A Comparison of the Vertical Transmission of High- and Low-Virulence Nucleopolyhedrovirus Strains in Lymantria Dispar L.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Virus
2.2. Virus Treatments and the Life-History Traits of Surviving Parents
2.3. Estimation of the Transmitted Virus in Offspring
2.4. Statistical Analysis
3. Results
3.1. Virus Treatment of the Stock Lymantria Dispar Population and the Life-History Traits of Surviving Parents
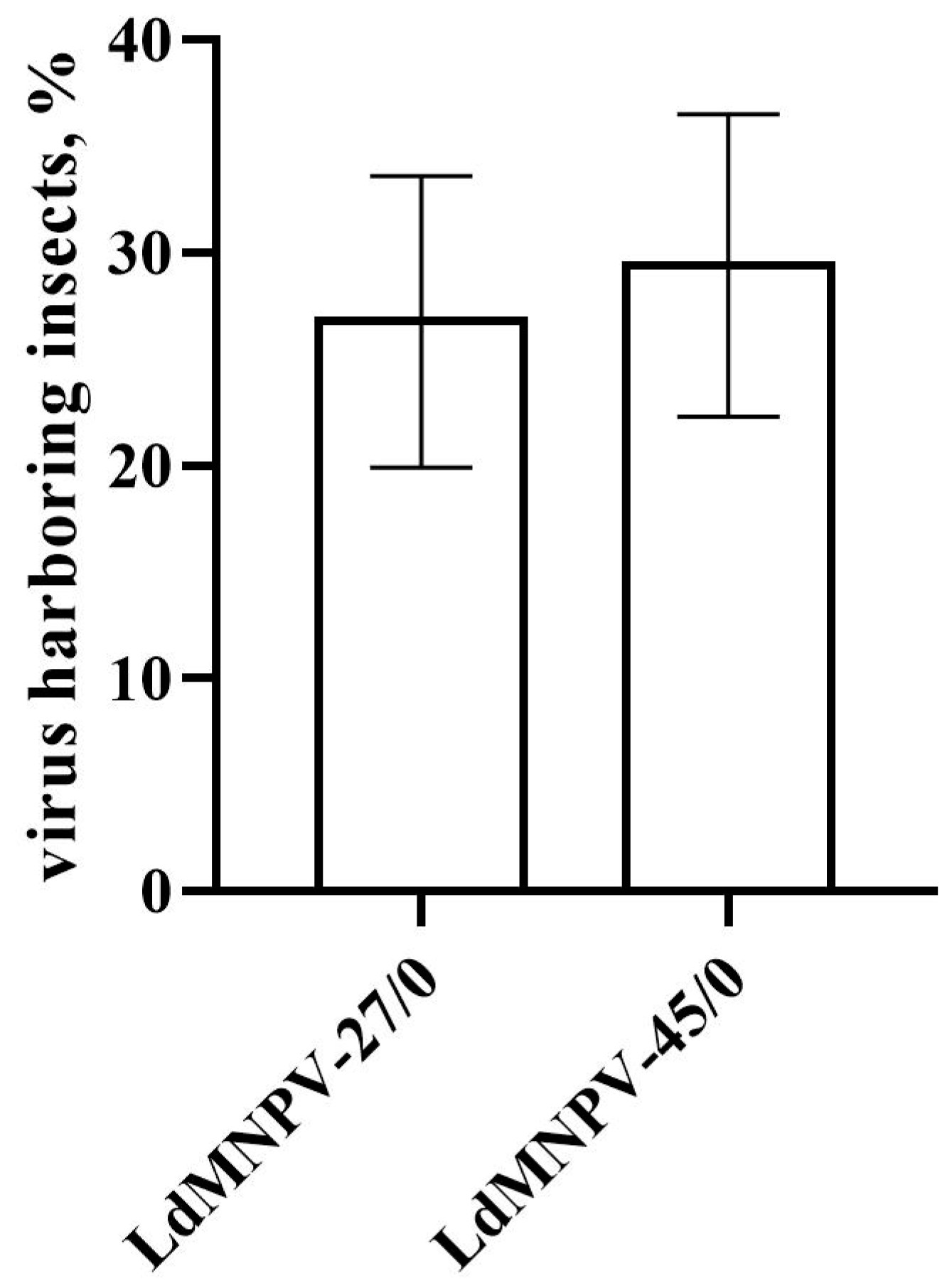
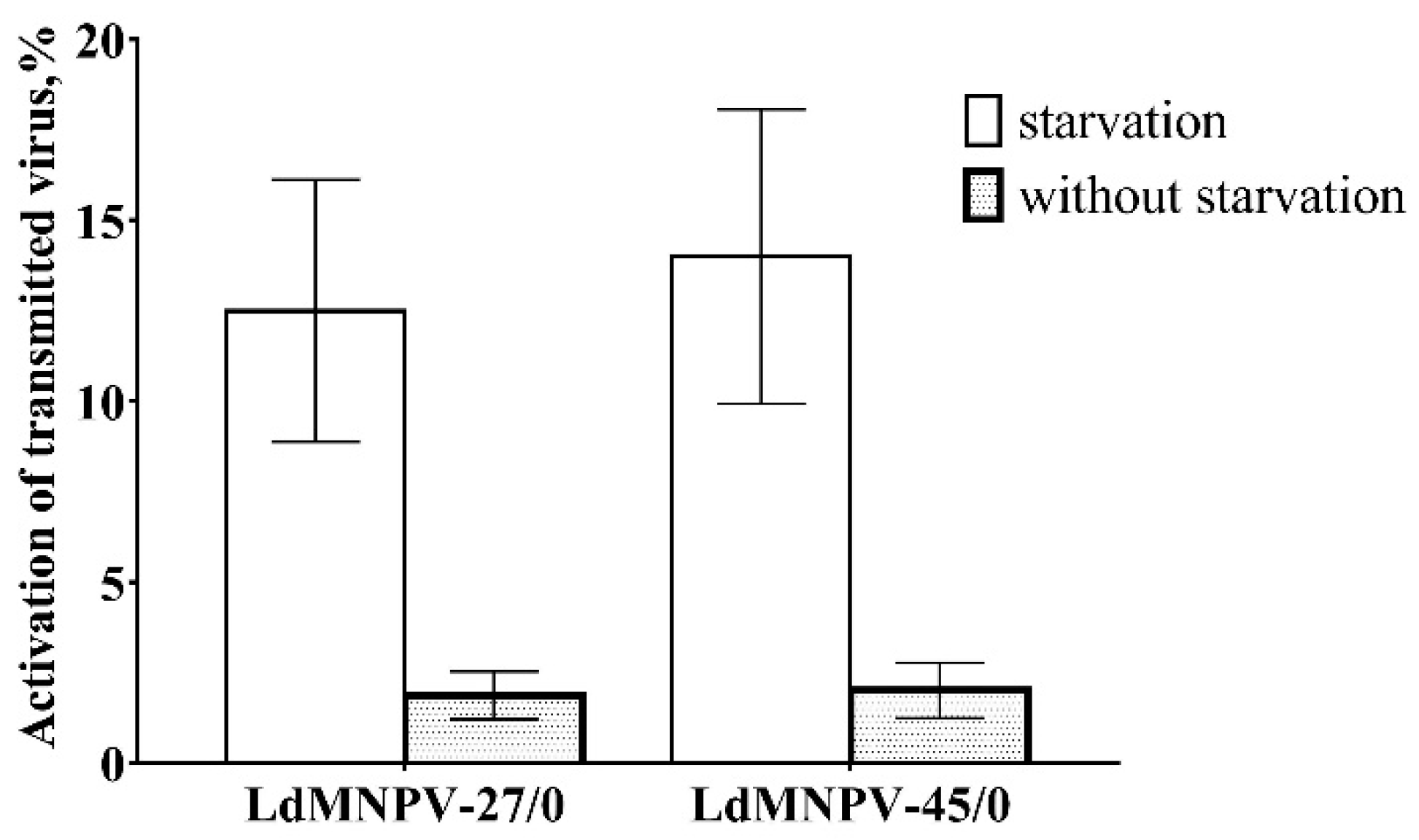
3.2. Estimation of the Transmitted Virus in Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kendall, B.E.; Briggs, C.J.; Murdoch, W.W.; Turchin, P.; Ellner, S.P.; Mccauley, E.; Nisbet, R.M.; Wood, S.N. Why do populations cycle? A synthesis of statistical and mechanistic modeling approaches. Ecology 1999, 80, 1789–1805. [Google Scholar] [CrossRef]
- Myers, J.H. Population cycles: Generalities, exceptions and remaining mysteries. Proc. R. Soc. B Biol. Sci. 2018, 285, e20172841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.H.; Cory, J.S. Ecology and evolution of pathogens in natural populations of Lepidoptera. Evol. Appl. 2016, 9, 231–247. [Google Scholar] [CrossRef] [Green Version]
- Fuxa, J.R. Ecology of insect nucleopolyhedroviruses. Agric. Ecosyst. Environ. 2004, 103, 27–43. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscardi, F. Assessment of the Application of Baculoviruses for Control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef] [PubMed]
- van Frankenhuyzen, K.; Reardon, R.C.; Dubois, N.R. Forest defoliators. In Field Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Kaya, H.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-5933-9. [Google Scholar]
- Elderd, B.D.; Rehill, B.J.; Haynes, K.J.; Dwyer, G. Induced plant defenses, host-pathogen interactions, and forest insect outbreaks. Proc. Natl. Acad. Sci. USA 2013, 110, 14978–14983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slack, J.; Arif, B.M. The Baculoviruses Occlusion-Derived Virus: Virion Structure and Function. Adv. Virus Res. 2006, 69, 99–165. [Google Scholar] [CrossRef]
- Cory, J.S.; Myers, J.H. The Ecology and Evolution of Insect Baculoviruses. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 239–272. [Google Scholar] [CrossRef] [Green Version]
