Unexpected Species Identities and Interspecific Relationships in a Subterranean Beetle Lineage, the Pterostichus macrogenys Species Group (Coleoptera, Carabidae), Revealed by Fine-Scale Field Sampling and Detailed Morphological Comparisons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
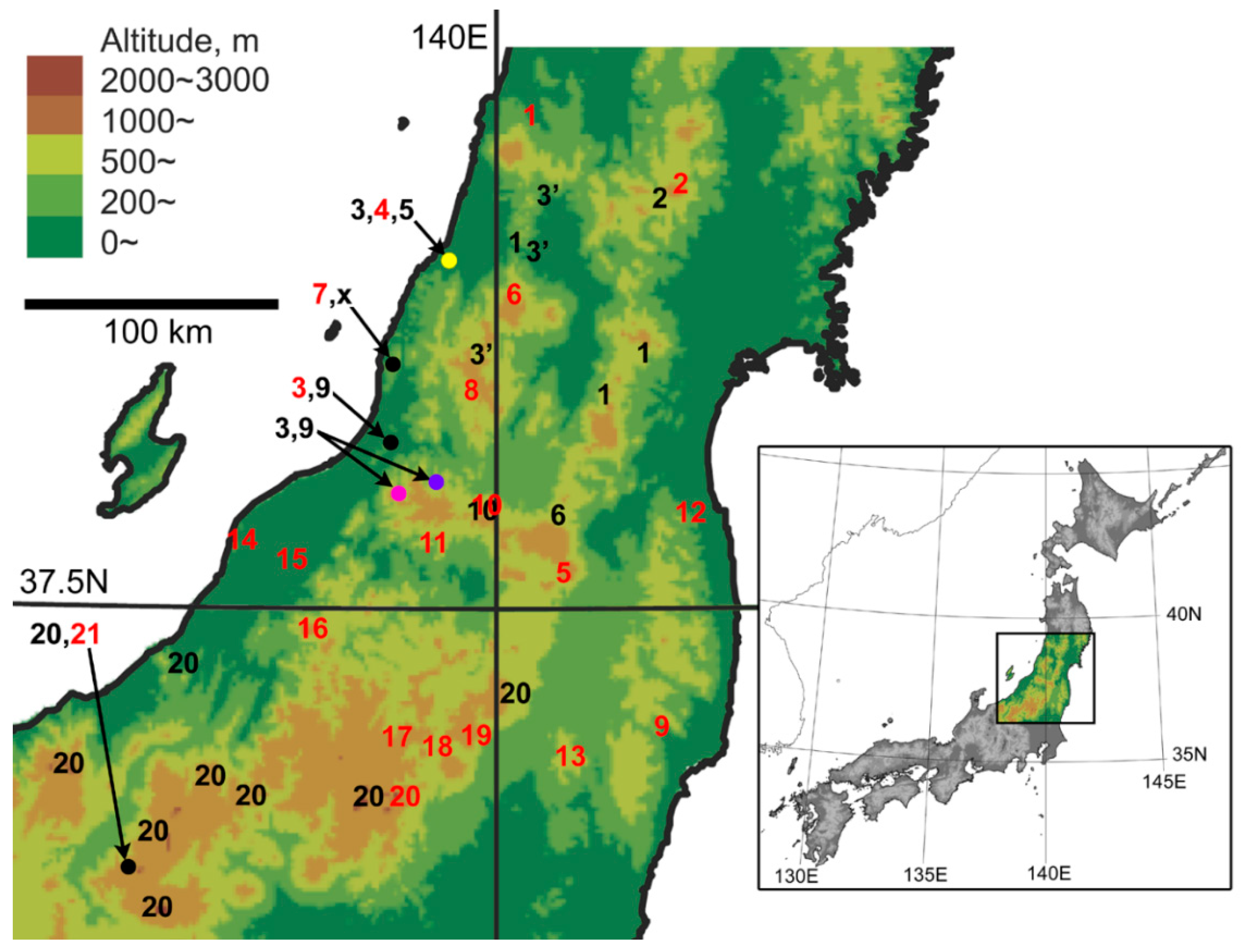
2. Materials and Methods
3. Results
3.1. Overview of Species Identification, Distribution, and Coexistence
3.2. Collection Records and Systematics
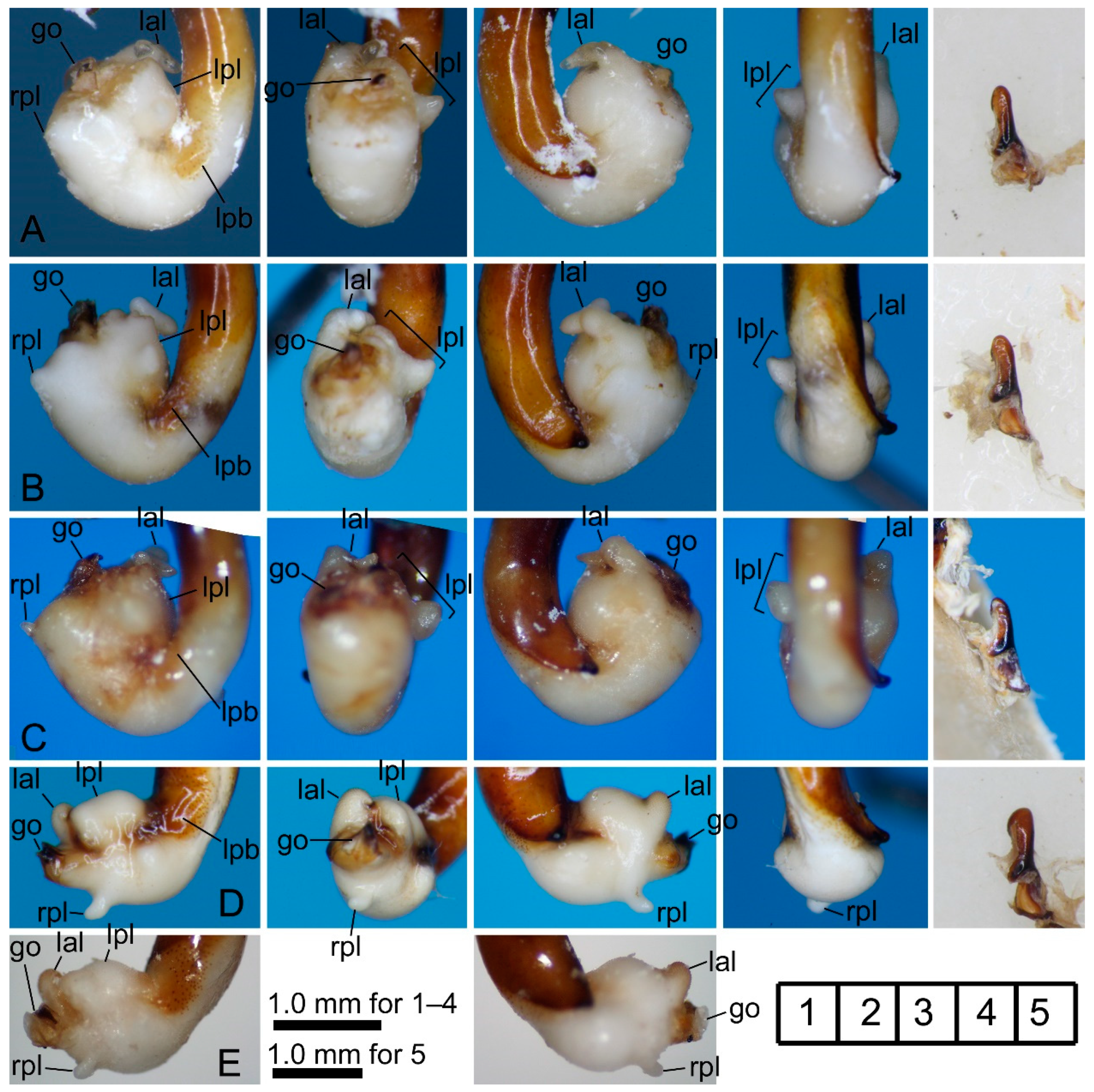
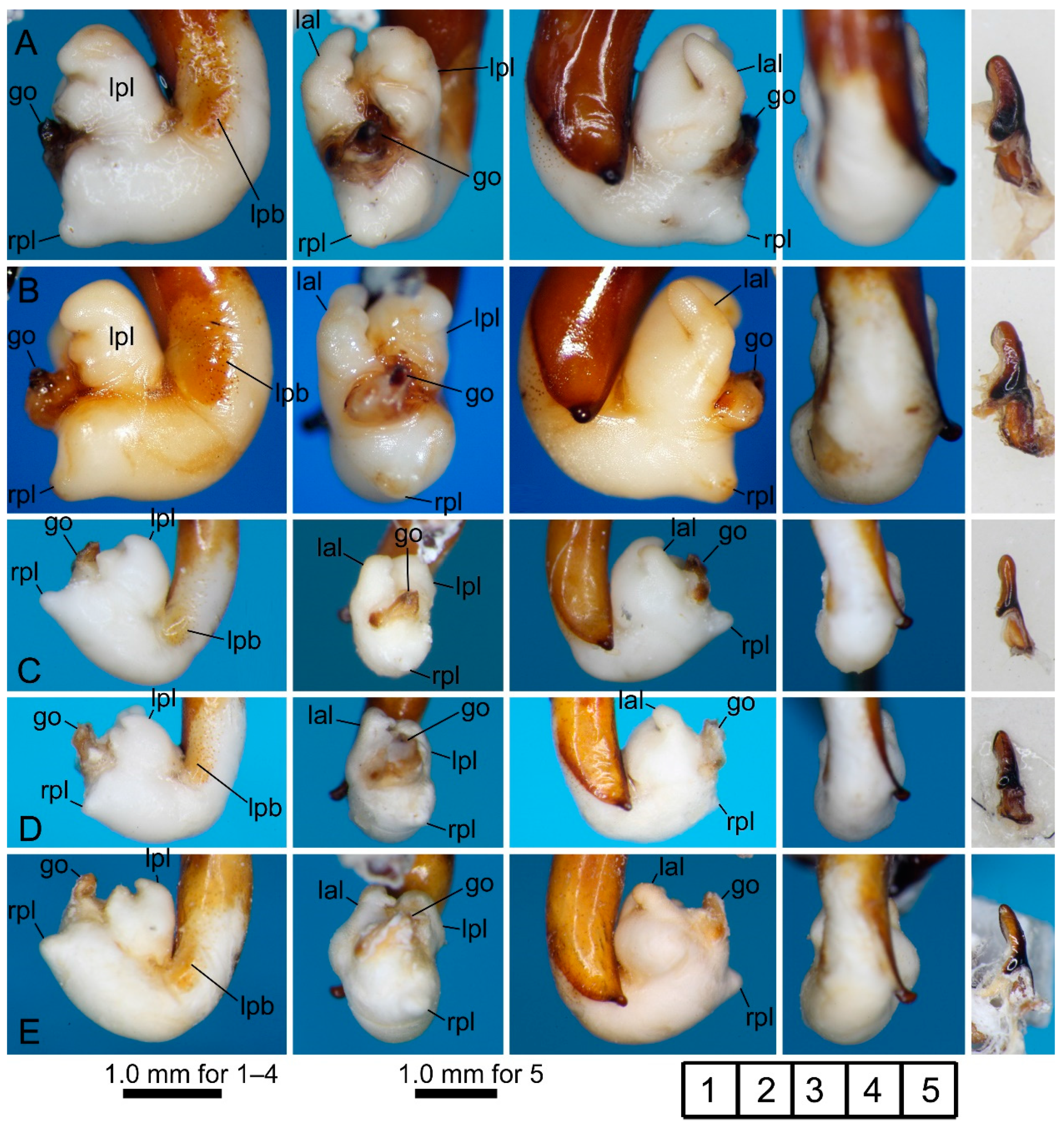
3.2.1. Pterostichus (Nialoe) chokaisanus Sasakawa, 2009
3.2.2. Pterostichus (Nialoe) kurikomasanus Sasakawa, 2005
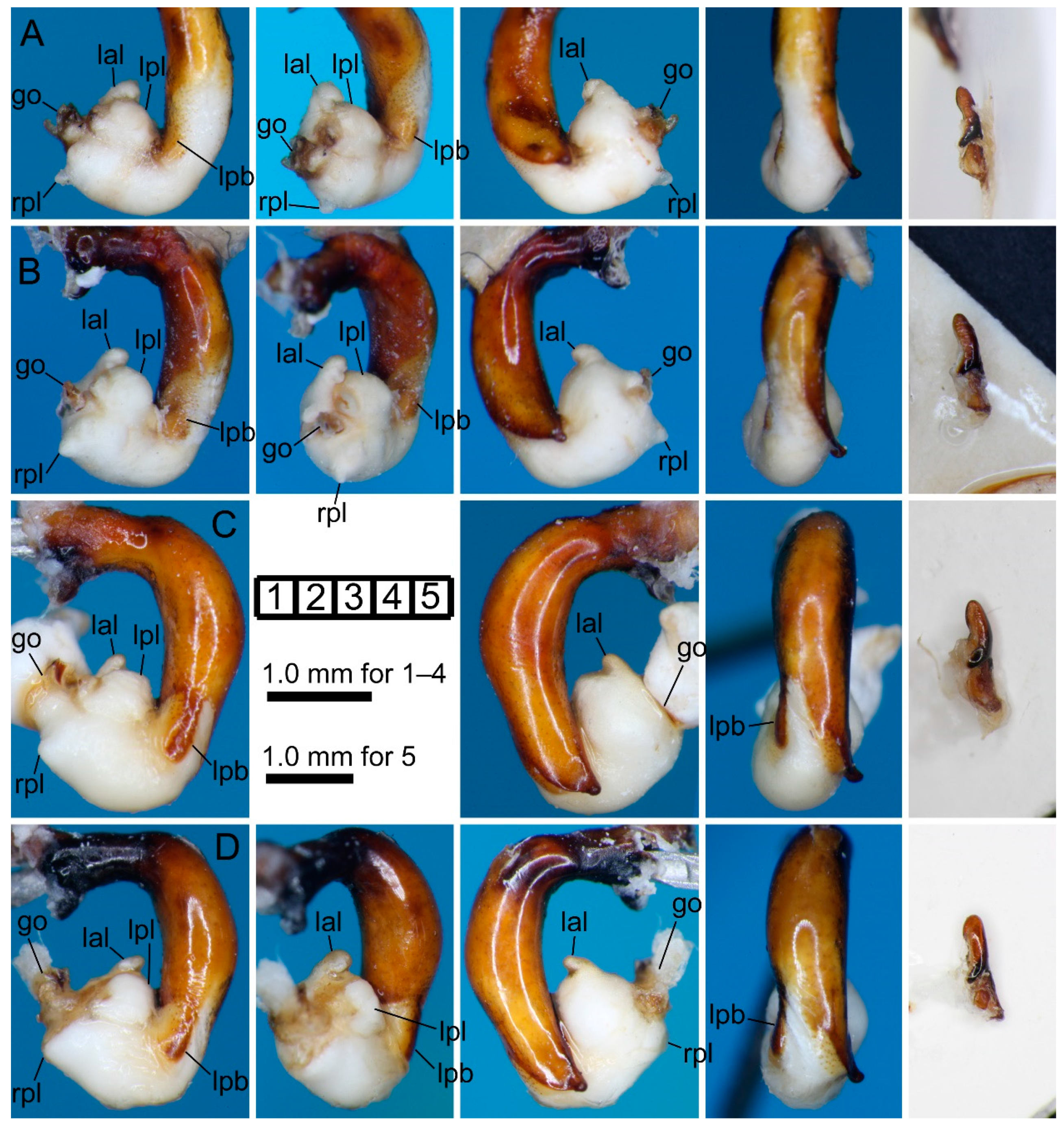
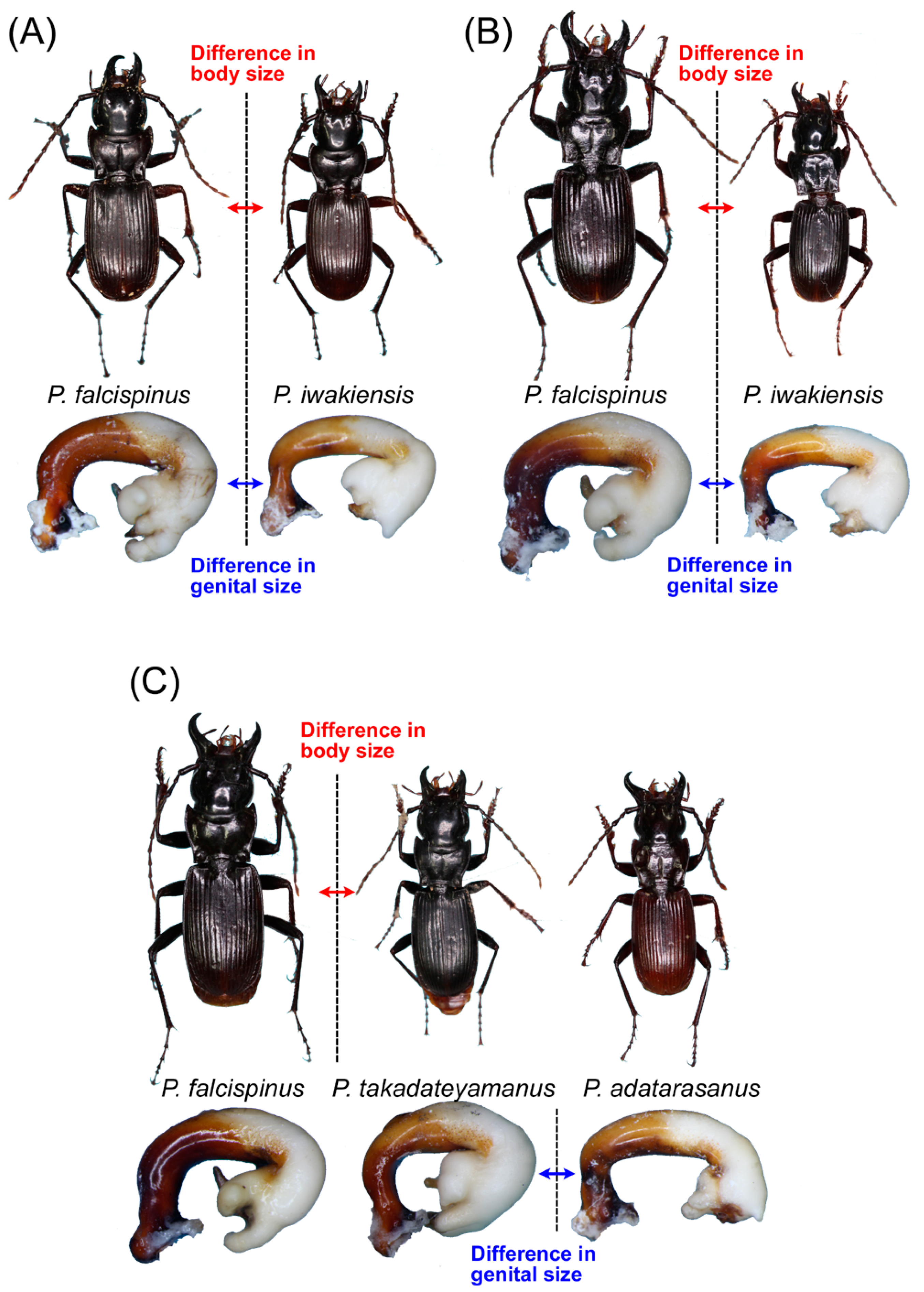
3.2.3. Pterostichus (Nialoe) falcispinus Sasakawa, 2005
