
Parasitoid Distribution and Parasitism of the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Maize Producing Regions of Uganda

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
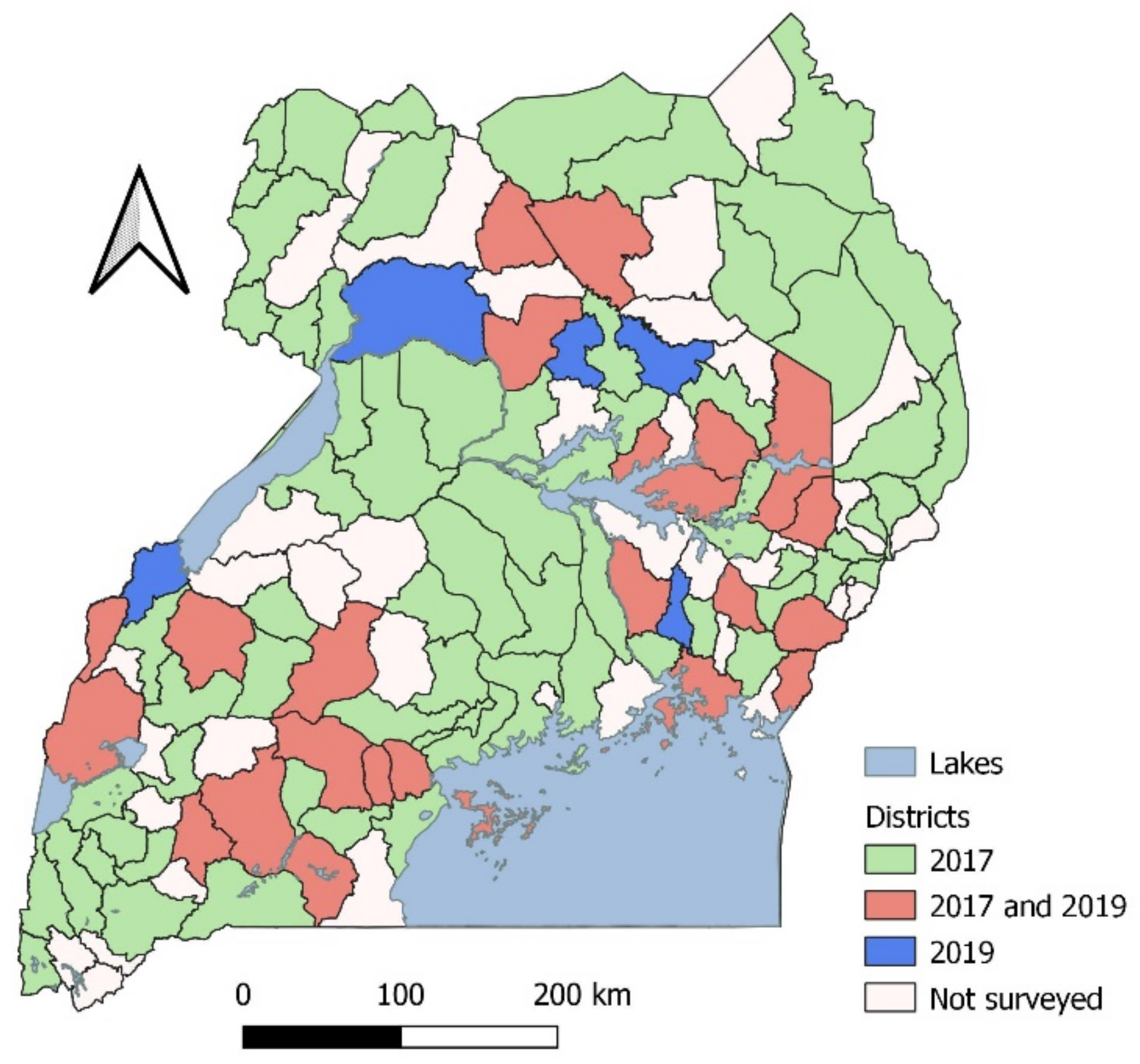
2.1. Survey for FAW and Natural Enemies
2.2. Rearing and Identification of Parasitoids
2.3. DNA Extraction
2.4. PCR Amplification of COI Segments
2.5. Sequencing and Sequence Analysis
2.6. Phylogenetic Analysis
2.6.1. Telenomus spp.
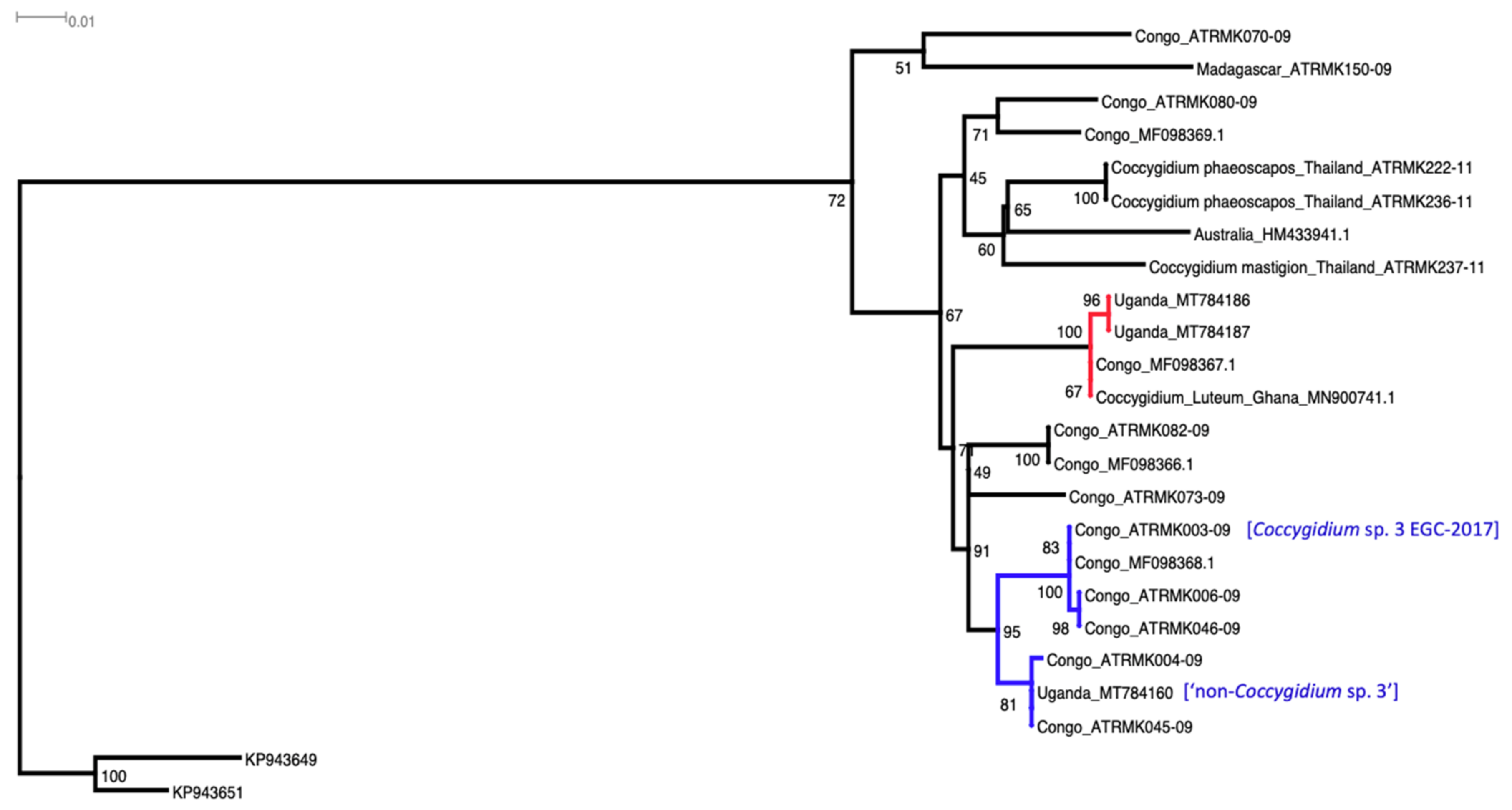
2.6.2. Coccygidium spp.
2.7. Relative Abundance and Parasitism Rates
3. Results
3.1. Distribution of Egg Parasitoids
3.2. Distribution of Egg/Larval Parasitoids of Spodoptera Frugiperda
Chelonus bifoveolatus
3.3. Distribution of Larval Parasitoids of Spodoptera Frugiperda
3.3.1. Coccygidium Luteum and Coccygidium sp.
3.3.2. Cotesia flavipes
3.3.3. Cotesia Icipe
3.3.4. Diolcogaster sp.
3.4. Distribution of Larval/Pupal Parasitoids of Spodoptera Frugiperda
3.4.1. Meteorus sp.
3.4.2. Charops cf. Diversipes
3.4.3. Sturmiopsis parasitica
3.4.4. Drino quadrizonula
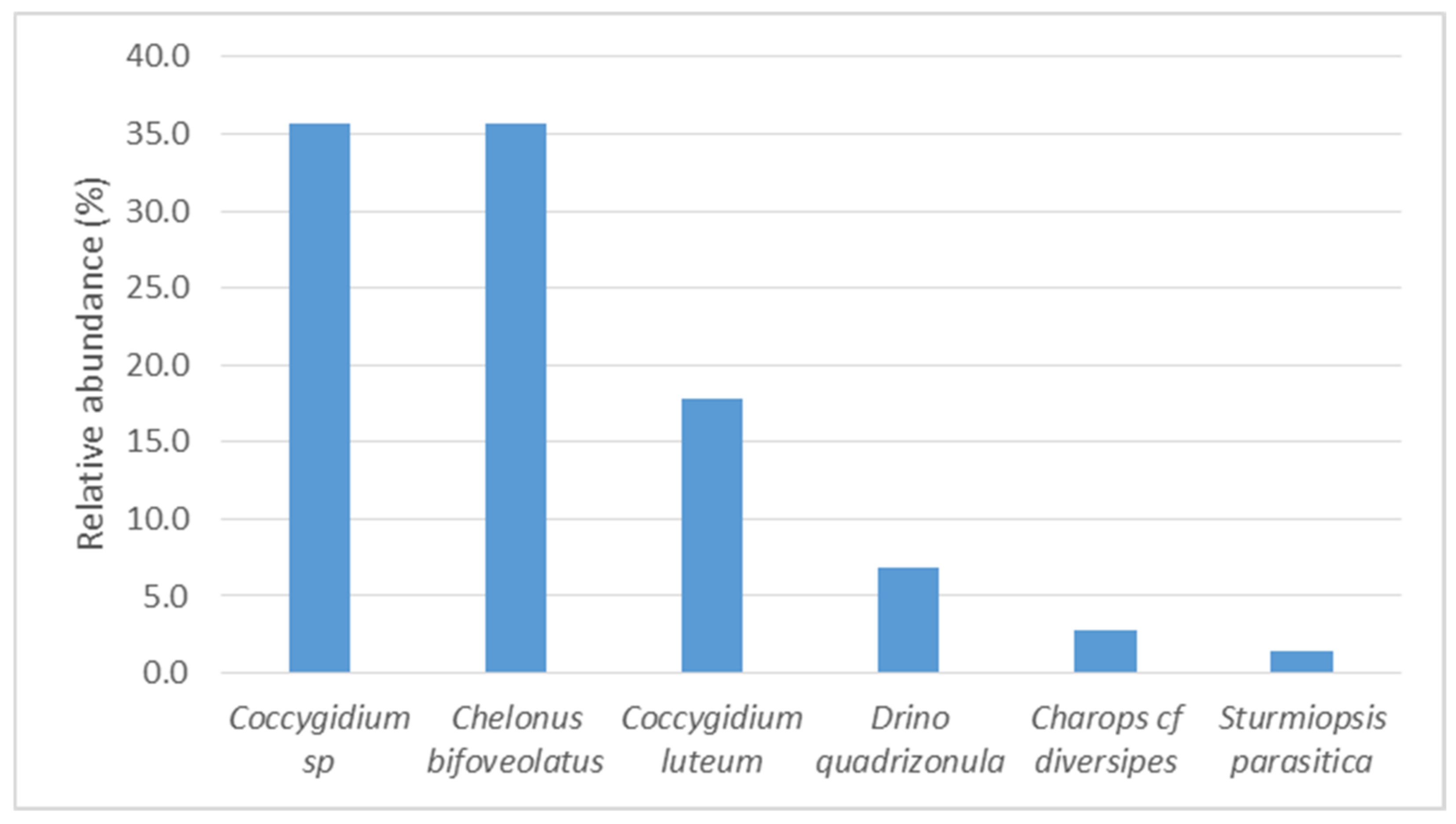
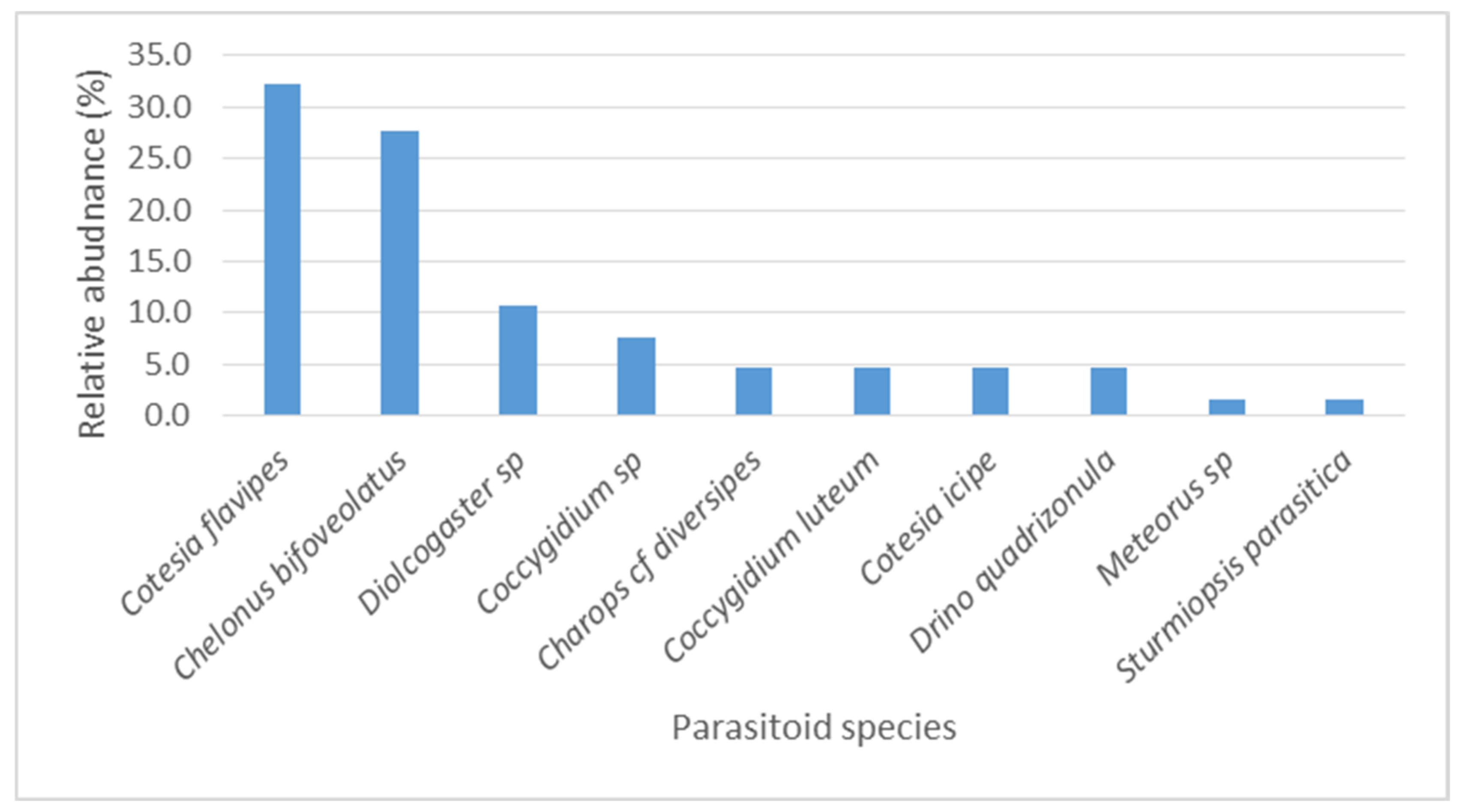
3.5. Relative Abundance and Parasitism of Spodoptera Frugiperda Eggs and Larvae
4. Discussions
4.1. Coccygidium spp.
4.2. Chelonus Bifoveolatus
4.3. Telenomus spp.
4.4. Tachinidae spp.
4.5. Cotesia spp.
4.6. Charops cf. Diversipes
4.7. Meteorus sp.
4.8. Diolcogaster sp.
4.9. Parasitism of FAW Larvae
5. Study Limitations
Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Briefing Note on FAO Actions on Fall Armyworm; Brief Note 03; FAO: Rome, Italy, 2018; pp. 1–6. [Google Scholar]
- Nagoshi, R.N.; Koffi, D.; Agboka, K.; Tounou, K.A.; Banerjee, R.; Jurat-Fuentes, J.L.; Meagher, R.L. Comparative molecular analyses of invasive fall armyworm in Togo reveal strong similarities to populations from the eastern United States and the Greater Antilles. PLoS ONE 2017, 12, e0181982. [Google Scholar] [CrossRef] [PubMed]
- Otim, M.H.; Tay, W.T.; Walsh, T.K.; Kanyesigye, D.; Adumo, S.; Abongosi, J.; Ochen, S.; Sserumaga, J.; Alibu, S.; Abalo, G.; et al. Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Uganda. PLoS ONE 2018, 13, e0194571. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Seo, B.Y.; Lee, J.; Kim, H.; Song, J.H.; Lee, W. First Report of the fall armyworm, Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae), a new migratory pest in Korea. Korean J. Appl. Entomol. 2020, 59, 73–78. [Google Scholar]
