Effect of Collection Month, Visible Light, and Air Movement on the Attraction of Male Agriotes obscurus L. (Coleoptera: Elateridae) Click Beetles to Female Sex Pheromone

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Beetle Collection and Pheromone Formulation
2.2. Experiment 1: Air Movement
2.3. Experiment 2: Visible Light
2.4. Experiment 3: Month of Beetle Collection
2.5. Experiment 4: Beetle Response Range
3. Results
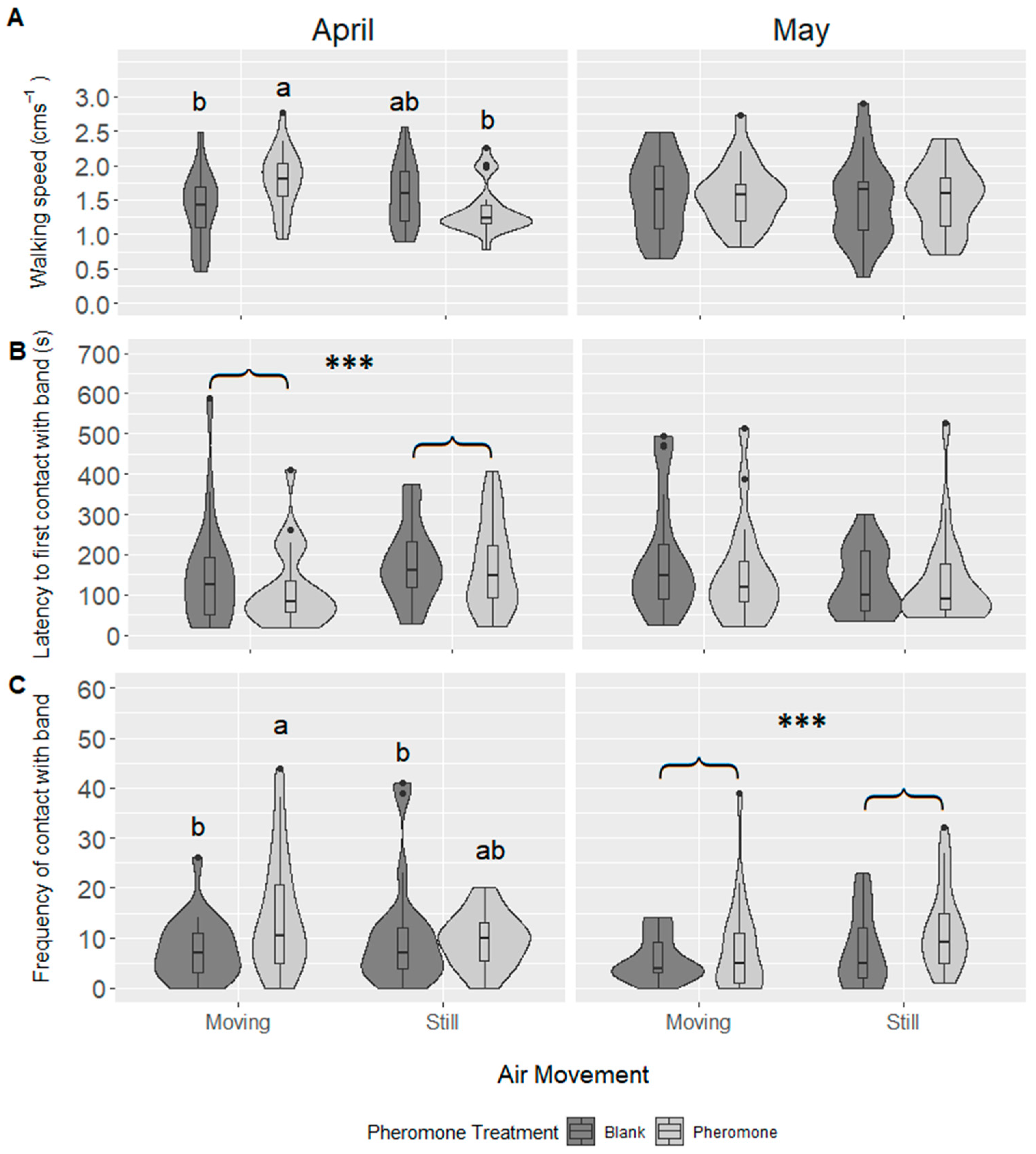
3.1. Experiment 1: Air Movement
3.2. Experiment 2: Visible Light
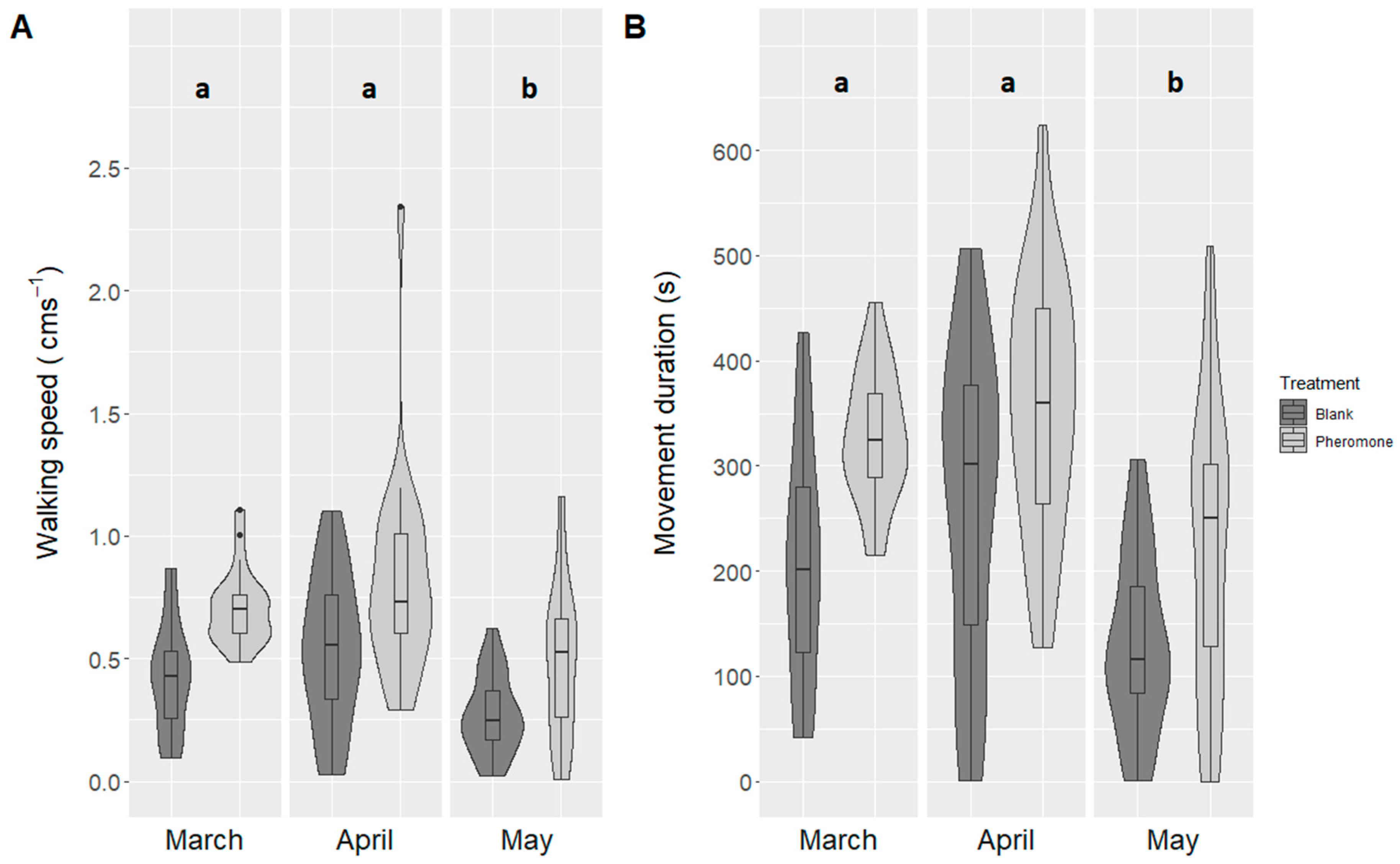
3.3. Experiment 3: Month of Beetle Collection
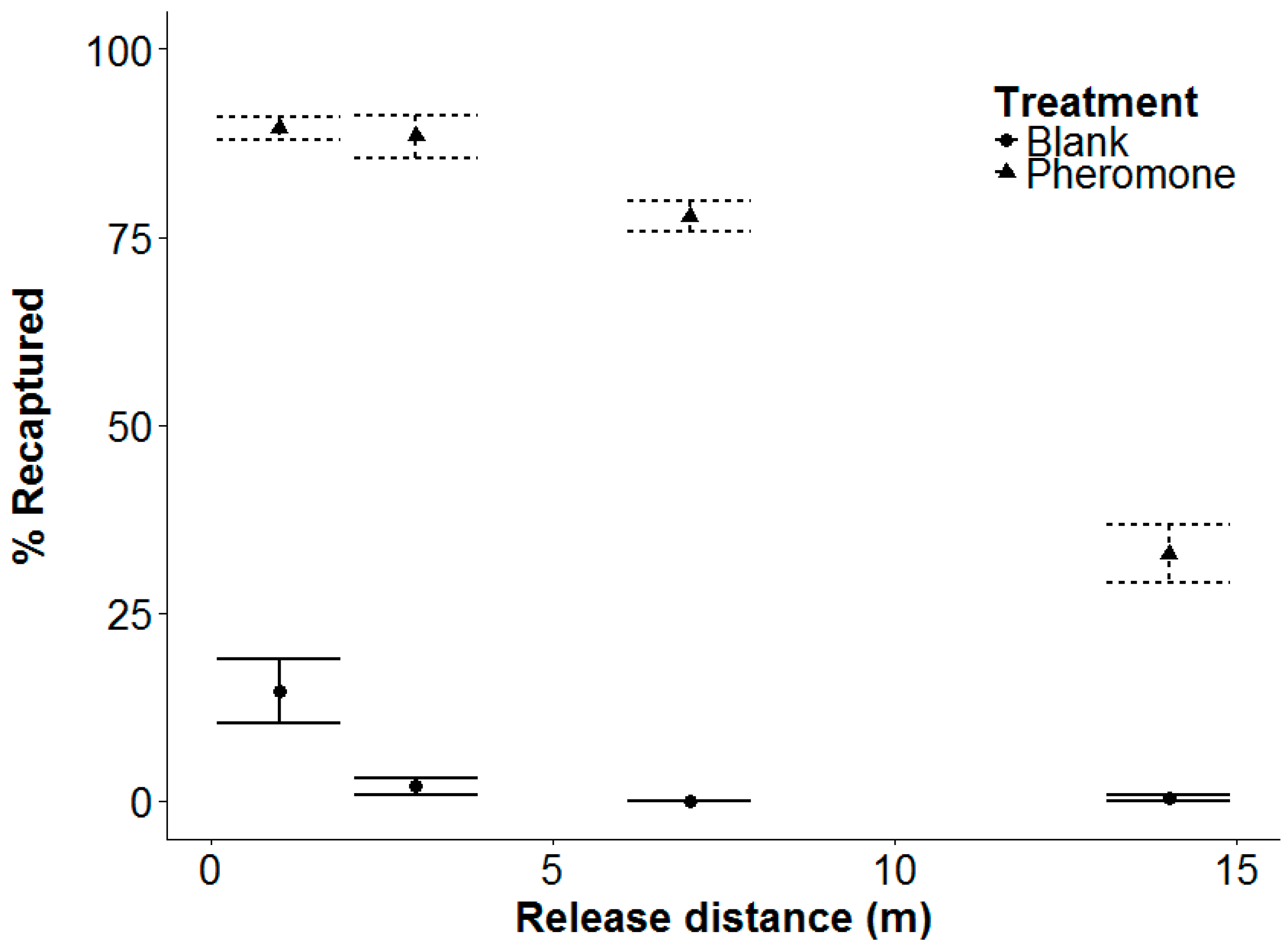
3.4. Experiment 4: Beetle Response Range
4. Discussion
4.1. Male Click Beetles Respond to Pheromone under Both Moving and Still Air Conditions
4.2. Visible Light Shows No Impact on Pheromone Response
4.3. Male Click Beetles More Active in March and April
4.4. Beetle Response Range and Recapture Rates
4.5. Implications for Pheromone-Based Management Strategies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eidt, D.C. European Wireworms 1 in Canada with Particular Reference to Nova Scotian Infestations 2. Can. Entomol. 1953, 85, 408–414. [Google Scholar] [CrossRef]
- Iwanaga, S.; Kawamura, F. Trapping efficacy of funnel-vane and water pan traps baited with synthetic sex pheromone of the sugarcane wireworms, Melanotus sakishimensis Ohira and M. okinawensis Ohira (Coleoptera: Elateridae). Appl. Entomol. Zool. 2000, 35, 283–285. [Google Scholar] [CrossRef] [Green Version]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Larval development of Agriotes obscurus under laboratory and semi-natural conditions. Bull. Insectol. 2014, 67, 227–235. [Google Scholar]
- Miles, H.W. Wireworms and agriculture, with special reference to Agriotes obscurus L. Ann. Appl. Biol. 1942, 29, 176–180. [Google Scholar] [CrossRef]
- Doane, J.F.; Lee, Y.W.; Klingler, J.; Westcott, N.D. Orientation response of Ctenicera destructor and other wireworms (Coleoptera: Elateridae) to germinating grain and carbon dioxide. Can. Entomol. 1975, 107, 1233–1252. [Google Scholar] [CrossRef]
