Evaluating Response of Mass-Reared and Irradiated Navel Orangeworm, Amyelois transitella (Lepidoptera: Pyralidae), to Crude Female Pheromone Extract


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Cultures
2.2. Gland Extraction
2.3. Wind Tunnel
2.4. Assay Setup
2.5. Data Analysis
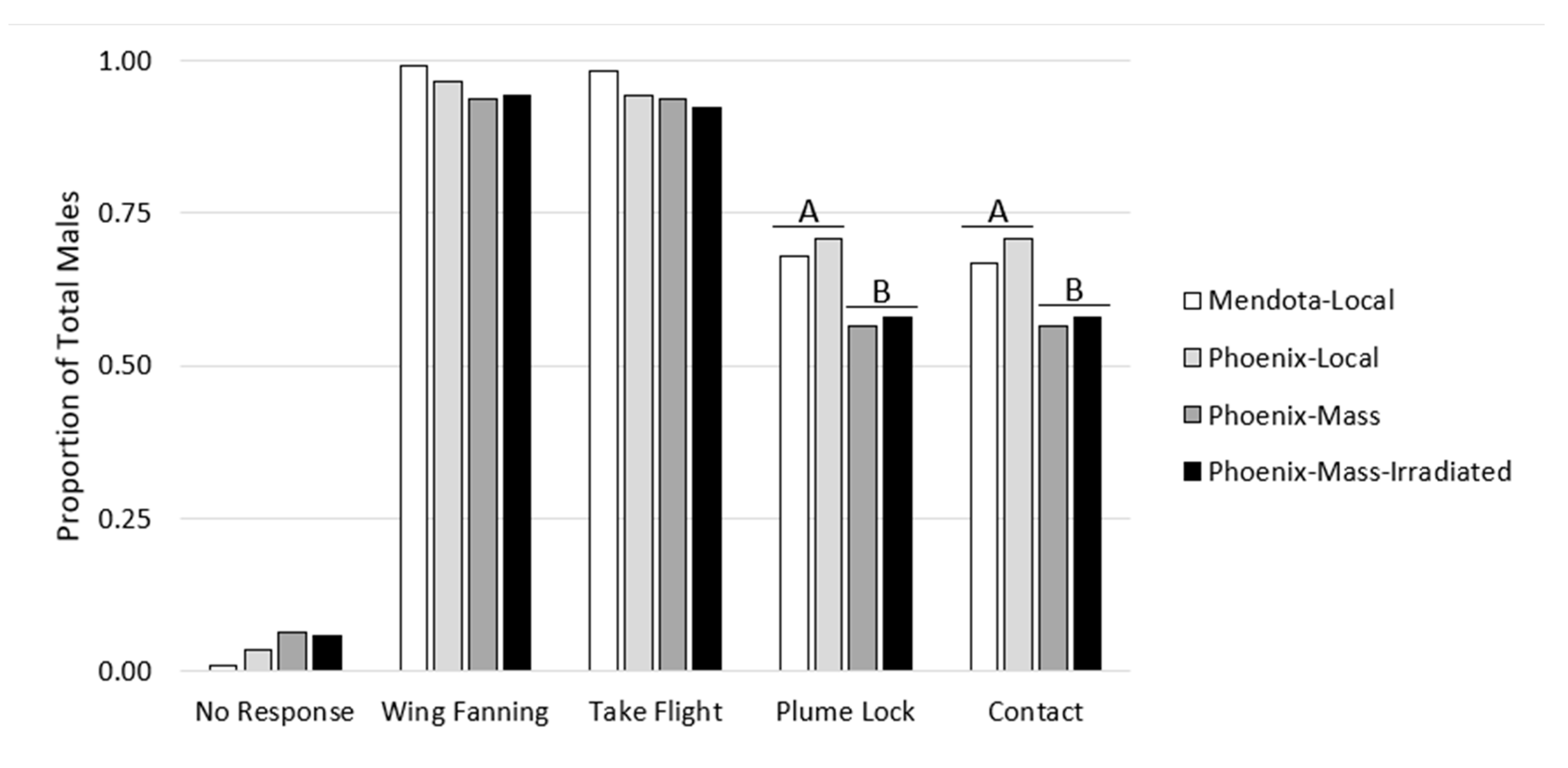
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klassen, W. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique; Springer: Berlin/Heidelberg, Germany, 2005; pp. 39–68. [Google Scholar]
- Knipling, E. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Bloem, K.; Bloem, S.; Carpenter, J. Impact of moth suppression/eradication programmes using the sterile insect technique or inherited sterility. In Sterile Insect Technique; Springer: Berlin/Heidelberg, Germany, 2005; pp. 677–700. [Google Scholar]
- Suckling, D.; Conlong, D.; Carpenter, J.; Bloem, K.; Rendon, P.; Vreysen, M. Global range expansion of pest Lepidoptera requires socially acceptable solutions. Biol. Invasions 2017, 19, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.; Klassen, W.; Carpenter, J.E. Overview of technological advances toward greater efficiency and efficacy in sterile insect-inherited sterility programs against moth pests. Fla. Entomol. 2016, 99, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thistlewood, H.; Judd, G.J. Twenty-five Years of Research Experience with the Sterile Insect Technique and Area-Wide Management of Codling Moth, Cydia pomonella (L.), in Canada. Insects 2019, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, A.; Barrington, A.; Fletcher, N.; Suckling, D. Irradiation conditions affect the quality of irradiated painted apple moth. N. Z. Plant Prot. 2006, 59, 119–124. [Google Scholar] [CrossRef]
- Walters, M.; Staten, R.; Roberson, R. Pink bollworm integrated management using sterile insects under field trial conditions, Imperial Valley, California. In Proceedings of the International Conference on Area-Wide Control of Insect Pests, Imperial Valley, CA, USA, 28 May–2 June 1998 and the Fifth International Symposium on Fruit Flies of Economic Importance, Penang, Malaysia, 1–5 June 1998; pp. 201–206. [Google Scholar]
- Simmons, G.; Suckling, D.; Carpenter, J.; Addison, M.; Dyck, V.; Vreysen, M. Improved quality management to enhance the efficacy of the sterile insect technique for lepidopteran pests. J. Appl. Entomol. 2010, 134, 261–273. [Google Scholar] [CrossRef]
- Doster, M.; Michailides, T. Aspergillus molds and aflatoxins in pistachio nuts in California. Phytopathology 1994, 84, 583–590. [Google Scholar] [CrossRef]
- Palumbo, J.D.; Mahoney, N.E.; Light, D.M.; Siegel, J.; Puckett, R.D.; Michailides, T.J. Spread of Aspergillus flavus by navel orangeworm (Amyelois transitella) on almond. Plant Dis. 2014, 98, 1194–1199. [Google Scholar] [CrossRef] [Green Version]
- Robens, J.; Richard, J. Aflatoxins in animal and human health. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 1992; pp. 69–94. [Google Scholar]
- Haviland, D.; Symmes, E.J.; Adaskaveg, J.E.; Duncan, R.A.; Roncoroni, J.A.; Gubler, W.D.; Hanson, B.; Hembree, K.J.; Holtz, B.A.; Stapleton, J.J.; et al. UC IPM Pest Management Guidelines: Almond. Available online: https://www2.ipm.ucanr.edu/agriculture/almond/Navel-Orangeworm/ (accessed on 15 September 2020).
- Walters, M. Pink bollworm integrated management using sterile insect under field trial conditions. In Programme Book of Abstracts; IAEA-CN-71, 17; Penerbit Universiti Sains Malaysia: Vienna, Austria, 1998. [Google Scholar]