- Uzayisenga, B.; Waweru, B.; Kajuga, J.; Karangwa, P.; Uwumukiza, B.; Edgington, S.; Thompson, E.; Offord, L.; Cafá, G.; Buddie, A. First record of the fall armyworm, Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae), in Rwanda. Afr. Entomol. 2018, 26, 244–246. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Chimweta, M.; Nyakudya, I.W.; Jimu, L.; Mashingaidze, A.B. Fall armyworm [Spodoptera frugiperda (J.E. Smith)] damage in maize: Management options for flood-recession cropping smallholder farmers. Int. J. Pest Manag. 2020, 66, 142–154. [Google Scholar] [CrossRef]
- Tambo, J.A.; Day, R.K.; Lamontagne-Godwin, J.; Silvestri, S.; Beseh, P.K.; Oppong-Mensah, B.; Phiri, N.A.; Matimelo, M. Tackling fall armyworm (Spodoptera frugiperda) outbreak in Africa: An analysis of farmers’ control actions. Int. J. Pest Manag. 2020, 66, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2019, 65, 1–9. [Google Scholar] [CrossRef]
- Midega, C.A.O.; Pittchar, J.O.; Pickett, J.A.; Hailu, G.W.; Khan, Z.R. A climate-adapted push-pull system effectively controls fall armyworm, Spodoptera frugiperda (J E Smith), in maize in East Africa. Crop Prot. 2018, 105, 10–15. [Google Scholar] [CrossRef]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the western hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Bateman, M.L.; Day, R.K.; Luke, B.; Edgington, S.; Kuhlmann, U.; Cock, M.J.W. Assessment of potential biopesticide options for managing fall armyworm (Spodoptera frugiperda) in Africa. J. Appl. Entomol. 2018, 142, 805–819. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.J. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Physiol. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutirrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; Difonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of target-site mutations conferring insecticide resistance in Spodoptera frugiperda. Insects 2020, 11, 545. [Google Scholar] [CrossRef]
- Guan, F.; Zhang, J.; Shen, H.; Wang, X.; Padovan, A.; Walsh, T.K.; Tay, W.T.; Gordon, K.H.J.; James, W.; Czepak, C.; et al. Whole-genome sequencing to detect mutations associated with resistance to insecticides and Bt proteins in Spodoptera frugiperda. Insect Sci. 2020, 1–12. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A.; et al. Genetic structure and insecticide resistance characteristics of fall armyworm populations invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef]
