Cellular Localization of Two Rickettsia Symbionts in the Digestive System and within the Ovaries of the Mirid Bug, Macrolophous pygmaeus

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Insect Rearing
2.2. Microbiota Characterization
2.3. Screening for Rickettsia bellii and Rickettsia limoniae Prevalance
2.4. Location of R. bellii and R. limoniae in M. pygmaeus
2.4.1. Morphology of the Digestive System
2.4.2. PCR for Ovaries and Digestive System
2.4.3. Fluorescence in Situ Hybridization (FISH)
3. Results
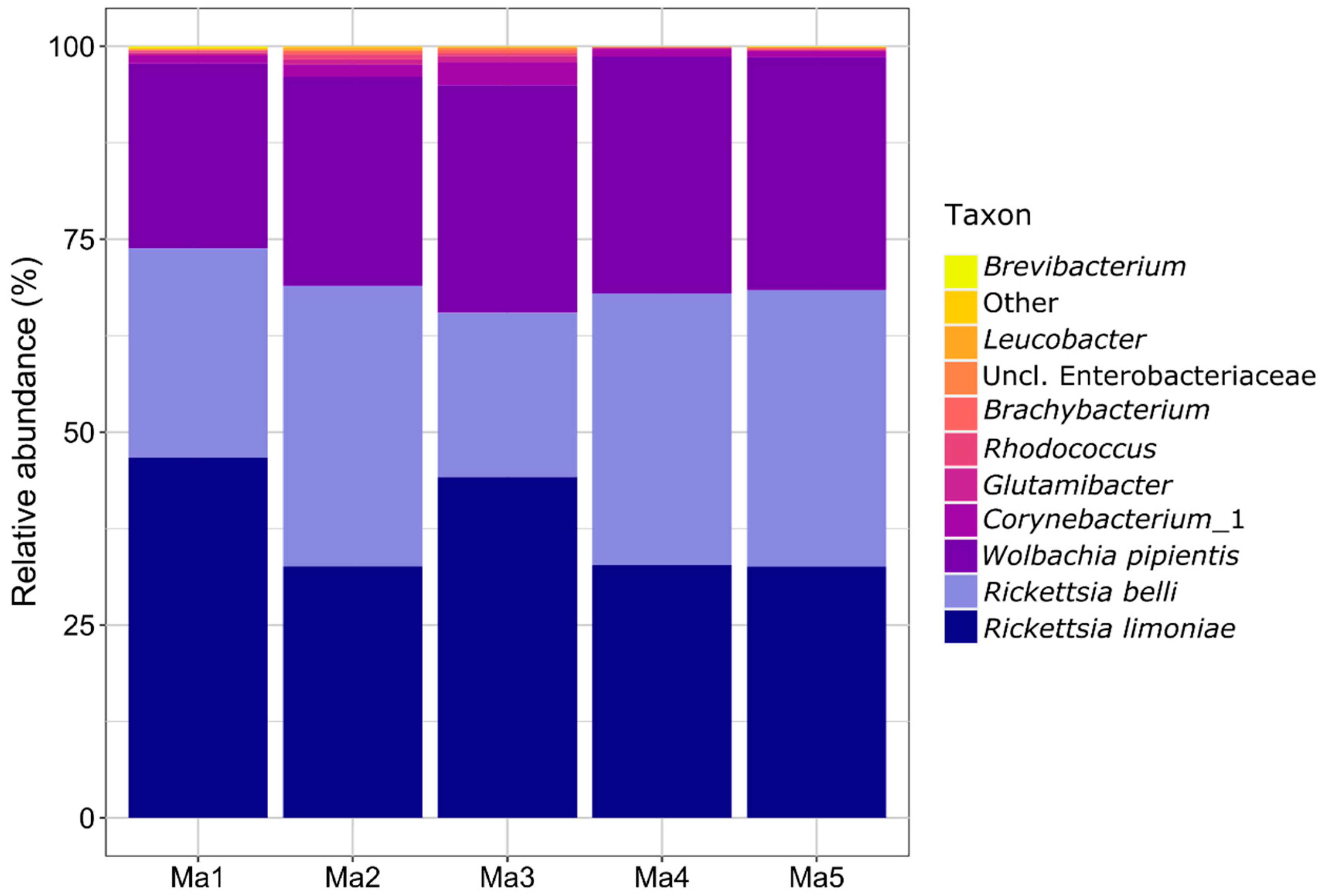
3.1. Microbiota Characterization
3.2. Location of R. bellii and R. limoniae in M. pygmaeus Body
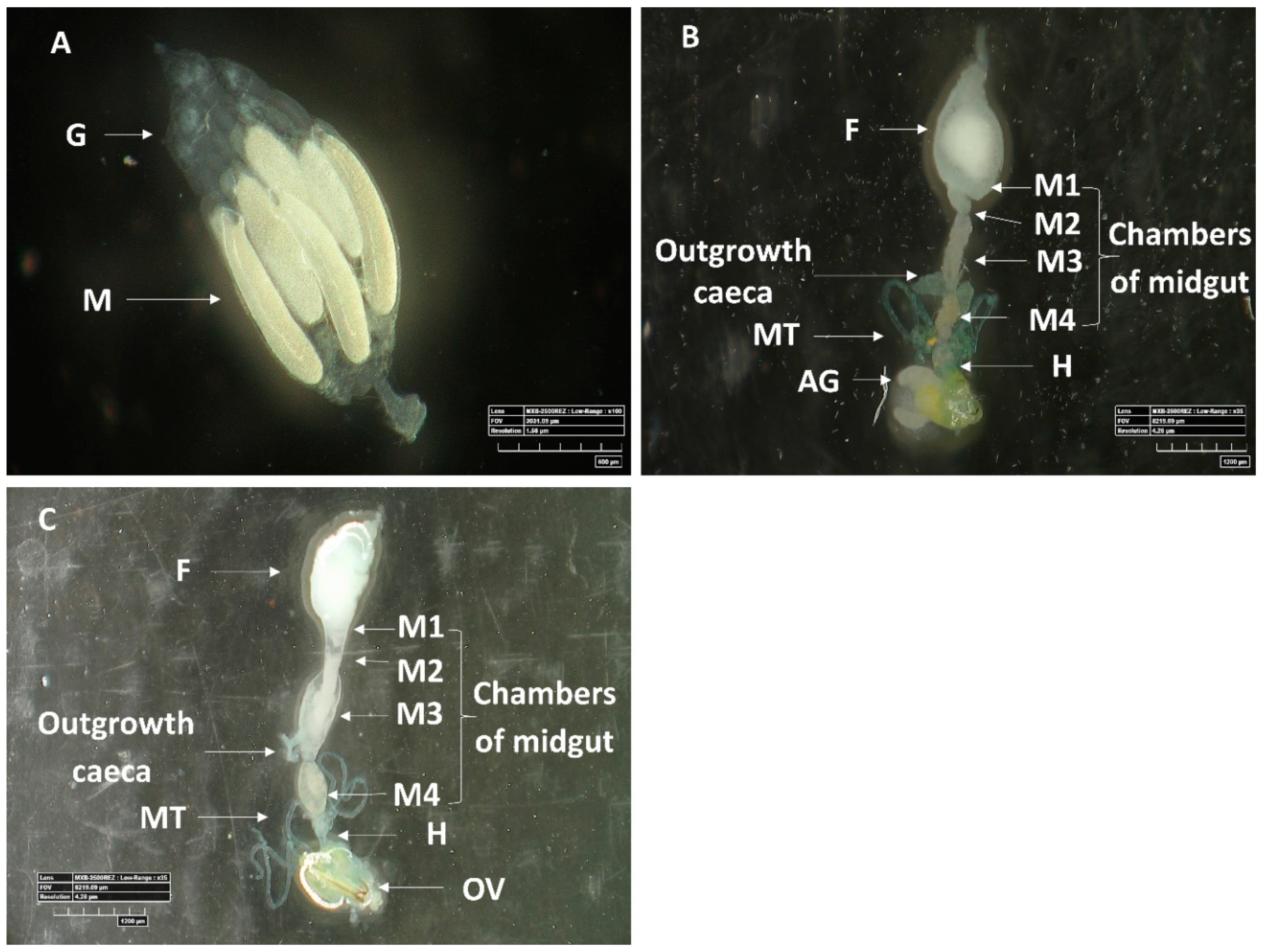
3.2.1. Morphology of Digestive System
3.2.2. PCR for Ovaries and Digestive System
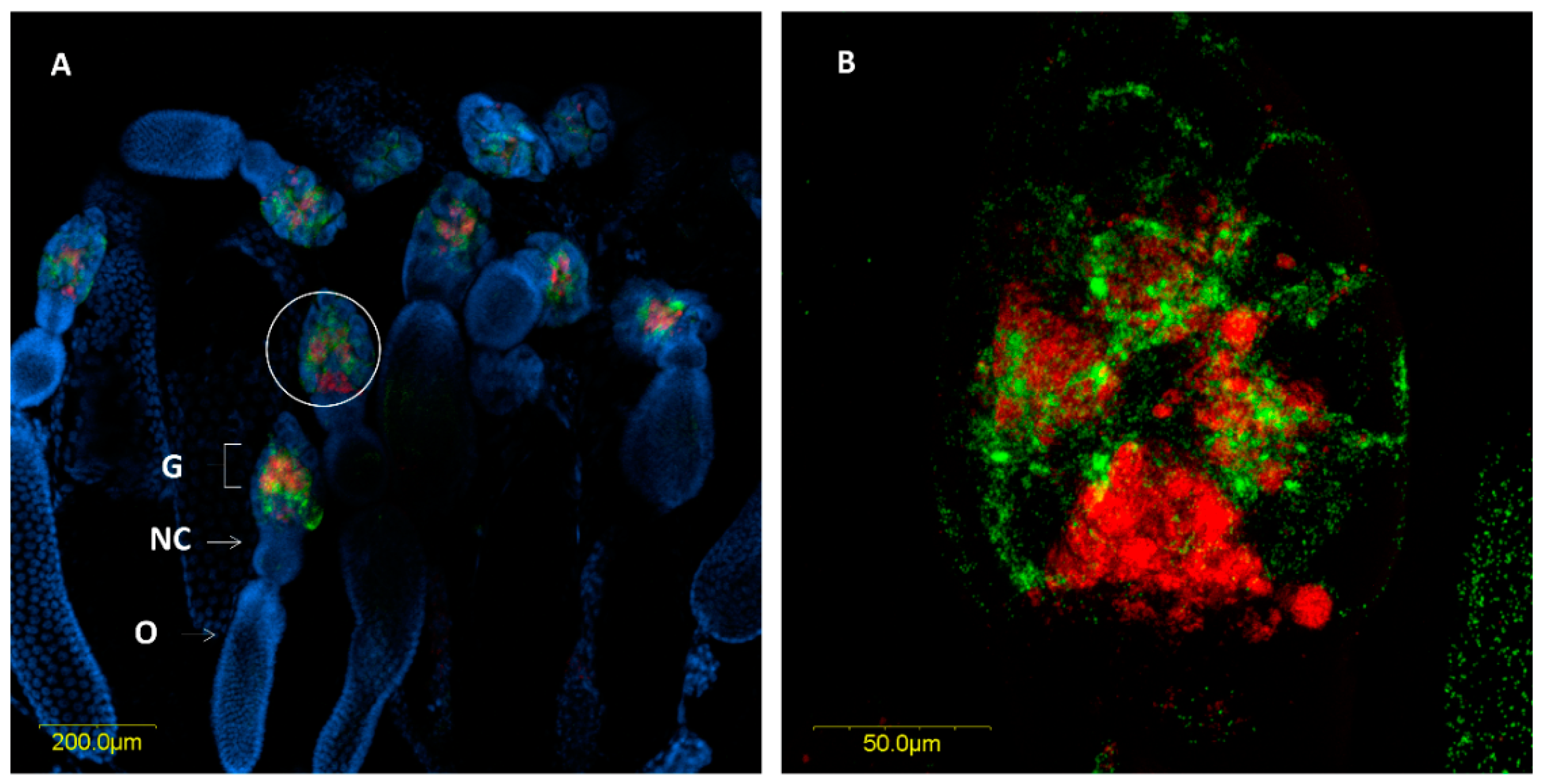
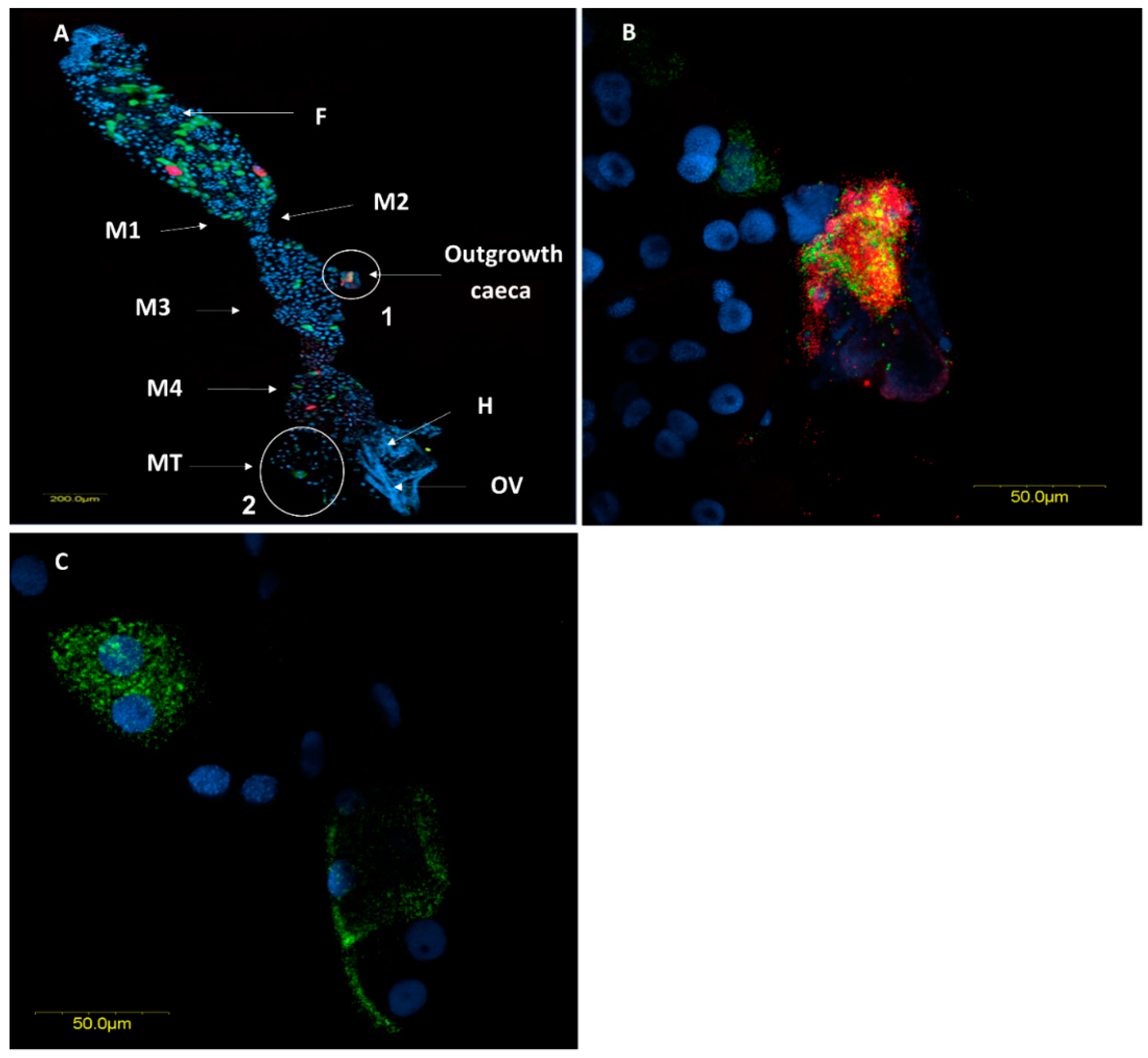
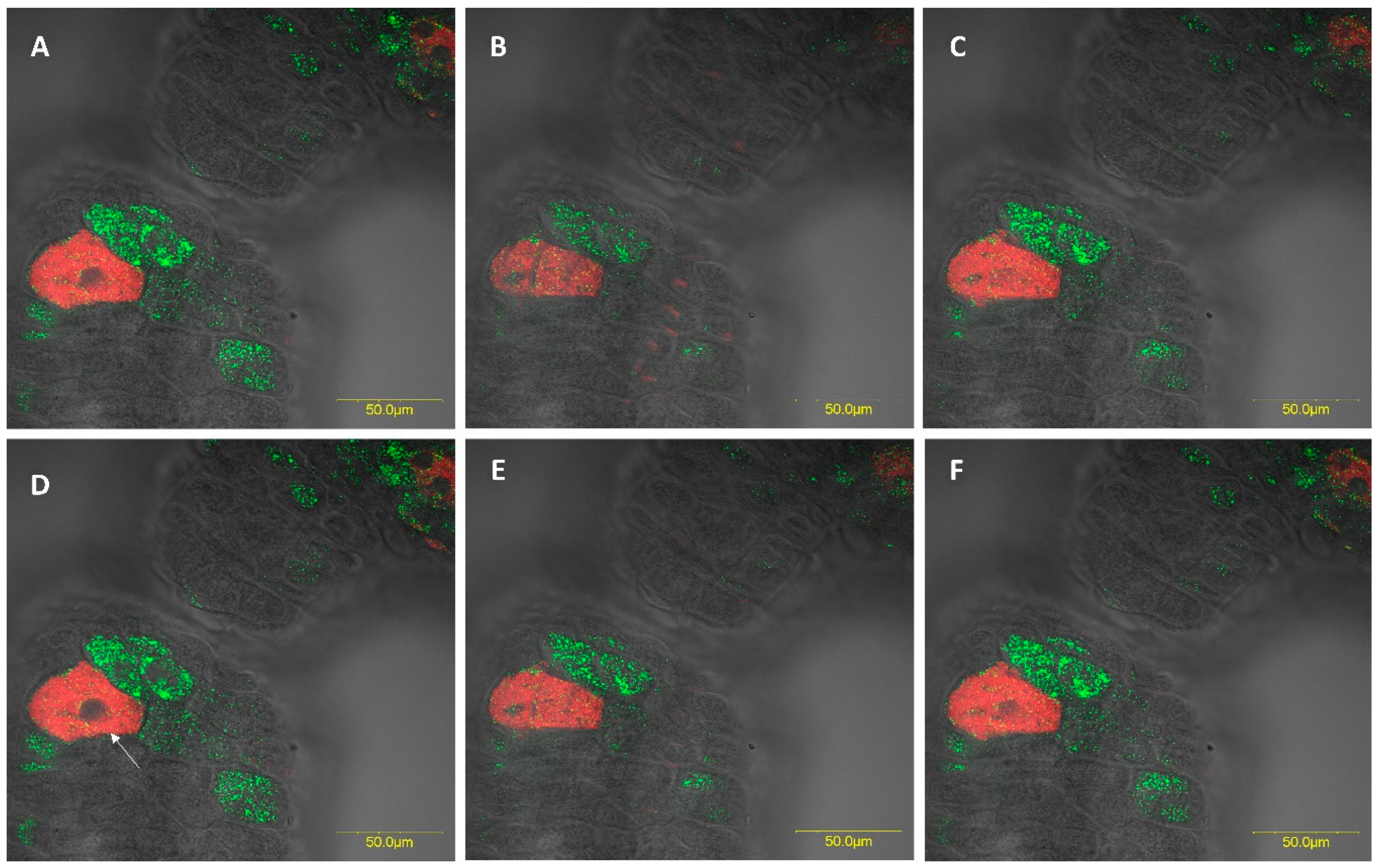
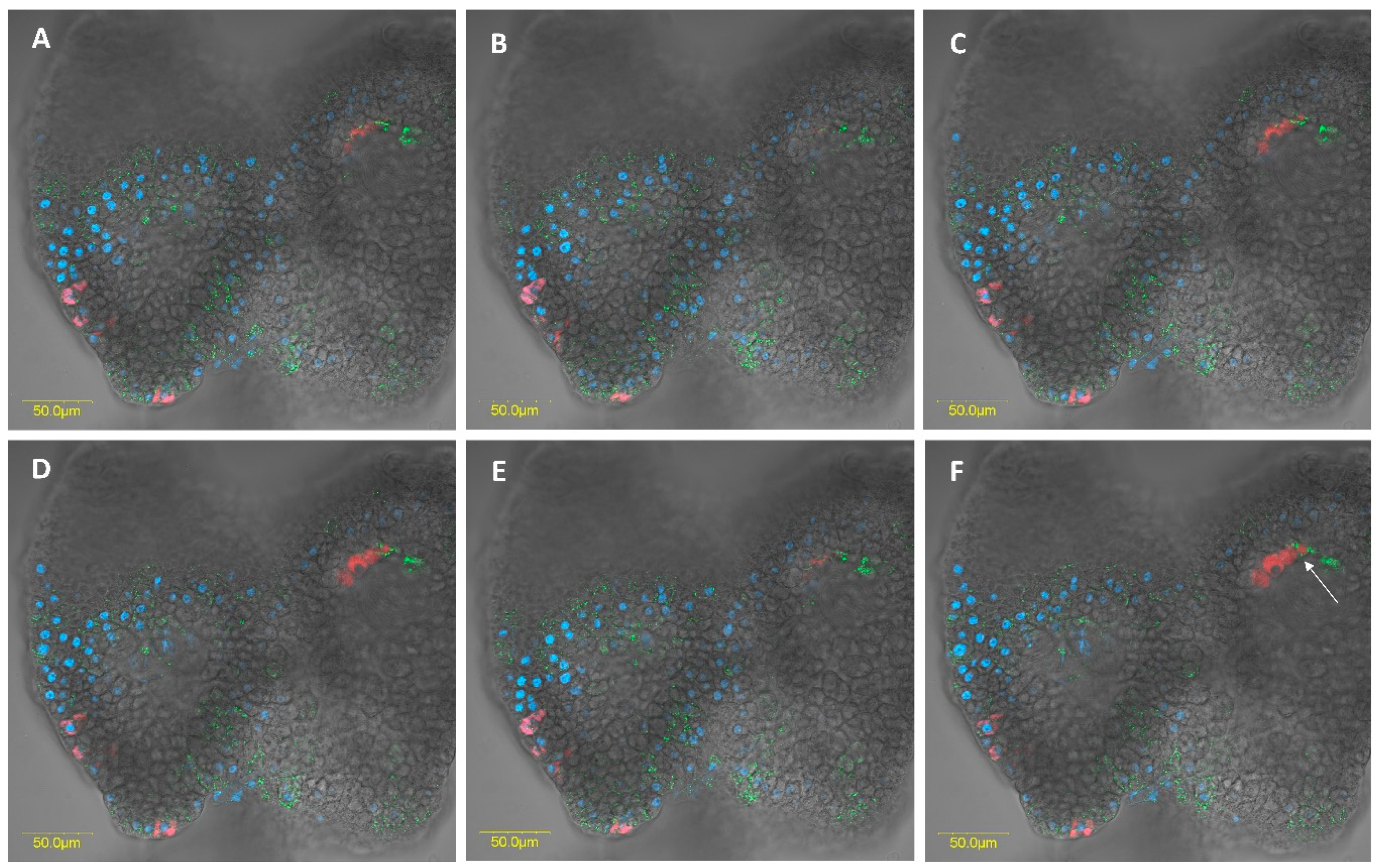
3.2.3. Fluorescence in Situ Hybridization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, S.; Arya, S.K.; Kaur, G.; Saxena, G.; Verma, P.C. Role of endosymbionts in nutritional uptake of sap sucking insects. In Molecular Approaches in Plant Biology and Environmental Challenges. Energy, Environment, and Sustainability; Singh, S.P., Upadhyay, S.K., Pandey, A., Kumar, S., Eds.; Springer: Singapore, 2019; pp. 487–499. [Google Scholar]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shropshire, J.D.; Bordenstein, S.R. Speciation by symbiosis: The microbiome and behavior. MBio 2016, 