- Cory, J.S. Insect virus transmission: Different routes to persistence. Curr. Opin. Insect Sci. 2015, 8, 130–135. [Google Scholar] [CrossRef]
- Kukan, B. Vertical Transmission of Nucleopolyhedrovirus in Insects. J. Invertebr. Pathol. 1999, 74, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doane, C.C. Trans-ovum transmission of a nuclear-polyhedrosis virus in the gypsy moth and the inducement of virus susceptibility. J. Invertebr. Pathol. 1969, 14, 199–210. [Google Scholar] [CrossRef]
- Burand, J.P.; Horton, H.M.; Retnasami, S.; Elkinton, J.S. The use of polymerase chain reaction and shortwave UV irradiation to detect baculovirus DNA on the surface of gypsy moth eggs. J. Virol. Methods 1992, 36, 141–149. [Google Scholar] [CrossRef]
- Burden, J.P.; Nixon, C.P.; Hodgkinson, A.E.; Possee, R.D.; Sait, S.M.; King, L.A.; Hails, R.S. Covert infections as a mechanism for long-term persistence of baculoviruses. Ecol. Lett. 2003, 6, 524–531. [Google Scholar] [CrossRef]
- Cabodevilla, O.; Villar, E.; Virto, C.; Murillo, R.; Williams, T.; Caballero, P. Intra- and intergenerational persistence of an insect nucleopolyhedrovirus: Adverse effects of sublethal disease on host development, reproduction, and susceptibility to superinfection. Appl. Environ. Microbiol. 2011, 77, 2954–2960. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.; Virto, C.; Murillo, R.; Caballero, P. Covert infection of insects by baculoviruses. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Graham, R.I.; Tummala, Y.; Rhodes, G.; Cory, J.S.; Shirras, A.; Grzywacz, D.; Wilson, K. Development of a real-time qPCR assay for quantification of covert baculovirus infections in a major african crop pest. Insects 2015, 6, 746–759. [Google Scholar] [CrossRef] [Green Version]
- Burden, J.P.; Griffiths, C.M.; Cory, J.S.; Smith, P.; Sait, S.M. Vertical transmission of sublethal granulovirus infection in the Indian meal moth, Plodia interpunctella. Mol. Ecol. 2002, 11, 547–555. [Google Scholar] [CrossRef]
- Kane, M.; Golovkina, T. Common Threads in Persistent Viral Infections. J. Virol. 2010, 84, 4116–4123. [Google Scholar] [CrossRef] [Green Version]
- Fuxa, J.R.; Sun, J.Z.; Weidner, E.H.; Lamotte, L.R. Stressors and Rearing Diseases of Trichoplusia ni: Evidence of Vertical Transmission of NPV and CPV. J. Invertebr. Pathol. 1999, 74, 149–155. [Google Scholar] [CrossRef]