- Wilson, H.W.; University of California, Riverside, Calif., USA; Burks, C.; United States Department of Agriculture-Agricultural Research Service, Parlier, Calif., USA. Personal communication, 2020.
- Tebbets, J.; Curtis, C.; Fries, R. Mortality of immature stages of the navel orangeworm stored at 3.5 C. J. Econ. Entomol. 1978, 71, 875–876. [Google Scholar] [CrossRef]
- Kanno, H.; Kuenen, L.; Klingler, K.A.; Millar, J.G.; Cardé, R.T. Attractiveness of a four-component pheromone blend to male navel orangeworm moths. J. Chem. Ecol. 2010, 36, 584–591. [Google Scholar] [CrossRef] [Green Version]
- Kuenen, L.; Steven McElfresh, J.; Millar, J.G. Identification of critical secondary components of the sex pheromone of the navel orangeworm (Lepidoptera: Pyralidae). J. Econ. Entomol. 2010, 103, 314–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuenen, L.; Rowe, H.; Steffan, K.; Millar, J. Incomplete female sex pheromone of the navel orangeworm Amyelois transitella. Calif. Pist. Comm. Annu. Rep. Crop Year 2001, 2002, 96. [Google Scholar]
- Kuenen, L.; Rowe, H. Cowpea weevil flights to a point source of female sex pheromone: Analyses of flight tracks at three wind speeds. Physiol. Entomol. 2006, 31, 103–109. [Google Scholar] [CrossRef]
- Coffelt, J.; Vick, K.; Sower, L.; McClellan, W. Sex pheromone mediated behavior of the navel orangeworm, Amyelois transitella. Environ. Entomol. 1979, 8, 587–590. [Google Scholar] [CrossRef]
- Parra-Pedrazzoli, A.L.; Leal, W.S. Sexual behavior of the navel orangeworm, Amyelois transitella (Walker) (Lepidoptera: Pyralidae). Neotrop. Entomol. 2006, 35, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Calkins, C.; Parker, A. Sterile insect quality. In Sterile Insect Technique; Springer: Berlin/Heidelberg, Germany, 2005; pp. 269–296. [Google Scholar]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [Green Version]
- Evenden, M. Mating disruption of moth pests in integrated pest management a mechanistic approach. In Pheromone Communication in Moths: Evolution, Behavior, and Application; Allison, J.D., Carde, R.T., Eds.; Univ of California Press: Oakland, CA, USA, 2016; pp. 365–393. [Google Scholar]
- Girling, R.D.; Cardé, R.T. Analysis of the courtship behavior of the navel orangeworm, Amyelois transitella (Walker) (Lepidoptera: Pyralidae), with a commentary on methods for the analysis of sequences of behavioral transitions. J. Insect Behav. 2006, 19, 497. [Google Scholar] [CrossRef]
- Collins, R.; Cardé, R. Wing fanning as a measure of pheromone response in the male pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelechiidae). J. Chem. Ecol. 1989, 15, 2635–2645. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.C.; Cardé, R.T. Analysis of pheromone-mediated behaviors in male Grapholitha molesta, the oriental fruit moth (Lepidoptera: Tortricidae). Environ. Entomol. 1979, 8, 956–968. [Google Scholar] [CrossRef] [Green Version]
- Finney, G.L.; Brinkman, D. Rearing the navel orangeworm in the laboratory. J. Econ. Entomol. 1967, 60, 1109–1111. [Google Scholar] [CrossRef]
- Liu, Y.-B.; Haynes, K.F. Evolution of behavioral responses to sex pheromone in mutant laboratory colonies of Trichoplusia ni. J. Chem. Ecol. 1994, 20, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Evenden, M.; Haynes, K. Potential for the evolution of resistance to pheromone-based mating disruption tested using two pheromone strains of the cabbage looper, Trichoplusia ni. Entomol. Exp. et Appl. 2001, 100, 131–134. [Google Scholar] [CrossRef] [Green Version]
