Foraging Bumblebees Selectively Attend to Other Types of Bees Based on Their Reward-Predictive Value


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
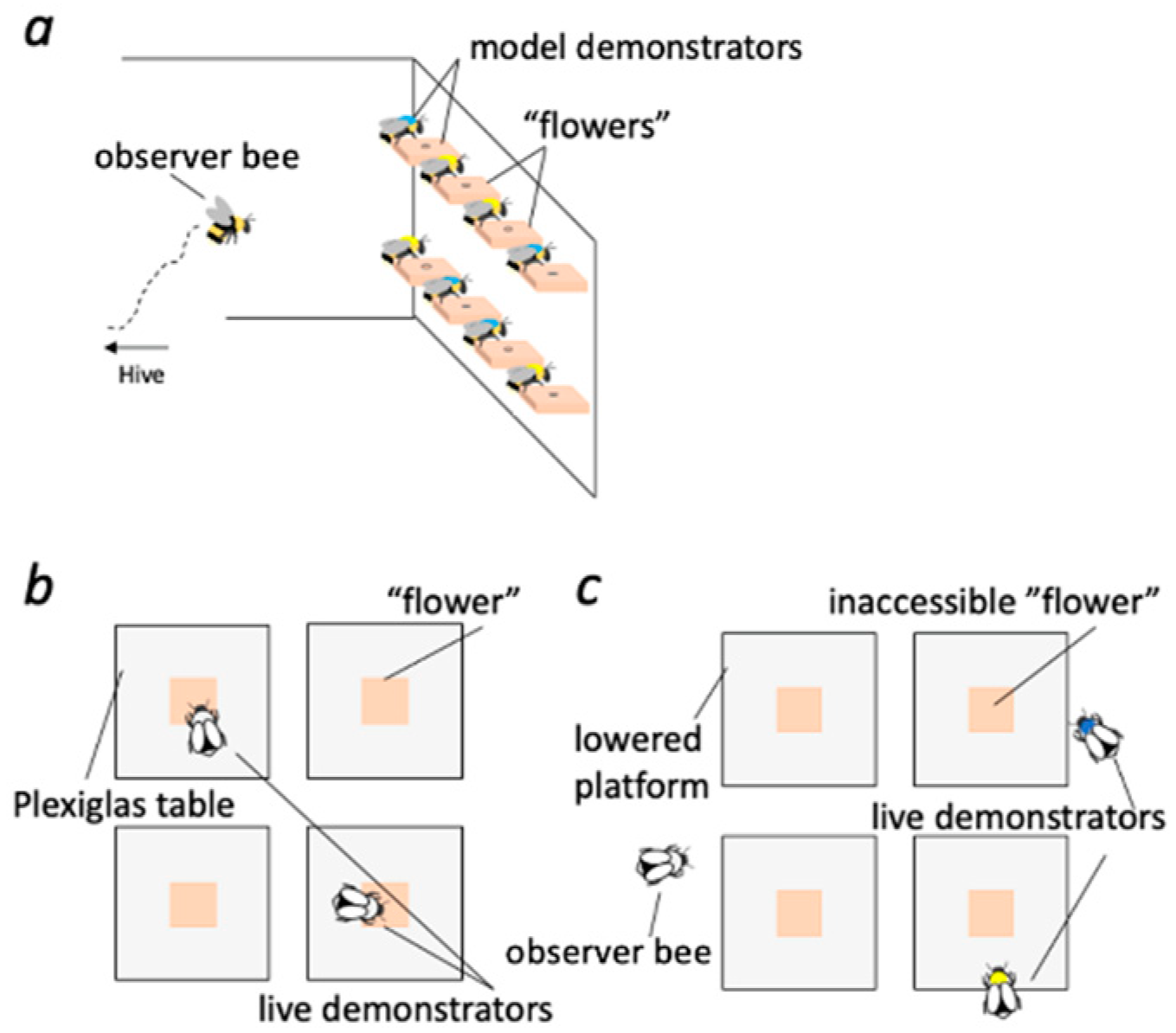
2.1. Set-Up
2.2. Pre-Training of Observer Bees
2.3. Training Observer Bees to Differentiate Model Demonstrators by Their Reliability to Predict Reward
2.4. Confirming Individual Learning of Observer Bees
2.5. Training of Live Demonstrators
2.6. Testing Observer Bees in a Novel Context with Live Demonstrators
2.7. Control Groups
2.8. Behaviors Measured in Test and Control Trials
2.9. Analysis
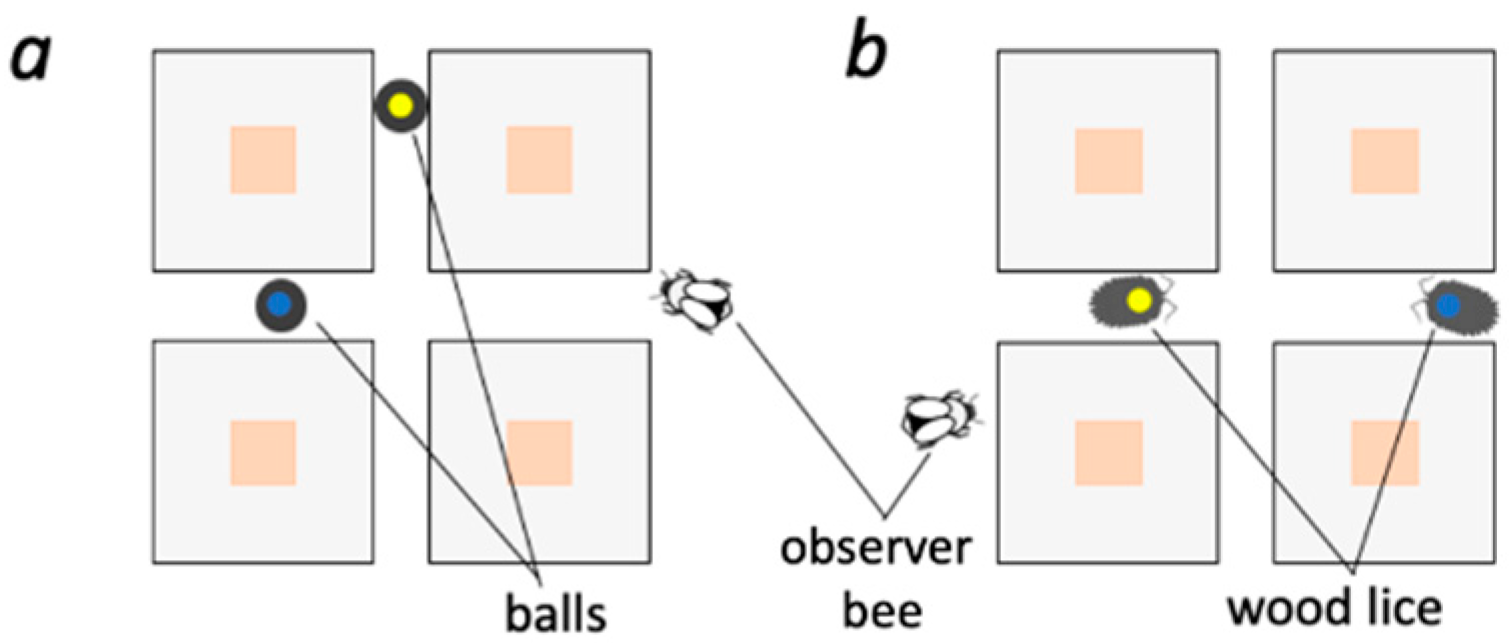
2.9.1. Do Bumblebees Preferentially Attend to Other Bees Rather than Simple Color Cues or Generic Insect-Shaped Cues?
2.9.2. Discriminating between Model Demonstrators of Differing Visual Appearance and Informative Value as Reward Predictors
2.9.3. The Effect of Experience on Observer Bees’ Attention to Live Demonstrators Differing in Their Informative Value as Reward Predictors
2.9.4. The Effect of Live Demonstrators’ Informative Value as Reward Predictors on in-Flight Decisions
3. Results
3.1. Do Bumblebees Preferentially Attend to Other Bees Rather Than Simple Color Cues or Generic Insect-Shaped Cues?
3.2. Discriminating between Model Demonstrators of Differing Visual Appearance and Informative Value as Reward Predictors
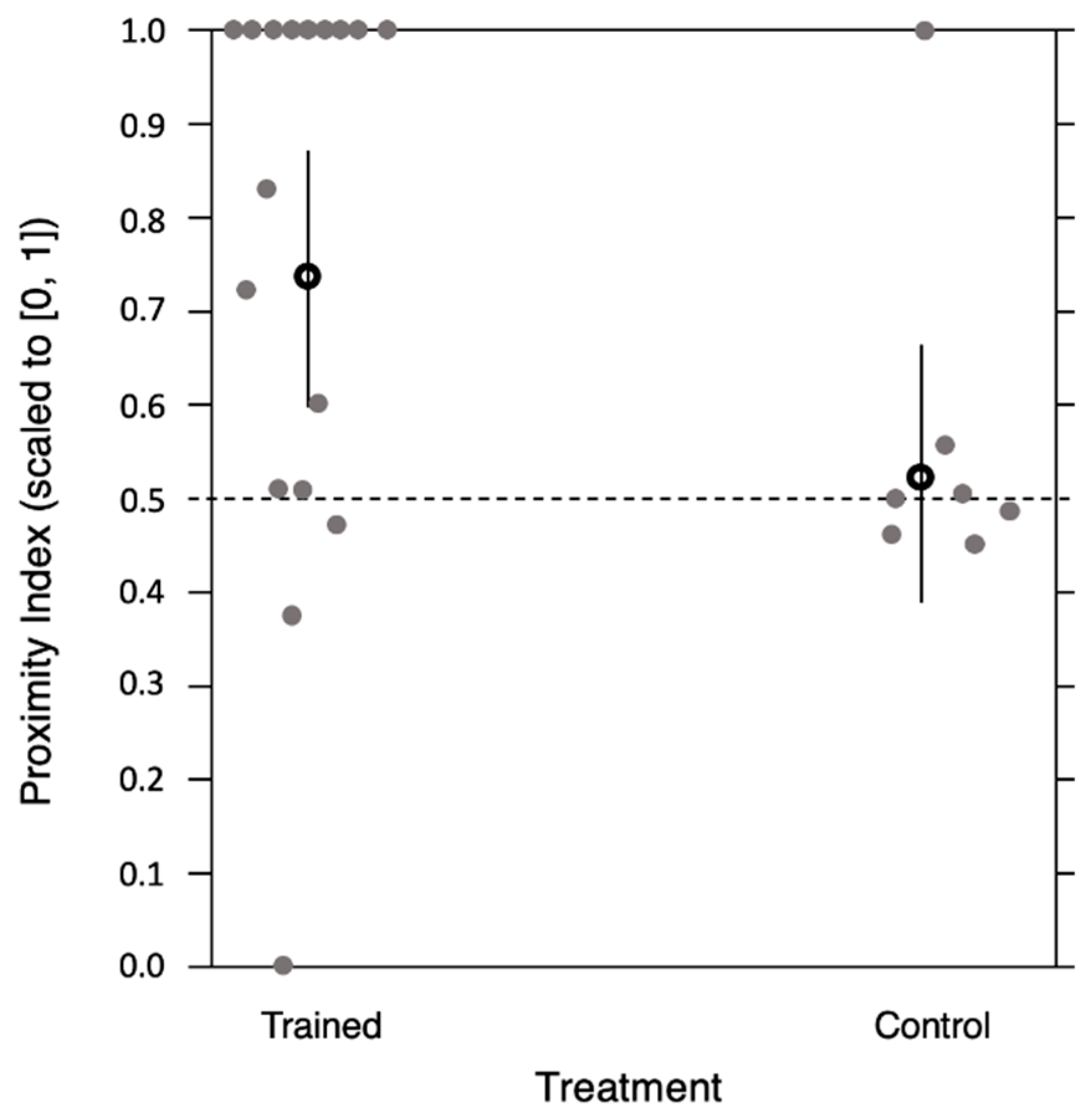
3.3. The Effect of Experience on Observer Bees’ Attention to Live Demonstrators Differing in Their Informative Value as Reward Predictors
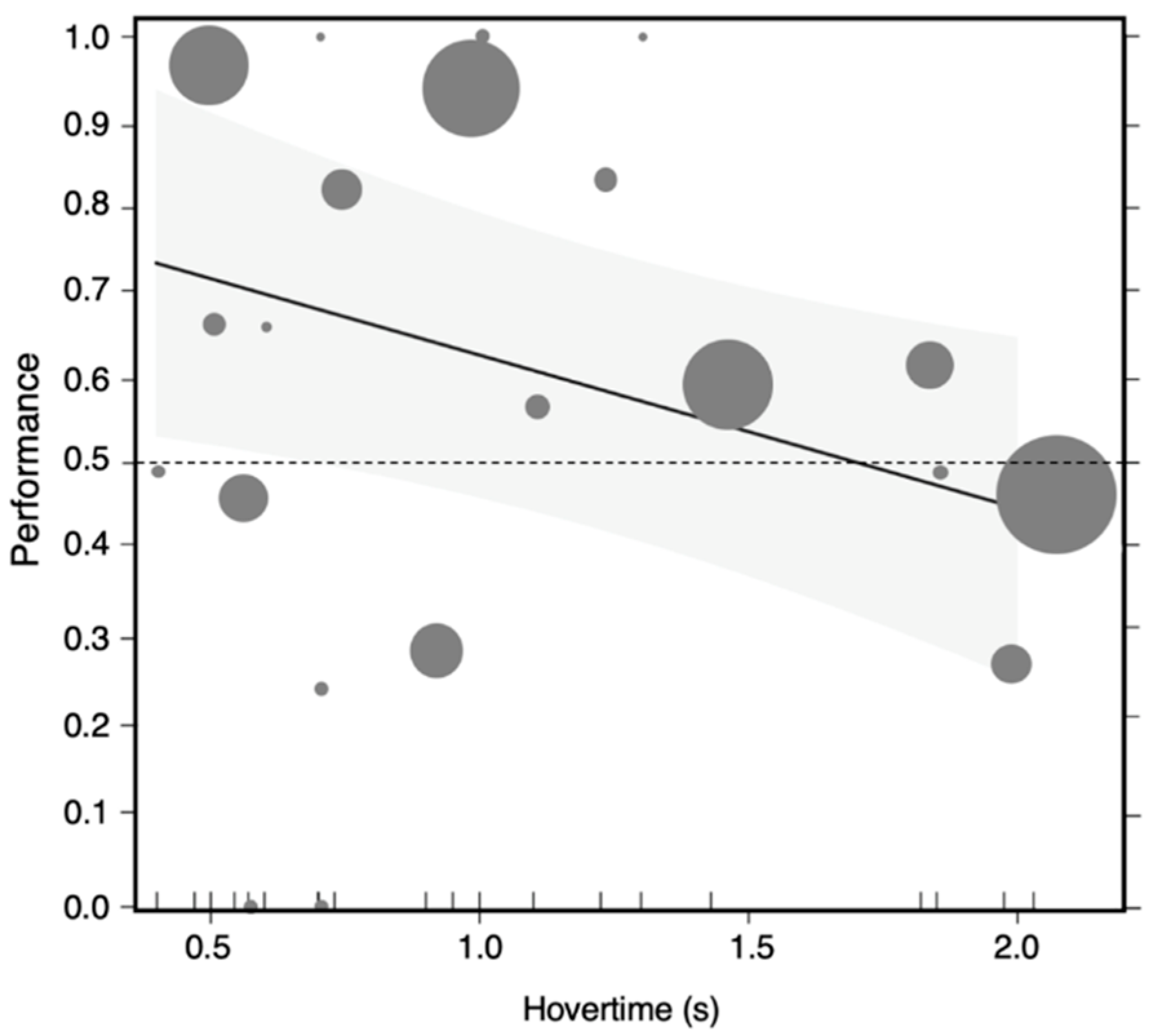
3.4. The Effect of Live Demonstrators’ Informative Value as Reward Predictors on In-Flight Decisions
4. Discussion
4.1. Benefits of Using Social Information
4.2. Associative Mechanisms Underlying Social Learning