- Humbert, P.; Vemmer, M.; Mavers, F.; Schumann, M.; Vidal, S.; Patel, A.V. Development of an attract-and-kill co-formulation containing Saccharomyces cerevisae and neem extract attractive towards wireworms. Pest Manag. Sci. 2018, 74, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Janson, R.K.; Seal, D.R. Biology and management of wireworm on potato. In Advances in Potato Pest Biology; Zehnder, G.W., Powelson, M.L., Janson, R.K., Raman, K.V., Eds.; APS Press: St. Paul, MN, USA; pp. 31–53.
- Barsics, F.; Haubruge, E.; Verheggen, F. Wireworms’ Management: An Overview of the Existing Methods, with Particular Regards to Agriotes spp. (Coleoptera: Elateridae). Insects 2013, 4, 117–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, W.E.; Howard, J.J. The biology and management of wireworms (Agriotes spp.) on potato with particular reference to the UK. Agric. For. Entomol. 2001, 3, 85–98. [Google Scholar] [CrossRef]
- Crozier, S.; Tanaka, A.; Vernon, R.S. Flight activity of Agriotes lineatus L. and A. obscurus L. (Coleoptera: Elateridae) in the field. J. Entomol. Soc. Br. Columbia 2003, 100, 91–92. [Google Scholar]
- Vernon, R.; van Herk, W.G.; Clodius, M.; Harding, C. Wireworm Management I: Stand Protection versus Wireworm Mortality with Wheat Seed Treatments. J. Econ. Entomol. 2009, 102, 212–213. [Google Scholar] [CrossRef]
- Vernon, B.O.B.; Lagasa, E.; Philip, H. Geographic and temporal distribution of Agriotes obscurus and A. lineatus (Coleoptera:Elateridae) in British Columbia and Washington as determined by pheromone trap surveys. J. Entomol. Soc. Br. Columbia 2001, 98, 257–265. [Google Scholar]
- Cohen, M. Observations on the biology of Agriotes obscurus L.: The Adult Insect. Ann. Appl. Biol. 1942, 29, 181–196. [Google Scholar] [CrossRef]
- Vernon, R.S.; Van Herk, W.G. Wireworms as pests of potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Giordanengo, P., Vincent, C., Alyokhin, A., Eds.; Academic Press: Amsterdam, The Netherlands, 2013; pp. 103–164. [Google Scholar]
- Traugott, M.; Benefer, C.M.; Blackshaw, R.P.; van Herk, W.G.; Vernon, R.S. Biology, ecology, and control of elaterid beetles in agricultural land. Annu. Rev. Entomol. 2015, 60, 313–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schallhart, N.; Wallinger, C.; Juen, A.; Traugott, M. Dispersal abilities of adult click beetles in arable land revealed by analysis of carbon stable isotopes. Agric. For. Entomol. 2009, 11, 333–339. [Google Scholar] [CrossRef]
