Positive Correlation between Pesticide Consumption and Longevity in Solitary Bees: Are We Overlooking Fitness Trade-Offs?

 , and
, and
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Pesticide Solutions
2.3. Endpoint Measurements
2.4. Statistical Analysis
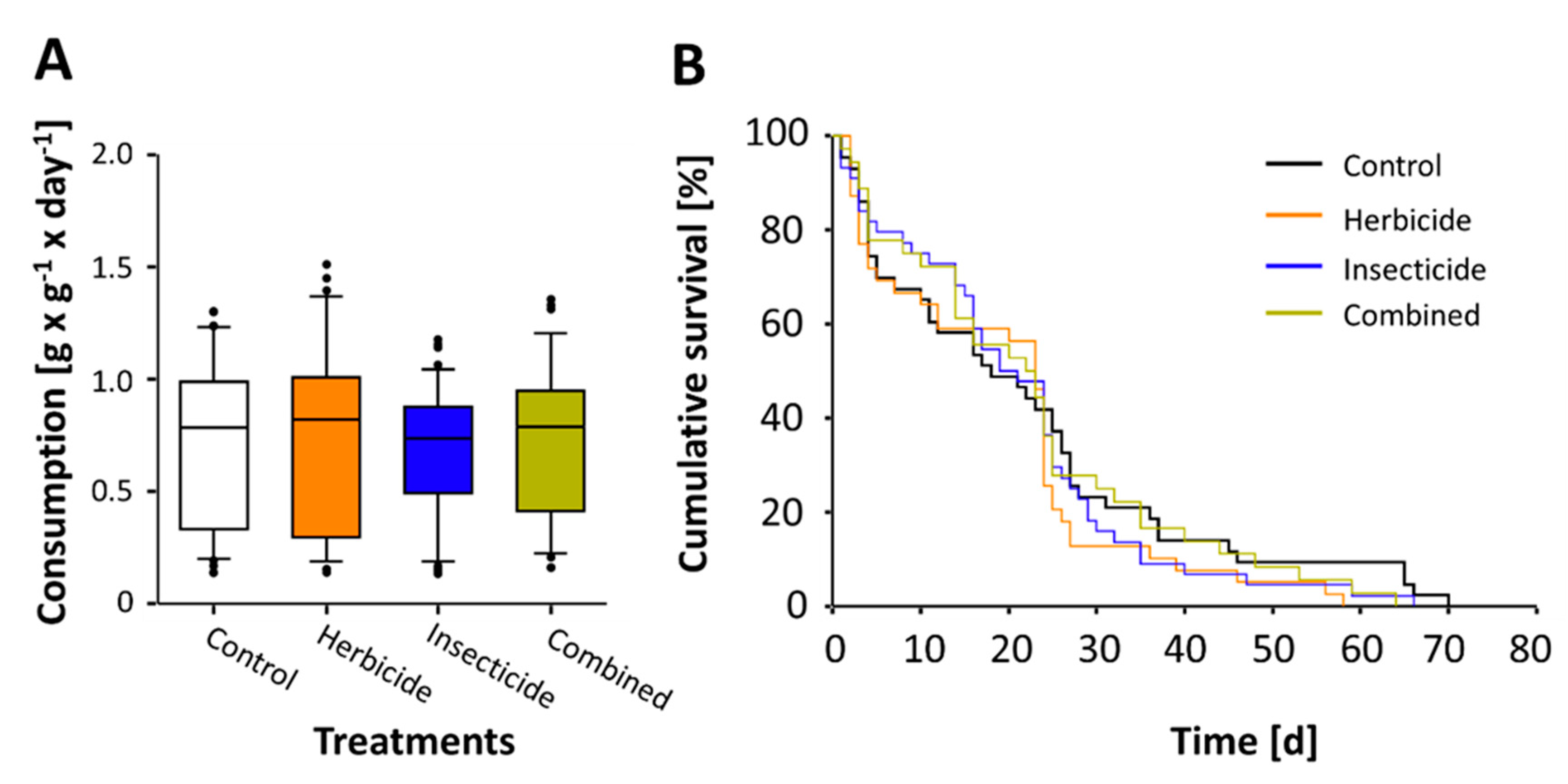
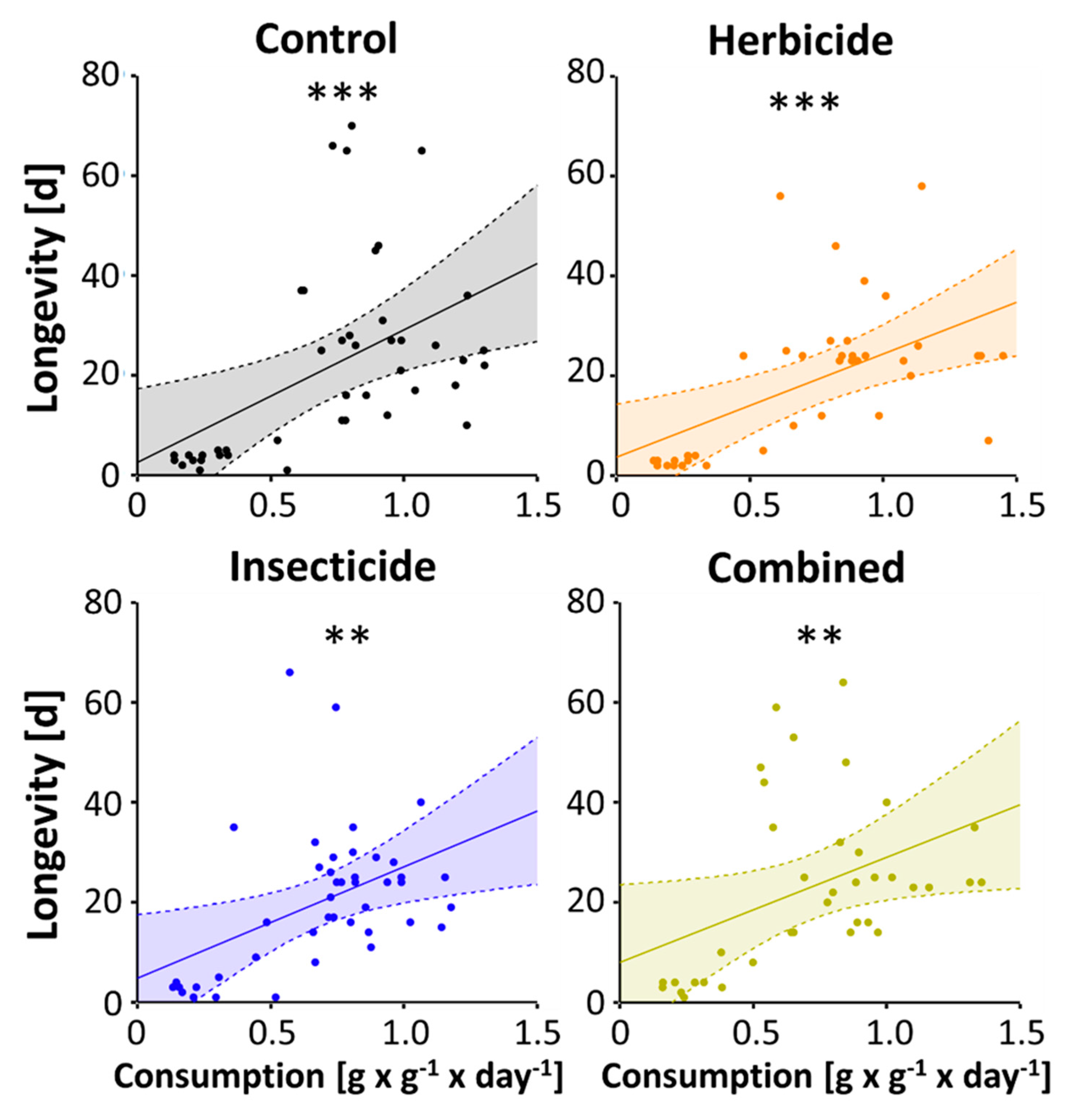
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Topping, C.J.; Aldrich, A.; Berny, P. Overhaul environmental risk assessment for pesticides. Science 2020, 367, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Frouz, J.; Helenius, J.; Sarthou, J.; Klein, A.; Genersch, E.; Kovács-Hostyánszki, A.; Samu, F.; Stout, J.; Pennacchio, F.; et al. Ecosystem Services, Agriculture and Neonicotinoids; EASAC policy report 26; German National Academy of Science: Halle, Germany, 2015; ISBN 9783804728974. [Google Scholar]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.; Strobl, V.; Neumann, P. The need for an evolutionary approach to ecotoxicology. Nat. Ecol. Evol. 2020, 4, 895. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [Green Version]
- Sandrock, C.; Tanadini, L.G.; Pettis, J.S.; Biesmeijer, J.C.; Potts, S.G.; Neumann, P. Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agric. For. Entomol. 2014, 16, 119–128. [Google Scholar] [CrossRef]
- Stuligross, C.; Williams, N.M. Pesticide and resource stressors additively impair wild bee reproduction. Proc. R. Soc. B 2020, 287. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goñalons, C.M.; Farina, W.M. Impaired associative learning after chronic exposure to pesticides in young adult honey bees. J. Exp. Biol. 2018, 221, jeb176644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid insecticides: Molecular targets, resistance, and toxicity. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Mogren, C.; Lundgren, J. Neonicotinoid-contaminated pollinator strips adjacent to cropland. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Humann-Guilleminot, S.; Binkowski, Ł.J.; Jenni, L.; Hilke, G.; Glauser, G.; Helfenstein, F. A nation-wide survey of neonicotinoid insecticides in agricultural land with implications for agri-environment schemes. J. Appl. Ecol. 2019, 56, 1502–1514. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [Green Version]