4.3. Suggestions for Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galef, B.G.; Laland, K.N. Social Learning in Animals: Empirical Studies and Theoretical Models. BioScience 2005, 55, 489–499. [Google Scholar] [CrossRef]
- Forsman, J.T.; Seppänen, J.-T.; Mönkkönen, M. Positive fitness consequences of interspecific interaction with a potential competitor. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1619–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danchin, É.; Giraldeau, L.-A.; Valone, T.J.; Wagner, R.H. Public information: From nosy neighbors to cultural evolution. Science 2004, 305, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall, S.R.X.; Giraldeau, L.-A.; Olsson, O.; McNamara, J.M.; Stephens, D.W. Information and its use by animals in evolutionary ecology. Trends Ecol. Evol. 2005, 20, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Seppänen, J.-T.; Forsman, J.T.; Mönkkönen, M.; Thomson, R.L. Social information use is a process across time, space, and ecology, reaching heterospecifics. Ecology 2007, 88, 1622–1633. [Google Scholar] [CrossRef]
- Laland, K.N.; Williams, K. Social transmission of maladaptive information in the guppy. Behav. Ecol. 1998, 9, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Giraldeau, L.-A.; Valone, T.J.; Templeton, J.J. Potential disadvantages of using socially acquired information. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 1559–1566. [Google Scholar] [CrossRef] [Green Version]
- Laland, K.N. Social learning strategies. Anim. Learn. Behav. 2004, 32, 4–14. [Google Scholar] [CrossRef]
- Grüter, C.; Leadbeater, E. Insights from insects about adaptive social information use. Trends Ecol. Evol. 2014, 29, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Kendal, R.L.; Boogert, N.J.; Rendell, L.; Laland, K.N.; Webster, M.; Jones, P.L. Social Learning Strategies: Bridge-Building between Fields. Trends Cogn. Sci. 2018, 22, 651–665. [Google Scholar] [CrossRef] [Green Version]