- Hruska, A.J. Fall armyworm (Spodoptera frugiperda) management by smallholders. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Molina-Ochoa, J.; Lezama-Gutierrez, R.; Gonzalez-Ramirez, M.; Lopez-Edwards, M.; Rodriguez-Vega, M.A.; Arceo-Palacios, F. Pathogens and parasitic nematodes associated with populations of fall armyworm (Lepidoptera: Noctuidae) larvae in Mexico. Fla. Entomol. 2003, 86, 244–253. [Google Scholar] [CrossRef]
- Ruiz-Nájera, R.E.; Ruiz-Estudillo, R.A.; Sánchez-Yáñez, J.M.; Molina-Ochoa, J.; Skoda, S.R.; Coutiño-Ruiz, R.; Pinto-Ruiz, R.; Guevara-Hernández, F.; Foster, J.E. Occurrence of entomopathogenic fungi and parasitic nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae collected in Central Chiapas, México. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Gardner, W.A.; Fuxa, J.R. Pathogens for the suppression of the fall armyworm. Fla. Entomol. 1980, 63, 439. [Google Scholar] [CrossRef]
- Camargo, A.M.; Castañera, P.; Farinós, G.P.; Huang, F. Comparative analysis of the genetic basis of Cry1F resistance in two strains of Spodoptera frugiperda originated from Puerto Rico and Florida. J. Invertebr. Pathol. 2017, 146, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Ashley, T.R. Geographical distribution and parasitization levels for parasitoids of the fall armyworm, Spodoptera frugiperda. Fla. Entomol. 1986, 69, 516–524. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Hamm, J.J.; Lezama-Gutiérrez, R.; López-Edwards, M.; González-Ramírez, M.; Pescador-Rubio, A. A survey of fall armyworm (Lepidoptera: Noctuidae) parasitoids in the Mexican states of Michoacán, Colima, Jalisco, and Tamaulipas. Fla. Entomol. 2001, 84, 31–36. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.G.; O’Neil, R.J. Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize. Crop Prot. 2006, 25, 1180–1190. [Google Scholar] [CrossRef]
- Murúa, M.G.; Molina-Ochoa, J.; Fidalgo, P. Natural distribution of parasitoids of larvae of the fall armyworm, Spodoptera frugiperda, in Argentina. J. Insect Sci. 2009, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Martínez, A.; Tolon-Becerra, A.; Lastra-Bravo, X.B. Biological control of Spodoptera frugiperda eggs using Telenomus remus Nixon in maize-bean-squash polyculture. Am. J. Agric. Biol. Sci. 2012, 7, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Hardke, J.T.; Lorenz, G.M.; Leonard, B.R. Fall armyworm (Lepidoptera: Noctuidae) ecology in Southeastern cotton. J. Integr. Pest Manag. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Hay-Roe, M.M.; Meagher, R.L.; Nagoshi, R.N.; Newman, Y. Distributional patterns of fall armyworm parasitoids in a corn field and a pasture field in Florida. Biol. Control 2016, 96, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Meagher, R.L.; Nuessly, G.S.; Nagoshi, R.N.; Hay-Roe, M.M. Parasitoids attacking fall armyworm (Lepidoptera: Noctuidae) in sweet corn habitats. Biol. Control 2016, 95, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.L.; Yang, B.; Xu, M.F.; Lin, W.; Wang, D.S.; Chen, K.W.; Chen, H.Y. First report of Telenomus remus parasitizing Spodoptera frugiperda and its field parasitism in southern China. J. Hymenopt. Res. 2019, 93, 95–102. [Google Scholar] [CrossRef]
- Firake, D.M.; Behere, G.T. Natural mortality of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in maize agroecosystems of northeast India. Biol. Control 2020, 148, 104303. [Google Scholar] [CrossRef]
- Firake, D.M.; Behere, G.T. Bioecological attributes and physiological indices of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) infesting ginger (Zingiber officinale Roscoe) plants in India. Crop Prot. 2020, 137. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Polaszek, A.; Kimani, S.W. Telenomus species (Hymenopetra: Scelionidae) attacking eggs of pyralid pests (Lepidoptera) in Africa: A review and guide to identification. Bull. Entomol. Res. 1990, 80, 57–71. [Google Scholar] [CrossRef]
- Shaw, M.R.; Huddleston, T. Classification and Biology of Braconid Wasps (Hymenoptera: Braconidae); Dolling, W.R., Askew, R., Eds.; The British Museum (Natural History): London, UK, 1991; Volume 7. [Google Scholar]
- Fiaboe, K.K.M.; Fernández-Triana, J.; Nyamu, F.W.; Agbodzavu, K.M. Cotesia icipe Sp. N., a new Microgastrinae wasp (Hymenoptera: Braconidae) of importance in the biological control of Lepidopteran pests in Africa. J. Hymenopt. Res. 2017, 61, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, H.; de Almeida, L.F.; Shaw, S.R.; Sarmiento, C.E. An illustrated key to Neotropical species of the genus Meteorus Haliday (Hymenoptera, Braconidae, Euphorinae). Zookeys 2015, 489, 33–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauld, I.D. An introduction to the Ichneumonidae of Australia; British Museum (Natural History): London, UK, 1984; 413p, ISBN 0565008951. [Google Scholar]
- O’Hara, J.E.; Shima, H.; Zhang, C. Annotated catalogue of the Tachinidae (Insecta: Diptera) of China. Zootaxa 2009, 2190, 1–236. [Google Scholar] [CrossRef]
