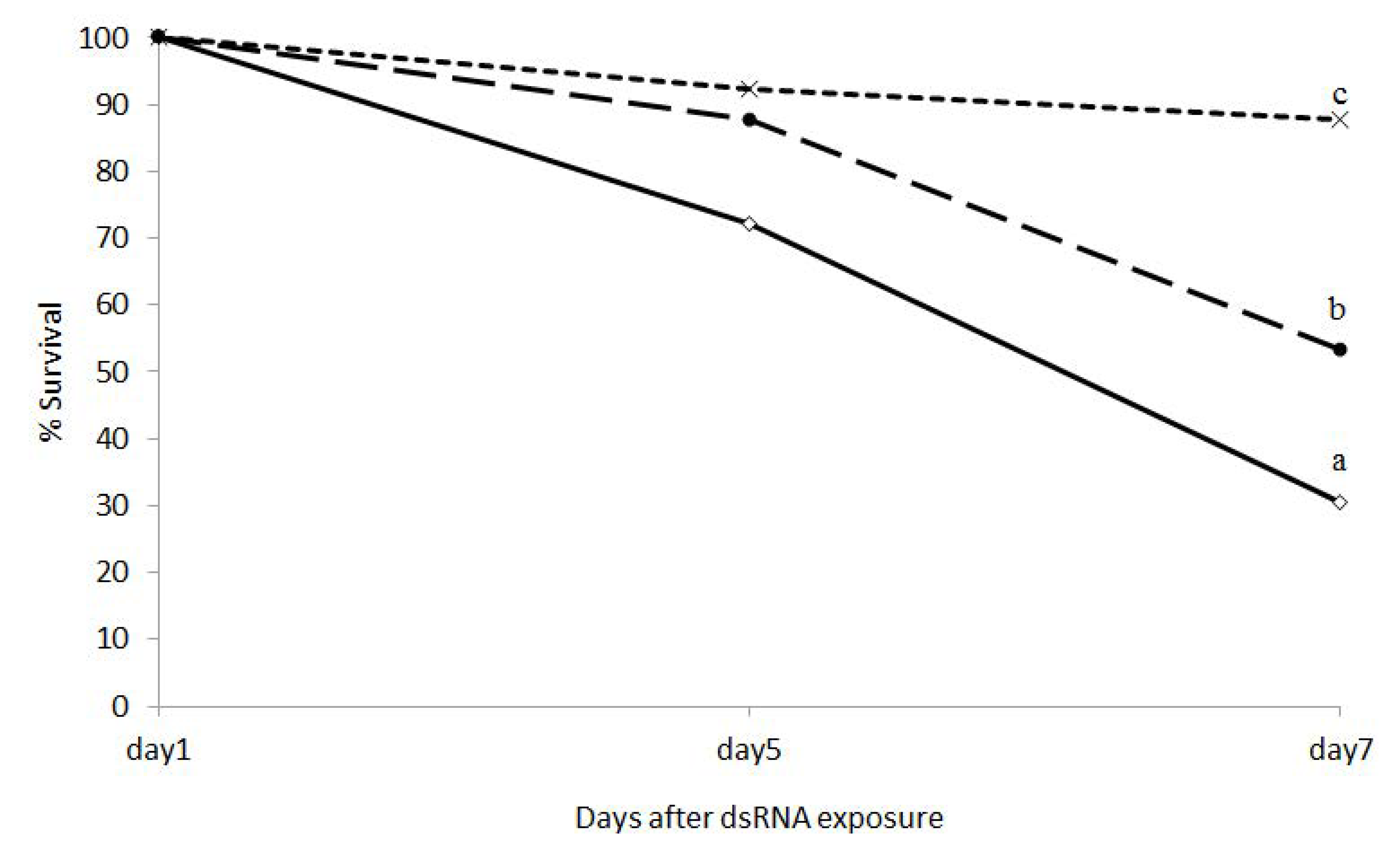
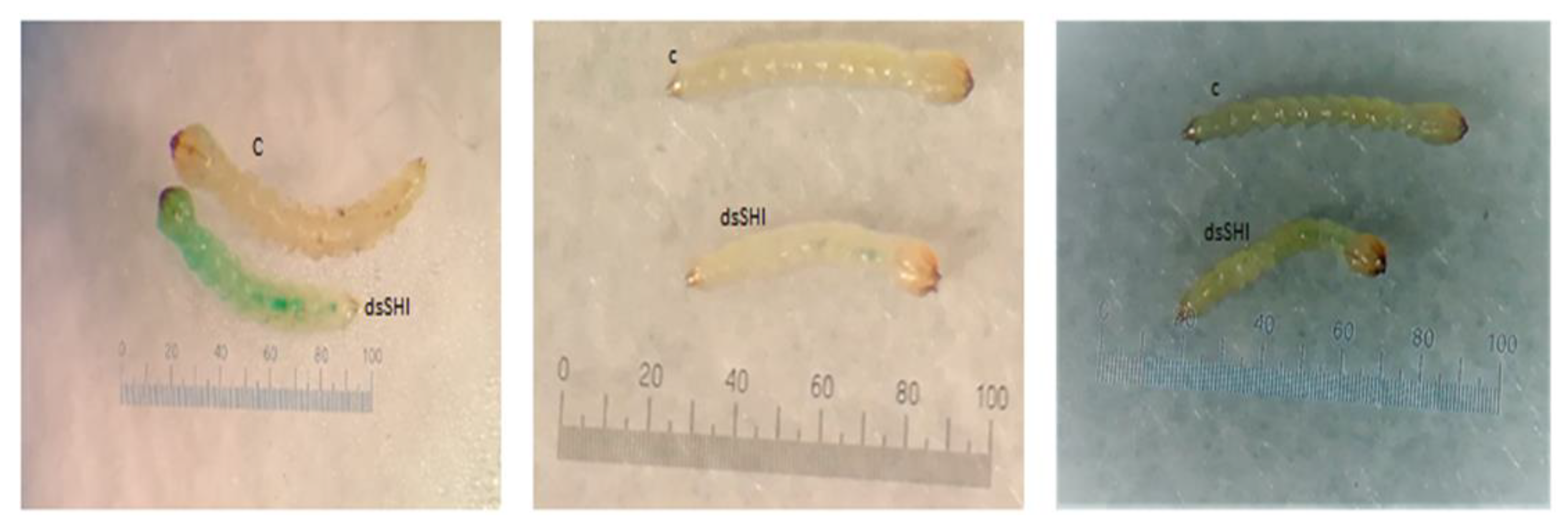
3.2. Biological Activity of dsRNA Expressing E. Coli Against EAB
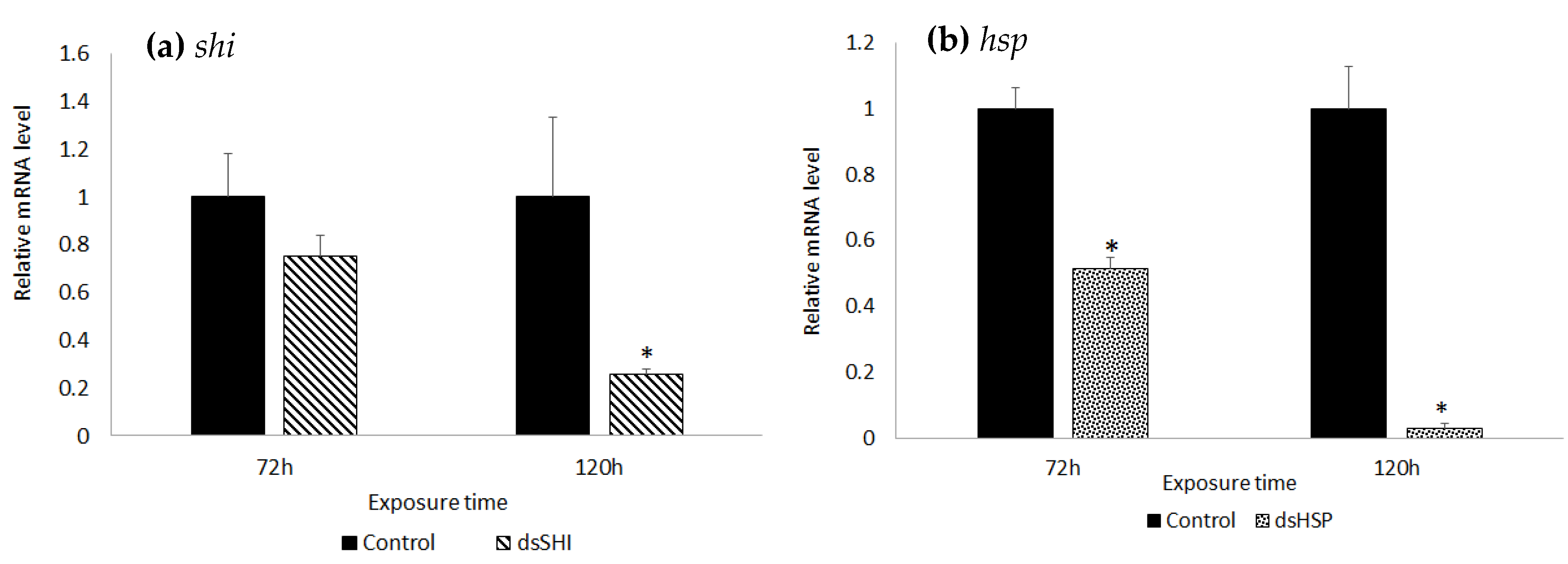
3.3. Molecular Validation of Gene Silencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgement
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Baum, J.A.; Thierry, B.; William, C.; Gregory, R.H.; Pascale, F.; Oliver, I.; Scott, J.; Robert, J. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef]
- Knorr, E.; Fishilevich, E.; Tenbusch, L.; Meghan, L.; Rangasamy, M.; Billion, A.; Sarah, E.; Gandhra, P.; Arora, K.; Narva, K.E. Gene silencing in Tribolium castaneum as a tool for the targeted identification of candidate RNAi targets in crop pests. Sci. Rep. 2018, 8, 2061. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.B.; Duan, J.J.; Palli, S.R.; Rieske, L.K. Identification of highly effective target genes for RNAi-mediated control of emerald ash borer, Agrilus planipennis. Sci. Rep. 2018, 8, 5020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terenius, O.; Paoanicolaou, A.; Garbutt, J.S.; Elefttherianos, I.; Huvenne, H.; Kanginakudru, S. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentice, K.; Pertry, I.; Christiaens, O.; Bauters, L.; Bailey, A.; Niblett, C.; Gislain, M.; Smagghe, G. Transcriptome analysis and systemic RNAi response in the African sweetpotato weevil (Cylas puncticollis, Coleoptera, Brentidae). PLoS ONE 2015, 10, e0115336. [Google Scholar] [CrossRef] [Green Version]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.; Telewski, F.W. Dendrochronological reconstruction of the epicenter and early spread of emerald ash borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]
- Haack, R.A.; Jendek, E.; Liu, H.; Merchant, K.R.; Petrice, T.R.; Poland, T.M.; Ye, H. The emerald ash borer: A new exotic pest in North America. Newsl. Michigan Entomol. Soc. 2012, 47, 1–7. [Google Scholar]