- Forsman, J.T.; Seppänen, J.-T. Learning what (not) to do: Testing rejection and copying of simulated heterospecific behavioural traits. Anim. Behav. 2011, 81, 879–883. [Google Scholar] [CrossRef]
- Loukola, O.J.; Seppänen, J.-T.; Krams, I.; Torvinen, S.S.; Forsman, J.T. Observed fitness may affect niche overlap in competing species via selective social information use. Am. Nat. 2013, 182, 474–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horner, V.; Proctor, D.; Bonnie, K.E.; Whiten, A.; de Waal, F.B.M. Prestige Affects Cultural Learning in Chimpanzees. PLoS ONE 2010, 5, e10625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, J.M.; Radford, A.N. Reduced social-information provision by immigrants and use by residents following dispersal. Curr. Biol. 2017, 27, R1266–R1267. [Google Scholar] [CrossRef] [PubMed]
- Nicol, C.J.; Pope, S.J. The effects of demonstrator social status and prior foraging success on social learning in laying hens. Anim. Behav. 1999, 57, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendal, R.; Hopper, L.M.; Whiten, A.; Brosnan, S.F.; Lambeth, S.P.; Schapiro, S.J.; Hoppitt, W. Chimpanzees copy dominant and knowledgeable individuals: Implications for cultural diversity. Evol. Hum. Behav. 2015, 36, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Choleris, E.; Guo, C.; Liu, H.; Mainardi, M.; Valsecchi, P. The effect of demonstrator age and number on duration of socially-induced food preferences in house mouse (Mus domesticus). Behav. Processes 1997, 41, 69–77. [Google Scholar] [CrossRef]
- Coelho, C.G.; Falótico, T.; Izar, P.; Mannu, M.; Resende, B.D.; Siqueira, J.O.; Ottoni, E.B. Social learning strategies for nut-cracking by tufted capuchin monkeys (Sapajus spp.). Anim. Cogn. 2015, 18, 911–919. [Google Scholar] [CrossRef]