- Fryer, J.C.F. Time of flight in beetles of the genus Agriotes. Entomol. Mon. Mag. 1941, 77, 280. [Google Scholar]
- Subklew, W. Agriotes lineatus L. und Agriotes obscurus L. J. Appl. Entomol. 1935, 21, 96–122. [Google Scholar] [CrossRef]
- Wilkinson, A.T.S. Controlling the European Wireworm Agriotes obscurus L. in corn in British Columbia. J. Entomol. Soc. Br. Columbia 1976, 73, 3–5. [Google Scholar]
- Tóth, M.; Furlan, L.; Yatsynin, V.G.; Ujváry, I.; Szarukán, I.; Imrei, Z.; Tolasch, T.; Francke, W.; Jossi, W. Identification of pheromones and optimization of bait composition for click beetle pests in central and Western Europe (Coleoptera: Elateridae). Pest Manag. Sci. 2003, 59, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Vernon, R.S.; Tóth, M. Evaluation of pheromones and a new trap for monitoring Agriotes lineatus and Agriotes obscurus in the Fraser Valley of British Columbia. J. Chem. Ecol. 2007, 33, 345–351. [Google Scholar] [CrossRef]
- Arakaki, N.; Nagayama, A.; Kobayashi, A.; Kishita, M.; Sadoyama, Y.; Mougi, N.; Kawamura, F.; Wakamura, S.; Yamamura, K. Control of the sugarcane click beetle Melanotus okinawensis Ohira (Coleoptera: Elateridae) by mass trapping using synthetic sex pheromone on Ikei Island, Okinawa, Japan. Appl. Entomol. Zool. 2008, 43, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara-Tsujii, N.; Yasui, H.; Arakaki, N. Chemical and Physical Cues Synergistically Affect Mating Behavior Sequences of Male Dasylepida ishigakiensis (Coleoptera: Scarabaeidae). Zool. Sci. 2014, 31, 553–558. [Google Scholar] [CrossRef]
- Kabaluk, T.J.; Lafontaine, J.P.; Borden, J.H. An attract and kill tactic for click beetles based on Metarhizium brunneum and a new formulation of sex pheromone. J. Pest Sci. 2015, 88, 707–716. [Google Scholar] [CrossRef]
- McNeil, J.N. Behavioural Ecology of Pheromone-Mediated Communication in Moths and Its Importance in the Use of Pheromone Traps. Annu. Rev. Entomol. 1991, 36, 407–430. [Google Scholar] [CrossRef]
- Bell, W.J. Searching Behaviour: The Behavioural Ecology of Finding Resources; Chapman and Hall: Cambridge, UK, 1991. [Google Scholar]