7, e01785-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.A.; Dubilier, N. Nature’s microbiome: Introduction. Mol. Ecol. 2014, 23, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Perlman, S.J.; Chiel, E.; Zchori-Fein, E. Rickettsia get around. In Manipulative Tenants: Bacteria Associated with Arthropods; Zchori-Fein, E., Bourtzis, K., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 191–206. [Google Scholar]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142957. [Google Scholar] [CrossRef] [Green Version]
- Sudakaran, S.; Kost, C.; Kaltenpoth, M. Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol. 2017, 25, 375–390. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The impact of microbial symbionts on host plant utilization by herbivorous insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef]
- Schmid, R.B.; Lehman, R.M.; Brözel, V.S.; Lundgren, J.G. An indigenous gut bacterium, Enterococcus faecalis (Lactobacillales: Enterococcaceae), increases seed consumption by Harpalus pensylvanicus (Coleoptera: Carabidae). Florida Entomol. 2014, 97, 575–584. [Google Scholar] [CrossRef]
- Barr, K.L.; Hearne, L.B.; Briesacher, S.; Clark, T.L.; Davis, G.E. Microbial symbionts in insects influence down-regulation of defense genes in maize. PLoS ONE 2010, 5, e11339. [Google Scholar] [CrossRef]
- Chung, S.H.; Rosa, C.; Scully, E.D.; Peiffer, M.; Tooker, J.F.; Hoover, K.; Luthe, D.S.; Felton, G.W. Herbivore exploits orally secreted bacteria to suppress plant defenses. Proc. Natl. Acad. Sci. USA 2013, 110, 15728–15733. [Google Scholar] [CrossRef] [Green Version]