- Duffy, G.A.; Pike, T.W.; Laland, K.N. Size-dependent directed social learning in nine-spined sticklebacks. Anim. Behav. 2009, 78, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Katz, M.; Lachlan, R.F. Social learning of food types in zebra finches (Taenopygia guttata) is directed by demonstrator sex and feeding activity. Anim. Cogn. 2003, 6, 11–16. [Google Scholar] [CrossRef]
- van de Waal, E.; Renevey, N.; Favre, C.M.; Bshary, R. Selective attention to philopatric models causes directed social learning in wild vervet monkeys. Proc. R. Soc. B Biol. Sci. 2010, 277, 2105–2111. [Google Scholar] [CrossRef] [PubMed]
- Coussi-Korbel, S.; Fragaszy, D.M. On the relation between social dynamics and social learning. Anim. Behav. 1995, 50, 1441–1453. [Google Scholar] [CrossRef]
- Schwab, C.; Bugnyar, T.; Schloegl, C.; Kotrschal, K. Enhanced social learning between siblings in common ravens, Corvus corax. Anim. Behav. 2008, 75, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyes, C.; Pearce, J.M. Not-so-social learning strategies. Proc. R. Soc. B Biol. Sci. 2015, 282, 20141709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.J.; Rand, A.S. Species Recognition and Sexual Selection as a Unitary Problem in Animal Communication. Evolution 1993, 47, 647–657. [Google Scholar] [CrossRef]
- Jaakkonen, T.; Kivelä, S.M.; Meier, C.M.; Forsman, J.T. The use and relative importance of intraspecific and interspecific social information in a bird community. Behav. Ecol. 2015, 26, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Avarguès-Weber, A.; Dawson, E.H.; Chittka, L. Mechanisms of social learning across species boundaries. J. Zool. 2013, 290, 1–11. [Google Scholar] [CrossRef]
- Parejo, D.; Avilés, J.M. Social information use by competitors: Resolving the enigma of species coexistence in animals? Ecosphere 2016, 7, e01295. [Google Scholar] [CrossRef]
- Leadbeater, E.; Dawson, E.H. A social insect perspective on the evolution of social learning mechanisms. Proc. Natl. Acad. Sci. USA 2017, 114, 7838–7845. [Google Scholar] [CrossRef] [Green Version]