- O’Hara, J.E.; Cerretti, P. Annotated catalogue of the Tachinidae (Insecta, Diptera) of the Afrotropical Region, with the description of seven new genera. Zookeys 2016, 2016, 1–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Song, J.; Liu, C.; Luo, K.; Han, J.; Li, Y.; Pang, X.; Xu, H.; Zhu, Y.; Xiao, P.; et al. Use of ITS2 region as the universal DNA barcode for plants and animals. PLoS ONE 2010, 5, e13102. [Google Scholar] [CrossRef] [Green Version]
- Staden, R.; Beal, K.F.; Bonfield, J.K. The Staden package, 1998. Methods Mol. Biol. 2000, 132, 115–130. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Brister, J.R.; Bolton, E.E.; Canese, K.; Comeau, D.C.; Funk, K.; Ketter, A.; Kim, S.; Kimchi, A.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2020, 48, D9–D16. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System: Barcoding. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramage, T.; Martins-Simoes, P.; Mialdea, G.; Allemand, R.; Duplouy, A.; Rousse, P.; Davies, N.; Roderick, G.K.; Charlat, S. A DNA barcode-based survey of terrestrial arthropods in the Society Islands of French Polynesia: Host diversity within the SymbioCode project. Eur. J. Taxon. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sow, A.; Brévault, T.; Delvare, G.; Haran, J.; Benoit, L.; d’Acier, A.C.; Galan, M.; Thiaw, C.; Soti, V.; Sembène, M. DNA sequencing to help identify crop pests and their natural enemies in agro-ecosystems: The case of the millet head miner Heliocheilus albipunctella (Lepidoptera: Noctuidae) in sub-Saharan Africa. Biol. Control 2018, 121, 199–207. [Google Scholar] [CrossRef]
- Sharkey, M.J.; Chapman, E.G. Phylogeny of the Agathidinae (Hymenoptera: Braconidae) with a revised tribal classification and the description of a new genus. In Proceedings of the Entomological Society of Washington, Washington, DC, USA, 1 December 2017; Volume 119, pp. 823–842. [Google Scholar]
- Sharkey, M.J.; Chapman, E.G. The Nearctic genera of Agathidinae (Hymenoptera: Braconidae) with a phylogenetic analysis, illustrated generic key, and the description of three new genera. Zootaxa 2015, 4000, 49–72. [Google Scholar] [CrossRef]
- Koffi, D.; Kyerematen, R.; Eziah, V.Y.; Agboka, K.; Adom, M.; Goergen, G.; Meagher, R.L. Natural enemies of the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in Ghana. Fla. Entomol. 2020, 103, 85. [Google Scholar] [CrossRef] [Green Version]
- Van Noort, S. WaspWeb: Hymenoptera of the Afrotropical Region. Iziko Museums of South Africa. Available online: www.waspweb.org (accessed on 23 July 2020).
