Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Occurrence Data
2.2. Environmental Variables
2.3. Modeling Procedure
2.4. Model Evaluation
3. Results
3.1. Model Performance
3.2. Effects of Environmental Variables
3.2.1. Contributions of Environmental Variables
3.2.2. Response to the Environmental Variables
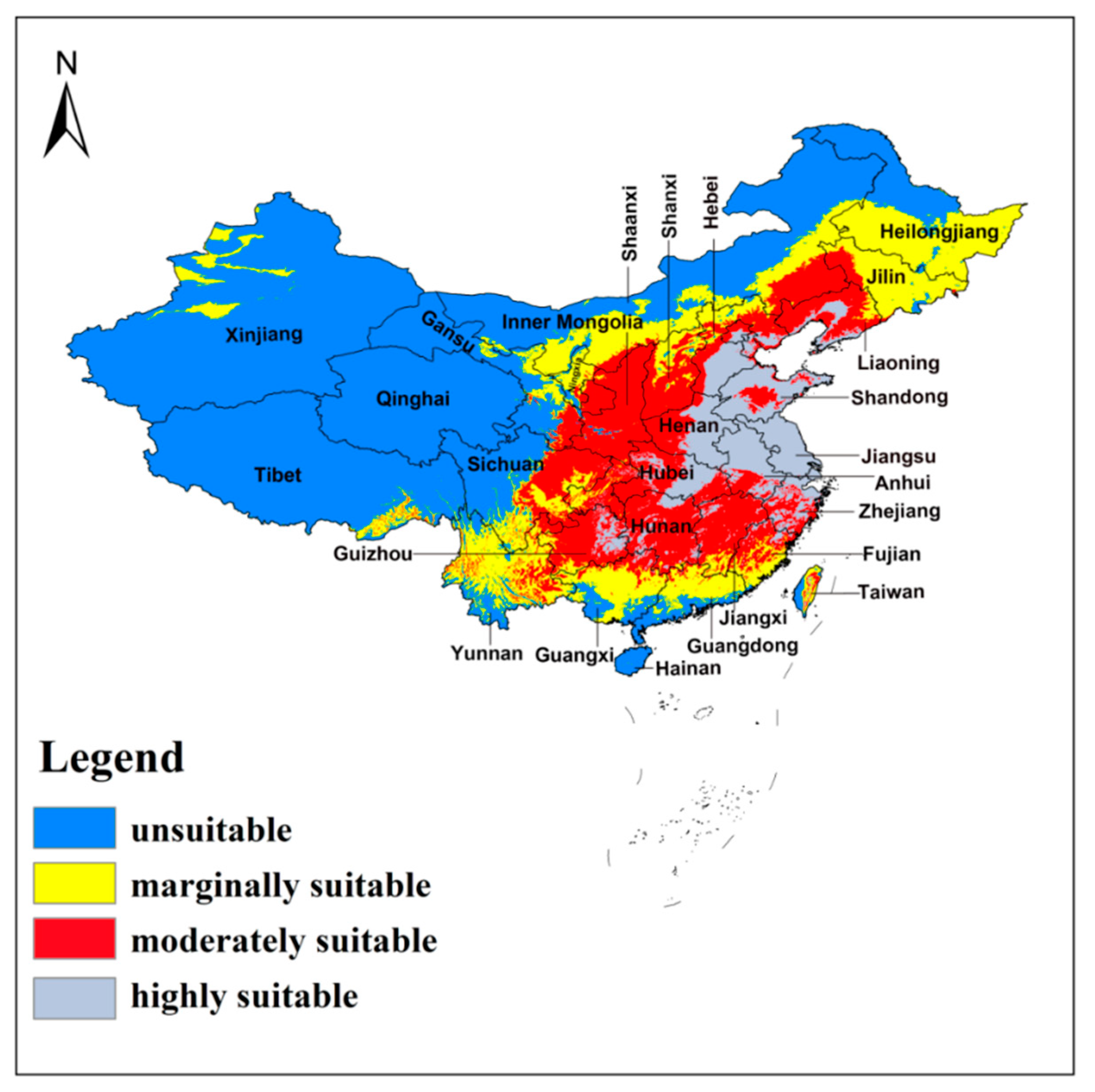
3.3. Current Potential Distribution
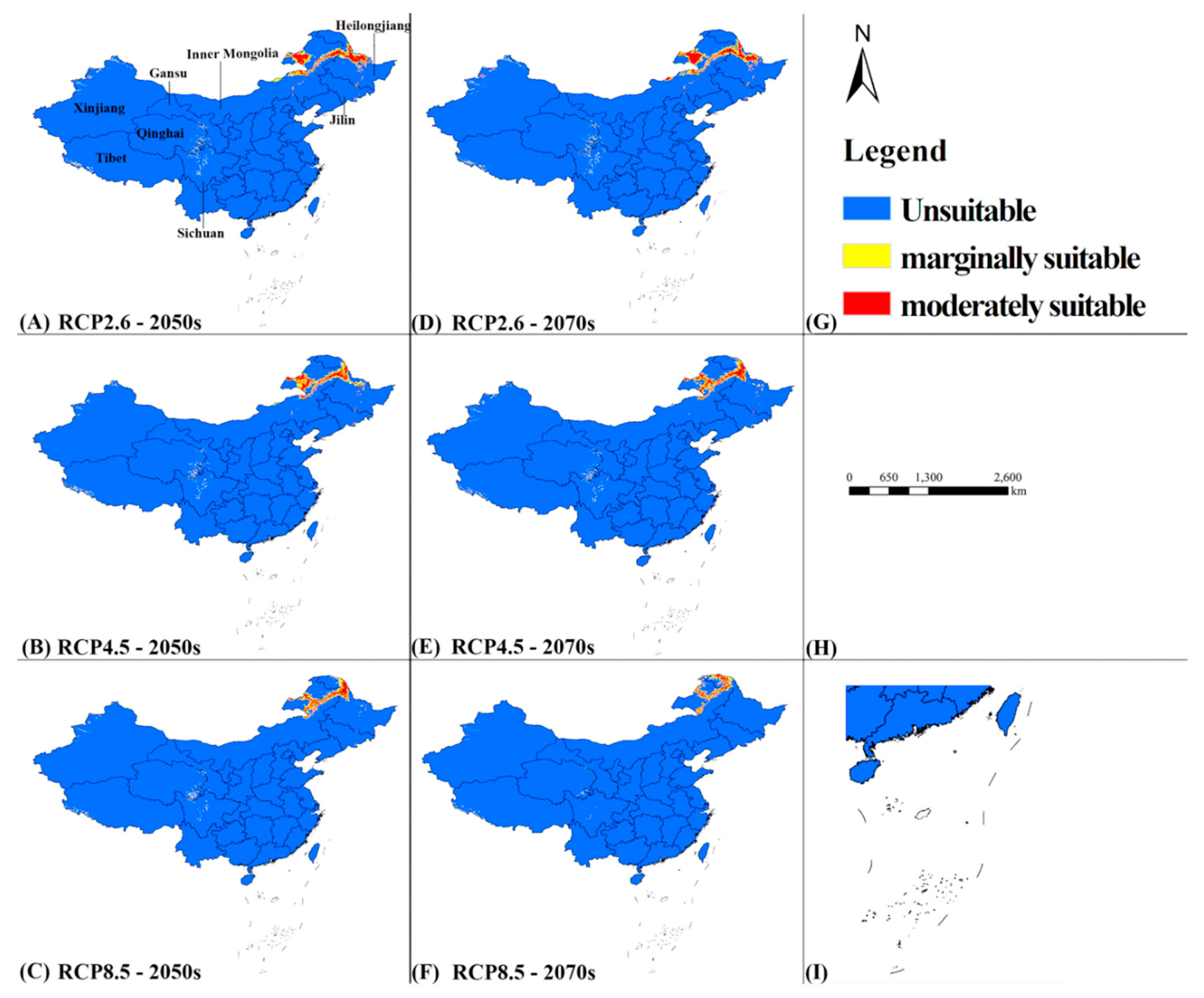
3.4. Future Potential Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zacarias, D.A. Global bioclimatic suitability for the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), and potential co-occurrence with major host crops under climate change scenarios. Clim. Chang. 2020, 1–12. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef] [Green Version]
- Montezano, A.D.G.; Specht, A.; Montezano, D.G.; Specht, A. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas Published by: Entomological Society of Southern Africa Review article Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Firake, D.M.; Behere, G.T. Bioecological attributes and physiological indices of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) infesting ginger (Zingiber officinale Roscoe) plants in India. Crop. Prot. 2020, 137, e105233. [Google Scholar] [CrossRef]
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Mallapur, C.P.; Naik, A.K.; Hagari, S.; Prabhu, S.T. Status of alien pest fall armyworm, Spodoptera frugiperda (J.E. Smith) on maize in Northern Karnataka. J. Entomol. Zool. Stud. 2018, 6, 432–436. [Google Scholar]