- Giurfa, M. Social learning in insects: A higher-order capacity? Front. Behav. Neurosci. 2012, 6. [Google Scholar] [CrossRef] [Green Version]
- Leadbeater, E.; Chittka, L. A new mode of information transfer in foraging bumblebees? Curr. Biol. CB 2005, 15, R447–R448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, E.H.; Chittka, L. Conspecific and Heterospecific Information Use in Bumblebees. PLoS ONE 2012, 7, e31444. [Google Scholar] [CrossRef] [PubMed]
- Leadbeater, E.; Chittka, L. Social learning in insect—From miniature brains to consensus building. Curr. Biol. CB 2007, 17, R703–R713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, N.; Chittka, L. The importance of experience in the interpretation of conspecific chemical signals. Behav. Ecol. Sociobiol. 2006, 61, 215–220. [Google Scholar] [CrossRef]
- Leadbeater, E.; Chittka, L. Bumble-bees learn the value of social cues through experience. Biol. Lett. 2009, 5, 310–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, A.S.; Nielsen, M.E.; Dornhaus, A.; Papaj, D.R. Foraging Bumble Bees Weigh the Reliability of Personal and Social Information. Curr. Biol. 2016, 26, 1195–1199. [Google Scholar] [CrossRef] [Green Version]
- Smolla, M.; Alem, S.; Chittka, L.; Shultz, S. Copy-when-uncertain: Bumblebees rely on social information when rewards are highly variable. Biol. Lett. 2016, 12, 20160188. [Google Scholar] [CrossRef]
- Baude, M.; Dajoz, I.; Danchin, É. Inadvertent social information in foraging bumblebees: Effects of flower distribution and implications for pollination. Anim. Behav. 2008, 76, 1863–1873. [Google Scholar] [CrossRef]
- Baracchi, D.; Vasas, V.; Jamshed Iqbal, S.; Alem, S. Foraging bumblebees use social cues more when the task is difficult. Behav. Ecol. 2018, 29, 186–192. [Google Scholar] [CrossRef]