- Agboyi, L.K.; Mensah, S.A.; Clottey, V.A.; Beseh, P.; Glikpo, R.; Rwomushana, I.; Day, R.; Kenis, M. Evidence of leaf consumption rate decrease in fall armyworm, Spodoptera frugiperda, larvae parasitized by Coccygidium luteum. Insects 2019, 10, 410. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Soujanya, P.L.; van Achterberg, C.; Sekhar, J.C. Coccygidium transcaspicum (Kokujev) (Hymenoptera: Braconidae) parasitizing larvae of invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in India. Zootaxa 2020, 4750, 293–297. [Google Scholar] [CrossRef]
- Sharanabasappa, D.; Kalleshwaraswamy, C.M.; Poorani, J.; Maruthi, M.S.; Pavithra, H.B.; Diraviam, J. Natural enemies of Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae), a recent invasive pest on maize in South India. Fla. Entomol. 2019, 102, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.V.R.; Wightman, J.A.; Rao, D.V.R. World review of the natural enemies and diseases of Spodoptera litura (F.) (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 1993, 14, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Stevens, N.B.; Austin, A.D.; Jennings, J.T. Synopsis of Australian Agathidine wasps (Hymenoptera: Braconidae: Agathidinae). Zootaxa 2010, 2480, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Ngangambe, M.H.; Mwatawala, M.W. Effects of entomopathogenic fungi (EPFs) and cropping systems on parasitoids of fall armyworm (Spodoptera frugiperda) on maize in eastern central, Tanzania. Biocontrol Sci. Technol. 2020, 30, 418–430. [Google Scholar] [CrossRef]
- Zenner, I.; Álvarez, A.; Barreto, S. Influence of parasitism by Chelonus insularis Cresson (Hymenoptera: Braconidae) on the susceptibility of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) to insecticides. Neotrop. Entomol. 2006, 35, 818–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittel, R.N.; Austin, A.D.; Klopfstein, S. Molecular and morphological phylogenetics of Chelonine parasitoid wasps (Hymenoptera: Braconidae), with a critical assessment of divergence time estimations. Mol. Phylogenet. Evol. 2016, 101, 224–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Laminou, S.A.; Ba, M.N.; Karimoune, L.; Doumma, A.; Muniappan, R. Parasitism of locally recruited egg parasitoids of the fall armyworm in Africa. Insects 2020, 11, 430. [Google Scholar] [CrossRef] [PubMed]
- Kankonda, O.M.; Akaibe, B.D.; Sylvain, N.M.; Le Ru, B.P. Response of maize stemborers and associated parasitoids to the spread of grasses in the rainforest zone of Kisangani, DR Congo: Effect on stemborers biological control. Agric. Entomol. 2018, 20, 150–161. [Google Scholar] [CrossRef]
- Hordzi, W.H.K. Lepidopterous stem borers of maize (Zea mays): Agro-ecological and regional composition and distribution of their parasitoids in Southern Ghana. J. Biosci. Biotechnol. Discov. 2018, 3, 46–58. [Google Scholar] [CrossRef]
- Smith, M.A.; Fernández-Triana, J.L.; Eveleigh, E.; Gómez, J.; Guclu, C.; Hallwachs, W.; Hebert, P.D.N.; Hrcek, J.; Huber, J.T.; Janzen, D.; et al. DNA barcoding and the taxonomy of Microgastrinae wasps (Hymenoptera, Braconidae): Impacts after 8 years and nearly 20 000 sequences. Mol. Ecol. Resour. 2013, 13, 168–176. [Google Scholar] [CrossRef]
- Muirhead, K.A.; Murphy, N.P.; Sallam, N.; Donnellan, S.C.; Austin, A.D. Phylogenetics and genetic diversity of the Cotesia flavipes complex of parasitoid wasps (Hymenoptera: Braconidae), biological control agents of Lepidopteran stemborers. Mol. Phylogenet. Evol. 2012, 63, 904–914. [Google Scholar] [CrossRef]
- Jourdie, V.; Alvarez, N.; Turlings, T.C.J. Identification of seven species of Hymenopteran parasitoids of Spodoptera frugiperda, using polymerase chain reaction amplification and restriction enzyme digestion. Agric. Entomol. 2008, 10, 129–136. [Google Scholar] [CrossRef] [Green Version]
