Species Separation within, and Preliminary Phylogeny for, the Leafhopper Genus Anoscopus with Particular Reference to the Putative British Endemic Anoscopus duffieldi (Hemiptera: Cicadellidae)

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
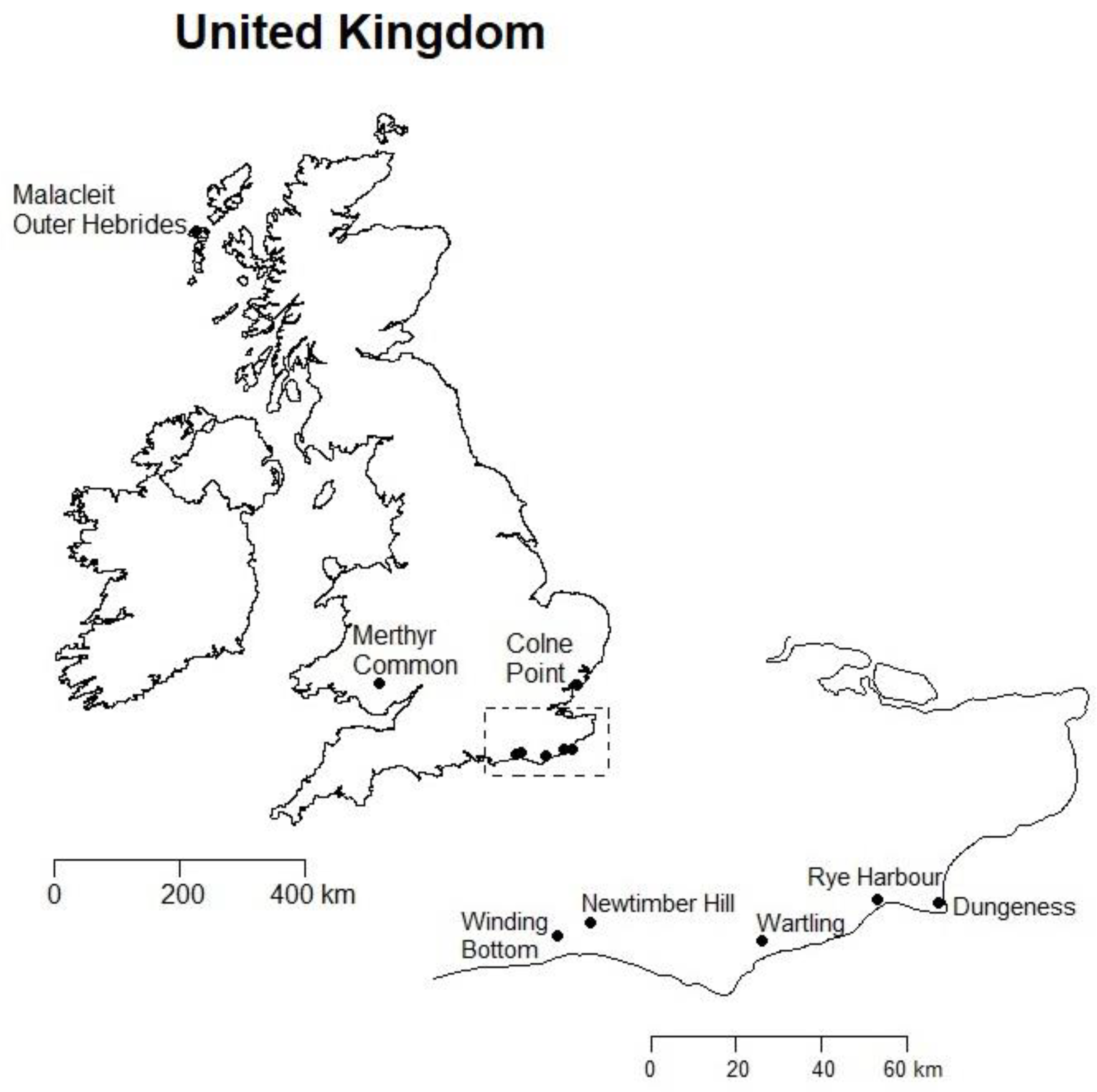
2.1. Specimen Collection
2.2. Choice of Molecular Markers
2.3. DNA Extraction and PCR Amplification
2.4. Sequencing Analysis
2.5. Phylogenetic Analyses
2.6. Population-Level Analyses
3. Results
3.1. Nucleotide and Haplotype Diversity
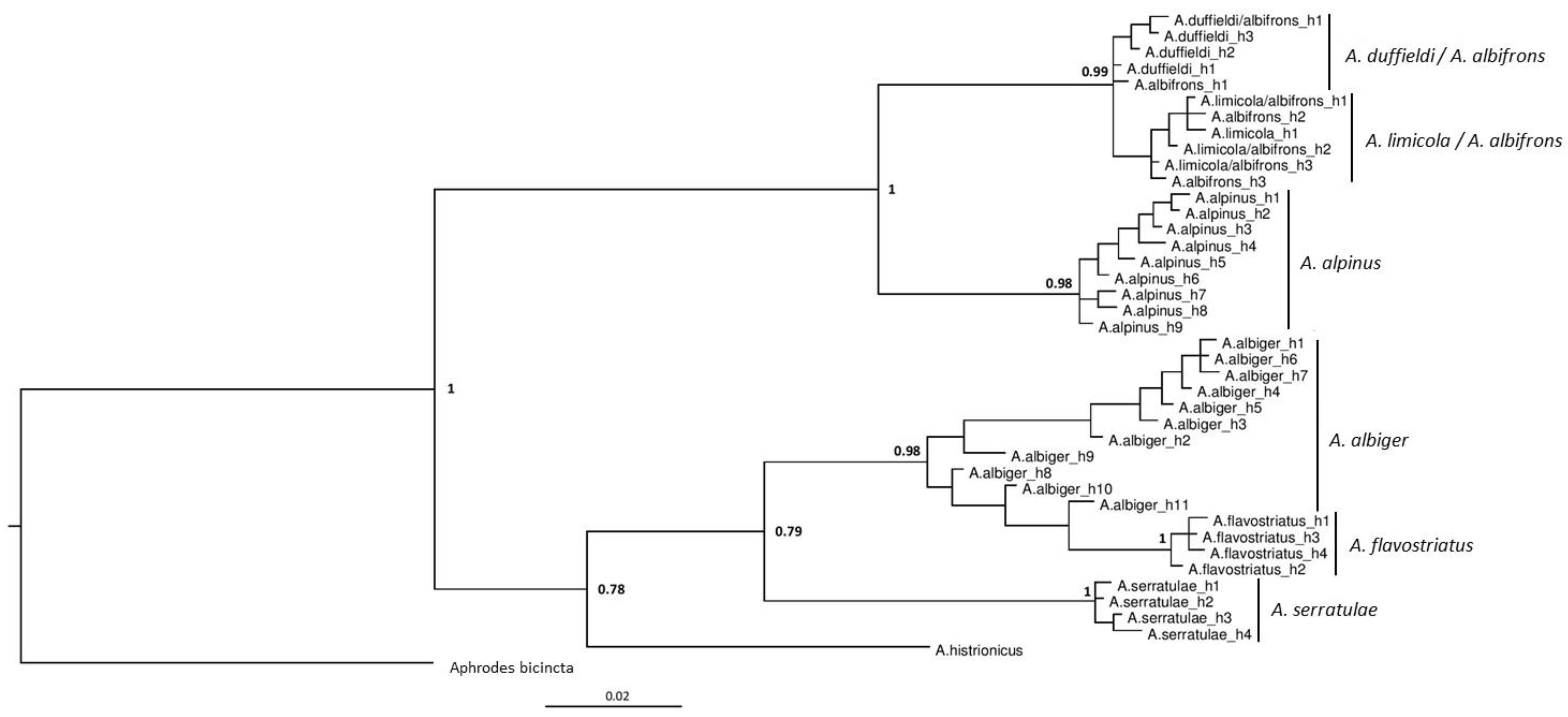
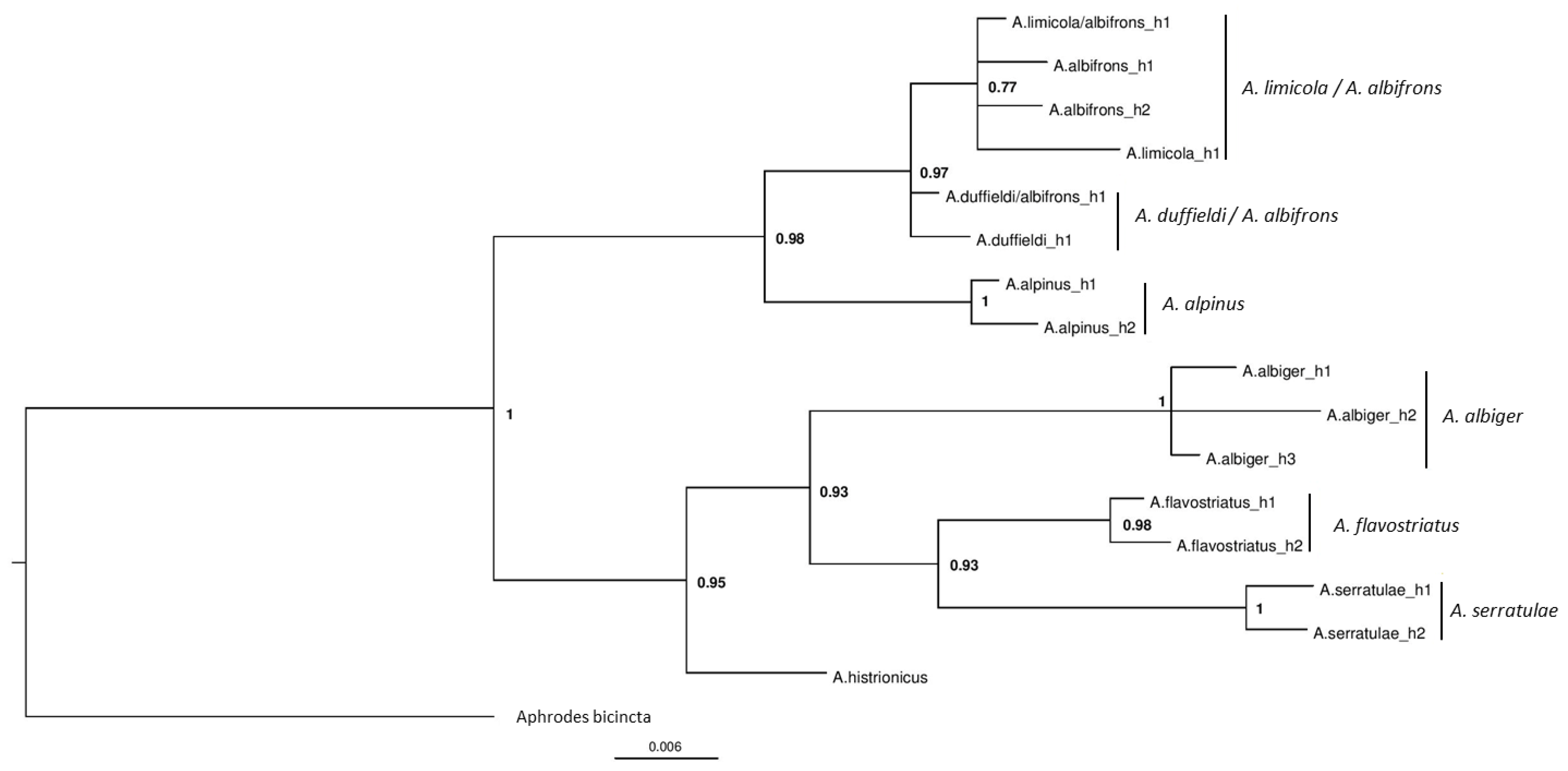
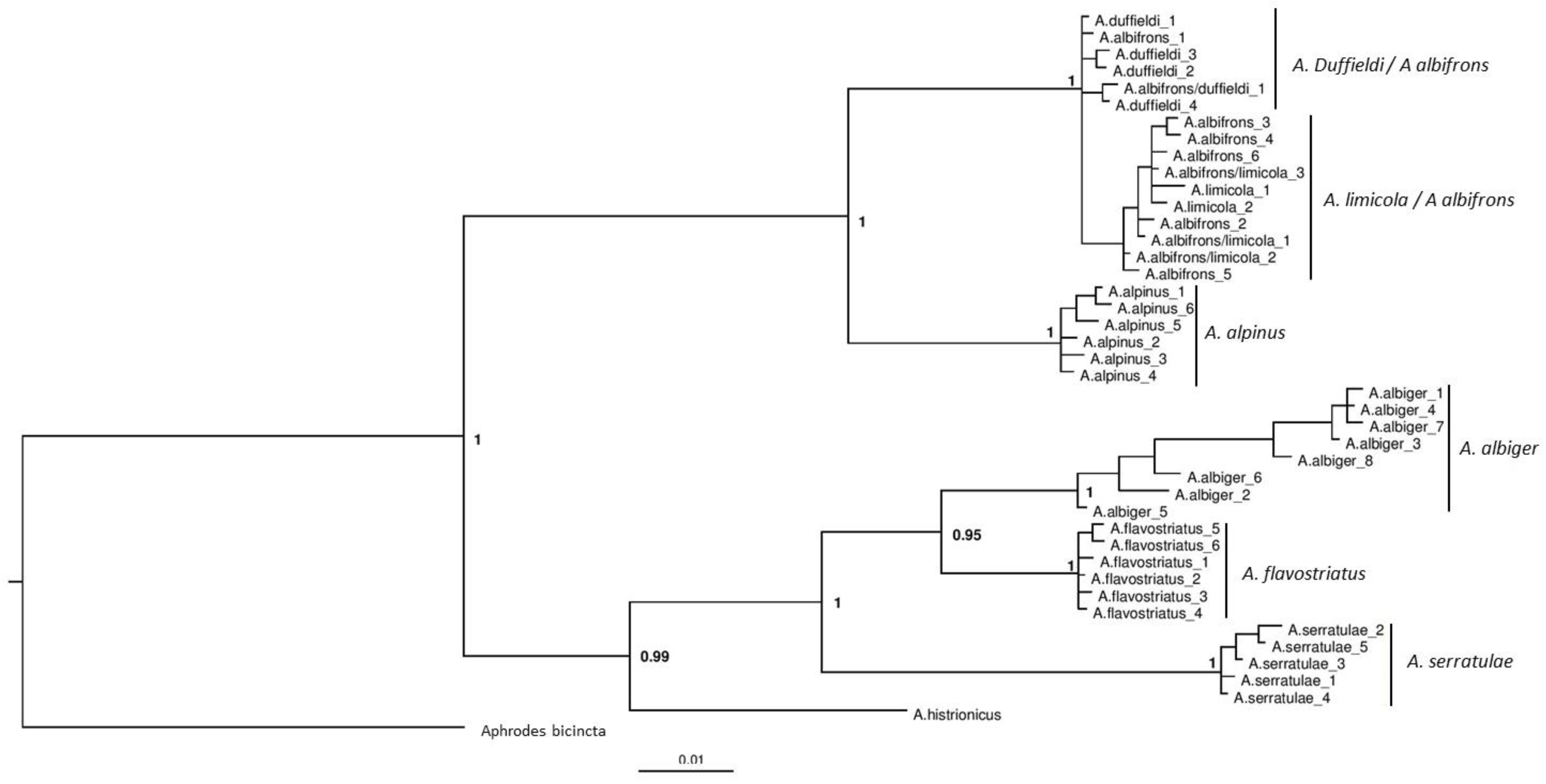
3.2. Phylogenetic Analyses
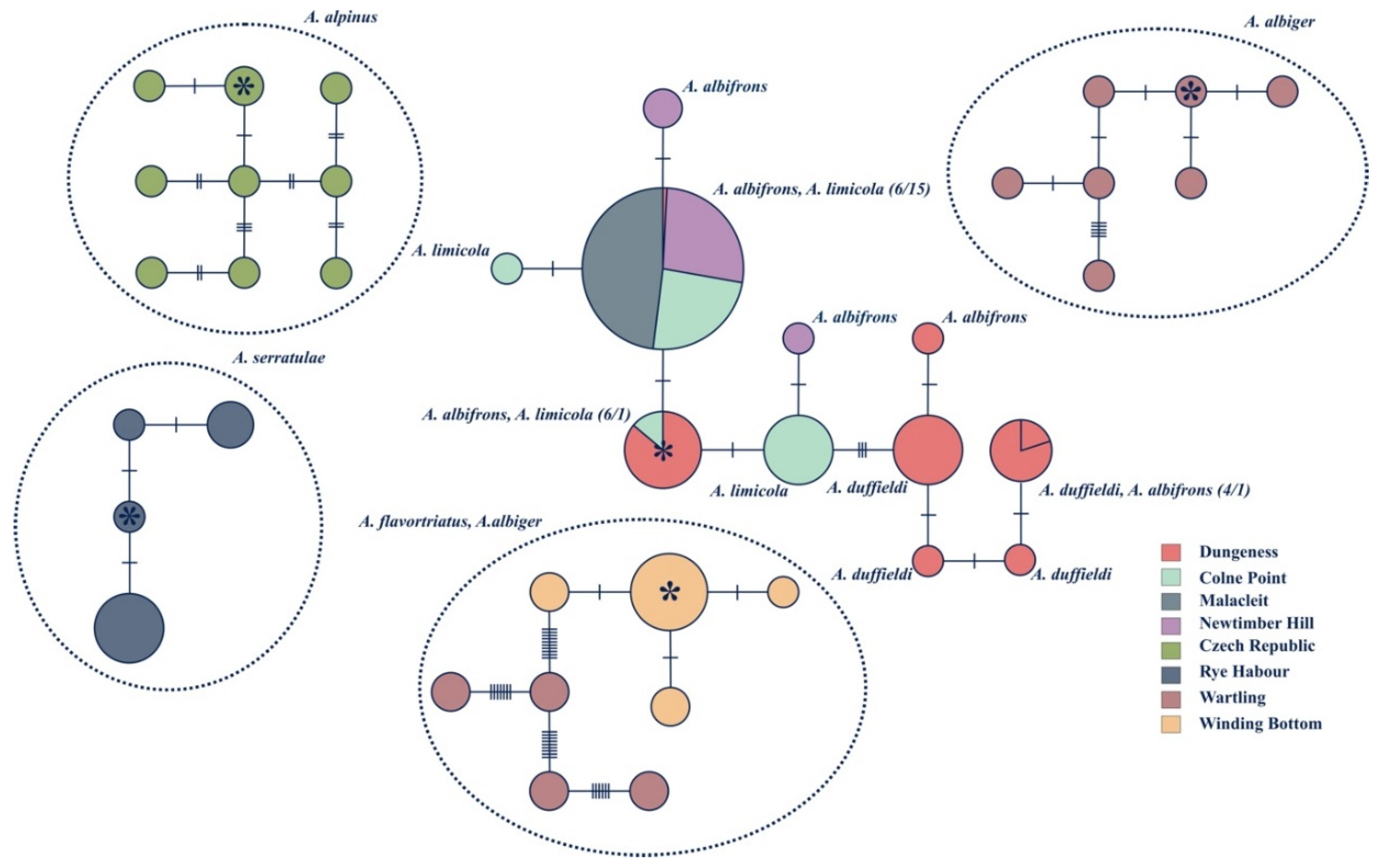
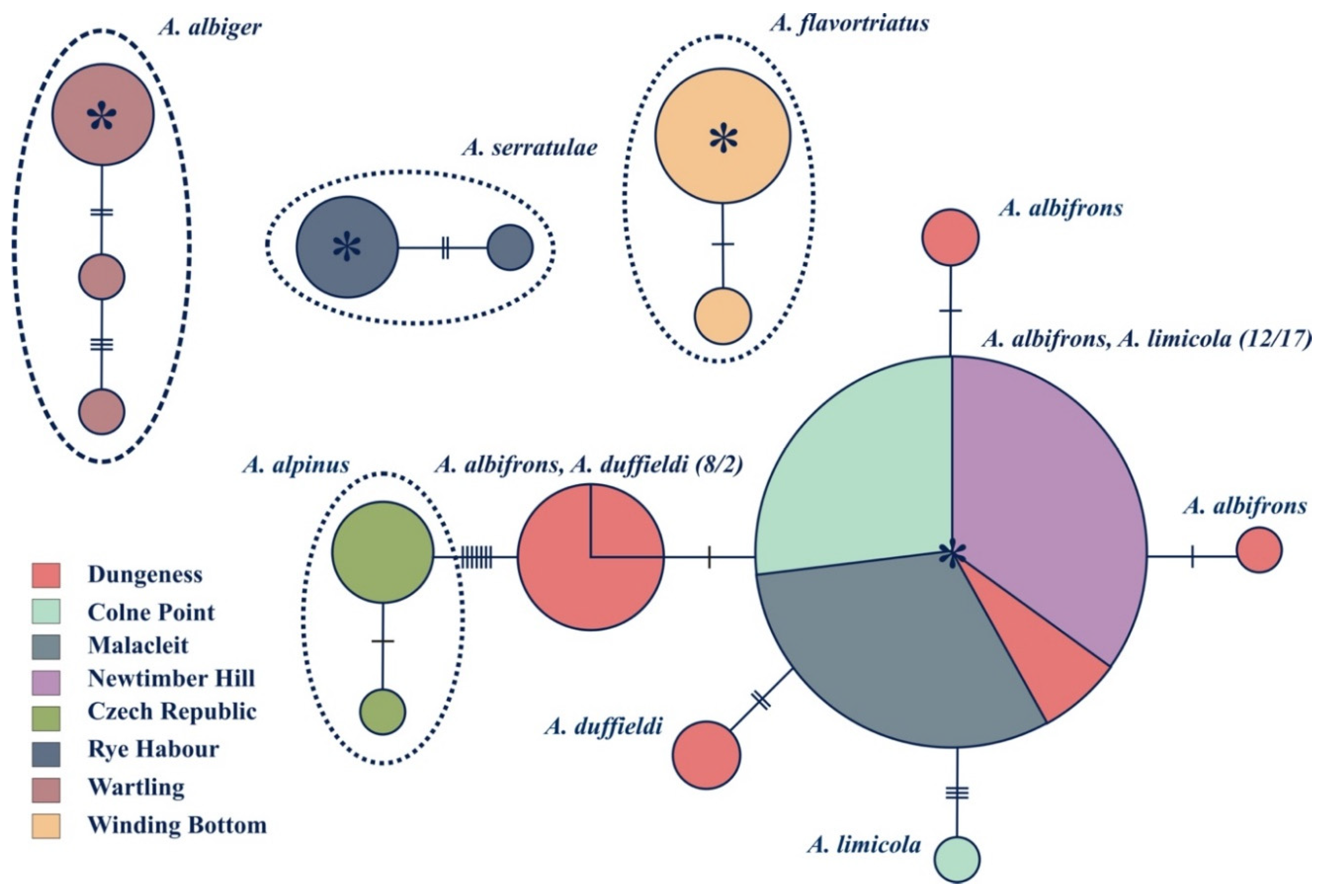
3.3. Population Level Analyses
4. Discussion
4.1. Anoscopus duffieldi and Related Taxa (Albifrons Subgroup)
4.2. Other Anoscopus Species (Albiger Subgroup)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Material
References
- Austin, J.J.; Melville, J. Incorporating historical museum specimens into molecular systematic and conservation genetics research. Mol. Ecol. Notes 2006, 6, 1089–1092. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Dexter, K.G.; Pennington, T.D.; Cunningham, C.W. Using DNA to assess errors in tropical tree identifications: How often are ecologists wrong and when does it matter? Ecol. Monogr. 2010, 80, 267–286. [Google Scholar] [CrossRef] [Green Version]