- Brian, A.D. Differences in the Flowers Visited by Four Species of Bumble-Bees and their Causes. J. Anim. Ecol. 1957, 26, 71–98. [Google Scholar] [CrossRef]
- Faegri, K.; Van Der Pijl, L. Principles of Pollination Ecology; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Morse, D.H. Foraging relationships within a guild of bumble bees. Insectes Sociaux 1982, 29, 445–454. [Google Scholar] [CrossRef]
- Kevan, P.G.; Baker, H.G. Insects as flower visitors and pollinators. Annu. Rev. Entomol. 1983, 28, 407–453. [Google Scholar] [CrossRef]
- Herrera, C.M. Pollinator abundance, morphology, and flower visitation rate: Analysis of the “quantity” component in a plant-pollinator system. Oecologia 1989, 80, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Goodale, E.; Beauchamp, G.; Magrath, R.D.; Nieh, J.C.; Ruxton, G.D. Interspecific information transfer influences animal community structure. Trends Ecol. Evol. 2010, 25, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Heyes, C. Who knows? Metacognitive social learning strategies. Trends Cogn. Sci. 2016, 20, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Bernays, E.A. Selective attention and host-plant specialization. Entomol. Exp. Appl. 1996, 80, 125–131. [Google Scholar] [CrossRef]
- Kamil, A.; Bond, A. Selective Attention, Priming, and Foraging Behavior. Pap. Behav. Biol. Sci. 2006. [Google Scholar]
- Morawetz, L.; Spaethe, J. Visual attention in a complex search task differs between honeybees and bumblebees. J. Exp. Biol. 2012, 215, 2515–2523. [Google Scholar] [CrossRef] [Green Version]
- Paulk, A.C.; Stacey, J.A.; Pearson, T.W.J.; Taylor, G.J.; Moore, R.J.D.; Srinivasan, M.V.; van Swinderen, B. Selective attention in the honeybee optic lobes precedes behavioral choices. Proc. Natl. Acad. Sci. USA 2014, 111, 5006–5011. [Google Scholar] [CrossRef] [Green Version]
- de Bivort, B.L.; van Swinderen, B. Evidence for selective attention in the insect brain. Curr. Opin. Insect Sci. 2016, 15, 9–15. [Google Scholar] [CrossRef]