3.2.4. Pterostichus (Nialoe) takadateyamanus Sasakawa, 2009
3.2.5. Pterostichus (Nialoe) adatarasanus Sasakawa, 2005
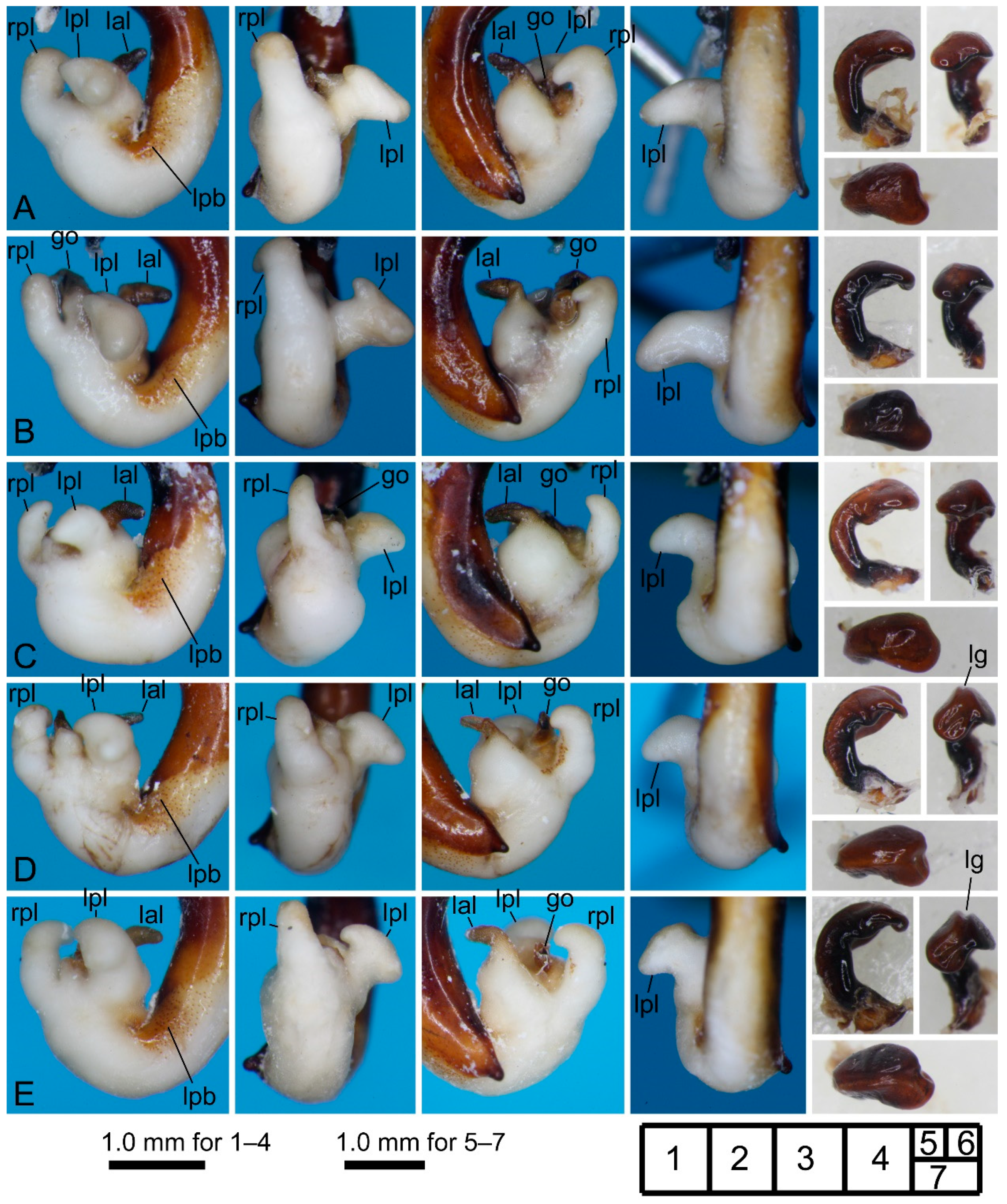
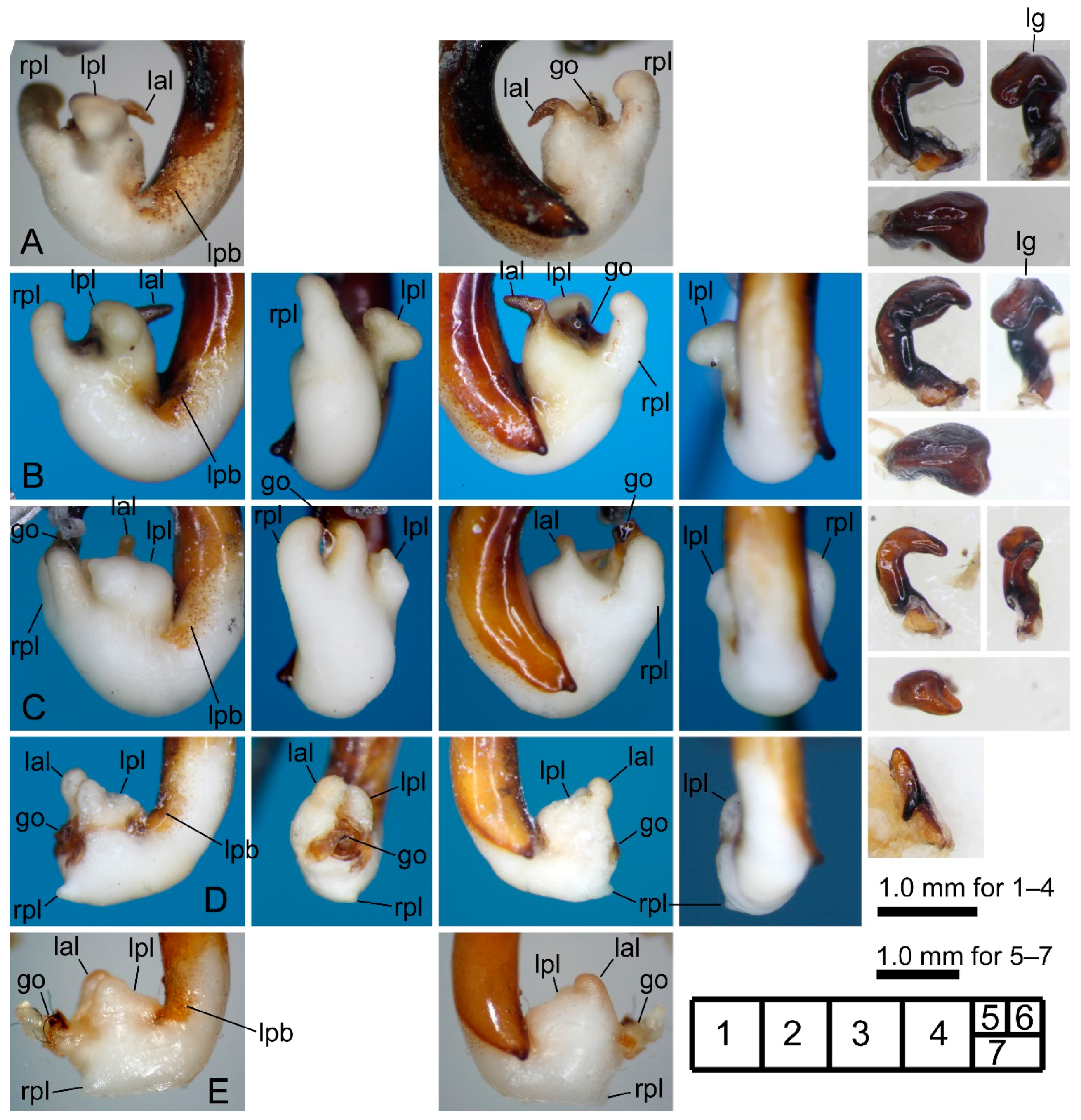
3.2.6. Pterostichus (Nialoe) gassanus Sasakawa, 2009
3.2.7. Pterostichus (Nialoe) iwakiensis Sasakawa, 2009
3.2.8. Pterostichus (Nialoe) eboshiyamanus Sasakawa, 2009
3.2.9. Pterostichus (Nialoe) monolineatus sp. n.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Culver, D.C.; Pipan, T. Insects in Caves. In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; Willey-Blackwell: Chichester, UK, 2018; Volume 3, pp. 123–152. [Google Scholar]
- Faille, A.; Ribera, I.; Deharveng, L.; Bourdeau, C.; Garnery, L.; Quéinnec, E.; Deuve, T. A molecular phylogeny shows the