- Hughes, D.S.; Possee, R.D.; King, L.A. Activation and Detection of a Latent Baculovirus Resembling Mamestra brassicae Nuclear Polyhedrosis Virus in M. brassicae Insects. Virology 1993, 194, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Kasianov, N.S.; Belousova, I.A.; Pavlushin, S.V.; Dubovskiy, I.M.; Podgwaite, J.D.; Martemyanov, V.V.; Bakhvalov, S.A. The activity of phenoloxidase in haemolymph plasma is not a predictor of Lymantria dispar resistance to its baculovirus. PLoS ONE 2017, 12, e0183940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlushin, S.; Belousova, I.; Chertkova, E.; Akhanaev, Y.; Martemyanov, V.; Glupov, V. Effect of starvation as a population stress-factor on activation of covert baculovirus infection in gypsy moth. Zhu. Obsh. Bio. 2020, 81, 31–36. [Google Scholar] [CrossRef]
- Myers, J.H.; Cory, J.S.; Ericsson, J.D.; Tseng, M.L. The effect of food limitation on immunity factors and disease resistance in the western tent caterpillar. Oecologia 2011, 167, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Martemyanov, V.V.; Pavlushin, S.V.; Dubovskiy, I.M.; Yushkova, Y.V.; Morosov, S.V.; Chernyak, E.I.; Efimov, V.M.; Ruuhola, T.; Glupov, V.V. Asynchrony between host plant and insects-defoliator within a tritrophic system: The role of herbivore innate immunity. PLoS ONE 2015. [Google Scholar] [CrossRef] [Green Version]
- Podgwaite, J.D.; Mazzone, H.M. Latency of Insect Viruses. Adv. Virus Res. 1986, 31, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Podgwaite, J.D. Gypchek: Biological Insecticide for the Gypsy Moth. J. For. 1999, 97, 16–19. [Google Scholar] [CrossRef]
- Elkinton, J.S. Gypsy Moth. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 435–439. ISBN 9780123741448. [Google Scholar]
- Harrison, R.L.; Keena, M.A.; Rowley, D.L. Classification, genetic variation and pathogenicity of Lymantria dispar nucleopolyhedrovirus isolates from Asia, Europe, and North America. J. Invertebr. Pathol. 2014, 116, 27–35. [Google Scholar] [CrossRef]
- Harrison, R.L.; Rowley, D.L.; Keena, M.A. Geographic isolates of Lymantria dispar multiple nucleopolyhedrovirus: Genome sequence analysis and pathogenicity against European and Asian gypsy moth strains. J. Invertebr. Pathol. 2016, 137, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.L.; Lynn, D.E. Genomic sequence analysis of a nucleopolyhedrovirus isolated from the diamondback moth, Plutella xylostella. Virus Genes 2007, 35, 857–873. [Google Scholar] [CrossRef]