- Riedl, H.; Howell, I.F.; McNally, P.S.; Westigard, P.H. Codling moth management: Use and standardization of pheromone trapping systems. In Bulletin 1918; University of California Division of Agriculture and Natural Resources: Oakland, CA, USA, 1986. [Google Scholar]
- Sappington, T.W.; Spurgeon, D.W. Variation in boll weevil (Coleoptera: Curculionidae) captures in pheromone traps arising from wind speed moderation by brush lines. Environ. Entomol. 2000, 29, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Rosén, W.Q.; Han, G.-B.; Löfstedt, C. The circadian rhythm of the sex-pheromone-mediated behavioral response in the turnip moth, Agrotis segetum, is not controlled at the peripheral level. J. Biol. Rhythm. 2003, 18, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landolt, P.J.; Curtis, C.E. Effects of temperature on the circadian rhythm of navel orangeworm sexual activity. Environ. Entomol. 1982, 11, 107–110. [Google Scholar] [CrossRef]
- Linn, C.E., Jr.; Roelofs, W.L. Modulatory effects of octopamine and serotonin on male sensitivity and periodicity of response to sex pheromone in the cabbage looper moth Trichoplusia ni. Arch. Insect Biochem. Physiol. 1986, 3, 161–171. [Google Scholar] [CrossRef]
- Hicks, H.; Blackshaw, R.P. Differential responses of three Agriotes click beetle species to pheromone traps. Agric. For. Entomol. 2008, 10, 443–448. [Google Scholar] [CrossRef]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Assessment of the range of attraction of pheromone traps to Agriotes lineatus and Agriotes obscurus. Agric. For. Entomol. 2011, 13, 313–319. [Google Scholar] [CrossRef]
- Van Herk, W.G.; Kabaluk, J.T.; Lam, V.W.; Vernon, R.S. Survival of male click beetles, Agriotes obscurus L., (Coleoptera: Elateridae) during and after storage at different temperatures. J. Entomol. Soc. Br. Columbia 2008, 105, 113–114. [Google Scholar]
- LeNaour, E.; Kabaluk, T. A comprehensive analysis of wind speed and direction during the spring months 2010–2013 at the Pacific Agri-Food Research Centre, Agassiz, British Columbia. In Internal Report; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2014. [Google Scholar]
- Gibson, G. A behavioural test of the sensitivity of a nocturnal mosquito, Anopheles gambiae, to dim white, red and infra-red light. Physiol. Entomol. 1995, 20, 224–228. [Google Scholar] [CrossRef]