single origin of the Pyrenean subterranean Trechini ground beetles (Coleoptera: Carabidae). Mol. Phylogenet. Evol. 2010, 54, 97–1106. [Google Scholar] [CrossRef] [PubMed]
- Boyd, O.F.; Philips, T.K.; Johnson, J.R.; Nixon, J.J. Geographically structured genetic diversity in the cave beetle Darlingtonea kentuckensis Valentine, 1952 (Coleoptera, Carabidae, Trechini, Trechina). Subterr. Biol. 2020, 34, 1–23. [Google Scholar] [CrossRef]
- Balke, M.; Ribera, I. A subterranean species of Exocelina diving beetle from the Malay Peninsula filling a 4,000 km distribution gap between Melanesia and southern China. Subterr. Biol. 2020, 34, 25–37. [Google Scholar] [CrossRef]
- Sánchez-Fernández, D.; Rizzo, V.; Bourdeau, C.; Cieslak, A.; Comas, J.; Faille, A.; Fresneda, J.; Lleopart, E.; Millán, A.; Montes, A.; et al. The deep subterranean environment as a potential model system in ecological, biogeographical and evolutionary research. Subterr. Biol. 2018, 25, 1–7. [Google Scholar] [CrossRef]
- Mammola, S.; Isaia, M. The ecological niche of a specialized subterranean spider. Invertebr. Biol. 2016, 135, 20–30. [Google Scholar] [CrossRef]
- Ortuño, V.M.; Gilgado, J.D.; Jiménez-Valverde, A.; Sendra, A.; Pérez-Suárez, G.; Herrero-Borgoñón, J.J. The “alluvial mesovoid shallow substratum”, a new subterranean habitat. PLoS ONE 2013, 8, e76311. [Google Scholar] [CrossRef] [Green Version]
- Sugaya, K.; Yamasako, J. A new species of the genus Ishikawatrechus (Coleoptera, Trechinae) from Japan. Zootaxa 2008, 3768, 189–195. [Google Scholar] [CrossRef]