- Shapiro, M.; Robertson, J.L.; Injac, M.G.; Katagiri, K.; Bell, R.A. Comparative Infectivities of Gypsy Moth (Lepidoptera: Lymantriidae) Nucleopolyhedrosis Virus Isolates from North America, Europe, and Asia. J. Econ. Entomol. 1984, 77, 153–156. [Google Scholar] [CrossRef]
- Podgwaite, J.D.; Martemyanov, V.V.; Slavicek, J.M.; Bakhvalov, S.A.; Pavlushin, S.V.; Hayes-Plazolles, N.; Zerillo, R.T. Potency of Nucleopolyhedrovirus Genotypes for European and Asian Gypsy Moth (Lepidoptera: Lymantriidae). J. Entomol. Sci. 2013, 48, 332–344. [Google Scholar] [CrossRef]
- Narang, N.; Hérard, F.; Dougherty, E.M.; Chen, K.; Vega, F.E. A gypsy moth (Lymantria dispar, Lepidoptera: Llymantriidae) multinucleocapsid nuclear polyhedrosis virus from France: Comparison with a North American and a Korean strain. Eur. J. Entomol. 2001. [Google Scholar] [CrossRef] [Green Version]
- Martemyanov, V.V.; Podgwaite, J.D.; Belousova, I.A.; Pavlushin, S.V.; Slavicek, J.M.; Baturina, O.A.; Kabilov, M.R.; Ilyinykh, A.V. A comparison of the adaptations of strains of Lymantria dispar multiple nucleopolyhedrovirus to hosts from spatially isolated populations. J. Invertebr. Pathol. 2017, 146, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Ilyinykh, A.V.; Polenogova, O.V. Demonstration of the remote effect of baculovirus vertical transmission, with gypsy moth Lymantria dispar L. (Lepidoptera, Lymantriidae)as an example. Zh. Obshch. Biol. 2012, 73, e389. [Google Scholar] [CrossRef]
- Castro, M.E.B.; Ribeiro, Z.M.A.; Santos, A.C.B.; Souza, M.L.; Machado, E.B.; Sousa, N.J.; Moscardi, F. Identification of a new nucleopolyhedrovirus from naturally-infected Condylorrhiza vestigialis (Guenée) (Lepidoptera: Crambidae) larvae on poplar plantations in South Brazil. J. Invertebr. Pathol. 2009, 102, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Akhanaev, Y.B.; Belousova, I.A.; Ershov, N.I.; Nakai, M.; Martemyanov, V.V.; Glupov, V.V. Comparison of tolerance to sunlight between spatially distant and genetically different strains of Lymantria dispar nucleopolyhedrovirus. PLoS ONE 2017, 12, e0189992. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, e9. [Google Scholar]
- R Development Core Team. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 19 July 2020).