- Lis, B.; Lis, B.; Ziaja, D.J. In BOLD we trust? A commentary on the reliability of specimen identification for DNA barcoding: A case study on burrower bugs (Hemiptera: Heteroptera: Cydnidae). Zootaxa 2016, 4114, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Bluemel, J.K.; King, R.; Virant-Doberlet, M.; Symondson, W.O.C. Primers for identification of type and other archived specimens of Aphrodes leafhoppers (Hemiptera, Cicadellidae). Mol. Ecol. Resour. 2011, 11, 770–774. [Google Scholar] [CrossRef]
- Seabra, S.G.; Pina-Martins, F.; Marabuto, E.; Yurtsever, S.; Halkka, O.; Quartau, J.A.; Paulo, O.S. Molecular phylogeny and DNA barcoding in the meadow-spittlebug Philaenus spumarius (Hemiptera, Cercopidae) and its related species. Mol. Phylogenet. Evol. 2010, 56, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.M. Reliability of long vs short COI markers in identification of forensically important flies. Croat. Med. J. 2014, 55, 19–26. [Google Scholar] [CrossRef] [Green Version]
- De Mandal, S.; Chhakchhuak, L.; Gurusubramanian, G.; Kumar, N.S. Mitochondrial markers for identification and phylogenetic studies in insects—A review. DNA Barcodes 2014, 2, 1–9. [Google Scholar] [CrossRef]
- Bluemel, J.K.; Derlink, M.; Pavlovčič, P.; Russo, I.-R.M.; King, R.A.; Corbett, E.; Sherrard-Smith, E.; Blejec, A.; Wilson, M.R.; Stewart, A.J.A.; et al. Integrating vibrational signals, mitochondrial DNA and morphology for species determination in the genus Aphrodes (Hemiptera: Cicadellidae). Syst. Entomol. 2014, 39, 304–324. [Google Scholar] [CrossRef]
- Kirby, P.; Stewart, A.J.; Wilson, M.R. True bugs, leaf-and planthoppers, and their allies. In The Changing Wildlife of Great Britain and Ireland; Hawksworth, D.L., Ed.; CRC Press: Boco Raton, FL, USA, 2001; Volume 17, pp. 262–299. [Google Scholar]