- De Freitas, J.G.; Takahashi, T.A.; Figueiredo, L.L.; Fernandes, P.M.; Camargo, L.F.; Watanabe, I.M.; Foerster, L.A.; Fernandez-Triana, J.; Shimbori, E.M. First record of Cotesia scotti (Valerio and Whitfield, 2009) (Hymenoptera: Braconidae: Microgastrinae) comb. nov. parasitising Spodoptera cosmioides (Walk, 1858) and Spodoptera eridania (Stoll, 1782) (Lepidoptera: Noctuidae) in Brazil. Rev. Bras. Entomol. 2019, 63, 238–244. [Google Scholar] [CrossRef]
- Desneux, N.; Ramírez-Romero, R.; Bokonon-Ganta, A.H.; Bernal, J.S. Attraction of the parasitoid Cotesia marginiventris to host (Spodoptera frugiperda) frass is affected by transgenic maize. Ecotoxicology 2010, 19, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Van Nouhuys, S.; Tay, W.T. Causes and consequences of small population size for a specialist parasitoid wasp. Oecologia 2001, 128, 126–133. [Google Scholar] [CrossRef] [PubMed]
- López, M.A.; Martínez-Castillo, A.M.; García-Gutiérrez, C.; Cortez-Mondaca, E.; Escobedo-Bonilla, C.M. Parasitoids and entomopathogens associated with fall armyworm, Spodoptera frugiperda, in Northern Sinaloa. Southwest. Entomol. 2018, 43, 867–881. [Google Scholar] [CrossRef]
- Yu, D.S.K.; van Achterberg, C.; Horstmann, K. Taxapad 2016, Ichneumonoidea 2015; Database on Flash-Drive: Nepean, ON, Canada, 2016; Available online: http://www.taxapad.com (accessed on 4 September 2020).
- Fernandez-Triana, J. Ten unique and charismatic new species of microgastrinae wasps (Hymenoptera: Braconidae) from North America. Zookeys 2018, 2018, 123–150. [Google Scholar] [CrossRef] [Green Version]
- Caniço, A.; Mexia, A.; Santos, L. First report of native parasitoids of fall armyworm Spodoptera frugiperda smith (Lepidoptera: Noctuidae) in Mozambique. Insects 2020, 11, 615. [Google Scholar] [CrossRef]
- Tendeng, E.; Labou, B.; Diatte, M.; Djiba, S.; Diarra, K. The fall armyworm Spodoptera frugiperda (J.E. Smith), a new pest of maize in Africa: Biology and first native natural enemies detected. Int. J. Biol. Chem. Sci. 2019, 13, 1011. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order, Family, and Species | GenBank Accession No. of Ugandan Parasitoids | Host Stage Attacked | Species with Closest NucleoTide Sequence Match | Percent Identity, and Reference GenBank Accession Number and iBoL Entries |
|---|---|---|---|---|
| Hymenoptera: Platygastridae | ||||
| Telenomus remus Dixon | MT780201, MT780202 | Eggs | Telenomus remus | 100% (MK533751) [38] |
| Telenomus sp. | MT782153 and MT782154 | Eggs | Telenomus remus | 96.6% (MK533751) [38] |
| Platygastridae sp. | MT784162 | Eggs | Telenomus sp. | 87.4% (KR878931) |
| Hymenoptera: Braconidae | ||||
| Chelonus bifoveolatus | MT776316 to MT776320 | Egg/larvae | Chelonus bifoveolatus | 99.8–100% (KX051941) [56]; MN900744, MN900743 [39]) |
| Coccygidium luteum | MT784186, MT784187 | Larvae | Coccygidium. luteum | 99.64%; (MN900741) [39]) |
| Coccygidium sp. | MT784160 | Larvae | Coccygidium sp. 3 | 98.2%; (MF098368) |
| Cotesia flavipes | MT780220 | Larvae | Cotesia flavipes | 100% (JF865973) and 99.6% (MN900741; MN900728) [39] |
| Cotesia icipe | MT780217 to MT780219 | Larvae | Cotesia icipe | 100% (iBoL ID JMIC0355) and 99.6% (MN900741; MN900728) [39] and 100% (JMIC0355) |
| Diolcogaster sp. | MT784194 | Larvae | Diolcogaster sp. | 98.38% (HM397609) |
| Meteorus sp. | MT784161 | Larvae/pupae | Meteorus sp. | 99.84% (MF673599) [57]) |
| Hymenoptera: Ichneumonidae | ||||
| Charops cf. diversipes | MT784179 to MT784183 | Larvae | Charops cf. diversipes | 99.8–100% (MN900729) [39] |
| Diptera: Tachinidae | ||||
| Sturmiopsis parasitica(Curran) | MT784184 and MT784185 | Larvae/pupae | Sturmiopsisparasitica | 99.35% (DQ336399) |
| Drino quadrizonula | MT784175 to MT784178 | Larvae/pupae | Drino quadrizonula | 99.28–99.8% (MN907776) [39]) |
| Region | District | Species | Relative Abundance (%) (n = 35) |
|---|---|---|---|
| Eastern | Bukedea | Telenomus remus | 28.6 |
| Soroti | Telenomus sp. † | 14.3 | |
| Western | Ntoroko | Platygastridae | 2.9 |
