Impact of Horizontal Edge–Interior and Vertical Canopy–Understory Gradients on the Abundance and Diversity of Bark and Woodboring Beetles in Survey Traps

 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Trapping Experiments: General Background
2.2. Experiment 1—Effect of Horizontal and Vertical Gradients on Trap Catches
2.3. Experiment 2—Effect of Horizontal Trap Position on Catches in Canopy Traps
2.4. Data Analysis
3. Results
3.1. General Results
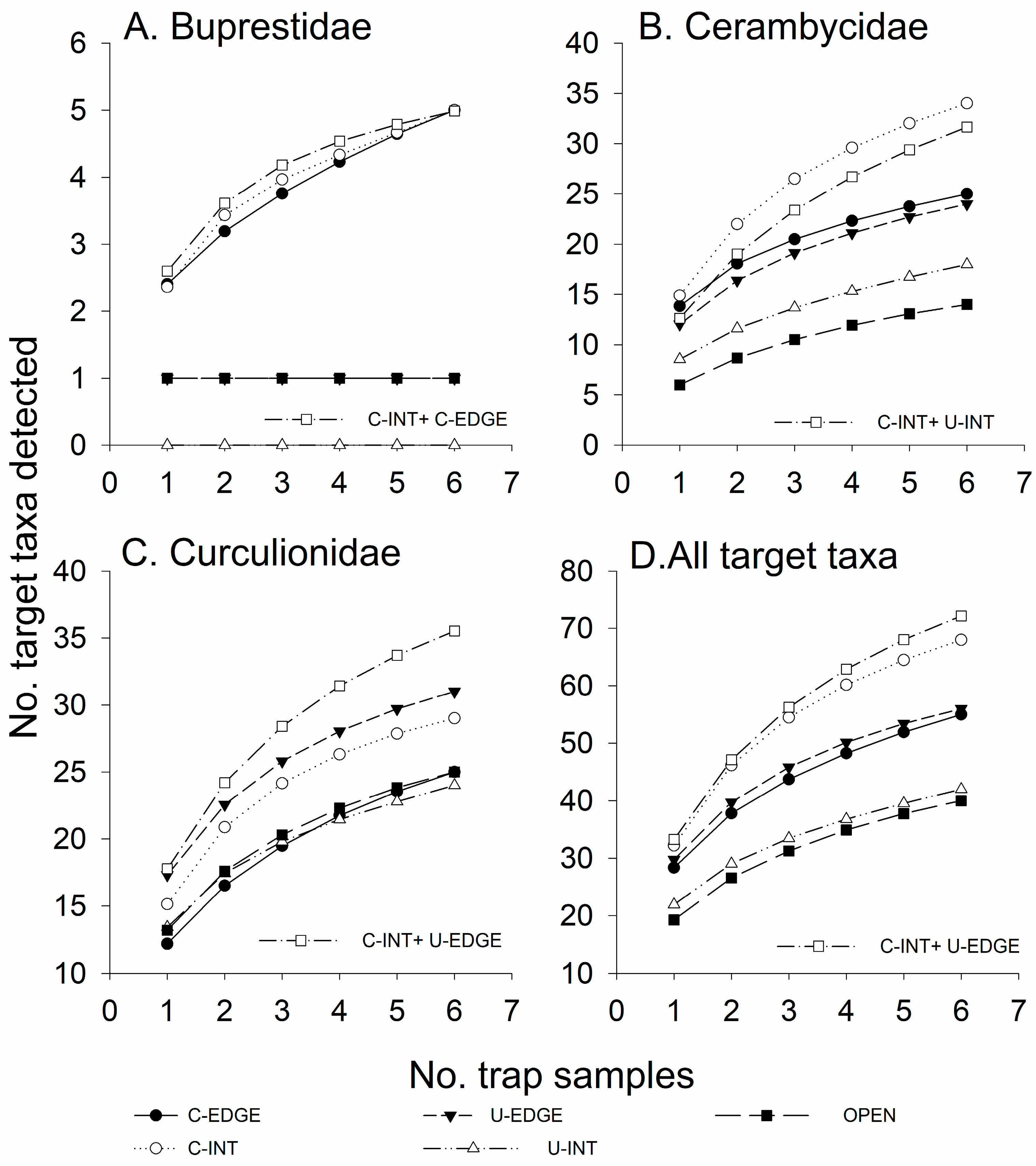
3.2. Experiment 1—Effect of Horizontal and Vertical Gradients on Trap Catches
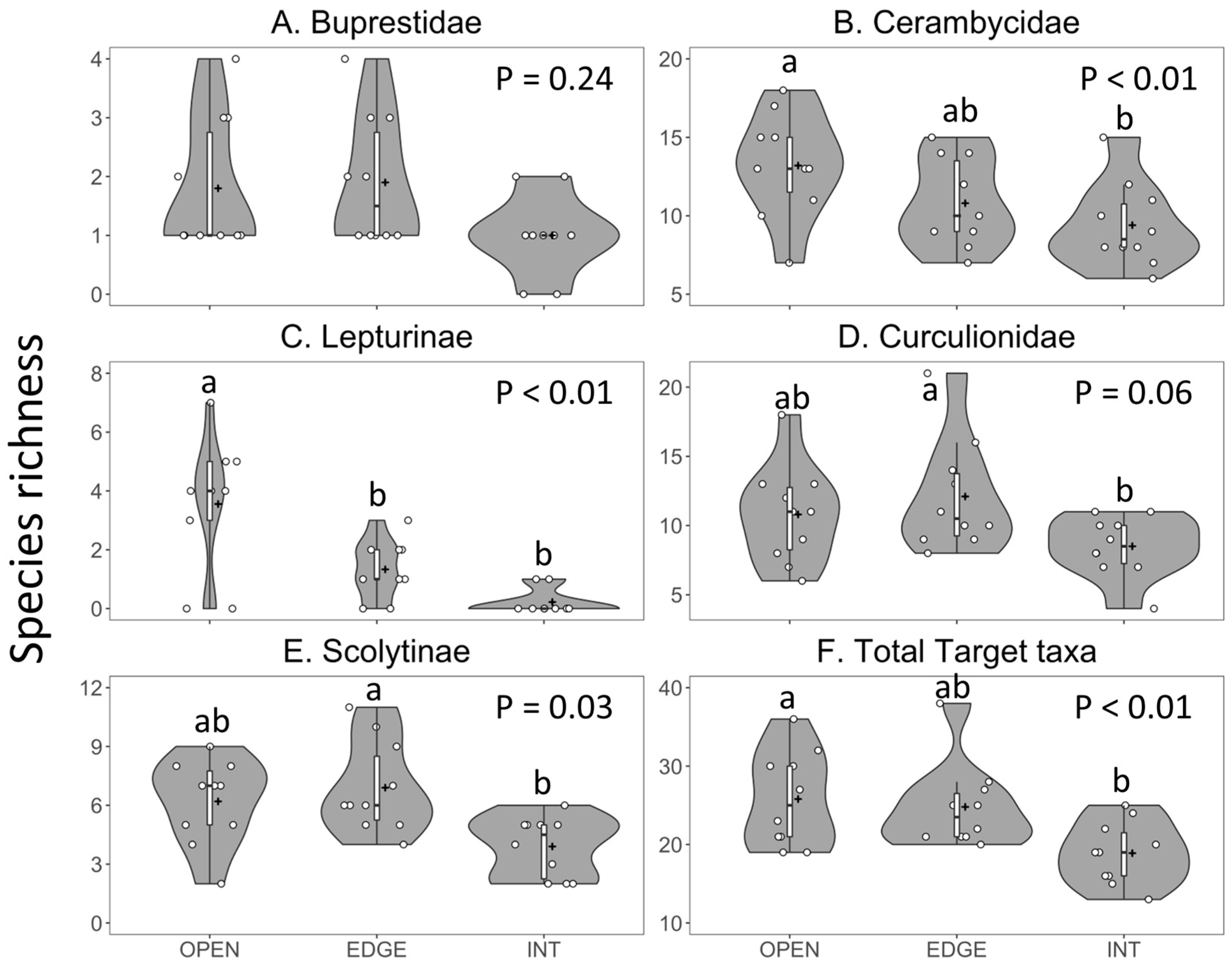
3.2.1. Species Richness
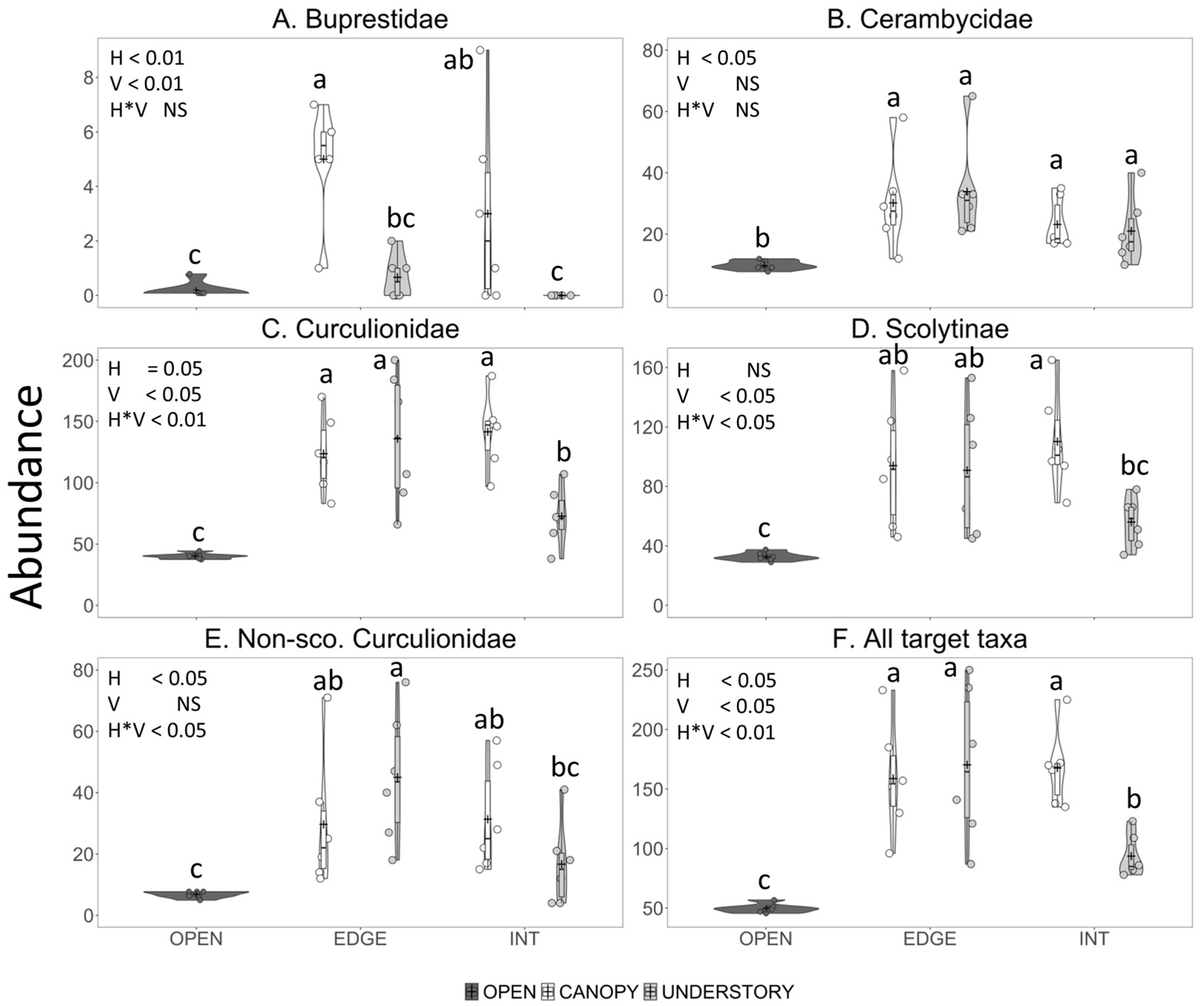
3.2.2. Abundance
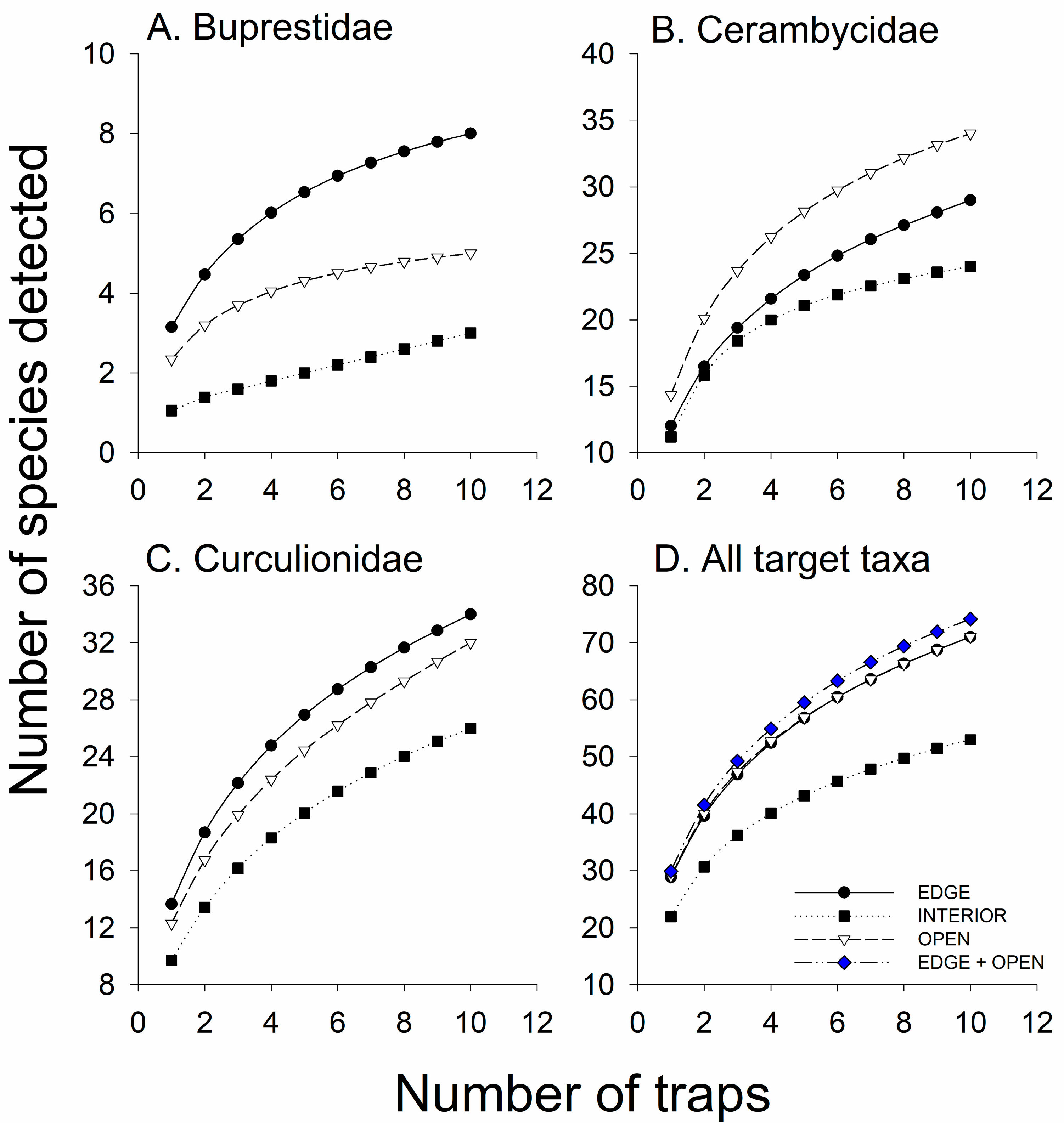
3.3. Experiment 2—Effect of Horizontal Trap Position on Catches in Canopy Traps
3.3.1. Species Richness
3.3.2. Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Binns, M.R.; Nyrop, J.P. Sampling insect populations for the purpose of IPM decision making. Annu. Rev. Entomol. 1992, 37, 427–453. [Google Scholar] [CrossRef]
- Pedigo, L.P. Entomology and Pest Management; Macmillan: New York, NY, USA, 1989. [Google Scholar]
- Evenden, M. Mating disruption of moth pests in integrated pest management: A mechanistic approach. In Pheromone Communication in Moths: Evolution, Behavior and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 365–393. [Google Scholar]
- Cork, A. Pheromones as management tools: Mass trapping and lure-and-kill. In Pheromone Communication in Moths: Evolution, Behavior and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 349–364. [Google Scholar]
- Liebhold, A.M.; Tobin, P.C. Population ecology of insect invasions and their management. Ann. Rev. Entomol. 2008, 53, 387–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.H.; Simberloff, D.; Kuris, A.M.; Carey, J.R. Eradication revisited: Dealing with exotic species. Trends Ecol. Evol. 2000, 15, 316–320. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Allison, J.D.; McKenney, J.L.; Millar, J.G.; McElfresh, J.S.; Mitchell, R.F.; Hanks, L.M. Response of the woodborers Monochamus carolinensis and Monochamus titillator (Coleoptera: Cerambycidae) to known cerambycid pheromones in the presence and absence of the host plant volatile α-pinene. Environ. Entomol. 2012, 41, 1587–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanks, L.M.; Millar, J.G. Sex and aggregation-sex pheromones of cerambycid beetles: Basic science and practical applications. J. Chem. Ecol. 2016, 42, 631–654. [Google Scholar] [CrossRef]