- Forero, D. The systematics of the Hemiptera. Rev. Colomb. Entomol. 2008, 34, 1–21. [Google Scholar]
- Nickel, H. The Leafhoppers and Planthoppers of Germany (Hemiptera: Auchenorrhyncha): Patterns and Strategies in a Highly Diverse Group of Phytophagous Insects; Pensoft: Sofia, Bulgaria; Goecke & Evers: Keltern, Germany, 2003. [Google Scholar]
- Bartlett, C.R.; Deitz, L.L.; Dmitriev, D.A.; Sanborn, A.F.; Soulier-Perkins, A.; Wallace, M.S. The diversity of the true hoppers (Hemiptera: Auchenorrhyncha). In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons: Chichester, UK, 2018; Volume 2, pp. 501–590. [Google Scholar]
- Biedermann, R.; Achtziger, R.; Nickel, H.; Stewart, A.J.A. Conservation of grassland leafhoppers: A brief review. J. Insect Conserv. 2005, 9, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Zahniser, J.N.; Dietrich, C.H. Phylogeny of the leafhopper subfamily Deltocephalinae (Insecta: Auchenorrhyncha: Cicadellidae) and related subfamilies based on morphology. Syst. Biodivers. 2008, 6, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C.H. Overview of the phylogeny, taxonomy and diversity of the leafhopper (Hemiptera: Auchenorrhyncha: Cicadomorpha: Membracoidea: Cicadellidae) vectors of plant pathogens. In Proceedings of the 2013 International Symposium on Insect Vectors and Insect-Borne Diseases, Taichung, Taiwan, August 2013; Chang, C.-I., Lee, C.-Y., Hsien-Tzung Shih, H.-T., Eds.; Special Publication of TARI; Taiwan Agricultural Research Institute: Taichung, Taiwan, 2013; Volume 173, pp. 47–70. [Google Scholar]