- Rebek, E.J.; Herms, D.A.; Smitley, D.R. Interspecific variation in resistance to emerald ash borer (Coleoptera: Buprestidae) among North American and Asian ash (Fraxinus spp.). Environ. Entomol. 2008, 37, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, W.; Rieske, L.K. Establishment of classical biological control targeting emerald ash borer is facilitated by use of insecticides, with little effect on native arthropod communities. Biol. Control 2016, 101, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Miller, F.; Gould, J.; Fierke, M.; Jones, M.; Kaltenbach, J. Compatibility of insecticides and biocontrol for controlling EAB in urban environments. In Proceedings of the Emerald Ash Borer National Research and Technology Development Meeting, Wooster, OH, USA, 15–16 October 2014. [Google Scholar]
- Duan, J.J.; Abell, J.; Bauer, L.S.; Gould, J.; Van Driesche, R. Natural enemies implicated in the regulation of an invasive pest: A life table analysis of the population dynamics of the emerald ash borer. Agric. Forest Entomol. 2014, 16, 406–416. [Google Scholar] [CrossRef]
- Rodrigues, T.B.; Rieske, L.K.; Duan, J.J.; Mogilicherla, K.; Palli, S.R. Development of RNAi method for screening candidate genes to control emerald ash borer, Agrilus planipennis. Sci. Rep. 2017, 7, 7379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobus, A.P.; Gross, J. Optimal cloning of PCR fragments by homologous recombination in Escherichia coli. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solis, F.C.; Julien, S.R.; Doranda, P.; Christian, W.; Nancy, G. Use of bacterially expressed dsRNA to downregulate Entamoeba histolytica gene expression. PLoS ONE 2009, 4, e8424. [Google Scholar] [CrossRef] [PubMed]
- Al Baki, A.; Jung, J.K.; Kim, Y. Alteration of insulin signaling to control insect pest by using transformed bacteria expressing dsRNA. Pest Manag. Sci. 2020, 76, 1020–1030. [Google Scholar] [CrossRef]