- Straub, L.; Villamar-Bouza, L.; Bruckner, S.; Chantawannakul, P.; Gauthier, L.; Khongphinitbunjong, K.; Retschnig, G.; Troxler, A.; Vidondo, B.; Neumann, P.; et al. Neonicotinoid insecticides can serve as inadvertent insect contraceptives. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160506. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent—Relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate. EFSA J. 2015, 13, 4302. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Raymann, K.; Moran, N.A. Glyphosate perturbs the gut microbiota of honey bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seide, V.E.; Bernardes, R.C.; Pereira, E.J.G.; Lima, M.A.P. Glyphosate is lethal and Cry toxins alter the development of the stingless bee Melipona quadrifasciata. Environ. Pollut. 2018, 243, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Farina, W.M.; Balbuena, M.S.; Herbert, L.T.; Goñalons, C.M.; Vázquez, D.E. Effects of the herbicide glyphosate on honey bee sensory and cognitive abilities: Individual impairments with implications for the hive. Insects 2019, 10, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez, D.E.; Balbuena, M.S.; Chaves, F.; Gora, J.; Menzel, R.; Farina, W.M. Sleep in honey bees is affected by the herbicide glyphosate. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Kessler, S.C.; Tiedeken, E.J.; Simcock, K.L.; Derveau, S.; Mitchell, J.; Softley, S.; Stout, J.C.; Wright, G.A. Bees prefer foods containing neonicotinoid pesticides. Nature 2015, 521, 74–76. [Google Scholar] [CrossRef]
- Sgolastra, F.; Arnan, X.; Cabbri, R.; Isani, G.; Medrzycki, P.; Teper, D.; Bosch, J.; Mediche, S. Combined exposure to sublethal concentrations of an insecticide and a fungicide affect feeding, ovary development and longevity in a solitary bee. Proc. R. Soc. B 2018, 285, 1–8. [Google Scholar] [CrossRef]
- Castañeda, L.E.; Figueroa, C.C.; Fuentes-Contreras, E.; Niemeyer, H.M.; Nespolo, R.F. Energetic costs of detoxification systems in herbivores feeding on chemically defended host plants: A correlational study in the grain aphid, Sitobion avenae. J. Exp. Biol. 2009, 212, 1185–1190. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, B.C.; Verhulst, S. Ecological immunology: Costy parasite defences and trade-offs in evolutionary ecology. Trends Ecol. Evol. 1996, 22, 317–321. [Google Scholar] [CrossRef]