- Rothenbücher, J.; Schaefer, M. Conservation of leafhoppers in floodplain grasslands—Trade-off between diversity and naturalness in a northern German national park. J. Insect Cons. 2005, 9, 335–349. [Google Scholar] [CrossRef]
- Schuch, S.; Wesche, K.; Schaefer, M. Long-term decline in the abundance of leafhoppers and planthoppers (Auchenorrhyncha) in Central European protected dry grasslands. Biol. Conserv. 2012, 149, 75–83. [Google Scholar] [CrossRef]
- Endrestøl, A.; Elven, H. Two species of Aphrodinae (Hemiptera, Cicadellidae) new to the Norwegian fauna. Norweg. J. Entomol. 2009, 56, 24–27. [Google Scholar]
- Guglielmino, A.; Bückle, C. Revision of Errhomeninae and Aphrodinae (Hemiptera, Cicadomorpha) in Italy with remarks on their variability and distribution in adjacent regions and description of three new taxa. Zootaxa 2015, 3906, 1–66. [Google Scholar] [CrossRef]
- Wilson, M.R.; Stewart, A.; Biedermann, R.; Nickel, H.; Niedringhaus, R. The Planthoppers and Leafhoppers of Britain and Ireland. Identification Keys to All Families and Genera and all British and Irish Species not Recorded from Germany. Cicadina, Supplement 2; WABV Fründ: Scheeßel, Germany, 2015. [Google Scholar]
- Ribaut, H. Homoptères Auchénorhinques. II. (Jassidae); Faune de France: Paris, France, 1952; Volume 57, p. 474. [Google Scholar]
- Le Quesne, W.J. Hemiptera (Cicadomorpha), excluding Deltocephalinae and Typhlocybinae. Handbooks for the Identification of British Insects; Royal Entomological Society: London, UK, 1965. [Google Scholar]