- Krinsky, W.L. Hemiptera (True Bugs). In Medical and Veterinary Entomology; Mullen, G.R., Durden, L.A., Eds.; Academic Press: New Haven, CT, USA, 2019; pp. 107–127. [Google Scholar]
- Schaefer, C.W.; Panizzi, A.R. Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Washington, DC, USA, 2000; pp. 3–7. [Google Scholar]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. Insect-microbe mutualism without vertical transmission: A stinkbug acquires a beneficial gut symbiont from the environment every generation. Appl. Environ. Microbiol. 2007, 73, 4308–4316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, A.G. Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists, 1st ed.; Cornell University Press: Ithaca, NY, USA, 2001; pp. 1–4. [Google Scholar]
- Perdikis, D.; Fantinou, A.A.; Lykouressis, D.P. Enhancing pest control in annual crops by conservation of predatory Heteroptera. Biol. Control 2011, 59, 13–21. [Google Scholar] [CrossRef]
- Perdikis, D.; Lykouressis, D. Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 2000, 17, 55–60. [Google Scholar] [CrossRef]
- Urbaneja, A.; Tapia, G.; Stansly, P. Influence of host plant and prey availability on developmental time and survivorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Technol. 2005, 15, 513–518. [Google Scholar] [CrossRef]
- Caspi-Fluger, A.; Inbar, M.; Steinberg, S.; Friedmann, Y.; Freund, M.; Mozes-Daube, N.; Zchori-Fein, E. Characterization of the symbiont Rickettsia in the mirid bug Nesidiocoris tenuis (Reuter) (Heteroptera: Miridae). Bull. Entomol. Res. 2014, 104, 681–688. [Google Scholar] [CrossRef] [PubMed]