- Rajarapu, S.P.; Mamidala, P.; Mittapalli, O. Validation of reference genes for gene expression studies in the emerald ash borer. Insect Sci. 2011, 19, 41–46. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Timmons, L.; Fire, A. Specific interference by ingested dsRNA. Nature 1998, 395, 854. [Google Scholar] [CrossRef]
- Zhu, F.; Xu, J.; Palli, R.; Ferguson, J.; Palli, S.R. Ingested RNA interference for managing the populations of the Colorado potato beetle, Leptinotarsa decemlineata. Pest Manag. Sci. 2010, 67, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Park, Y.; Kim, Y. A transformed bacterium expressing double-stranded RNA specific to integrin β1enhances Bt toxin efficacy against a polyphagous insect pest, Spodoptera exigua. PLoS ONE 2015, 10, 7. [Google Scholar] [CrossRef]
- Wang, Y.; Zangh, H.; Li, X.; Miao, X. Second generation sequencing supply an effective way to screen RNAi targets in large-scale for potential application in pest insect control. PLoS ONE 2011, 6, e18644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavaria, M.; Gabriele, T.; Kola, I.; Anderson, R.L. A hitchhiker’s guide to the human Hsp70 family. Cell Stress Chaperon. 1996, 1, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, T.; Ikeda, K. Reversible blockage of membrane retrieval and endocytosis in the garland cell of the temperature-sensitive mutant of Drosophila melanogaster, shribirets 1. J. Cell Biol. 1983, 97, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Pampolini, F.B.; Rodrigues, T.B.; Leelesh, R.S.; Kawashima, T.; Rieske, L.K. Confocal microscopy provides visual evidence and confirms the feasibility of dsRNA delivery to emerald ash borer through plant tissues. J. Pest Sci. 2020. [Google Scholar] [CrossRef]
- Tenllado, F.; Martínez-García, B.; Vargas, M.; Diaz-Ruiz, J.R. Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC Biotechnol. 2003, 3. [Google Scholar] [CrossRef] [Green Version]