- Moret, Y.; Schmid-Hempel, P. Survival for immunity: The price of immune system activation for bumblebee workers. Science 2000, 290, 1166–1168. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Cutler, G.C. Insecticide-induced hormesis and arthropod pest management. Pest Manag. Sci. 2013, 70, 690–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azpiazu, C.; Bosch, J.; Viñuela, E.; Medrzycki, P.; Teper, D.; Sgolastra, F. Chronic oral exposure to field-realistic pesticide combinations via pollen and nectar: Effects on feeding and thermal performance in a solitary bee. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Heard, M.S.; Baas, J.; Dorne, J.L.; Lahive, E.; Robinson, A.G.; Rortais, A.; Spurgeon, D.J.; Svendsen, C.; Hesketh, H. Comparative toxicity of pesticides and environmental contaminants in bees: Are honey bees a useful proxy for wild bee species? Sci. Total Environ. 2017, 578, 357–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westrich, P. Die Wildbienen Baden-Württembergs; Eugen Ulmer Verlag: Stuttgart, Germany, 1989. [Google Scholar]
- Straub, L.; Williams, G.R.G.R.; Pettis, J.; Fries, I.; Neumann, P. Superorganism resilience: Eusociality and susceptibility of ecosystem service providing insects to stressors. Curr. Opin. Insect Sci. 2015, 12, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Colgan, T.J.; Fletcher, I.K.; Arce, A.N.; Gill, R.J.; Rodrigues, A.R.; Stolle, E.; Chittka, L.; Wurm, Y. Caste- and pesticide-specific effects of neonicotinoid pesticide exposure on gene expression in bumblebees. Mol. Ecol. 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schläppi, D.; Kettler, N.; Straub, L.; Glauser, G.; Neumann, P. Long-term effects of neonicotinoid insecticides on ants. Commun. Biol. 2020, 3, 1–9. [Google Scholar] [CrossRef]
- EFSA. Guidance on the risk assessment of plant protection products on bees (Apis mellifera, Bombus spp. and solitary bees). EFSA J. 2014, 11, 268. [Google Scholar] [CrossRef]
- EFSA. Outline of the revision of the Guidance on the risk assessment of plant protection products on bees (Apis mellifera, Bombus spp. and solitary bees) (EFSA, 2013). Eur. Food Saf. Auth. 2020, 11, 1–6. Available online: https://www.efsa.europa.eu (accessed on 26 October 2020).