- Miller, D.R.; Crowe, C.M.; Mayo, P.D.; Reid, L.S.; Silk, P.J.; Sweeney, J.D. Interactions between ethanol, syn-2,3-hexanediol, 3-hydroxyhexan-2-one, and 3-hydroxyoctan-2-one lures on trap catches of hardwood longhorn beetles in southeastern United States. J. Econ. Entomol. 2017, 110, 2119–2128. [Google Scholar] [CrossRef]
- Allison, J.D.; Bhandari, B.D.; McKenney, J.L.; Millar, J.G. Design factors that influence the performance of flight intercept traps for the capture of longhorned beetles (Coleoptera: Cerambycidae) from the subfamilies Lamiinae and Cerambycinae. PLoS ONE 2014, 9, e93203. [Google Scholar] [CrossRef] [Green Version]
- Dodds, K.J.; Allison, J.D.; Miller, D.R.; Hanavan, R.P.; Sweeney, J. Considering species richness and rarity when selecting optimal survey traps: Comparisons of semiochemical-baited flight intercept traps for Cerambycidae in eastern North America. Agric. For. Entomol. 2015, 17, 36–47. [Google Scholar] [CrossRef]
- Allison, J.D.; Redak, R.A. The impact of trap type and design features on survey and detection of bark and woodboring beetles and their associates: A review and meta-analysis. Ann. Rev. Entomol. 2017, 62, 127–146. [Google Scholar] [CrossRef]
- Harman, D.M.; Harman, A.L. Distribution pattern of adult locust borers, (Coleoptera: Cerambycidae) on nearby goldenrod, Solidago spp. (Asteraceae), at a forest-field edge. Proc. Entomol. Soc. Wash. 1987, 89, 706–710. [Google Scholar]
- Vance, C.C.; Kirby, K.R.; Malcolm, J.R.; Smith, S.M. Community composition of longhorned beetles (Coleoptera: Cerambycidae) in the canopy and understory of sugar maple and white pine stands in south-central Ontario. Environ. Entomol. 2003, 32, 1066–1074. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L.; Horn, S.; Kilgo, J.C.; Moorman, C.E. Spatial and temporal patterns of beetles associated with coarse woody debris in managed bottomland hardwood forests. For. Ecol. Manag. 2004, 199, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Wermelinger, B.; Flückiger, P.F.; Obrist, M.K.; Duelli, P. Horizontal and vertical distribution of saproxylic beetles (Col., Buprestidae, Cerambycidae, Scolytinae) across sections of forest edges. J. Appl. Entomol. 2007, 131, 104–114. [Google Scholar] [CrossRef]