- Boersma, N.; Carpenter, J.E. Influence of holding temperature and irradiation on field performance of mass-reared Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Fla. Entomol. 2016, 99, 215–221. [Google Scholar] [CrossRef]
- Boersma, N.; Boardman, L.; Gilbert, M.; Terblanche, J.S. Sex-dependent thermal history influences cold tolerance, longevity and fecundity in false codling moth Thaumatotibia leucotreta (Lepidoptera: T ortricidae). Agric. For. Entomol. 2018, 20, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Matveev, E.; Kwon, J.; Judd, G.; Evenden, M. The effect of cold storage of mass-reared codling moths (Lepidoptera: Tortricidae) on subsequent flight capacity. Can. Entomol. 2017, 149, 391–398. [Google Scholar] [CrossRef]
- Johnson, J. Survival of Indianmeal moth and navel orangeworm (Lepidoptera: Pyralidae) at low temperatures. J. Econ. Entomol. 2007, 100, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Nepgen, E.; Hill, M.; Moore, S. The effect of long-distance transportation on the fitness of irradiated false codling Moth (Lepidoptera: Tortricidae) for use in a sterile insect release program. J. Econ. Entomol. 2015, 108, 2610–2619. [Google Scholar] [CrossRef] [PubMed]
- Blomefield, T.; Carpenter, J.; Vreysen, M. Quality of mass-reared codling moth (Lepidoptera: Tortricidae) after long-distance transportation: 1. Logistics of shipping procedures and quality parameters as measured in the laboratory. J. Econ. Entomol. 2011, 104, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Husseiny, M.; Madsen, H. Sterilization of the navel orangeworm, Paramyelois transitella (Walker), by gamma radiation (Lepidoptera: Phycitidae). Hilgardia 1964, 36, 113–137. [Google Scholar] [CrossRef] [Green Version]
- Light, D.M.; Ovchinnikova, I.; Jackson, E.S.; Haff, R.P. Effects of x-ray irradiation on male navel orangeworm moths (Lepidoptera: Pyralidae) on mating, fecundity, fertility, and inherited sterility. J. Econ. Entomol. 2015, 108, 2200–2212. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Fixed Effect | Behavioral Response | ||||
|---|---|---|---|---|---|
| No Response | Wing Fanning | Take Flight | Plume Lock | Contact | |
| Strain | 1.22 | 1.22 | 1.59 | 1.19 | 1.19 |
| Rearing | 1.12 | 1.12 | 0.36 | 4.09 * | 4.09 * |
| Irradiation | 0.09 | 0.09 | 0.00 | 0.04 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reger, J.; Wenger, J.; Brar, G.; Burks, C.; Wilson, H. Evaluating Response of Mass-Reared and Irradiated Navel Orangeworm, Amyelois transitella (Lepidoptera: Pyralidae), to Crude Female Pheromone Extract. Insects 2020, 11, 703. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100703
Reger J, Wenger J, Brar G, Burks C, Wilson H. Evaluating Response of Mass-Reared and Irradiated Navel Orangeworm, Amyelois transitella (Lepidoptera: Pyralidae), to Crude Female Pheromone Extract. Insects. 2020; 11(10):703. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100703
Chicago/Turabian StyleReger, Joshua, Jacob Wenger, Gurreet Brar, Charles Burks, and Houston Wilson. 2020. "Evaluating Response of Mass-Reared and Irradiated Navel Orangeworm, Amyelois transitella (Lepidoptera: Pyralidae), to Crude Female Pheromone Extract" Insects 11, no. 10: 703. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100703