- Hamilton, K.G.A. A review of the northern hemisphere aphrodina (Rhynchota: Homoptera: Cicadellidae), with special reference to the nearctic fauna. Can. Entomol. 1975, 107, 1009–1027. [Google Scholar] [CrossRef]
- Biedermann, R.; Niedringhaus, R. The Plant- and Leafhoppers of Germany: Identification Key to All Species; WABV Fründ: Scheeßel, Germany, 2009. [Google Scholar]
- Barnett, D.E. Some new preparation techniques used in leafhopper identification. Fla. Entomol. 1976, 59, 321–323. [Google Scholar] [CrossRef]
- Remane, R.; Fröhlich, W. Beiträge zur Chorologie einiger Zikaden-Arten (Homoptera Auchenorrhyncha) in der Westpaläarktis. Marbg. Entomol. Publik. 1994, 2, 131–188. [Google Scholar]
- Tishechkin, D.Y. On the taxonomy and distribution of Aphrodes bicincta (Schrank, 1776) species group (Homoptera: Auchenorrhyncha: Cicadellidae: Aphrodinae) in Eastern Palaearctic. Zootaxa 2017, 4318, 167–176. [Google Scholar] [CrossRef]
- Duffield, C.A.W. The genus aphrodes (Homoptera; Auchenorhyncha). Trans. Kent Field Club 1963, 1, 155–160. [Google Scholar]
- Le Quesne, W.J. Some taxonomic changes and additions in the British Cicadellidae (Hemiptera) including a new species and subspecies. Proc. R. Entomol. Soc. B 1964, 33, 73–116. [Google Scholar] [CrossRef]
- Nast, J. Palaearctic Auchenorrhyncha (Homoptera) an Annotated Check List; Polish Scientific Publishers: Warsaw, Poland, 1992. [Google Scholar]
- Key, R.S.; Drake, C.M.; Sheppard, D.A. Conservation of Invertebrates in England: A Review and Framework; English Nature Science Report 35; English Nature: Peterborough, UK, 2000. [Google Scholar]