- Sasakawa, K.; Kubota, K. Phylogeny of ground beetles subgenus Nialoe (s. lat.) Tanaka (Coleoptera: Carabidae; genus Pterostichus): A molecular phylogenetic approach. Entomol. Sci. 2009, 12, 308–313. [Google Scholar] [CrossRef]
- Sasakawa, K.; Itô, H. Two new species of the Pterostichus macrogenys species group (Coleoptera, Carabidae) discovered in shallow subterranean habitats in northern Honshu, Japan. Subterr. Biol. 2017, 21, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Sasakawa, K. A new species of ground beetle and a revised list of the East Asian endemic subgenus Nialoe (s. lat.) (Coleoptera: Carabidae: Pterostichus). Orient. Insects 2020. [Google Scholar] [CrossRef]
- Habu, A.; Baba, K. A new subspecies of Pterostichus macrogenys Bates from Niigata Prefecture, Japan. Akitu 1960, 9, 62. [Google Scholar]
- Habu, A. On two Pterostichus species found in caves in Niigata Prefecture, Japan (Coleoptera, Carabidae). Entomol. Rev. Jpn. 1977, 30, 13–17. [Google Scholar]
- Nakane, T. Harpalidae. In Iconographia Insectorum Japonicorum Colore Naturali Edita; Nakane, T., Ohbayashi, K., Nomura, S., Kurosawa, Y., Eds.; Hokuryukan: Tokyo, Japan, 1963; Volume 2, pp. 22–54. [Google Scholar]
- Tanaka, K. Pterostichinae. In The Coleoptera of Japan in Color; Uéno, S., Kurosawa, Y., Satô, M., Eds.; Hoikusha: Osaka, Japan, 1985; Volume 2, pp. 105–135. [Google Scholar]
- Ito, N.; Ogai, T. A new species of macrocephalic carabid from Nagano Prefecture, Japan (Coleoptera: Carabidae: Pterostichini). Jpn. J. Syst. Entomol. 2015, 21, 271–275. [Google Scholar]
- Sasakawa, K. Pterostichus macrogenys Bates, 1883 (Coleoptera, Carabidae) and its allied species of northern Japan. Biogeography 2005, 7, 69–78. [Google Scholar]
- Sasakawa, K. Pterostichus macrogenys Bates (Coleoptera: Carabidae) and its allied species of northern Japan: Descriptions of seven additional species and possible evidence supporting species status. Zool. Stud. 2009, 48, 262–269. [Google Scholar]
- Yoshida, M. How to collect insects living in upper hypogean by using underground traps. Nat. Insects 2012, 47, 6–12. (In Japanese) [Google Scholar]
- Berlov, O. Preparati permanenti a secco dell’endofallo nel genere Carabus, L. (Coleoptera, Carabidae). Boll. Soc. Entomol. Ital. 1992, 124, 141–143. [Google Scholar]
- Rasband, W.S. ImageJ, Version 1.50i. 2016. Available online: https://imagej.nih.gov/ij/ (accessed on 16 February 2018).