- Murray, K.D.; Shields, K.S.; Burand, J.P.; Elkinton, J.S. The effect of gypsy moth metamorphosis on the development of nuclear polyhedrosis virus infection. J. Invertebr. Pathol. 1991, 57, 352–361. [Google Scholar] [CrossRef]
- Shapiro, M.; Robertson, J.L. Yield and Activity of Gypsy Moth (Lepidoptera: Lymantriidae) Nucleopolyhedrosis Virus Recovered from Survivors of Viral Challenge. J. Econ. Entomol. 1987, 80, 901–905. [Google Scholar] [CrossRef]
- Myers, J.H.; Malakar, R.; Cory, J.S. Sublethal Nucleopolyhedrovirus Infection Effects on Female Pupal Weight, Egg Mass Size, and Vertical Transmission in Gypsy Moth (Lepidoptera: Lymantriidae). Environ. Entomol. 2000, 29, 1268–1272. [Google Scholar] [CrossRef] [Green Version]
- Cabodevilla, O.; Ibañez, I.; Simón, O.; Murillo, R.; Caballero, P.; Williams, T. Occlusion body pathogenicity, virulence and productivity traits vary with transmission strategy in a nucleopolyhedrovirus. Biol. Control 2010, 56, 184–192. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Selection for an increased rate of vertical transmission of Spodoptera frugiperda (Lepidoptera: Noctuidae) nuclear polyhedrosis virus. Environ. Entomol. 1991, 20, 603–609. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Maruniak, J.E.; Richter, A.R. Characterization of the DNA of a nuclear polyhedrosis virus selected for an increased rate of vertical transmission. J. Invertebr. Pathol. 1994, 64, 1–5. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Virulence and multigeneration passage of a nuclear polyhedrosis virus selected for an increased rate of vertical transmission. Biol. Control 1992, 2, 171–175. [Google Scholar] [CrossRef]
- Martemyanov, V.V.; Kabilov, M.R.; Tupikin, A.E.; Baturina, O.A.; Belousova, I.A.; Podgwaite, J.D.; Ilynykh, A.V.; Vlassov, V.V. The enhancin gene: One of the genetic determinants of population variation in baculoviral virulence. Dokl. Biochem. Biophys. 2015, 465, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Cory, J.S.; Theilmann, D.A.; Myers, J.H. Nucleopolyhedroviruses of forest and western tent caterpillars: Cross-infectivity and evidence for activation of latent virus in high-density field populations. Ecol. Entomol. 2003, 28, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Fuxa, J.R.; Richter, A.R.; Ameen, A.O.; Hammock, B.D. Vertical transmission of TnSNPV, TnCPV, AcMNPV, and possibly recombinant NPV in Trichoplusia ni. J. Invertebr. Pathol. 2002, 79, 44–50. [Google Scholar] [CrossRef]
- Murillo, R.; Hussey, M.S.; Possee, R.D. Evidence for covert baculovirus infections in a spodoptera exigua laboratory culture. J. Gen. Virol. 2011, 92, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.M.; Zhai, W.J.; Li, B.C.; Zhang, W.; Zhang, Z.Q.; Li, M.L.; Zhang, Y.A. Covert LdMNPV detected in Lymantria dispar larvae during a survey of potential hosts for DekiNPV production. Int. J. Pest Manag. 2015, 61, 26–29. [Google Scholar] [CrossRef]
- Pavlushin, S.V.; Belousova, I.A.; Chertkova, E.A.; Kryukova, N.A.; Glupov, V.V.; Martemyanov, V.V. The effect of population density of Lymantria dispar (Lepidoptera: Erebidae) on its fitness, physiology and activation of the covert nucleopolyhedrovirus. Eur. J. Entomol. 2019, 116, 85–91. [Google Scholar] [CrossRef]