- Ossiannilsson, F. The auchenorrhyncha (homoptera) of fennoscandia and Denmark. In Fauna Entomologica Scandinavica—Part 2; Scandinavian Science Press: Klampenborg, Denmark, 1981; Volume 7. [Google Scholar]
- Pentinsaari, M.; Salmela, H.; Mutanen, M.; Roslin, T. Molecular evolution of a widely-adopted taxonomic marker (COI) across the animal tree of life. Sci. Rep. 2016, 6, 35275. [Google Scholar] [CrossRef] [PubMed]
- Percy, D.M. Making the most of your host: The Metrosideros-feeding psyllids (Hemiptera, Psylloidea) of the Hawaiian Islands. ZooKeys 2007, 649, 1–163. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C.; Rakitov, R.; Holmes, J.; Black, W. Phylogeny of the major lineages of membracoidea (Insecta: Hemiptera: Cicadomorpha) based on 28S rDNA sequences. Mol. Phylogenet. Evol. 2001, 18, 293–305. [Google Scholar] [CrossRef]
- Dai, R.H.; Chen, X.X.; Li, Z.Z. Phylogeny of Deltocephalinae (Hemiptera: Cicadellidae) from China based on partial 16S rDNA and 28S rDNA D2 sequences combined with morphological characters. Acta Entomol. Sin. 2008, 51, 1055–1064. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Fu, J.-Y.; Han, B.-Y.; Xiao, Q. Mitochondrial COI and 16sRNA evidence for a single species hypothesis of E. vitis, J. formosana and E. onukii in East Asia. PLoS ONE 2014, 9, e115259. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef] [PubMed]
- Zahniser, J.N. Systematics of the Leafhopper Subfamily Deltocephalinae (Hemiptera: Cicadellidae) and the Tribe Chiasmini: Phylogeny, Classification, and Biogeography. Ph.D Thesis, University of Illinois, Champaign, IL, USA, 2008. Available online: http://hdl.handle.net/2142/86469 (accessed on 28 October 2020).