- Dodds, K.J. Effects of habitat type and trap placement on captures of bark (Coleoptera: Scolytidae) and longhorned (Coleoptera: Cerambycidae) beetles in semiochemical-baited traps. J. Econ. Entomol. 2011, 104, 879–888. [Google Scholar] [CrossRef]
- Dodds, K.J. Effect of trap height on captures of arboreal insects in pine stands of northeastern United States of America. Can. Entomol. 2014, 146, 80–89. [Google Scholar] [CrossRef]
- Graham, E.E.; Poland, T.M.; McCullough, D.G.; Millar, J.G. A comparison of trap type and height for capturing cerambycid beetles (Coleoptera). J. Econ. Entomol. 2012, 105, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Vodka, Š.; Cizek, L. The effects of edge-interior and understory-canopy gradients on the distribution of saproxylic beetles in a temperate lowland forest. For. Ecol. Manag. 2013, 304, 33–41. [Google Scholar] [CrossRef]
- Maguire, D.Y.; Robert, K.; Brochu, K.; Larrivée, M.; Buddle, C.M.; Wheeler, T.A. Vertical stratification of beetles (Coleoptera) and flies (Diptera) in temperate forest canopies. Environ. Entomol. 2014, 43, 9–17. [Google Scholar] [CrossRef]
- Su, J.C.; Woods, S.A. Importance of sampling along a vertical gradient to compare the insect fauna in managed forests. Environ. Entomol. 2014, 30, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Schmeelk, T.C.; Millar, J.G.; Hanks, L.M. Influence of trap height and bait type on abundance and species diversity of cerambycid beetles captured in forest of east-central Illinois. J. Econ. Entomol. 2016, 109, 1750–1757. [Google Scholar] [CrossRef]
- Li, Y.; Meng, Q.; Silk, P.; Gao, W.; Mayo, P.; Sweeney, J. Effect of semiochemicals and trap height on catch of Neocerambyx raddei in Jilin province, China. Entomol. Exp. Appl. 2017, 164, 94–101. [Google Scholar] [CrossRef]
- Flaherty, L.; Gutowski, J.M.G.; Hughes, C.; Mayo, P.; Mokrzycki, T.; Pohl, G.; Silk, P.; Sweeney, J. Pheromone-enhanced lure blends and multiple trap heights improve detection of bark and wood-boring beetles potentially moved in solid wood packaging. J. Pest Sci. 2019, 92, 309–325. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Sheehan, T.N. Trap height considerations for detecting two economically important forest beetle guilds in southeastern US forests. J. Pest Sci. 2019, 92, 253–265. [Google Scholar] [CrossRef]