- Allema, A.B.; Rossing, W.A.H.; Van der Werf, W.; Heusinkveld, B.G.; Bukovinszky, T.; Steingröver, E.; Van Lenteren, J.C. Effect of light quality on movement of Pterostichus melanarius (Coleoptera: Carabidae). J. Appl. Entomol. 2012, 136, 793–800. [Google Scholar] [CrossRef]
- Leung, J.P.S. The Response of Agriotes Obscurus Click Beetles to Pheromone and Its Impact on Acquisition of a Fungal Pathogen. Master’s Thesis, Simon Fraser University, Burnaby, BC, Canada, 2018. [Google Scholar]
- Kramer, E. Orientation of the male silkmoth to the sex attractant bombykol. In Mechanisms in Insect Olfaction; Denton, D.A., Coghlan, J.P., Eds.; Academic Press: New York, NY, USA, 1975; pp. 329–335. [Google Scholar]
- Willis, M.A.; Avondet, J.L.; Finnell, A.S. Effects of altering flow and odor information on plume tracking behavior in walking cockroaches, Periplaneta americana (L.). J. Exp. Biol. 2008, 211, 2317–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, T.R.; Bell, W.J. Chemo-orientation of male Trogoderma variabile (Coleoptera, Dermestidae) in a simulated corridor of female sex pheromone. J. Comp. Physiol. 1986, 158, 729–739. [Google Scholar] [CrossRef]
- Blackshaw, R.P.; van Herk, W.G.; Vernon, R.S. Determination of Agriotes obscurus (Coleoptera: Elateridae) sex pheromone attraction range using target male behavioural responses. Agric. For. Entomol. 2017, 20, 228–233. [Google Scholar] [CrossRef]
- Cardé, R.T.; Willis, M.A. Navigational strategies used by insects to find distant, wind-borne sources of odor. J. Chem. Ecol. 2008, 34, 854–866. [Google Scholar] [CrossRef]
- Brian, M.V. On the ecology of beetles of the genus Agriotes with special reference to A. obscurus. J. Anim. Ecol. 1947, 16, 210–224. [Google Scholar] [CrossRef]
- Ridgel, A.L.; Ritzmann, R.E.; Schaefer, P.L. Effects of aging on behavior and leg kinematics during locomotion in two species of cockroach. J. Exp. Biol. 2003, 206, 4453–4465. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.R.; Grant, M.D.; Tulli, N.M.; Karkowski, L.M.; McClearn, G.E. Differences in locomotor activity across the lifespan of Drosophila melanogaster. Exp. Gerontol. 1999, 34, 621–631. [Google Scholar] [CrossRef]
- Tofilski, A. Senescence and learning in honeybee (Apis mellifera) workers. Acta Neurobiol. Exp. 2000, 60, 35–39. [Google Scholar]