- MacHtelinckx, T.; Van Leeuwen, T.; Van De Wiele, T.; Boon, N.; De Vos, W.H.; Sanchez, J.A.; Nannini, M.; Gheysen, G.; De Clercq, P. Microbial community of predatory bugs of the genus Macrolophus (Hemiptera: Miridae). BMC Microbiol. 2012, 12, 1–14. [Google Scholar]
- Castañé, C.; Arnó, J.; Gabarra, R.; Alomar, O. Plant damage to vegetable crops by zoophytophagous mirid predators. Biol. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Chen, H.; Rangasamy, M.; Tan, S.Y.; Wang, H.; Siegfried, B.D. Evaluation of five methods for total DNA extraction from western corn rootworm beetles. PLoS ONE 2010, 5, e11963. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved bacterial 16S rRNA gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 2015, 1, e0009-15. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Jiang, P.; Green, S.J.; Chlipala, G.E.; Turek, F.W.; Vitaterna, M.H. Reproducible changes in the gut microbiome suggest a shift in microbial and host metabolism during spaceflight. Microbiome 2019, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Frohlich, D.R.; Torres-Jerez, I.; Bedford, I.D.; Markham, P.G.; Brown, J.K. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V.; Steinberg, S.; Tzuri, G.; Horowitz, A.R.; Belausov, E.; Mozes-Daube, N.; et al. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Environ. Microbiol 2006, 72, 3646–3652. [Google Scholar] [CrossRef] [Green Version]