- Heinrich, B. Bumblebees out in the cold. In The Hot-Blooded Insects; Springer: Berlin/Heidelberg, Germany, 1993; pp. 227–276. [Google Scholar]
- Leadbeater, E.; Chittka, L. The dynamics of social learning in an insect model, the bumblebee (Bombus terrestris). Behav. Ecol. Sociobiol. 2007, 61, 1789–1796. [Google Scholar] [CrossRef]
- Dawson, E.H.; Avarguès-Weber, A.; Chittka, L.; Leadbeater, E. Learning by Observation Emerges from Simple Associations in an Insect Model. Curr. Biol. 2013, 23, 727–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avarguès-Weber, A.; Chittka, L. Local enhancement or stimulus enhancement? Bumblebee social learning results in a specific pattern of flower preference. Anim. Behav. 2014, 97, 185–191. [Google Scholar] [CrossRef]
- Jones, P.L.; Ryan, M.J.; Chittka, L. The influence of past experience with flower reward quality on social learning in bumblebees. Anim. Behav. 2015, 101, 11–18. [Google Scholar] [CrossRef]
- Dyer, A.G.; Spaethe, J.; Prack, S. Comparative psychophysics of bumblebee and honeybee colour discrimination and object detection. J. Comp. Physiol. A 2008, 194, 617. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. Available online: https://www.R-project.org (accessed on 5 May 2018).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:14065823. [Google Scholar]
- Chittka, L.; Dyer, A.G.; Bock, F.; Dornhaus, A. Bees trade off foraging speed for accuracy. Nature 2003, 424, 388. [Google Scholar] [CrossRef]
- Ings, T.C.; Chittka, L. Speed-Accuracy Tradeoffs and False Alarms in Bee Responses to Cryptic Predators. Curr. Biol. 2008, 18, 1520–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, B. Resource heterogeneity and patterns of movement in foraging bumblebees. Oecologia 1979, 40, 235–245. [Google Scholar] [CrossRef]