- Jiang, Y.Y.; Liu, J.; Zhu, X.M. Analysis on the occurrence dynamics and future trend of the invasion of Spodoptera frugiperda in China. China Plant Prot. 2019, 39, 33–35. [Google Scholar]
- Jing, D.P.; Guo, J.F.; Jiang, Y.Y.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2020, 27, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, K.W.; Lu, Y.Y. The trends and dynamics of the invasion and expansion of Spodoptera frugiperda in China. J. Environ. Entomol. 2019, 41, 683–694. [Google Scholar]
- Koffi, D.; Kyerematen, R.; Eziah, V.Y.; Agboka, K.; Adom, M.; Goergen, G.; Meagher, R.L. Natural Enemies of the Fall Armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in Ghana. Fla. Entomol. 2020, 103, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Da, R.B.; Cruz, I.; Maria, D.L.C. Efficiency of chemical pesticides to control Spodoptera frugiperda and validation of pheromone trap as a pest management tool in maize crop. Rev. Bras. Milho Sorgo 2010, 9, 107–122. [Google Scholar]
- Yu, S.J.; Nguyen, S.N.; Elghar, G.E. Biochemical characteristics of insecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Physiol. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Gouge, D.H.; Piggott, S.J.; Fife, J.P. Application technology and environmental considerations for use of entomopathogenic nematodes in biological control. Biol. Control. 2006, 38, 124–133. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Integrated Management of the Fall Armyworm on Maize. A Guide for Farmer Field Schools in Africa; FAO of the United Nations: Rome, Italy, 2018; pp. 1–139. [Google Scholar]