- Perlman, S.J.; Hunter, M.S.; Zchori-Fein, E. The emerging diversity of Rickettsia. Proc. R. Soc. B Biol. Sci. 2006, 273, 2097–2106. [Google Scholar] [CrossRef] [Green Version]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc. R. Soc. B Biol. Sci. 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [Green Version]
- Chrostek, E.; Pelz-Stelinski, K.; Hurst, G.D.D.; Hughes, G.L. Horizontal transmission of intracellular insect symbionts via plants. Front. Microbiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gehrer, L.; Vorburger, C. Parasitoids as vectors of facultative bacterial endosymbionts in aphids. Biol. Lett. 2012, 8, 613–615. [Google Scholar] [CrossRef]
- Kuechler, S.M.; Dettner, K.; Kehl, S. Characterization of an obligate intracellular bacterium in the midgut epithelium of the bulrush bug Chilacis typhae (Heteroptera, Lygaeidae, Artheneinae). Appl. Environ. Microbiol. 2011, 77, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Brumin, M.; Levy, M.; Ghanim, M. Transovarial transmission of Rickettsia spp. and organ-specific infection of the Whitefly Bemisia tabaci. Appl. Environ. Microbiol. 2012, 78, 5565–5574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardi, J.B.; Miller, L.A.; Bee, C.M. Interfaces between microbes and membranes of host epithelial cells in hemipteran midguts. J. Morphol. 2019, 280, 1046–1060. [Google Scholar] [CrossRef] [PubMed]