| Ntoroko | Telenomus remus | 31.4 | |
| Mbarara | Telenomus sp. † | 22.9 |
| Host Stage Attacked, Parasitoid Species | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Egg/larva | Larva | Larva/pupa | |||||||
| Region/District | No. Larvae Collected | No. Parasitoids Recovered | Overall Parasitism, % | Chb | Col | Cos | Cha | Spa | Dri |
| Central | |||||||||
| Sembabule | 2 | 1 | 50 | 50 | |||||
| Eastern | |||||||||
| Amuria | 33 | 1 | 3 | 3 | |||||
| Bukedea | 67 | 1 | 1.5 | 1.5 | |||||
| Kaberamaido | 29 | 3 | 10.3 | 6.9 | 3.4 | ||||
| Kamuli | 15 | 1 | 6.7 | 6.7 | |||||
| Katakwi | 35 | 4 | 11.4 | 8.6 | 2.9 | ||||
| Kumi | 42 | 5 | 11.9 | 4.8 | 4.8 | 2.4 | |||
| Mayuge | 46 | 2 | 4.3 | 4.3 | |||||
| Namutumba | 22 | 4 | 18.1 | 13.6 | 4.5 | ||||
| Pallisa | 58 | 1 | 1.7 | 1.7 | |||||
| Serere | 32 | 1 | 3.1 | 3.1 | |||||
| Sironko | 40 | 1 | 2.5 | 2.5 | |||||
| Soroti | 30 | 3 | 10 | 3.3 | 6.7 | ||||
| Northern | |||||||||
| Adjumani | 14 | 2 | 14.3 | 14.3 | |||||
| Alebtong | 10 | 2 | 20 | 20 | |||||
| Dokolo | 32 | 2 | 6.3 | 3.1 | 3.1 | ||||
| Gulu | 24 | 1 | 4.2 | 4.2 | |||||
| Kaabong | 37 | 3 | 8.1 | 5.4 | 2.7 | ||||
| Kole | 30 | 1 | 3.3 | 3.3 | |||||
| Kotido | 42 | 4 | 9.5 | 9.5 | |||||
| Lira | 40 | 4 | 10 | 10 | |||||
| Maracha | 75 | 1 | 1.3 | 1.3 | |||||
| Moroto | 9 | 1 | 11.1 | 11.1 | |||||
| Moyo | 23 | 3 | 13 | 13 | |||||
| Nebbi | 36 | 5 | 13.9 | 13.9 | |||||
| Nwoya | 26 | 2 | 7.6 | 3.8 | 3.8 | ||||
| Oyam | 19 | 2 | 10.6 | 5.3 | 5.3 | ||||
| Pader | 35 | 4 | 11.4 | 11.4 | |||||
| Pakwach | 24 | 1 | 4.2 | 4.2 | |||||
| Yumbe | 39 | 2 | 5.1 | 5.1 | |||||
| Zombo | 36 | 2 | 5.6 | 5.6 | |||||
| Western | |||||||||
| Kasese | 35 | 3 | 8.6 | 8.6 | |||||
| Host Stage Attacked, Parasitoid Species | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Egg/Larva | Larva | Larva/Pupa | |||||||||||
| Region/District | No. Larvae Collected | No. Parasitoids Recovered | Overall Parasitism, % | Chb | Col | Cos | Cof | Coi | Dio | Cha | Met | Spa | Dri |
| Central | |||||||||||||
| Mubende | 20 | 1 | 5 | 5 | |||||||||
| Sembabule | 3 | 1 | 33.3 | 33.3 | |||||||||
| Eastern | |||||||||||||
| Bukedea | 21 | 1 | 4.8 | 4.8 | |||||||||
| Busia | 32 | 2 | 6.2 | 3.1 | 3.1 | ||||||||
| Kaberamaido | 24 | 1 | 4.2 | 4.2 | |||||||||
| Kamuli | 48 | 2 | 4.1 | 2.1 | 2.1 | ||||||||
| Katakwi | 19 | 1 | 5.3 | 5.3 | |||||||||
| Kumi | 27 | 3 | 11.1 | 3.7 | 7.4 | ||||||||
| Luuka | 74 | 1 | 1.4 | 1.4 | |||||||||
| Mayuge | 49 | 1 | 2 | 2 | |||||||||
| Namutumba | 30 | 3 | 10 | 6.7 | 3.3 | ||||||||
| Serere | 28 | 1 | 3.6 | 3.6 | |||||||||
| Soroti | 30 | 6 | 20 | 16.7 | 3.3 | ||||||||
| Northern | |||||||||||||
| Alebtong | 119 | 1 | 0.8 | 0.8 | |||||||||
| Gulu | 58 | 1 | 1.7 | 1.7 | |||||||||
| Kole | 59 | 10 | 17 | 5.1 | 11.9 | ||||||||
| Nwoya | 42 | 4 | 9.5 | 4.8 | 2.4 | 2.4 | |||||||
| Western | |||||||||||||
| Kasese | 31 | 5 | 16.1 | 9.7 | 3.2 | 3.2 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otim, M.H.; Adumo Aropet, S.; Opio, M.; Kanyesigye, D.; Nakelet Opolot, H.; Tek Tay, W. Parasitoid Distribution and Parasitism of the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Maize Producing Regions of Uganda. Insects 2021, 12, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020121
Otim MH, Adumo Aropet S, Opio M, Kanyesigye D, Nakelet Opolot H, Tek Tay W. Parasitoid Distribution and Parasitism of the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Maize Producing Regions of Uganda. Insects. 2021; 12(2):121. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020121
Chicago/Turabian StyleOtim, Michael Hilary, Stella Adumo Aropet, Moses Opio, Dalton Kanyesigye, Henry Nakelet Opolot, and Wee Tek Tay. 2021. "Parasitoid Distribution and Parasitism of the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Maize Producing Regions of Uganda" Insects 12, no. 2: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020121