- Chittka, L.; Thomson, J.D.; Waser, N.M. Flower Constancy, Insect Psychology, and Plant Evolution. Naturwissenschaften 1999, 86, 361–377. [Google Scholar] [CrossRef]
- Loukola, O.J.; Gatto, E.; Híjar-Islas, A.C.; Chittka, L. Selective interspecific information use in the nest choice of solitary bees. Anim. Biol. 2020, 70, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Pearce, R.F.; Giuggioli, L.; Rands, S.A. Bumblebees can discriminate between scent-marks deposited by conspecifics. Sci. Rep. 2017, 7, 43872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieh, J.C.; Barreto, L.S.; Contrera, F.A.; Imperatriz–Fonseca, V.L. Olfactory eavesdropping by a competitively foraging stingless bee, Trigona spinipes. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 1633–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alem, S.; Perry, C.J.; Zhu, X.; Loukola, O.J.; Ingraham, T.; Søvik, E.; Chittka, L. Associative Mechanisms Allow for Social Learning and Cultural Transmission of String Pulling in an Insect. PLoS Biol. 2016, 14, e1002564. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-González, J.E.; Royka, A.L.; MaBouDi, H.; Solvi, C.; Seppänen, J.-T.; Loukola, O.J. Foraging Bumblebees Selectively Attend to Other Types of Bees Based on Their Reward-Predictive Value. Insects 2020, 11, 800. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110800
Romero-González JE, Royka AL, MaBouDi H, Solvi C, Seppänen J-T, Loukola OJ. Foraging Bumblebees Selectively Attend to Other Types of Bees Based on Their Reward-Predictive Value. Insects. 2020; 11(11):800. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110800
Chicago/Turabian StyleRomero-González, Jose E., Amanda L. Royka, HaDi MaBouDi, Cwyn Solvi, Janne-Tuomas Seppänen, and Olli J. Loukola. 2020. "Foraging Bumblebees Selectively Attend to Other Types of Bees Based on Their Reward-Predictive Value" Insects 11, no. 11: 800. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110800