- Machtelinckx, T.; Van Leeuwen, T.; Vanholme, B.; Gehesquière, B.; Dermauw, W.; Vandekerkhove, B.; Gheysen, G.; De Clercq, P. Wolbachia induces strong cytoplasmic incompatibility in the predatory bug Macrolophus pygmaeus. Insect Mol. Biol. 2009, 18, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Morel, O.; Noël, V.; Buysse, M.; Binetruy, F.; Lancelot, R.; Loire, E.; Ménard, C.; Bouchez, O.; Vavre, F.; et al. Tick-bacteria mutualism depends on B vitamin synthesis pathways. Curr. Biol. 2018, 28, 1896–1902.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perotti, M.A.; Clarke, H.K.; Turner, B.D.; Braig, H.R. Rickettsia as obligate and mycetomic. FASEB J. 2006, 20, 2372–2374. [Google Scholar] [CrossRef]
- Kölsch, G.; Matz-Grund, C.; Pedersen, B.V. Ultrastructural and molecular characterization of endosymbionts of the reed beetle genus Macroplea (Chrysomelidae, Donaciinae), and proposal of “Candidatus Macropleicola appendiculatae” and ‘‘Candidatus Macropleicola muticae”. Can. J. Microbiol. 2009, 55, 1250–1260. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hosokawa, T.; Nikoh, N.; Meng, X.Y.; Kamagata, Y.; Fukatsu, T. Host-symbiont co-speciation and reductive genome evolution in gut symbiotic bacteria of acanthosomatid stinkbugs. BMC Biol. 2009, 7, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Kaltenpoth, M.; Winter, S.A.; Kleinhammer, A. Localization and transmission route of Coriobacterium glomerans, the endosymbiont of pyrrhocorid bugs. FEMS Microbiol. Ecol. 2009, 69, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Kuechler, S.M.; Renz, P.; Dettner, K.; Kehl, S. Diversity of symbiotic organs and bacterial endosymbionts of lygaeoid bugs of the families Blissidae and Lygaeidae (Hemiptera: Heteroptera: Lygaeoidea). Appl. Environ. Microbiol. 