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; Mcgettigan, P.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Goldman, N.; Friday, A. Comparison of models from nucleotide substitution used in maximum likelihood phylogenetic estimation. Mol. Biol. Evol. 1994, 11, 316–324. [Google Scholar] [PubMed] [Green Version]
- Yang, Z. Among-site rate variation and its impact on phylogenetic analyses. Trends Ecol. Evol. 1996, 144, 1941–1950. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.; Xie, W.; Drummond, A. Tracer v. 1.6; Institute of Evolutionary Biology, University of Edinburgh. 2014. Available online: https://github.com/beast-dev/tracer/releases/tag/v1.6 (accessed on 28 October 2020).
- Rambaut, A. FigTree 1.4. 2; University of Edinburgh: Edinburgh, Scotland. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 28 October 2020).
- Nei, M.; Tajima, F. Genetic drift and estimation of effective population size. Genetics 1981, 98, 625–640. [Google Scholar]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; p. 512. [Google Scholar]
- Rozas, J.; Rozas, R. DnaSP version 3: An integrated program for molecular population genetics and molecular evolution analysis. Bioinformatics 1999, 15, 174–175. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, UK, 2000. [Google Scholar]
- Gwiazdowski, R.A.; Foottit, R.G.; Maw, H.E.L.; Hebert, P.D.N. The Hemiptera (Insecta) of Canada: Constructing a reference library of DNA barcodes. PLoS ONE 2015, 10, e0125635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albre, J.; Gibernau, M. Diversity and temporal variations of the Hemiptera Auchenorrhyncha fauna in the Ajaccio region (France, Corsica). Ann. Soc. Entomol. Fr. 2019, 55, 497–508. [Google Scholar] [CrossRef]
- Lindberg, H. Hemiptera insularum canariensium. systematik, ökologie und verbreitung der kanarischen heteropteren und cicadinen. Comment. Biol. 1954, 14, 1–304. [Google Scholar]
- Malenovský, I. New records of Auchenorrhyncha (Hemiptera) for the Czech Republic. Acta Musei Morav. Scient. Boil. 2013, 98, 235–263. [Google Scholar]
- Della Giustina, W. Homoptères Cicadellidae. Compléments aux ouvrages d’Henri Ribaut. In Faune de France, 73; Fédération Française des Sociétés de Sciences Naturelles et INRA: Paris, France, 1989; Volume 3. [Google Scholar]
- Bennett, G.M.; O’Grady, P.M. Review of the native Hawaiian leafhopper genus Nesophrosyne (Hemiptera: Cicadellidae: Deltocephalinae) with description of eight new species associated with Broussaisia arguta (Hydrangeaceae). Zootaxa 2011, 2805, 1–25. [Google Scholar] [CrossRef]
- Derlink, M.; Pipan, B.; Pavlovčič, P.; Jones, L.; Meglič, V.; Symondson, W.O.C.; Virant-Doberlet, M. Characterization of eleven polymorphic microsatellite markers for leafhoppers of the genus Aphrodes (Hemiptera: Cicadellidae). Conserv. Genet. Resour. 2014, 6, 933–935. [Google Scholar] [CrossRef]
- Morris, R.K.A.; Parsons, M.S. A survey of invertebrate communities on the shingle of Dungeness, Rye Harbour and Orford Ness. In JNCC Report 77; Joint Nature Conservation Committee: Peterborough, UK, 1992. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anoscopus Species | Collection Site | Ordnance Survey + | Co-Ordinates (N, E) | Year Collected | Storage |
|---|---|---|---|---|---|
| albifrons | Dungeness, Kent | TR076190 | 50.933074, 0.95310771 | 2013 | 100% Ethanol * |
| Newtimber Hill, Sussex | TQ268119 | 50.892674, −0.19846861 | 2015 | 100% Ethanol | |
| albiger | Wartling, Sussex | TQ666085 | 50.852027, 0.36542503 | 2014/15 | Dried/100% Ethanol |
| alpinus | Mt Kralicky Sneznik, Czech Rep. | 50.206401, 16.849404 | 2015 | 100% Ethanol | |
| duffieldi | Dungeness, Kent | TR076190 | 50.933074, 0.95310771 | 2014/15 | 100% Ethanol |
| flavostriatus | Winding Bottom, Sussex | TQ191087 | 50.865548, −0.30893968 | 2013 | Dried |
| 2014/15 | 100% Ethanol | ||||