- Shylesha, A.N.; Jalali, S.K.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K. Studies on new invasive pest Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) and its natural enemies. J. Biol. Control. 2018, 32, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, R.E.; Ruiz, R.A.; Sánchez, J.M.; Molina, J.; Skoda, S.R.; Coutiño, R.; Pinto, R.; Guevara, F.; Foster, J.E. Occurrence of Entomopathogenic Fungi and Parasitic Nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae Collected in Central Chiapas, México. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Jakka, S.R.K.; Knight, V.R.; Jurat, J.L. Fitness Costs Associated With Field-Evolved Resistance to Bt Maize in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen, K.; Gilioli, G.; Gregoire, J.C.; Jaques, J.A.; Navarro, M.N.; et al. Pest categorisation of Spodoptera frugiperda. EFSA J. 2017, 15, 4927. [Google Scholar]
- Leyva, H.A.; Garciá, C.; Ruíz, J.; Calderón, C.L.; Luna, A.; Garciá, S. Evaluation of the Virulence of Steinernema riobrave and Rhabditis blumi against Third Instar Larvae of Spodoptera frugiperda. Southwest. Entomol. 2018, 43, 189–197. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Joseph, E.H. Fall Armyworm in Africa: A Guide for Integrated Pest Management; USAID: Washington, DC, USA, 2018; pp. 1–109.