2012, 78, 2648–2659. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.P.; Musgrave, A.J. Ultrastructure of Rickettsia-like microorganisms in the midgut of a plant bug, Stenotus binotatus Jak. (Heteroptera: Miridae). Can. J. Microbiol. 1970, 16, 621–622. [Google Scholar] [CrossRef]
- Fukatsu, T.; Hosokawa, T. Capsule-transmitted gut symbiotic bacterium of the Japanese common plataspid stinkbug, Megacopta punctatissima. Appl. Environ. Microbiol. 2002, 68, 389–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 2007, 5, 48. [Google Scholar]
- Maes, S.; Machtelinckx, T.; Moens, M.; Grégoire, J.C.; De Clercq, P. The influence of acclimation, endosymbionts and diet on the supercooling capacity of the predatory bug Macrolophus pygmaeus. BioControl 2012, 57, 643–651. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Wernegreen, J.J. Bacteriophage flux in endosymbionts (Wolbachia): Infection frequency, lateral transfer, and recombination rates. Mol. Biol. Evol. 2004, 21, 1981–1991. [Google Scholar]
- Vautrin, E.; Vavre, F. Interactions between vertically transmitted symbionts: Cooperation or conflict? Trends Microbiol. 2009, 17, 95–99. [Google Scholar]
- Vautrin, E.; Genieys, S.; Charles, S.; Vavre, F. Do vertically transmitted symbionts co-existing in a single host compete or cooperate? A modelling approach. Evol. Biol. 2008, 21, 145–161. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Gene/Probes | Name | Sequence | Reference |