- Rassati, D.; Marini, L.; Marchioro, M.; Rapuzzi, P.; Magnani, G.; Poloni, R.; Di Giovanni, F.; Mayo, P.; Sweeney, J. Developing trapping protocols for wood-boring beetles associated with broadleaf trees. J. Pest Sci. 2019, 92, 267–279. [Google Scholar] [CrossRef]
- Allison, J.D.; Strom, B.; Sweeney, J.; Mayo, P. Trap deployment along linear transects perpendicular to forest edges: Impact of capture of longhorned beetles (Coleoptera: Cerambycidae). J. Pest Sci. 2019, 92, 299–308. [Google Scholar] [CrossRef]
- Sheehan, T.N.; Ulyshen, M.D.; Horn, S.; Hoebeke, E.R. Vertical and horizontal distribution of bark and woodboring beetles by feeding guild: Is there an optimal trap location for detection? J. Pest Sci. 2019, 92, 327–341. [Google Scholar] [CrossRef]
- Haack, R.A. Exotic bark- and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can. J. For. Res. 2006, 36, 269–288. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Allison, J.D.; Graham, E.E.; Poland, T.M.; Strom, B.L. Dilution of fluon before trap surface treatment has no effect on longhorned beetle (Coleoptera: Cerambycidae) captures. J. Econ. Entomol. 2016, 109, 1215–1219. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.C.; Johns, R.C.; Sweeney, J.D. A technical guide to installing beetle traps in the upper crown of trees. J. Acad. Entomol. Soc. 2014, 10, 12–18. [Google Scholar]
- Yanega, D. Field Guide to Northeastern Longhorned Beetles (Coleoptera: Cerambycidae); Illinois Natural History Survey Manual 6: Champaign, IL, USA, 1996. [Google Scholar]
- Bright, D.E., Jr. The Insects and Arachnids of Canada, Part 2: The Bark Beetles of Canada and Alaska (Coleoptera: Scolytidae); Biosystematics Research Institute, Research Branch, Canada Department of Agriculture: Ottawa, ON, Canada, 1976; Publication No. 1576. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9.1.0 User’s guide and application. Available online: http://viceroy.eeb.uconn.edu/estimates/ (accessed on 24 August 2020).
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef] [Green Version]
- Statistical Tools for High-Throughput Data Analysis, Ggplot2 Violin Plot: Quick Start Guide—R Software and Data Visualization. Available online: http://www.sthda.com/english/wiki/ggplot2-violin-plot-quick-start-guide-r-software-and-data-visualization (accessed on 11 August 2020).