| limicola | Colne Point, Essex | TM108124 | 51.770566, 1.0538762 | 2011 | 100% Ethanol * |
| Malacleit, Outer Hebrides | NF790730 | 56.632199, −7.3805002 | 2012 | Dried | |
| serratulae | Rye Harbour, Sussex | TQ931192 | 50.939904, 0.74711850 | 2015 | 100% Ethanol |
| histrionicus | Merthyr Common, Wales | SO071058 | 51.743173, −3.3469341 | 2015 | Ethanol § |
| Locus | Primer Name | Primer Sequence (5′–3′) | Reference |
|---|---|---|---|
| CO1 | LCO1490 | GGTCAACAAATCATAAAGATATTGG | Folmer et al., 1994 [40] |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | ||
| 16S | LR-J-12887 | CCGGTYTGAACTCARATCAWGT | Fu et al., 2014 [41] |
| LR-N-13398 | CTGTTTAWCAAAAACATTTC | ||
| 28S | 28SP | AGTCGKGTTGCTTGAKAGTGCAG | Zahniser, 2008 [43] |
| 28SM2 | TTCGGGTCCCAACGTGTACG | ||
| 28SII’(F) | GGGACCCGTCTTGAAACAC | Dietrich et al., 2001 [38] | |
| 28SII’(R) | ACCCTCCTACTCGTCAAGG |
| Species | CO1 | 16S | ||
|---|---|---|---|---|
| d (%) | S.E. (%) | d (%) | S.E. (%) | |
| albifrons | 0.8 | 0.2 | 0.4 | 0.2 |
| albiger | 2.4 | 0.4 | 0.7 | 0.3 |
| alpinus | 0.6 | 0.2 | 0.3 | 0.3 |
| duffieldi | 0.3 | 0.2 | 0.3 | 0.3 |
| flavostriatus | 0.3 | 0.2 | 0.3 | 0.3 |
| limicola | 0.3 | 0.1 | 0.8 | 0.5 |
| serratulae | 0.4 | 0.2 | 0.5 | 0.3 |
| Species | CO1 | 16S rRNA | ||
|---|---|---|---|---|
| Haplotype Diversity | Nucleotide Diversity | Haplotype Diversity | Nucleotide Diversity | |
| A. albifrons | 0.81 | 0.004 | 0.5 | 0.001 |
| A. albiger | 1.00 | 0.024 | 0.46 | 0.003 |
| A. alpinus | 0.98 | 0.006 | 0.29 | 0.0007 |
| A. duffieldi | 0.68 | 0.003 | 0.44 | 0.001 |
| A. flavostriatus | 0.65 | 0.001 | 0.33 | 0.0009 |
| A. limicola | 0.38 | 0.001 | 0.11 | 0.0009 |
| A. serratulae | 0.81 | 0.004 | 0.29 | 0.001 |
| Loci and Species Name | Mean Genetic Distance/Standard Error (%) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| CO1 | ||||||
| 1. albifrons | 1 | 0.9 | 0.3 | 1.2 | 0.2 | |
| 2. albiger | 10.2 | 1.2 | 1 | 0.7 | 1.1 | |
| 3. alpinus | 6.2 | 10.6 | 0.9 | 1.3 | 0.9 | |
| 4. duffieldi | 0.35 | 10.05 | 6.35 | 1.2 | 0.4 | |
| 5. flavostriatus | 13.25 | 4.15 | 13.25 | 13.1 | 1.2 | |
| 6. limicola | 0.05 | 10.65 | 6.55 | 0.8 | 13.7 | |
| 7. serratulae | 11.4 | 6.7 | 11.7 | 11.15 | 8.25 | 11.85 |
| 16S | ||||||
| 1. albifrons | 1.2 | 0.7 | 0.2 | 1 | 0.1 | |
| 2. albiger | 5.25 | 1.1 | 1.2 | 0.8 | 1.2 | |
| 3. alpinus | 1.95 | 5.2 | 0.6 | 1 | 0.6 | |
| 4. duffieldi | 0.15 | 5.2 | 1.9 | 1 | 0.3 | |
| 5. flavostriatus | 5.25 | 3.6 | 4.9 | 5.2 | 1 | |
| 6. limicola | 0 | 5.35 | 2.15 | 0.15 | 5.35 | |
| 7. serratulae | 5.05 | 4.1 | 5.2 | 4.9 | 2.6 | 5.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redihough, J.; Russo, I.-R.M.; Stewart, A.J.A.; Malenovský, I.; Stockdale, J.E.; Moorhouse-Gann, R.J.; Wilson, M.R.; Symondson, W.O.C. Species Separation within, and Preliminary Phylogeny for, the Leafhopper Genus Anoscopus with Particular Reference to the Putative British Endemic Anoscopus duffieldi (Hemiptera: Cicadellidae). Insects 2020, 11, 799. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110799
Redihough J, Russo I-RM, Stewart AJA, Malenovský I, Stockdale JE, Moorhouse-Gann RJ, Wilson MR, Symondson WOC. Species Separation within, and Preliminary Phylogeny for, the Leafhopper Genus Anoscopus with Particular Reference to the Putative British Endemic Anoscopus duffieldi (Hemiptera: Cicadellidae). Insects. 2020; 11(11):799. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110799
Chicago/Turabian StyleRedihough, Joanna, Isa-Rita M. Russo, Alan J. A. Stewart, Igor Malenovský, Jennifer E. Stockdale, Rosemary J. Moorhouse-Gann, Michael R. Wilson, and William O. C. Symondson. 2020. "Species Separation within, and Preliminary Phylogeny for, the Leafhopper Genus Anoscopus with Particular Reference to the Putative British Endemic Anoscopus duffieldi (Hemiptera: Cicadellidae)" Insects 11, no. 11: 799. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110799