- Kalantidis, K.; Psaradakis, S.; Tabler, M.; Tsagris, M. The occurrence of CMV-specific short RNAs in transgenic tobacco expressing virus-derived double-stranded RNA is indicative of resistance to the virus. Mol. Plant-Microbe Inter. 2002, 15, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C.; et al. Development and characterization of the first dsRNA-resistant insect population from western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE. 2018, 13, e0197059. [Google Scholar] [CrossRef] [Green Version]
- William, M.; Clark, G.; Sathasivan, K.; Islam, A.S. RNA interference and its application in crop improvement. Plant Tissue Cult. Biotechnol. 2004, 1, 18. [Google Scholar]
- Burand, J.P.; Hunter, W.B. RNAi: Future in insect management. J. Invert. Pathol. 2013, 112, S68–S74. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Andrade, E.C.; Hunter, W.B.; Christiaens, O.; Smagghe, G. Asian citrus psyllid RNAi pathway—RNAi evidence. Sci. Rep. 2016, 6, 38082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyre, B.R.; Rodrigues, T.B.; Rieske, L.K. RNA interference and validation of reference genes for gene expression analyses using RT-qPCR in southern pine beetle, Dendroctonus frontalis. Sci. Rep. 2019, 9, 5640. [Google Scholar] [CrossRef]
- Kyre, B.R.; Bentz, B.J.; Rieske, L.K. Susceptibility of mountain pine beetle (Dendroctonus ponderosae Hopkins) to gene silencing through RNAi provides potential as a novel management tool. For. Ecol. 2020, in press. [Google Scholar]
- Hunter, W.B.; Sinisterra-Hunter, X. Emerging RNA suppression technologies to protect citrus trees from citrus greening disease bacteria. Adv. Insect Physiol. 2018, 55, 163–199. [Google Scholar] [CrossRef]