|---|---|---|---|
| Targeted Genes | |||
| 16S rRNA gene of R. limoniae and R. bellii | Rick-1F | 5′-ATACCGAGTGRGTGAYGAAG-3′ | [21] |
| 16S rRNA gene of R. limoniae | Ricklimoniae-F | 5′-CGGTACCTGACCAAGAAAGC-3′ | [21] |
| 16S rRNA gene of R. bellii | Rickbellii-R | 5′-TCCACGTCGCCGTCTTGC-3′ | [30] |
| 16S rRNA Rickettsia | 1513R | 5′-ACGGYTACCTTGTTACGACTT-3′ | [30] |
| Probes | |||
| Rb1-Cy3 | R. bellii-specific probe | 5′-TCCACGTCGCCGTCTTGC-3′ | [30] |
| Rl1-Cy5 | R. limoniae-specific probe | 5′-GCTTTCTTGGTCAGGTACCG-3′ | [21] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dally, M.; Lalzar, M.; Belausov, E.; Gottlieb, Y.; Coll, M.; Zchori-Fein, E. Cellular Localization of Two Rickettsia Symbionts in the Digestive System and within the Ovaries of the Mirid Bug, Macrolophous pygmaeus. Insects 2020, 11, 530. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080530
Dally M, Lalzar M, Belausov E, Gottlieb Y, Coll M, Zchori-Fein E. Cellular Localization of Two Rickettsia Symbionts in the Digestive System and within the Ovaries of the Mirid Bug, Macrolophous pygmaeus. Insects. 2020; 11(8):530. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080530
Chicago/Turabian StyleDally, Maria, Maya Lalzar, Eduard Belausov, Yuval Gottlieb, Moshe Coll, and Einat Zchori-Fein. 2020. "Cellular Localization of Two Rickettsia Symbionts in the Digestive System and within the Ovaries of the Mirid Bug, Macrolophous pygmaeus" Insects 11, no. 8: 530. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080530