- Jesus, F.G.; Boiça, A.L.; Alves, G.C.S.; Zanuncio, J.C. Behavior, Development, and Predation of Podisus nigrispinus (Hemiptera: Pentatomidae) on Spodoptera frugiperda (Lepidoptera: Noctuidae) Fed Transgenic and Conventional Cotton Cultivars. Ann. Entomol. Soc. Am. 2014, 107, 601–606. [Google Scholar] [CrossRef]
- Shapiro, J.P.; Legaspi, J.C. Assessing Biochemical Fitness of Predator Podisus maculiventris (Heteroptera: Pentatomidae) in Relation to Food Quality: Effects of Five Species of Prey. Ann. Entomol. Soc. Am. 2006, 99, 321–326. [Google Scholar] [CrossRef]
- Chen, Z.M.; Zhao, L.C.; Liu, H. Parasitic behavior and effect of Microplitis similis on Spodoptera frugiperda larvae. Plant Prot. 2019, 45, 71–74. [Google Scholar]
- Li, F.; Wang, L.K.; Lu, B.Q. The Report of Chelonus munakatae Parasitizing Fall Armworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Hainan, China. Chin. J. Biol. Control. 2019, 35, 992–996. [Google Scholar]
- Xiao, G.R. Forest Insects of China; China Forestry Publishing House: Beijing, China, 1992; pp. 304–306. [Google Scholar]
- Thomas, D. Taxonomic synopsis of the Old World asopine genera (Heteroptera. Pentatomidae). Insecta Mundi 1994, 8, 145–212. [Google Scholar]
- Zou, D.; Wang, M.; Zhang, L.; Zhang, Y.; Zhang, X.; Chen, H. Taxonomic and bionomic notes on Arma chinensis (Fallou) (Hemiptera: Pentatomidae: Asopinae). Zootaxa 2012, 52, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Chai, X.M.; He, Z.H.; Jiang, P. Studies on natural enemies of Dendrolimus punctatus in Zhejiang Province. J. Zhejiang For. Sci. Technol. 2000, 20, 1–56. [Google Scholar]
- Liang, Z.P.; Zhang, X.X.; Song, A.D. Biology of Clostera anachoreta and its control methods. Chin. Bull. Entomol. 2006, 43, 147–152. [Google Scholar]
- Gao, C.Q.; Wang, Z.M.; Yu, E.Y. Studies on artificial rearing of Arma chinensis Fallou. J. Jilin For. Sci. Technol. 1993, 103, 16–18. [Google Scholar]
- Yan, J.H.; Tang, W.Y.; Zhang, H. Bionomics of the leafhopper Macropsis matsudanis. Chin. Bull. Entomol. 2006, 43, 245–249. [Google Scholar]
- Gao, Z. Biological Characteristics and Releasing Techniques of Arma chinensis. Master’s Thesis, Heilongjiang University, Harbin, Heilongjiang, China, 5 July 2010. [Google Scholar]
- Xu, C.H.; Yan, J.J.; Yao, D.F. The relation between the development of Arma chinensis and temperatures. Scientia Silvae Sinicae 1984, 1, 96–99. [Google Scholar]
- Wang, Y.; Zhang, H.M.; Yin, Y.Q. Predation of adult of Arma chinensis to larvae of Spodoptera frugiperda. Plant Prot. 2019, 45, 42–46. [Google Scholar]
- Tang, Y.T.; Li, Y.Y.; Liu, C.X. Predation and Behavior of Arma chinensis to Spodoptera frugiperda. Plant Prot. 2019, 45, 65–68. [Google Scholar]
- Zhang, J.; Zhang, X.; Liu, C.; Meng, L.; Zhou, Y. Fine structure and distribution of antennal sensilla of stink bug Arma chinensis (Heteroptera: Pentatomidae). Entomol. Fenn. 2014, 25, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Zou, D.Y.; Wu, H.H.; Coudron, T.A.; Zhang, L.S.; Wang, M.Q.; Liu, C.X.; Chen, H.Y. A meridic diet for continuous rearing of Arma chinensis (Hemiptera: Pentatomidae: Asopinae). Biol. Control. 2013, 67, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Li, X.P.; Song, L.W.; Coudron, T.A.; Zuo, T.T.; Chen, Y.Q.; Zhang, Y.; Wu, S.A. Effects of two natural diets on the response of the predator Arma chinensis (Hemiptera: Pentatomidae: Asopinae) to cold storage. Appl. Ecol. Environ. Res. 2019, 17, 15329–15347. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, D.; Zhang, W.H. Study on Predatory Function of Arma chinensis on Spodoptera litura (Fabricius). Chin. Tob. Sci. 2019, 40, 55–59. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Kadmon, R.; Farber, O.; Danin, A. Effect of Roadside Bias on the Accuracy of Predictive Maps Produced by Bioclimatic Models. Ecol. Appl. 2004, 14, 401–413. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.F.; Zhao, Q.; Zhao, W.Q.; Zhang, H.F. Predicting the potential distributions of the invasive cycad scale Aulacaspis yasumatsui (Hemiptera: Diaspididae) under different climate change scenarios and the implications for management. PeerJ 2018. [Google Scholar] [CrossRef]