- Torchio, P.F.; Asensio, E.; Thorp, R.W. Introduction of the European bee, Osmia cornuta, into California almond orchards (Hymenoptera: Megachilidae). Environ. Entomol. 1987, 16, 664–667. [Google Scholar] [CrossRef]
- Sgolastra, F.; Medrzycki, P.; Bortolotti, L.; Renzi, M.T.; Tosi, S.; Bogo, G.; Teper, D.; Porrini, C.; Molowny-Horas, R.; Bosch, J. Synergistic mortality between a neonicotinoid insecticide and an ergosterol-biosynthesis-inhibiting fungicide in three bee species. Pest Manag. Sci. 2017, 73, 1236–1243. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Effects of fungicide and herbicide chemical exposure on Apis and non-Apis bees in agricultural landscape. Front. Environ. Sci. 2020, 8, 1–10. [Google Scholar] [CrossRef]
- Strobl, V.; Straub, L.; Bruckner, S.; Albrecht, M.; Maitip, J.; Kolari, E.; Chantawannakul, P.; Williams, G.R.; Neumann, P. Not every sperm counts: Male fertility in solitary bees, Osmia cornuta. PLoS ONE 2019, 14, e0214597. [Google Scholar] [CrossRef] [PubMed]
- Torchio, P.F.; Asensio, E. The introduction of the European bee, Osmia cornuta Latr., into the U.S. as a potential pollinator of orchard crops, and a comparison of its manageability with Osmia lignaria propinqua Cresson (Hymenoptera: Megachilidae). J. Kansas Entomol. Soc. 1985, 58, 42–52. [Google Scholar]
- Nicholls, E.; Fowler, R.; Niven, J.E.; Gilbert, J.D.; Goulson, D. Larval exposure to field-realistic concentrations of clothianidin has no effect on development rate, over-winter survival or adult metabolic rate in a solitary bee, Osmia bicornis. PeerJ 2017, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assalin, M.R.; de Moraes, S.G.; Queiroz, S.C.N.; Ferracini, V.L.; Duran, N. Studies on degradation of glyphosate by several oxidative chemical processes: Ozonation, photolysis and heterogeneous photocatalysis. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2010, 45, 89–94. [Google Scholar] [CrossRef]
- Rubio, F.; Guo, E.; Kamp, L. Survey of glyphosate residues in honey, corn and soy products. J. Environ. Anal. Toxicol. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Botias, C.; David, A.; Horwood, J.; Abdul-Sada, A.; Nicholls, E.; Hill, E.; Goulson, D. Neonicotinoid residues in wildflowers, a potential route of chronic exposure for bees. Environ. Sci. Technol. 2015, 49, 12731–12740. [Google Scholar] [CrossRef]
- Ladurner, E.; Bosch, J.B.; Kemp, W.P.; Maini, S. Evaluation of a standard artificial flower design to feed individual bees known amounts of pesticides. Apidologie 2005, 36, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Harshman, L.G.; Zera, A.J. The cost of reproduction: The devil in the details. Trends Ecol. Evol. 2007, 22, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Isaac, N.J.B.; Bullock, J.M.; Roy, D.B.; Garthwaite, D.G.; Crowe, A.; Pywell, R.F. Impacts of neonicotinoid use on long-term population changes in wild bees in England. Nat. Commun. 2016, 7, 12459. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmehl, D.R.; Teal, P.E.A.; Frazier, J.L.; Grozinger, C.M. Genomic analysis of the interaction between pesticide exposure and nutrition in honey bees (Apis mellifera). J. Insect Physiol. 2014, 71, 177–190. [Google Scholar] [CrossRef]
- Dickel, F.; Münch, D.; Amdam, G.V.; Mappes, J.; Freitak, D. Increased survival of honeybees in the laboratory after simultaneous exposure to low doses of pesticides and bacteria. PLoS ONE 2018, 13, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Turturro, A.; Hass, B.S.; Hart, R.W. Does caloric restriction induce hormesis? Hum. Exp. Toxicol. 2000, 19, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Arena, M.; Sgolastra, F. A meta-analysis comparing the sensitivity of bees to pesticides. Ecotoxicology 2014, 23, 324–334. [Google Scholar] [CrossRef]