- Webster, R.P.; de Tonnancour, P.; Sweeney, J.D.; Webster, V.L.; Kostanowicz, C.A.; Hughes, C.; Anderson, R.S.; Klymko, J.; Chantal, C.; Vigneault, R. New Coleoptera records from eastern Canada, with additions to the fauna of Manitoba, British Columbia, and Yukon Territory. Zookeys 2020, 946, 53–112. [Google Scholar] [CrossRef] [PubMed]
- McCann, J.M.; Harman, D.M. Influence of the intrastand position of black locust trees on attack rate of the locust borer (Coleoptera: Cerambycidae). Ann. Entomol. Soc. Am. 1990, 83, 705–711. [Google Scholar] [CrossRef]
- Dunn, E.; Hough-Goldstein, J.; Hanks, L.M.; Millar, J.G.; D’Amico, V. Range of attraction of pheromone lures and dispersal behavior of cerambycid beetles. Ann. Entomol. Soc. Am. 2016, 109, 872–880. [Google Scholar] [CrossRef]
- Guimarães, C.; Viana, J.; Cornelissen, T. A meta-analysis of the effects of fragmentation on herbivorous insects. Environ. Entomol. 2014, 43, 537–545. [Google Scholar] [CrossRef]
- Peltonen, M.; Heliövaara, K. Incidence of Xylechinus pilosus and Cryphalus saltuarius (Scolytidae) in forest-clearcut edges. For. Ecol. Manag. 1998, 103, 141–147. [Google Scholar] [CrossRef]
- Flückiger, P.F. Der Bietrag von Waldrandstrukturen zur Regionalen Biodiversität. Ph.D. Thesis, -Natur-wiss. Fakultät Univ., Basel, Switzerland, 1999. [Google Scholar]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L. A comparison of the beetle (Coleoptera) fauna captured at two heights above the ground in a North American temperate deciduous forest. Am. Midl. Nat. 2007, 158, 260–278. [Google Scholar] [CrossRef]
- Hirao, T.; Murakami, M.; Kashizaki, A. Importance of the understory stratum to entomofaunal diversity in temperate deciduous forest. Ecol. Res. 2009, 24, 263–272. [Google Scholar] [CrossRef]
- Schroeder, B.; Buddle, C.M.; Saint-Germain, M. Activity of flying beetles (Coleoptera) at two heights in canopy gaps and intact forests in a hardwood forest in Quebec. Can. Entomol. 2009, 141, 515–520. [Google Scholar] [CrossRef]
- Gossner, M.M. Light intensity affects spatial distribution of Heteroptera in deciduous forests. Eur. J. Entomol. 2009, 106, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Vodka, S.; Konvicka, M.; Cizek, L. Habitat preferences of oak-feeding xylophagous beetles in a temperate woodland: Implications for forest history and management. J. Insect Conserv. 2009, 13, 553–562. [Google Scholar] [CrossRef]
- Bouget, C.; Brin, A.; Brustel, H. Exploring the “last biotic frontier”: Are temperate forest canopies special for saproxylic beetles? For. Ecol. Manag. 2011, 261, 211–220. [Google Scholar] [CrossRef]
- Miller, D.R.; Crowe, C.M.; Sweeney, J.D. Trap height affects catches of bark and woodboring beetles (Coleoptera: Curculionidae, Cerambycidae) in baited multiple-funnel traps in Southeastern United States. J. Econ. Entomol. 2019, 113, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aukema, J.E.; McCullough, D.G.; von Holle, B.; Liebhold, A.M.; Britton, K.; Frankel, S.J. Historical accumulation of nonindigenous forest pests in the continental United States. Bioscience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef]
- Gaston, K.J. Rarity; Chapman Hall: London, UK, 1994. [Google Scholar]
- Williamson, M. Biological Invasions; Chapman Hall: London, UK, 1996. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Semiochemical | Release Device | Purity (%) | Release Rate (mg/d) at 20 °C | Source 3 |
|---|---|---|---|---|
| Racemic 3-hydroxyhexan-2-one | Pouch | 99% 1 | 20–25 | Bedoukian Research Danbury, CT/Contech Enterprises (Scott’s Canada), Delta, BC |
| Racemic 3-hydroxyoctan-2-one | Pouch | 99% 1 | 20–25 | Bedoukian Research/Contech Enterprises (Scott’s Canada) |