- Virto, C.; Navarro, D.; Tellez, M.M.; Murillo, R.; Williams, T.; Caballero, P. Chemical and biological stress factors on the activation of nucleopolyhedrovirus infections in covertly infected Spodoptera exigua. J. Appl. Entomol. 2017, 141, 384–392. [Google Scholar] [CrossRef]
- Ilyinykh, A.V.; Shternshis, M.V.; Kuzminov, S.V. Exploration into a mechanism of transgenerational transmission of nucleopolyhedrovirus in Lymantria dispar L. in Western Siberia. BioControl 2004, 49, 441–454. [Google Scholar] [CrossRef]
- Cory, J.S.; Green, B.M.; Paul, R.K.; Hunter-Fujita, F. Genotypic and phenotypic diversity of a baculovirus population within an individual insect host. J. Invertebr. Pathol. 2005, 89, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Rothman, L.D.; Myers, J.H. Debilitating effects of viral diseases on host Lepidoptera. J. Invertebr. Pathol. 1996, 67, 1–10. [Google Scholar] [CrossRef]
- Miller, W.E. Extrinsic effects on fecundity-maternal weight relations in capital-breeding Lepidoptera. J. Lepid. Soc. 2005, 59, 143–160. [Google Scholar]
- Rothman, L.D.; Myers, J.H. Nuclear polyhedrosis virus treatment effect on reproductive potential of western tent caterpillar (Lepidoptera: Lasiocampidae). Environ. Entomol. 1994, 23, 864–869. [Google Scholar] [CrossRef]
- Kemp, E.M.; Woodward, D.T.; Cory, J.S. Detection of single and mixed covert baculovirus infections in eastern spruce budworm, Choristoneura fumiferana populations. J. Invertebr. Pathol. 2011, 107, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Vilaplana, L.; Wilson, K.; Redman, E.M.; Cory, J.S. Pathogen persistence in migratory insects: High levels of vertically-transmitted virus infection in field populations of the African armyworm. Evol. Ecol. 2010, 24, 147–160. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Cases | Median Development Time, Days | Mean ± SE Pupal Weight, g | Sex Ratio, % of Male | Mean ± SE Fecundity, Egg | Mean ± SE Fertility, % | |||
|---|---|---|---|---|---|---|---|---|
| Larvae | Pupae | |||||||
| Male | Female | Male | Female | |||||
| LdMNPV-45/0 | 31 (n = 119) C | 34 (n = 93) C | 12 (n = 119) A | 11 (n = 93) A | 1.06 ± 0.03 (n = 215) C | 64.6 (n = 113) B | 129.5 ± 16.6 (n = 29) B | 37.4 ± 4.8 (n = 26) B |
| LdMNPV-27/0 | 33 (n = 41) B | 35 (n = 66) B | 13 (n = 45) A | 11 (n = 72) A | 1.18 ± 0.04 (n = 114) B | 63.8 (n = 72) B | 110.2 ± 18.1 (n = 15) B | 43.4 ± 7.7 (n = 15) B |
| Untreated insects | 30 (n = 108) A | 32 (n = 140) A | NA | NA | 0.82 ± 0.01 (n = 248) A | 44 (n = 248) A | 429.2 ± 56.2 (n = 10) A | 91.6 ± 2 (n = 6) A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhanaev, Y.B.; Belousova, I.A.; Lebedeva, D.A.; Pavlushin, S.V.; Martemyanov, V.V. A Comparison of the Vertical Transmission of High- and Low-Virulence Nucleopolyhedrovirus Strains in Lymantria Dispar L. Insects 2020, 11, 455. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070455
Akhanaev YB, Belousova IA, Lebedeva DA, Pavlushin SV, Martemyanov VV. A Comparison of the Vertical Transmission of High- and Low-Virulence Nucleopolyhedrovirus Strains in Lymantria Dispar L. Insects. 2020; 11(7):455. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070455
Chicago/Turabian StyleAkhanaev, Yuriy B., Irina A. Belousova, Darya A. Lebedeva, Sergey V. Pavlushin, and Vyacheslav V. Martemyanov. 2020. "A Comparison of the Vertical Transmission of High- and Low-Virulence Nucleopolyhedrovirus Strains in Lymantria Dispar L." Insects 11, no. 7: 455. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070455