- Wei, J.F.; Peng, L.F.; He, Z.Q.; Lu, Y.; Wang, F. Potential distribution of two invasive pineapple pests under climate change. Pest. Manag. Sci. 2020, 76, 1652–1663. [Google Scholar] [CrossRef]
- Xin, X.; Wu, T.; Li, J. How Well does BCC_CSM1.1 Reproduce the 20th Century Climate Change over China? Atmos. Ocean. Sci. Lett. 2013, 6, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Li, W.; Ji, J.; Xin, X.; Li, L.; Wang, Z.; Zhang, Y.; Li, J.; Zhang, F.; Wei, M.; et al. Global carbon budgets simulated by the Beijing Climate Center Climate System Model for the last century. J. Geophys. Res. Atmos. 2013, 118, 4326–4347. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; ISBN 978-1-107-05799-1. [Google Scholar]
- Van, D.P.; Bouwman, A.F.; Beusen, A.H.W. Phosphorus demand for the 1970–2100 period: A scenario analysis of resource depletion. Glob. Environ. Chang. 2010, 20, 428–439. [Google Scholar] [CrossRef]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 35, 1–20. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, P.R.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, J.R.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Xu, Z.L.; Peng, H.H.; Peng, S.H. The development and evaluation of species distribution models. Shengtai Xuebao/Acta Ecologica Sinica 2015, 35, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.Y.; Li, X.H.; Zou, H.F. Optimizing MaxEnt model in the prediction of species distribution. Chin. J. Appl. Ecol. 2019, 30, 2116–2128. [Google Scholar]
- Bosso, L.; Febbraro, M.D.; Cristinzio, G. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean basin. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Cobos, M.E.; Osorio-Olvera, L.; Peterson, A.T. Assessment and representation of variability in ecological niche model predictions. BioRxiv 2019, 603100. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Ahmadi, M. Assessing accuracy methods of species distribution models: AUC, Specificity, Sensitivity and the True Skill Statistic. Glob. J. Hum. Soc. Sci. 2018, 18, 6–18. [Google Scholar]
- Gilfillan, D.; Joyner, T.A.; Scheuerman, P. Maxent estimation of aquatic Escherichia coli stream impairment. PeerJ 2018, 6, e5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarzo-Arias, A.; Penteriani, V.; Delgado, M.; Torre, P.P.; García-González, R.; Mateo-Sánchez, M.C.; García, P.V.; Dalerum, F. Identifying potential areas of expansion for the endangered brown bear (Ursus arctos) population in the cantabrian mountains (NW Spain). PLoS ONE 2019, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Zhou, S.L.; Zhou, C.Y.; Ding, H.L. Influence of different temperature on the growth and development of Arma chinensis. J. Jilin For. Sci. Technol. 2012, 41, 19–21. [Google Scholar] [CrossRef]
- Liao, P.; Shi, X.R.; Guo, Y.; Yin, Y.F.; Zhu, Y.J. Influence of Low Temperature on Growth and Development of Arma chinensis Fallou (Hemiptera: Pentatomidae). Chin. J. Biol. Control. 2020, 36, 340–346. [Google Scholar] [CrossRef]
- Qin, Y.J.; Lan, S.; Zhao, Z.H.; Sun, H.Y. Potential geographical distribution of the fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2019, 45, 43–47. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M.; et al. Climate Change 2013: The Physical Science Basis; Columbia University Press: New York, NY, USA, 2013; pp. S1–S9. [Google Scholar]
- Xie, D.J.; Tang, J.H.; Zhang, L.; Cheng, Y.X.; Jiang, X.F. Annual Generation Numbers Prediction and Division of Fall Armyworm, Spodoptera frugiperda in China. Available online: http://kns.cnki.net/kcms/detail/11.1982.S.20200604.1800.001.html (accessed on 5 June 2020).