- Sasakawa, K. Taxonomic studies of the ground beetle subgenus Falcinebria Ledoux & Roux, 2005. (Coleoptera, Carabidae, Nebria) from Honshû, Japan. ZooKeys 2020, 902, 37–60. [Google Scholar]
- Morita, S.; Hirasawa, H. Macrocephalic pterostichines (Coleoptera, Carabidae) from central Honshu, Japan. Elytra 1996, 24, 21–30. [Google Scholar]
- Morita, S.; Kurosa, Y. A new macrocephalic species of the genus Pterostichus (Coleoptera, Carabidae). Elytra 1998, 24, 69–73. [Google Scholar]
- Sugimura, A. A new species of the genus Pterostichus (Coleoptera, Carabidae) from Mt. Fukube-ga-take, Chûnô District of Gifu Prefecture, central Japan. Elytra 2005, 33, 641–647. [Google Scholar]
- Sugimura, A. A new species of the genus Pterostichus (Coleoptera, Carabidae) from Mt. Shirakusa-yama in Gero-shi of Gifu Prefecture, central Japan. Elytra 2006, 34, 33–39. [Google Scholar]
- Sugimura, A. A new species of the genus Pterostichus (Coleoptera, Carabidae) from the Neo-dani, Seinô District of Gifu Prefecture, central Japan. Elytra 2007, 35, 279–286. [Google Scholar]
- Morita, S.; Ohkawa, H. A new Pterostichus (Coleoptera, Carabidae) from Gifu Prefecture, central Japan. Elytra 2010, 38, 99–103. [Google Scholar]
- Toda, N. Two new macrocephalic pterostichines (Coleoptera, Carabidae) from central Japan. Elytra New Ser. 2012, 2, 119–125. [Google Scholar]
- Morita, S.; Ohkawa, H.; Kurihara, T. Two new macrocephalic pterostichine carabids (Coleoptera, Carabidae) from central Japan. Elytra New Ser. 2013, 3, 9–17. [Google Scholar]
- Sota, T.; Vogler, A.P. Reconstructing species phylogeny of the carabid beetles Ohomopterus using multiple nuclear DNA sequences: Heterogeneous information content and the performance of simultaneous analyses. Mol. Phylogenet. Evol. 2003, 26, 139–154. [Google Scholar] [CrossRef]
- Fujisawa, T.; Sasabe, M.; Nagata, N.; Takami, Y.; Sota, T. Genetic basis of species-specific genitalia reveals role in species diversification. Sci. Adv. 2019, 5, eaav9939. [Google Scholar] [CrossRef] [Green Version]
- Sota, T.; Takami, Y.; Kubota, K.; Ujiie, M.; Ishikawa, R. Interspecific body size differentiation in species assemblages of the carabid subgenus Ohomopterus in Japan. Popul. Ecol. 2000, 42, 279–291. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasakawa, K.; Mitsuduka, Y.; Itô, H. Unexpected Species Identities and Interspecific Relationships in a Subterranean Beetle Lineage, the Pterostichus macrogenys Species Group (Coleoptera, Carabidae), Revealed by Fine-Scale Field Sampling and Detailed Morphological Comparisons. Insects 2020, 11, 803. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110803
Sasakawa K, Mitsuduka Y, Itô H. Unexpected Species Identities and Interspecific Relationships in a Subterranean Beetle Lineage, the Pterostichus macrogenys Species Group (Coleoptera, Carabidae), Revealed by Fine-Scale Field Sampling and Detailed Morphological Comparisons. Insects. 2020; 11(11):803. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110803
Chicago/Turabian StyleSasakawa, Kôji, Yoshiji Mitsuduka, and Hirotarô Itô. 2020. "Unexpected Species Identities and Interspecific Relationships in a Subterranean Beetle Lineage, the Pterostichus macrogenys Species Group (Coleoptera, Carabidae), Revealed by Fine-Scale Field Sampling and Detailed Morphological Comparisons" Insects 11, no. 11: 803. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110803