| Racemic syn-2,3-hexanediols | Pouch | 99% 1 | 1–2 | Atlantic Forestry Centre, Fredericton, NB/Contech Enterprises (Scott’s Canada) |
| (E/Z)-fuscumol | Rubber septa | 99% 2 | 0.5–2 | Sylvar Technologies, Fredericton, NB |
| (E/Z)-fuscumol acetate | Rubber septa | 99% 2 | 0.5–2 | Sylvar Technologies |
| Ethanol ultra high release rate (UHR) lure | Pouch | 99% 2 | 300–400 | Contech Enterprises (Scott’s Canada) |
| Figure | Variable | Horizontal | Vertical | Hor x Vert | Model | |||
|---|---|---|---|---|---|---|---|---|
| F1, 20 | p | F1, 20 | p | F1, 20 | p | |||
| Buprestidae | Richness | 6.8 | 0.02 | 33.7 | 0.00 | 0.1 | 0.79 | Gaussian, log |
| Abundance | 10.3 | 0.01 | 46.3 | 0.00 | 0.6 | 0.45 | Gaussian, log | |
| Cerambycidae | Richness | 0.4 | 0.53 | 4.5 | 0.05 | 3.1 | 0.09 | Poisson |
| Abundance | 5.1 | 0.04 | 0.0 | 0.99 | 0.4 | 0.53 | Neg. binomial | |
| Lamiinae | Richness | 0.1 | 0.78 | 6.1 | 0.02 | 0.6 | 0.47 | Poisson |
| Abundance | 2.7 | 0.12 | 2.6 | 0.12 | 0.5 | 0.51 | Neg. binomial | |
| Cerambycinae | Richness | 5.3 | 0.03 | 1.2 | 0.28 | 2.5 | 0.13 | Gaussian, log |
| Abundance | 3.9 | 0.06 | 1.5 | 0.24 | 0.1 | 0.81 | Neg. binomial | |
| Lepturinae | Richness | 0.3 | 0.57 | 0.3 | 0.57 | 2.8 | 0.11 | Poisson |
| Abundance | 1.9 | 0.19 | 0.0 | 0.97 | 1.9 | 0.19 | Neg. binomial | |
| Curculionidae | Richness | 2.0 | 0.17 | 1.9 | 0.18 | 23.8 | 0.00 | Gaussian, log |
| Abundance | 4.2 | 0.05 | 5.8 | 0.03 | 10.2 | 0.01 | Neg. binomial | |
| Scolytinae | Richness | 2.0 | 0.17 | 1.3 | 0.26 | 3.7 | 0.07 | Gaussian |
| Abundance | 1.2 | 0.29 | 5.5 | 0.03 | 4.5 | 0.05 | Neg. binomial | |
| Non-Scolytine | Richness | 0.7 | 0.40 | 0.7 | 0.40 | 16.5 | 0.00 | Gaussian |
| Curculionidae | Abundance | 5.2 | 0.03 | 0.1 | 0.81 | 7.2 | 0.01 | Neg. binomial |
| All target taxa | Richness | 2.1 | 0.16 | 2.6 | 0.12 | 10.6 | 0.00 | Poisson |
| Abundance | 6.7 | 0.02 | 5.9 | 0.03 | 9.6 | 0.01 | Neg. binomial | |
| Family | Species | Canopy | Understory | Horizontal | Vertical | Hor*Vert | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Edge | Interior | Edge | Interior | Open | df | F | p | F | p | F | p | ||
| Buprestidae | Dicerca divaricata | 3.17 ± 0.87 a | 0.83 ± 0.48 a | 0.67 ± 0.33 a | 0 ± 0 b | 0.17 ± 0.17 a | 1,20 | 10.90 | <0.01 | 12.70 | <0.01 | 1.05 | 0.32 |
| Cerambycidae | Clytus marginicollis | 1.50 ± 0.56 | 1.33 ± 0.33 | 0.33 ± 0.21 | 0 ± 0 | 1.50 ± 0.62 | 1,20 | 0.64 | 0.43 | 15.90 | <0.01 | 0.07 | 0.79 |
| Cerambycidae | Clytus ruricola | 1.83 ± 0.70 | 1.00 ± 0.52 | 2.50 ± 0.62 | 1.17 ± 0.31 | 0.33 ± 0.21 | 1,20 | 4.01 | 0.06 | 0.46 | 0.50 | 0.05 | 0.82 |
| Cerambycidae | Cyrtophorus verrucosus | 4.33 ± 1.58 a | 1.00 ± 0.37 b | 1.67 ± 0.88 ab | 0.17 ± 0.17 b | 1.67 ± 0.49 ab | 1,20 | 10.88 | <0.01 | 5.78 | 0.03 | 0.54 | 0.47 |
| Cerambycidae | Microclytus compressicollis | 0.67 ± 0.49 ab | 0.83 ± 0.48 ab | 1.33 ± 0.42 ab | 2.67 ± 0.88 a | 0.00 b | 1,20 | 1.32 | 0.26 | 5.41 | 0.03 | 0.35 | 0.56 |
| Cerambycidae | Neoclytus acuminatus | 2.00 ± 0.58 | 1.75 ± 1.44 | 0 ± 0 | 0 ± 0 | 0.25 ± 0.25 | 1,12 | 0.77 | 0.40 | 15.37 | <0.01 | 0.77 | 0.40 |
| Cerambycidae | Phymatodes maculicollis | 3.50 ± 2.74 | 2.67 ± 0.99 | 10.17 ± 1.92 | 7.17 ± 2.80 | 2.83 ± 1.17 | 1,20 | 0.44 | 0.52 | 3.04 | 0.10 | 0.01 | 0.92 |
| Cerambycidae | Aegomorphus modestus | 1.80 ± 0.91 | 1.00 ± 0.55 | 0 ± 0 | 0 ± 0 | 0.6 ± 0.24 | 1,16 | 0.77 | 0.39 | 9.42 | 0.01 | 0.77 | 0.39 |
| Cerambycidae | Astylopsis macula | 3.17 ± 1.11 | 1.50 ± 0.56 | 7.33 ± 4.65 | 1.00 ± 0.52 | 1.17 ± 0.48 | 1,20 | 7.69 | 0.01 | 0.19 | 0.67 | 1.59 | 0.22 |
| Cerambycidae | Monochamus scutellatus | 1.17 ± 0.54 | 0.83 ± 0.40 | 1.50 ± 1.02 | 0 ± 0 | 0 ± 0 | 1,20 | 2.68 | 0.12 | 0.20 | 0.66 | 1.08 | 0.31 |