- Soberón, J. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Fern, R.R.; Morrison, M.L.; Wang, H.H.; Grant, W.E.; Campbell, T.A. Incorporating biotic relationships improves species distribution models: Modeling the temporal influence of competition in conspecific nesting birds. Ecol. Model. 2019, 408, 108743. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, P.; Shang, Y. The potential global distribution and dynamics of wheat under multiple climate change scenarios. Sci. Total Environ. 2019, 688, 1308–1318. [Google Scholar] [CrossRef]
- Ghareghan, F.; Ghanbarian, G.; Pourghasemi, H.R.; Safaeian, R. Prediction of habitat suitability of Morina persica L. species using artificial intelligence techniques. Ecol. Indic. 2020, 112, 106096. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Percent Contribution (%) |
|---|---|
| Annual Mean Temperature (bio1) | 58.4 |
| Annual Precipitation (bio12) | 24.8 |
| Precipitation Seasonality (bio15) | 1.9 |
| Precipitation of Wettest Quarter (bio16) | 4 |
| Precipitation of Driest Quarter (bio17) | 8.6 |
| Altitude (alt) | 2.3 |
| Suitability Grade | Current Climate | RCP2.6 2050s | RCP4.5 2050s | RCP8.5 2050s | RCP2.6 2070s | RCP4.5 2070s | RCP8.5 2070s |
|---|---|---|---|---|---|---|---|
| None | ~3.50 × 106 (36.46%) | ~9.27 × 106 (96.66%) | ~9.33 × 106 (97.19%) | ~9.39 × 106 (97.78%) | ~9.24 × 106 (96.23%) | ~9.34 × 106 (97.30%) | 9.43 × 106 (98.21%) |
| Marginal | ~2.50 × 106 (26.04%) | ~1.40 × 105 (1.46%) | ~1.20 × 105 (1.25%) | ~8.84 × 104 (0.92%) | ~1.62 × 105 (1.69%) | ~1.24 × 105 (1.29%) | ~7.73 × 104 (0.81%) |
| Moderate | ~2.50 × 106 (26.04%) | ~1.80 × 105 (1.88%) | ~1.50 × 105 (1.56%) | ~1.25 × 105 (1.30%) | ~1.99 × 105 (2.08%) | ~1.35 × 105 (1.41%) | ~9.41 × 104 (0.98%) |
| High | ~1.10 × 106 (11.46%) | 0 | 0 | 0 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, S.; Chen, C.; Zhao, Q.; Wei, J.; Zhang, H. Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change. Insects 2020, 11, 674. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100674
Fan S, Chen C, Zhao Q, Wei J, Zhang H. Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change. Insects. 2020; 11(10):674. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100674
Chicago/Turabian StyleFan, Shiyu, Chao Chen, Qing Zhao, Jiufeng Wei, and Hufang Zhang. 2020. "Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change" Insects 11, no. 10: 674. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100674