- Goldansaz, S.H.; McNeil, J.N. Effect of wind speed on the pheromone-mediated behavior of sexual morphs of the potato aphid, Macrosiphum euphorbiae (Thomas) under laboratory and field conditions. J. Chem. Ecol. 2006, 32, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Hardee, D.D.; McKibben, G.H.; Gueldner, R.C.; Mitchell, E.B.; Tumlinson, J.H.; Cross, W.H. Boll weevils in nature respond to grandlure, a synthetic pheromone. J. Econ. Entomol. 1972, 65, 97–100. [Google Scholar] [CrossRef]
- Duehl, A.J.; Arbogast, R.T.; Teal, P.E. Age and sex related responsiveness of Tribolium castaneum (Coleoptera: Tenebrionidae) in novel behavioral bioassays. Environ. Entomol. 2011, 40, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Boughton, A.; Fadamiro, H. Effect of Age and Sex on the Response of Walking Pvostephanus truncatus (Horn) (Coleoptera:Bostrichidae) to Its Male-Produced Aggregation Pheromone. J. Stored Prod. Res. 1996, 32, 13–20. [Google Scholar] [CrossRef]
- Kopanic, R.J.; Schal, C. Coprophagy Facilitates Horizontal Transmission of Bait among Cockroaches (Dictyoptera: Blattellidae). Environ. Entomol. 1999, 28, 431–438. [Google Scholar] [CrossRef]
- Roditakis, E.; Couzin, I.D.; Franks, N.R.; Charnley, A.K. Effects of Lecanicillium longisporum infection on the behaviour of the green peach aphid Myzus persicae. J. Insect Physiol. 2008, 54, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, S.A.; Parsons, N.M. The emergency life-history stage and immunity in the cricket, Gryllus texensis. Anim. Behav. 2006, 72, 235–244. [Google Scholar] [CrossRef]
- Doums, C.; Schmid-Hempel, P. Immunocompetence in workers of a social insect, Bombus terrestris L., in relation to foraging activity and parasitic infection. Can. J. Zool. 2000, 78, 1060–1066. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Response | Transformation | Fixed Effects | Random Effects |
|---|---|---|---|---|
| Experiment 1: Air movement | Walking speed | Air movement Pheromone Beetle collection | Date of experiment | |
| Distance walked | ||||
| Time to first contact with granule band | Natural log | |||
| Number of contacts with granule band | Square-root | |||
| Duration of contact with granule band | Natural log | |||
| Experiment 2: Light quality | Walking Speed | Light type Pheromone Beetle collection | Date of experiment | |