| Cerambycidae | Pogonocherus pencillatus | 4.00 ± 1.58 a | 0.60 ± 0.24 b | 0.20 ± 0.20 b | 0 ± 0 b | 0 ± 0 b | 1,16 | 13.00 | <0.01 | 28.40 | <0.01 | 7.53 | 0.01 |
| Cerambycidae | Urgleptes signatus | 2.17 ± 1.08 | 2.67 ± 1.02 | 2.17 ± 0.65 | 3.17 ± 0.83 | 0.17 ± 0.17 | 1,20 | 1.28 | 0.27 | 0.11 | 0.74 | 0.11 | 0.74 |
| Cerambycidae | Centrodera decolorata | 1.33 ± 1.15 | 0.83 ± 0.31 | 1.33 ± 0.61 | 1.17 ± 0.60 | 0 ± 0 | 1,20 | 0.31 | 0.59 | 0.10 | 0.76 | 0.10 | 0.76 |
| Cerambycidae | Tetropium cinnamopterum | 0 ± 0 | 0 ± 0 | 0.67 ± 0.33 | 3.67 ± 0.88 | 0.5 ± 0.50 | 1,8 | 19.20 | <0.01 | 40.03 | <0.01 | 19.2 | <0.01 |
| Curculionidae | Stenoscelis brevis | 4.17 ± 1.30 a | 2.00 ± 0.51 a | 4.17 ± 1.45 a | 3.33 ± 1.02 a | 0 ± 0 b | 1,20 | 2.46 | 0.13 | 0.70 | 0.41 | 0.70 | 0.41 |
| Curculionidae | Phyllobius intrusus | 1.60 ± 1.36 | 1.40 ± 0.24 | 0.20 ± 0.20 | 1.4 ± 1.17 | 0.40 ± 0.40 | 1,16 | 2.33 | 0.15 | 3.07 | 0.09 | 3.07 | 0.09 |
| Curculionidae | Phyllobius oblongus | 0.17 ± 0.17 | 0.33 ± 0.21 | 2.33 ± 1.17 | 2.17 ± 0.95 | 0.17 ± 0.17 | 1,20 | 0.16 | 0.69 | 8.52 | 0.01 | 0.25 | 0.62 |
| Curculionidae | Polydrusus formosus | 21.5 ± 9.10 a | 22.83 ± 6.01 a | 19.83 ± 4.60 a | 8.17 ± 5.17 ab | 1.33 ± 0.42 b | 1,20 | 2.42 | 0.14 | 2.38 | 0.14 | 4.16 | 0.05 |
| Curculionidae | Dryophthorus americanus | 0 ± 0 | 0.80 ± 0.20 | 5.80 ± 3.84 | 0.60 ± 0.40 | 1.83 ± 0.31 | 1,16 | 0.91 | 0.36 | 5.64 | 0.03 | 9.95 | 0.01 |
| Curculionidae | Anisandrus obesus | 0.20 ± 0.20 | 0.80 ± 0.58 | 1.80 ± 0.58 | 1.40 ± 0.51 | 1.67 ± 0.33 | 1,16 | 0.86 | 0.37 | 5.05 | 0.04 | 1.78 | 0.20 |
| Curculionidae | Anisandrus sayi | 63.5 ± 11.8 ab | 93.2 ± 11.2 a | 67.0 ± 16.7 ab | 39.5 ± 8.45 b | 10.17 ± 1.33 c | 1,20 | 0.01 | 0.94 | 7.11 | 0.01 | 6.31 | 0.02 |
| Curculionidae | Hylesinis aculeatus | 22.5 ± 9.27 a | 7.50 ± 3.85 ab | 6.50 ± 1.57 ab | 1.17 ± 0.65 b | 10.50 ± 2.57 a | 1,20 | 9.93 | 0.01 | 10.70 | <0.01 | 0.46 | 0.50 |
| Curculionidae | Orthotomicus caelatus | 0.40 ± 0.24 | 0 ± 0 | 1.80 ± 0.58 | 1.00 ± 0.45 | 2.20 ± 1.50 | 1,16 | 2.96 | 0.10 | 11.64 | <0.01 | 0.03 | 0.87 |
| Curculionidae | Phloeosinus canadensis | 1.40 ± 0.68 | 1.20 ± 0.97 | 0.60 ± 0.40 | 0.60 ± 0.40 | 0.40 ± 0.24 | 1,16 | 0.02 | 0.88 | 2.43 | 0.14 | 0.02 | 0.88 |
| Curculionidae | Pityokeines sparsus | 0.60 ± 0.40 | 2.00 ± 0.71 | 1.80 ± 1.36 | 5.20 ± 3.62 | 0.20 ± 0.20 | 1.16 | 2.97 | 0.10 | 1.86 | 0.19 | 0.04 | 0.85 |
| Curculionidae | Polygraphus rufipennis | 2.40 ± 0.68 | 1.80 ± 0.37 | 0 ± 0 | 0 ± 0 | 1.20 ± 0.58 | 1,16 | 0.81 | 0.38 | 39.85 | <0.01 | 0.81 | 0.38 |
| Curculionidae | Xyloterinus politus | 1.67 ± 1.09 bc | 1.00 ± 0.52 c | 8.50 ± 1.52 a | 4.83 ± 1.01 ab | 1.83 ± 0.75 bc | 1,20 | 3.61 | 0.07 | 32.00 | <0.01 | 0.01 | 0.93 |
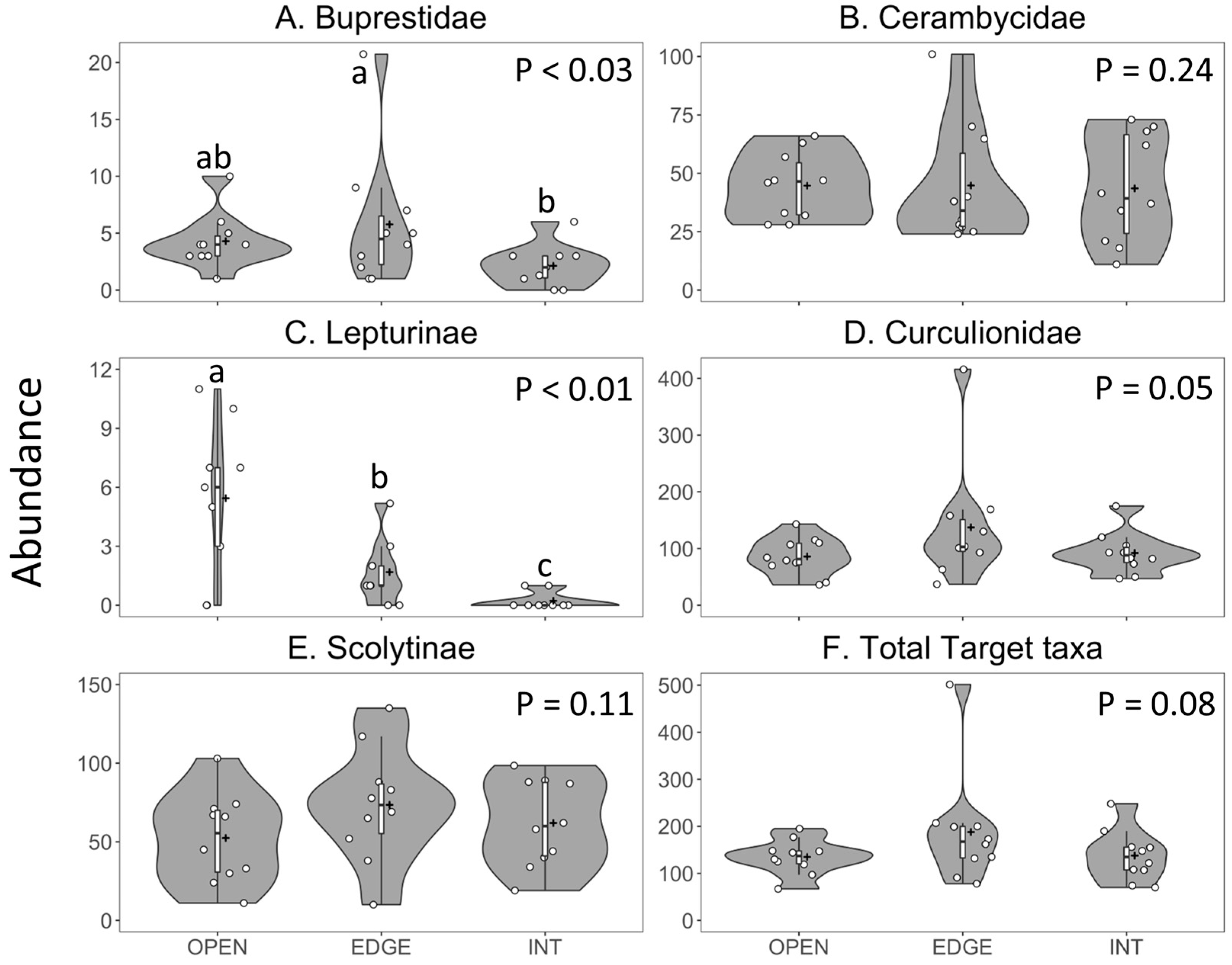
| Family/Subtaxa | Variable | F2, 27 | p | Model |
|---|---|---|---|---|
| Buprestidae | Richness | 1.5 | 0.24 | Poisson |
| Abundance | 4.1 | 0.03 | Negative binomial | |
| Cerambycidae | Richness | 7.1 | <0.01 | Gaussian |
| Abundance | 0.2 | 0.85 | Negative binomial | |
| Lamiinae | Richness | 0.6 | 0.54 | Gaussian log |
| Abundance | 0.3 | 0.76 | Negative binomial | |
| Cerambycinae | Richness | 1.1 | 0.36 | Poisson |
| Abundance | 1.5 | 0.24 | Negative binomial | |
| Lepturinae | Richness | 10.2 | <0.01 | Poisson |
| Abundance | 16.2 | <0.01 | Poisson | |
| Curculionidae | Richness | 3.2 | 0.06 | Poisson |
| Abundance | 3.4 | 0.05 | Negative binomial | |
| Scolytinae | Richness | 4.3 | 0.03 | Poisson |
| Abundance | 2.4 | 0.11 | Negative binomial | |
| Non-Scolytine | Richness | 0.3 | 0.78 | Poisson |
| Curculionidae | Abundance | 2.9 | 0.07 | Negative binomial |