- Baron, G.L.; Raine, N.E.; Brown, M.J.F. General and species-specific impacts of a neonicotinoid insecticide on the ovary development and feeding of wild bumblebee queens. Proc. R. Soc. B 2017, 284, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Siva-Jothy, M.T.; Tsubaki, Y.; Hooper, R.E. Decreased immune response as a proximate cost of copulation and oviposition in a damselfly. Physiol. Entomol. 1998, 23, 274–277. [Google Scholar] [CrossRef]
- Du Rand, E.E.; Smit, S.; Beukes, M.; Apostolides, Z.; Pirk, C.W.W.; Nicolson, S.W. Detoxification mechanisms of honey bees (Apis mellifera) resulting in tolerance of dietary nicotine. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–immunity trade-offs in insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berenbaum, M.R.; Johnson, R.M. Xenobiotic detoxification pathways in honey bees. Curr. Opin. Insect Sci. 2015, 10, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beadle, K.; Singh, K.S.; Troczka, B.J.; Randall, E.; Zaworra, M.; Zimmer, C.T.; Hayward, A.; Reid, R.; Kor, L.; Kohler, M.; et al. Genomic insights into neonicotinoid sensitivity in the solitary bee Osmia bicornis. PLoS Genet. 2019, 15, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinkevich, F.D.; Margotta, J.W.; Pittman, J.M.; Danka, R.G.; Tarver, M.R.; Ottea, J.A.; Healy, K.B. Genetics, synergists, and age affect insecticide sensitivity of the honey bee, Apis mellifera. PLoS ONE 2015, 10, e0139841. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Benbrook, C.; Antoniou, M.N. Insight into the confusion over surfactant co-formulants in glyphosate-based herbicides. Food Chem. Toxicol. 2019, 128, 137–145. [Google Scholar] [CrossRef]
- Cutler, G.C.; Rix, R.R. Can poisons stimulate bees? Appreciating the potential of hormesis in bee-pesticide research. Pest Manag. Sci. 2015, 71, 1368–1370. [Google Scholar] [CrossRef]
- Mallqui, K.S.V.; Vieira, J.L.; Guedes, R.N.C. Azadirachtin-induced hormesis mediating shift in fecundity-longevity trade-off in the Mexican bean weevil (Chrysomelidae: Bruchinae). J. Econ. Entomol. 2014, 107, 860–866. [Google Scholar] [CrossRef]
- Fauser-Misslin, A.; Sadd, B.M.; Neumann, P.; Sandrock, C. Influence of combined pesticide and parasite exposure on bumblebee colony traits in the laboratory. J. Appl. Ecol. 2014, 51, 450–459. [Google Scholar] [CrossRef]
- Brittain, C.A.; Vighi, M.; Bommarco, R.; Settele, J.; Potts, S.G. Impacts of a pesticide on pollinator species richness at different spatial scales. Basic Appl. Ecol. 2010, 11, 106–115. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strobl, V.; Camenzind, D.; Minnameyer, A.; Walker, S.; Eyer, M.; Neumann, P.; Straub, L. Positive Correlation between Pesticide Consumption and Longevity in Solitary Bees: Are We Overlooking Fitness Trade-Offs? Insects 2020, 11, 819. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110819
Strobl V, Camenzind D, Minnameyer A, Walker S, Eyer M, Neumann P, Straub L. Positive Correlation between Pesticide Consumption and Longevity in Solitary Bees: Are We Overlooking Fitness Trade-Offs? Insects. 2020; 11(11):819. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110819
Chicago/Turabian StyleStrobl, Verena, Domenic Camenzind, Angela Minnameyer, Stephanie Walker, Michael Eyer, Peter Neumann, and Lars Straub. 2020. "Positive Correlation between Pesticide Consumption and Longevity in Solitary Bees: Are We Overlooking Fitness Trade-Offs?" Insects 11, no. 11: 819. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110819