- Cagliari, D.; Dias, N.P.; Galdeano, D.M.; dos Santos, E.Á.; Smagghe, G.; Zotti, M.J. Management of pest insects and plant diseases by non-transformative RNAi. Front. Plant Sci. 2019, 10, 1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A perspective on RNAi-based biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herms, D.A.; McCullough, D.G.; Smitley, D.R.; Sadof, C.S.; Cranshaw, W. Insecticide options for protecting ash trees from emerald ash borer. North Central IPM Center Bull. 2009, 3–14. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Primer Sequence (5′–3′) |
|---|---|---|
| hsp—heat shock protein (70kDA) | F-dsRNA-HSP | CTAGTCTAGAGTTACGAGCCAGGGTGAAAA |
| R-dsRNA-HSP | TCCCCCCGGGCCTTTTGAACGGCACGGTTAT | |
| F-qRNA-HSP | GACAAAGGAACGGGAAACAA | |
| R-qRNA-HSP | TCTCGGCATCCCTTATCATC | |
| shi—shibire | F-dsRNA-SHI | CTAGTCTAGATGGCACATTTGTATGCCAGT |
| R-dsRNA-SHI | TCCCCCCGGGCTTGTTGCATTTGCTGAGGA | |
| F-qRNA-SHI | GGGATCTGCCCAAATTAACA | |
| R-qRNA-SHI | CCCGTCTGAGTTCTTTCTCG | |
| TEF-1α-Translation elongation factor 1 alpha | F-qRNA-TEF | CATTGAAACCTACGTTGTCGC |
| R-qRNA-TEF | ACTGGAGTGCTTAAACCTGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leelesh, R.S.; Rieske, L.K. Oral Ingestion of Bacterially Expressed dsRNA Can Silence Genes and Cause Mortality in a Highly Invasive, Tree-Killing Pest, the Emerald Ash Borer. Insects 2020, 11, 440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070440
Leelesh RS, Rieske LK. Oral Ingestion of Bacterially Expressed dsRNA Can Silence Genes and Cause Mortality in a Highly Invasive, Tree-Killing Pest, the Emerald Ash Borer. Insects. 2020; 11(7):440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070440
Chicago/Turabian StyleLeelesh, Ramya Shanivarsanthe, and Lynne K. Rieske. 2020. "Oral Ingestion of Bacterially Expressed dsRNA Can Silence Genes and Cause Mortality in a Highly Invasive, Tree-Killing Pest, the Emerald Ash Borer" Insects 11, no. 7: 440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070440