| All target taxa | Richness | 6.0 | <0.01 | Poisson |
| Abundance | 2.8 | 0.08 | Negative binomial |
| Family | Species | Mean Catch per Trap (±SE) | |||||
|---|---|---|---|---|---|---|---|
| Open | Edge | Interior | df | F | p | ||
| Buprestidae | Dicerca divaricata | 2.30 ± 0.45 | 2.70 ± 1.01 | 1.93 ± 0.56 | 2,27 | 0.64 | 0.54 |
| Cerambycidae | Glycobius speciosus | 2.20 ± 0.49 a | 1.30 ± 0.30 ab | 0.73 ± 0.22 b | 2,27 | 3.62 | 0.04 |
| Cerambycidae | Clytus ruricola | 1.33 ± 0.33 | 0.37 ± 0.19 | 0.81 ± 0.28 | 2,24 | 2.28 | 0.12 |
| Cerambycidae | Cyrtophorus verrucosus | 1.40 ± 0.56 | 1.96 ± 0.46 | 1.63 ± 0.42 | 2,27 | 0.47 | 0.63 |
| Cerambycinae | Anelaphus parallelus | 1.00 ± 0.22 | 0.43 ± 0.20 | 1.04 ± 0.54 | 2,18 | 0.94 | 0.41 |
| Cerambycinae | Anelaphus villosus | 0.63 ± 0.38 | 0.79 ± 0.37 | 0.88 ± 0.35 | 2,21 | 0.17 | 0.85 |
| Cerambycinae | Neoclytus a. acuminatus | 1.00 ± 0.27 | 1.36 ± 0.56 | 0.45 ± 0.33 | 2,21 | 1.66 | 0.21 |
| Cerambycinae | Phymatodes maculicollis | 0.67 ± 0.24 | 2.11 ± 0.75 | 1.24 ± 0.56 | 2,24 | 1.97 | 0.24 |
| Cerambycinae | Xylotrechus colonus | 1.67 ± 0.50 | 3.23 ± 1.06 | 0.92 ± 0.57 | 2,24 | 2.50 | 0.10 |
| Cerambycidae | Aegomorphus modestus | 0.60 ± 0.16 | 1.54 ± 0.99 | 0.96 ± 0.27 | 2,27 | 0.96 | 0.40 |
| Cerambycidae | Astylopsis macula | 5.20 ± 0.73 a | 2.62 ± 0.49 b | 2.73 ± 0.82 b | 2,27 | 5.84 | <0.01 |
| Cerambycidae | Sternidius rusticus | 1.75 ± 0.45 | 0.50 ± 0.19 | 1.79 ± 0.96 | 2,21 | 2.80 | 0.08 |
| Cerambycidae | Graphisurus fasciatus | 2.89 ± 0.84 | 2.21 ± 0.52 | 1.07 ± 0.36 | 2,24 | 2.69 | 0.09 |
| Cerambycidae | Urgleptes signatus | 19.8 ± 2.09 | 24.5 ± 7.31 | 27.6 ± 5.81 | 2,27 | 0.68 | 0.51 |
| Cerambycidae | Anthophylax cyaneus | 2.33 ± 0.76 | 0.17 ± 0.17 | 0 ± 0 | 2,15 | 3.25 | 0.07 |
| Curculionidae | Acoptus suturalis | 1.00 ± 0.49 | 2.78 ± 0.74 | 1.60 ± 0.56 | 2,27 | 2.06 | 0.15 |
| Curculionidae | Polydrusus cervinus | 15.7 ± 6.03 | 47.0 ± 27.1 | 13.7 ± 3.67 | 2,27 | 3.26 | 0.05 |
| Curculionidae | Phyllobius oblongus | 5.25 ± 2.72 a | 1.49 ± 0.62 b | 0.75 ± 0.37 b | 2,21 | 10.5 | <0.01 |
| Curculionidae | Polydrusus formosus | 11.4 ± 4.11 | 9.87 ± 3.65 | 11.8 ± 4.73 | 2,27 | 0.20 | 0.82 |
| Curculionidae | Crypturgus borealis | 0.80 ± 0.20 b | 2.19 ± 0.28 a | 0.53 ± 0.23 b | 2,27 | 15.2 | <0.01 |
| Curculionidae | Anisandrus sayi | 39.6 ± 8.2 | 50.7 ± 7.2 | 54.9 ± 8.9 | 2,27 | 1.63 | 0.21 |
| Curculionidae | Pseudopityophthorus minutissimus | 5.10 ± 1.44 b | 14.4 ± 5.53 a | 4.32 ± 1.06 b | 2,27 | 6.83 | <0.01 |
| Curculionidae | Xyleborinus attenuatus | 3.22 ± 0.66 a | 0.92 ± 0.43 b | 0.44 ± 0.24 b | 2,24 | 10.3 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sweeney, J.; Hughes, C.; Webster, V.; Kostanowicz, C.; Webster, R.; Mayo, P.; Allison, J.D. Impact of Horizontal Edge–Interior and Vertical Canopy–Understory Gradients on the Abundance and Diversity of Bark and Woodboring Beetles in Survey Traps. Insects 2020, 11, 573. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11090573
Sweeney J, Hughes C, Webster V, Kostanowicz C, Webster R, Mayo P, Allison JD. Impact of Horizontal Edge–Interior and Vertical Canopy–Understory Gradients on the Abundance and Diversity of Bark and Woodboring Beetles in Survey Traps. Insects. 2020; 11(9):573. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11090573
Chicago/Turabian StyleSweeney, Jon, Cory Hughes, Vincent Webster, Chantelle Kostanowicz, Reginald Webster, Peter Mayo, and Jeremy D. Allison. 2020. "Impact of Horizontal Edge–Interior and Vertical Canopy–Understory Gradients on the Abundance and Diversity of Bark and Woodboring Beetles in Survey Traps" Insects 11, no. 9: 573. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11090573