| Distance Walked | ||||
| Number of contacts with granules zone | Square-root | |||
| Duration of contact with granule zone | Natural log | |||
| Activity duration * | ||||
| Probability of reaching granule zone | Squared | |||
| Experiment 3: Seasonality | Walking speed | Pheromone Beetle collection | Date of experiment | |
| Distance walked | ||||
| Time spent moving | ||||
| Experiment 4: Response range | Proportion of beetles recovered ^ | Pheromone Release direction Release distance | Plot |
| Num DF | Den DF | F Value | Pr > F | |
|---|---|---|---|---|
| Walking speed | ||||
| Pheromone | 1 | 126 | 32.6 | <0.001 |
| Beetle collection | 2 | 126 | 9.64 | <0.001 |
| Pheromone: Beetle collection | 2 | 124 | 0.55 | 0.581 |
| Distance walked | ||||
| Pheromone | 1 | 126 | 32.75 | <0.001 |
| Beetle collection | 2 | 126 | 8.72 | <0.001 |
| Pheromone: Beetle collection | 2 | 124 | 0.47 | 0.624 |
| Movement duration | ||||
| Pheromone | 1 | 126 | 27.87 | <0.001 |
| Beetle collection | 2 | 126 | 6.35 | 0.002 |
| Pheromone: Beetle collection | 2 | 124 | 0.83 | 0.437 |
| Release Distance (m) | Mean Hours to Maximum Recapture (±SE) |
|---|---|
| 1 | 19.3 (4.2) |
| 3 | 13.1 (0.9) |
| 7 | 34.6 (2.6) |
| 14 | 38.1 (8.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leung, J.P.S.; Cory, J.S.; Kabaluk, J.T.; Janmaat, A.F. Effect of Collection Month, Visible Light, and Air Movement on the Attraction of Male Agriotes obscurus L. (Coleoptera: Elateridae) Click Beetles to Female Sex Pheromone. Insects 2020, 11, 729. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110729
Leung JPS, Cory JS, Kabaluk JT, Janmaat AF. Effect of Collection Month, Visible Light, and Air Movement on the Attraction of Male Agriotes obscurus L. (Coleoptera: Elateridae) Click Beetles to Female Sex Pheromone. Insects. 2020; 11(11):729. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110729
Chicago/Turabian StyleLeung, Joyce P. S., Jenny S. Cory, J. Todd Kabaluk, and Alida F. Janmaat. 2020. "Effect of Collection Month, Visible Light, and Air Movement on the Attraction of Male Agriotes obscurus L. (Coleoptera: Elateridae) Click Beetles to Female Sex Pheromone" Insects 11, no. 11: 729. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110729